256
ON THE CONTROL OF THE LEVEL OF ACTIVITY
OF THE ANIMAL BODY
I. THE ENDOCRINE CONTROL OF SEASONAL VARIATIONS
OF ACTIVITY IN THE FROG
BY G. S. CARTER.
(From the Sub-department of Experimental Zoology of the University of Cambridge.)
{Received 20th December, 1932.)
(With Eight Text-figures.)
HIBERNATION and the other seasonal variations of activity in the vertebrate have been . the subject of a very large amount of investigation and of an even larger literature. This has been recently summarised by Pocock (1926) from the standpoint of general biology, and by Gorer (1931) from that of comparative physiology. Here, it is only necessary to discuss previous work on those aspects of the subject with which this paper is immediately concerned.
Seasonal variations of activity are of many different types. An uninterrupted series may be found between the complete lethargy of hibernating mammals and reptiles and the relative inactivity which many animals, such as the frog, the subject of this paper, show in cold conditions. The series may, perhaps, be extended from these conditions of relative inactivity to the seasonal physiological changes, often unaccompanied by any obvious inactivity, which occur in many non-hibernating mammals. Nor is cold the only condition in which animals become inactive; aestivation is as well-known a phenomenon as hibernation.
It is by no means clear, without further evidence, that conditions of all these types are due to similar physiological changes within the body. However, in most of them the body passes into a more or less completely inactive state, and many show other, more definite, physiological resemblances. Whether it is or is not true that all such conditions are physiologically similar, it will be advisable, in discussing the previous work on the aspects of their physiology with which we are concerned, to consider evidence derived from all these conditions, since it is possible that any of this evidence may have some bearing on the changes which occur in the frog. This will be done, but it must not be forgotten that the experimental results to be given in the paper are concerned only with the seasonal changes of the frog. Generalisation from these results cannot be justified until other types of seasonal change have been investigated. It is hoped to extend the observations in later papers.
frog. It is not intended to draw from this evidence the further conclusion that other organs play no part in this control. It is, indeed, probable that the nervous system is also concerned, but the part played by it and other organs is outside the subject of the paper and will not be further discussed.
The belief that the control of seasonal variations is endocrine is by no means new. Even if we restrict ourselves to the more recent literature, we shall find that at one time or another almost every endocrine organ has been put forward as being actively concerned in the control of these changes.
Evidence of at least three distinct types has been used to support these con-clusions.
Firstly, extracts of the endocrine organs have been injected and their effects on the activity of the animal observed. In experiments of this type Adler (1920 b) and Schenk (1922) found that extracts of the thymus, thyroid, adrenal and anterior pituitary woke the hedgehog from its winter sleep, but their results were criticised by Zondek (1924) and have not been generally accepted as conclusive. Also, Dworkin and Finney (1927) found that large injections of insulin induced winter sleep in the marmot.
Secondly, seasonal changes have been observed in the condition of the endocrine organs. Adler (1920 b) found regressive changes during the winter in the thyroid of hedgehogs and, more markedly, of bats. In non-hibernating mammals and birds the thyroid is larger in the winter (Riddle and Fisher, 1925, who give evidence for the pigeon and summarise the literature), but, on the other hand, its iodine content is higher in the summer and to an extent which more than compensates for its larger size in the winter (Seidell and Fenger, 1913; Kendall and Simonsen, 1928). These last results show clearly the inadequacy of histological evidence alone, when the question at issue is, as it is here, the functional activity of a gland. Neither the size nor the histological appearance of a gland is necessarily correlated with the amount of its secretion which it is pouring into the circulation.
Mann (1916) was unable to find constant seasonal changes in any of the endocrine organs of the woodchuck. Inconstant changes in the pituitary and adrenal were observed.
258 G. S. CARTER
effects of the thymus and most of those of the pituitary (v. Bugbee, Simond and Grimes, 1931) show no definite correlation with the changes associated with hiber-nation. An exception with regard to the anterior pituitary will be discussed in the succeeding paper of this series.
At first sight one other line of evidence seems to bear on the subject under discussion. There is evidence that the condition of the endocrine organs varies in races living under different conditions. Thus, Adler (1920 a) found that frogs
(R. temporaria) living continuously in cold conditions have abnormally large and
active thyroids. But such races must remain active in the cold conditions in order to maintain their life, and their needs are very different from those of animals which can become inactive when the temperature falls, passing into hibernation. Adler's evidence cannot therefore be considered to indicate activity of the thyroid in hibernation. Rather, it indicates the opposite conclusion—that the thyroid is active when the animal is forced to be abnormally active to maintain its life in cold conditions.
From this survey of the literature it may be concluded that there is considerable evidence that the endocrine organs are concerned in the control of seasonal changes, but that the evidence is not yet sufficient to decide which of these organs is re-sponsible for their control. On the whole the most reliable evidence suggests that the thyroid and the adrenal are concerned, but it cannot be said that this has been established. Very probably more than one organ plays a part in this control, but it is hardly likely that all those for which evidence has been put forward are equally concerned. There can be little doubt that the reason why positive results have been obtained with so many of these organs is that most of the experimental work has been carried out within the body, where the interactions of the endocrine system are extraordinarily complex. When the endocrine balance in the body is disturbed in any way, it is extremely hard to decide which of the observed results are directly due to the primary alteration and which to secondary alterations induced in other organs. If evidence could be obtained from experiments carried out outside the body, it would seem to be of more value in the discussion of these problems. Recently an opportunity of gaining evidence of this type has been provided by the results of Barcroft and Izquierdo (1931). They found that the temperature-pulse rate curve of the excised heart of the frog is of different form according as the experiment is carried out in the summer or the winter. The curve given by the winter heart is exponential between 5 and 20° C , whereas that of a summer heart is almost straight over the same range of temperature and can be shown, when plotted logarithmically, to be composed of two exponential curves crossing at or slightly above io° C. They found the intact heart of the summer frog to give a curve of the same form as that given by the excised heart in the summer. In the winter they failed to get concordant results from the intact heart.
this paper consisted in determining the effects of various endocrine and related substances on the form of this curve. The results of such experiments should give, at least, some indication of the changes within the body which cause the difference in the condition of the heart.
MATERIAL AND METHODS.
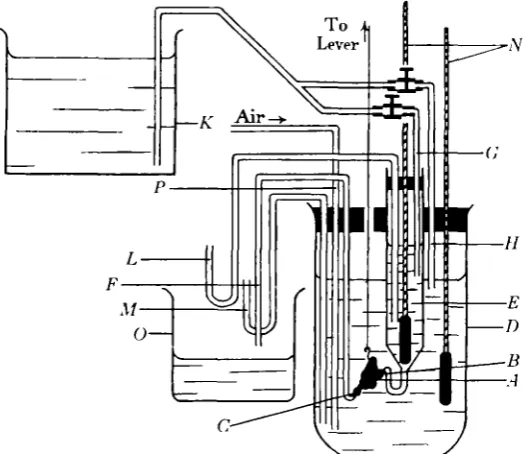
[image:4.595.167.429.293.520.2]Both the common frog (Rana temporaria) and the edible frog (R. esculenta) were used as material for these experiments. There is no difference in the behaviour of the hearts of these two species in the phenomena which were being investigated, except that the heart of R. esculenta is able to support a slightly higher temperature without
Fig. 1. Apparatus for circulating fluids through the frog's heart. The different parts of this figure are not drawn to the same scale.
damage. In most of the experiments a temperature range of 5-200 C. was used for the heart of R. temporaria and 5-240 C. for that of R. esculenta. Exposure to the highest of these temperatures for the necessary period did not alter the rate of beat at lower temperatures. It was therefore assumed that no damage had been done to
the hearts by the high temperature.
260 G. S. C A R T E R
were maintained constant by the siphons L and M. The heights of these siphons were adjusted so that the heart was under a venous pressure of about 3 in. The liquid in the vessel D could be stirred by a current of air through the tube P. The fluid which had passed through the apparatus was replaced from the dish O into K by a suction device. In the course of its passage between these two dishes it was well aerated in the tubes of this device. In most of the experiments a litre of fluid was used and this passed round the apparatus in 1-2 hours. The temperature of the heart was altered by immersing the vessel D in a much larger vessel of warmer or colder water. It was kept constant at any value by filling this larger vessel with water of the required temperature.
Barcroft and Izquierdo (loc. cit.) found difficulty in obtaining the same rate of beat at any given temperature when the temperature was rising as when it was falling. They found it necessary to allow the heart to beat outside the body for some hours before they could get this result. It was found that this difficulty was almost entirely overcome by keeping the temperature of the fluid constant for at least five minutes before the observations of rate of beat were made. All the experiments were carried out in this way. It was found necessary, however, to allow the heart to beat for at least one hour after the apparatus had been set up, and for at least half an hour after it had been disturbed in any way, as, for instance, in adding substances to the fluid.
The procedure of keeping the temperature constant for five minutes before each observation and the fact that several curves were needed in many of the experiments (and therefore not more than 1-2 hours could be given to each) made it necessary to observe fewer points on the curve than were observed by Barcroft and Izquierdo in some of their experiments, but the smaller errors in each observation at least compensated for this disadvantage. The experimental error is clearly proportional to the rate of beat. In several determinations it was found to be less than 0-5 beat per minute at the lowest temperatures and 2 beats per minute at the highest. An error of about this amount may be assumed throughout. On the logarithmic plot this is equivalent to an error of log"1 0-02 throughout the range of temperature.
The fluid circulating through the apparatus was a Ringer solution of the following constitution:
/
/o NaH
2PO4
Glucose Lecithin
/ U
o-ooi
O-2
O-OOO2
NaCl 0-65
KC1 0-014
CaCl2 0-012
The lecithin was added on the evidence of Clark (1913) that it prolongs the life of the heart. The/>H of this solution was brought to that required by adding the necessary amount of a strong solution of NajHPO4. The amount necessary to give a />H of 76,
which was used in almost all the experiments, was equivalent to about 0-012 per cent. NajHPOj.
in order that the control of />H might be more effective. In spite of this, there was always a slight movement of the />H in the acid direction during the experiments (to />H 7-3-4 after 2-3 hours). The />H was readjusted to pH 7-6, whenever the medium was altered, i.e. after each curve had been obtained. The effects of variations in pH were observed in experiments recorded below.
When the action of synthetic substances was to be observed, these substances were dissolved in N/10 NaOH and added to the medium in the correct proportion. Adrenaline alone was dissolved in N/10 HC1. ThepH of the medium was readjusted after these additions. Extracts of various organs were made by grinding the tissue in Ringer solution and precipitating the proteins by means of tri-chlor-acetic acid. The only commercial extract used was that of pituitrin. The extracts were added directly to the medium.
Each result was confirmed in a series of at least three experiments. Wherever the results were doubtful or contradictory, a longer series of experiments was carried out, and this was also done where the experiments seemed to be of especial import-ance, for example, the experiments on thyroxine and adrenaline, and on the effect of long-continued beat in unmodified Ringer solution.
EXPERIMENTAL RESULTS.
(1) The summer and winter form of the pulse rate-temperature curve.
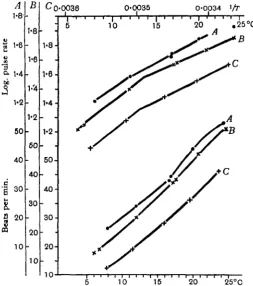
Typical examples of the two forms of the curve are given in Fig. 2. In this and all the succeeding figures the observations of pulse rate are directly plotted against the temperature in the lower part of the figure, and smoothed curves are drawn through these points. In the upper part of the figure these smoothed curves are redrawn on a plot of which the abscissa is the reciprocal of the absolute temperature and the ordinate the logarithm of the pulse rate. Although the lines of the upper part of the figure are drawn from the smoothed curves of the lower part, the observed points have been replotted in order that the goodness of the fit may be estimated. The procedure of plotting the figures logarithmically has been carried out because it seemed that the alteration of the shape of the curve could be most easily recognised in the logarithmic plot. It is not intended to discuss whether or not the form of the logarithmic curve has any biological meaning.
The curves of Fig. 2 are redrawn from results published by Barcroft and Izquierdo (loc. cit.). Both curves refer to isolated hearts.
262 G. S. CARTER
Results will be given below (Figs. 3,5,6) which show that no general increase or decrease in the rate of beat can be observed as the heart passes from one seasonal condition to the other. When plotted on the same ordinates and abscissae, the sum-mer and winter curves lie over each other. It is for this reason that in all the figures except Fig. 2 the ordinates have been stepped down for each curve, so that the curves may not become confused with each other.
0-0036
I
10 15 20 25°C
Fig. 2. Pulse-rate curves of the summer and winter forms, R. temporaria. Redrawn from Barcroft and Izquierdo, 1931, pp. 149, 152. S, summer; W, winter.
On the other hand, comparison of the figures will show that there is considerable variation in the rate of beat at the same temperature from one heart to another.
(2) Preliminary experiments.
(a) Long-continued beat in Ringer solution.
was hardly recognisable after 5-6 hours, but became evident after 12 hours (Fig.
3)-These results are explicable if some substance is present in the summer heart in greater concentration than in the winter heart, and is gradually reduced by being washed out of the heart or used up. They are very difficult to explain if the difference consists in greater concentration of a substance in the winter heart. Thus, if the endocrine theory is true, they are strongly in favour of greater activity of some organ in the summer.
0-0036 0-0035 0-0034 1/r
B ~"
1-8
1-6
•3
1-2
6 0
5 0
-g 40
I
30
20
1-8-
1-6-
1-4-
126 0
5 0
-40
3 0
2 0
-10
10 15 20 25°C
Fig. 3. R. esculenta. Pulse-rate curves of the same heart in Ringer solution. Curve B, I2i hours after curve A. A, 5.4.32, 10-11 p.m.; B, 6.4.32, 11.30 a.m.-i2.3O p.m.
(b) Effects of variation of the pH of the medium.
264 G. S. CARTER
equally bent, and the differences in form between them are probably within the experimental error. It may, therefore, be assumed that the variations of />H which occurred during the experiments had no appreciable effect on the form of the curves.
(c) The action of tri-chlor-acetic acid.
It has been mentioned that tri-chlor-acetic acid was used to precipitate proteins from the extracts of the endocrine organs used in these experiments. Since a
rat e puls e Log . | pe r
1
a A 1-8 1-6 1-4 1-2 50 4 0 30 20 10 B -1-8 -1-8 1-4 -1-2 -50 -40 -30 -20 -10C 0-0036 0-0035 0-0034 W
1-8- 1-6- 1-4- 1-2- 50- 40- 30- 20-10 10 15
10 15 20 25 °C
Fig. 4. R. esculenta. Pulse-rate curves of the same heart in Ringer solution of different pH. Curve A, pH 7-6; B, pH 7-2, z hours after curve A; C, pH 8-o, 1J hours after curve B.
failure of tri-chlor-acetic acid to produce any effect was not due to some damage of the heart caused by its high concentration.
(d) The action of tissue extracts of organs other than the endocrine organs.
It will be found that not all the extracts used in the following experiments had any effect upon the form of the curve given by the heart. Thus the results are prob-ably sufficient by themselves to exclude the possibility that tissue extracts of any organ might contain an effective substance. In order that this point might be more definitely established, a few experiments were carried out with extracts of rabbit muscle and salivary glands. No alteration of the form of the curve was observed, although the ability of the heart to react to thyroxine was not damaged by these extracts.
(3) The action of endocrine substances, (a) Thyroxine.
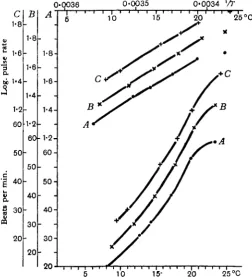
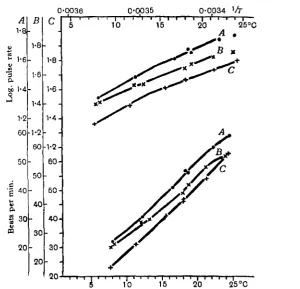
In Figs. 5 and 6 the effect of adding thyroxine to the winter hearts of R. esculenta and R. temporaria are shown. The same effect is shown in curve C of Fig. 8. These experiments are examples of a long series, all of which gave similar results.
A
1-8'1-e
1-6-3 a 1-4}.
,3
V21
3
60
40
30
2 0
10
B
1-8
1-6
1-2
60
40
30
20
10
C0-0036 0-0035 0-0034 </T
1-8-
1-6-
1-2-
50-
40-
30-
20-1 0 '
10 15 20 25°C
Fig. 5. R. etculenta. A, pulse-rate curve of a heart in the winter condition; B, curve given by the same heart after beating for 1 J hours in Ringer solution containing thyroxine, 1 in 5 x io'; C, 13 hours after return to Ringer solution.
[image:10.595.170.424.380.667.2]266 G. S. CARTER
It will be seen that the substance produces a definite alteration of the curve towards the summer form. The effect is produced rapidly and can always be obtained when the heart has been beating in the presence of thyroxine for one hour. No change in the form of the curve could be found when a heart in full summer condition was treated with thyroxine. When the heart was allowed to beat for some hours in unmodified Ringer solution after treatment with thyroxine there was some return towards the winter condition (Fig. 5), presumably due to loss of the thyroxine.
3
A
1-8
1-6
1-4
1-2 60
50
40
c •§
t 30
a S
« 20
10
B,0-0036 0-0035 0-0034
1-8-
1-6-
1-4- 1-2-
60-
50-
40-
30-20
10
25 °C
10 15 20 25 °C
Fig. 6. R. ttmporaria. A, pulse-rate curve of a heart in almost complete winter condition in Ringer solution; B, curve given by the same heart after beating for i \ hours in the same solution with the addition of thyroxine, i in 2 x 10*.
The lower limit of the effective concentration of thyroxine was approximately i in io7. At this concentration the substance produced a complete effect in two experiments, but in others it was ineffective. All concentrations between i in io6 and i in io7 were effective. When the concentration was above i in io6, the beat was slightly slowed and in some experiments the effect seemed to be less or absent.
therefore be expected, if the thyroid is concerned in the control of seasonal variations, that changes of at least 6y in the iodine content of the blood will occur. Thus, the evidence indicates that the protein secretion of the thyroid is no more effective than thyroxine in this effect upon the heart.
The figures also show that there is no general increase of the rate of beat in the presence of thyroxine. Barcroft and Izquierdo (1931) obtained similar results in untreated hearts of summer and winter frogs (e.g. Fig. 2). In this respect, as in the change of the form of the curve, the effect of thyroxine resembles the natural change. A general increase of the rate of beat is the effect which has most often been looked for in experiments on the action of thyroxine on the heart. These results are therefore in agreement with those of the large proportion of the previous experiments which have given negative results, when an increase of the rate of beat was expected.
(b) Adrenaline.
The addition of adrenaline to the heart results in a general increase in the rate of beat without any alteration of the form of the curve. This effect is shown in Fig. 7. The differences in the form of the logarithmic curves in this figure are within the experimental error. The increase amounted at a maximum to about 20 per cent, of
V puls e ra t
<f
•UIU l pe r!
&
c
1-8 1-6 1-4 1-2 60 50 40 30 20B
•
1-8 1-6 -1-4 -1-2 an ov 50 -40 -30 -20 Q.QO36 0-0035 1-8- 1-6- 1-4- 60- 50- 40- 30-20 O-op34 1010 15" 20 25 °C
Fig. 7. R. etculenta. Effect of adrenaline. Pulse-rate curves of the same heart in Ringer solution (A); the same solution containing adrenaline, 1 in 10* (B); and containing adrenaline, 1 in 5 x 10* (C). B, 2 hours after A; C, i\ hours after B.
[image:12.595.171.424.390.668.2]268 G. S. CARTER
the original rate of beat. A practically complete effect was obtained when the concentration of adrenaline was as high as i in io7, and a smaller effect could be recognised with a concentration of i in io8. All these effects were immediate.
These results are in apparent contradiction to those of Gellhorn (1924), who found a rise of the Qlo of the pulse rate of muscle strips of frog's heart on the
addition of adrenaline. It is impossible to give a complete explanation of this discrepancy, but it may be remarked that the concentration of adrenaline used in his experiments was very large (1 in 1-500,000), and very far outside the concentration in the blood (Schlossmann, 1927, 1 in 2 x io8 at the absolute maximum).
(c) Insulin.
A definite modification of the form of the curve towards the winter condition was observed in the presence of this substance. The effect was, therefore, opposite to that of thyroxine. The effective concentration was higher than that of thyroxine, and the effect was smaller. A concentration of 1 in io8 of the dry substance gave a slight effect in some, but not all, of the experiments: a concentration of 1 in 3 x io5 gave a somewhat larger, but still small, effect. The concentration of insulin in blood is 5-6 units per 100 c.c. (mammal, Brugsch and Horsters, 1930), equivalent to a concentration of 0-2-0-3 mg. of the dry substance per 100 c.c. or 1 in 3-500,000. The necessary change for the production of the effect is therefore large, but not im-possibly so. It is more important that the effect produced was always small, and necessitated an increase of the insulin content of the blood in the winter (cf. p. 263 above). These facts together with the lack of agreement, noted above (p. 257), between the physiological changes of hibernation and those to be expected from an increase of insulin in the circulation, and make it very unlikely that insulin is the natural effective agent in these changes.
(d) A general pituitary extract.
A Ringer extract of a whole pituitary of a frog was made and its effect determined in 250 c.c. of the medium. It produced a definite change in the form of the curve towards the winter condition, and therefore an opposite effect to that of thyroxine. This is shown in Fig. 8. It will be seen that the effect could be reversed by means of thyroxine. The effect was larger and more rapid than could be accounted for by the change in the form of the curve which results from continued beat in Ringer solution alone (p. 262 above), and was considerably greater than the experimental error. It must therefore be believed to be a real effect, but the dilution was great and the substance present must be very active.
(e) Pituitrin.
0-02 unit per c.c. in the Ringer solution circulating through a heart produced no alteration of the curve towards the winter form. The heart responded to potassium bromide, which will be shown in the succeeding paper of this series to transform the curve from the summer into the winter form, after treatment with this pituitary extract.
(/) Extract of the thymus gland.
It would seem improbable that this organ, which normally decreases in size in the vertebrate after puberty, is the effective organ in controlling changes which continue
0-0036 0-0035 0-0034
V
cd i *
<u
3
a
M
pe
r
= i4 1-8
1-6
1-4
1-2
60
50
40
30
20
B
1-8
1-6
1-4
1-2
60
50
-4 0
-30
-20
1-8-
16V 4
- 1-2-
60-
50-
40-
30-2 0
10 25°C
T
10 15 20 25°CFig. 8. R. esculenta. Effect of pituitary extract. Curves given by a heart in Ringer solution (A); the same solution containing pituitary extract (B); the same solution with the addition of thyroxine, i in z x io* (C). B, 2 hours after A; C, three hours after B.
throughout the life of the animal. It may also be remarked that the thymus is also very small in the frog.
[image:14.595.166.467.262.551.2]270 G. S. CARTER
(4) Substances chemically related to thyroxine.
A series of experiments with various substances related more or less closely to thyroxine gave the following results:
(a) Des-iodo-thyroxine. This substance gave a definite effect of the same type as
the thyroxine effect at a concentration of 1 in io8, doubtful effects at 1 in 2 x io6 and no effect at 1 in 5 x io8. At 1 in io8 its effect was as large as the effect normally produced by thyroxine. It was therefore effective, but at a concentration about five times as high as the effective concentration of thyroxine.
(b) Inorganic iodine. Iodine dissolved in five times its concentration of potassium
iodide produced definite effects of the same kind as those of thyroxine in concentra-tions of iodine of 1 in io8 and 1 in 2 x io8, but none, or a doubtful effect, at 1 in 5 x io8. Inorganic iodine was therefore, like des-iodo-thyroxine, effective but at a higher concentration than thyroxine.
(c) Di-iodo-tyrosine. This substance was used in concentrations between 1 in
2-5 x io6 and 1 in 2 x i o \ Its effects were never more than doubtful.
(d) Tyrosine and tyramine had no effect at a concentration of 1 in 2 x io5,
phenyl-ethyl-amine none at concentrations of 1 in 2 x io6 and 1 in io5.
DISCUSSION.
1. The experiments on the effect upon the heart of long-continued beat in unmodified Ringer solution (p. 262 above) show clearly that, if the difference be-tween the summer and winter conditions is due to a change in the endocrine system, the organ responsible must be more active in the summer.
2. Of the endocrine substances and extracts used in the experiments the only one which produced a condition of the heart similar to the natural summer condition by an increase of its concentration in the medium was thyroxine. Thus, the thyroid is the only endocrine organ of those investigated which is capable of controlling the difference between the summer and winter condition in agreement with the observed facts of this difference. Further, the exactness of the similarity between the effect of increased concentration of thyroxine and the observed difference between the summer and winter conditions makes it very probable that the thyroid is not only capable of controlling this change, but does actually do so. This conclusion is further strengthened by the fact that the concentration of thyroxine which was found to be effective in these experiments is approximately equivalent to the change in the concentration of the thyroid secretion which is likely to occur in the circulation.
We must conclude that the control of this seasonal change is endocrine and that the thyroid is the endocrine organ which controls it. This last conclusion is, as we have seen, in agreement with the balance of the evidence provided by previous work. 3. Both insulin and some secretion of the pituitary were found to be capable of producing the opposite effect—a change towards the winter condition by increase of their concentration in the circulation.
should be able to produce changes of the same nature as those produced by thyroxine, although opposite in direction, is a subject into which we need not enter here.
Nor can the pituitary secretion control these changes. The experiments, however, provide evidence for the presence of a substance in the pituitary, distinct from pituitrin, which is somehow related to the thyroid secretion in its action. It will be shown in the succeeding paper of this series that there is evidence for interaction between the thyroid and the pituitary, especially the anterior pituitary. It need not therefore cause surprise that secretions of these glands produce effects which are similar, although opposite.
4. So far we have only considered the temperature-pulse rate curve of the heart and its seasonal changes, and this is by no means the only seasonal change in the physio-logy of the animal. Many of the other seasonal changes are those which might be expected to result from a reduction in the amount of the thyroid secretion in the circu-lation, but it is possible that some of them are controlled in other ways. The experi-ments give no evidence for or against seasonal variations in the activity of any organ of which the secretion has no effect on the form of the temperature-pulse rate curve. However, our consideration of the previous evidence led to the conclusion that there is no reliable evidence for seasonal variations in the activity of any endocrine organ except the thyroid and, perhaps, the adrenal, which Britton believes to play a part in the causation of hibernation in the mammal. The adrenal is therefore the only other organ which we need consider.
It has been stated that no general increase in the rate of beat was observed as the heart passed into the summer condition. If, as Britton suggests for the mammal, the adrenal is in an inactive condition in the winter animal, such an increase in the rate of beat should occur, for our experiments with adrenaline showed that the pulse rate was increased over the whole range of temperature by the addition of this substance. It is possible, however, that the experimental conditions were not suffi-ciently exact to allow us to form a definite conclusion on this point. The action of adrenaline is very rapid, and it is possible that the handling of the frog in killing it, although this was done as quickly as possible, may have stimulated the adrenal to activity before death. If so, hearts taken from winter frogs would have contained more adrenaline than is normal in the winter condition. Thus, it is possible, and may seem probable in view of the general inactivity of the animal in the winter, that the winter condition is characterised by lowered activity of the adrenal, but the experi-ments of this paper give no proof that this is so.
272 G. S. CARTER
SUMMARY.
1. Barcroft and Izquierdo(i93i) have shown that the form of the temperature-pulse rate curve of the excised heart of the frog differs seasonally. The effects produced by endocrine substances and extracts of endocrine glands on the form of this curve have been investigated with a view to determining the nature of the seasonal change within the body.
2. When the heart of a winter frog is allowed to beat for some hours in Ringer solution, there is no alteration in the form of the curve. If the heart is that of a summer frog, there is a gradual alteration of the curve towards the winter form. It is concluded that, if the control of this change is endocrine, the organ responsible is more active in the summer.
3. Thyroxine, at a concentration equivalent to the possible change in the amount of the thyroid secretion in the circulation, transforms the curve given by a winter heart into the summer form, but has no effect on the curve of a summer heart.
4. Adrenaline, pituitrin, thymus extract and extracts of non-endocrine organs, such as muscle and salivary glands, have no effect on the form of the curve.
5. Insulin, in large doses, and a general extract of the pituitary have an effect opposite to that of thyroxine, altering the curve from the summer to the winter form.
6. It is concluded (1) that the control of this phenomenon is endocrine and (2) that the thyroid is the effective organ, by increase of its activity in the summer. Although the adrenal does not control the phenomenon investigated, it is not excluded that it may control other seasonal changes in the frog and other vertebrates. 7. Of several chemical substances related to thyroxine, des-iodo-thyroxine and inorganic iodine produced effects on the form of the curve similar to those of thyroxine, but higher concentrations of these substances were necessary.
REFERENCES.
ADLER, L. (1920 a). Arch. get. Phytiol. 183, 23. (1920 b). Arch. exp. Path. Pharmakol. 86, 159.
BARCROFT, J. and IZQUIERDO, J. J. (1931). Journ. Phytiol. 71, 14s. BRITTON, S. W. (1928). Amer. Journ. Phytiol. 84, 119.
(1930). Amer. Journ. Phytiol. 94, 686. (1931). Amer. Journ. Phytiol. 99, 9.
BRUGSCH, H. and HORSTERS, H. (1930). Arch. exp. Path. Pharmakol. 148, 295. BUGBEE, E. P., SIMOND, A. E. and GRIMES, H. M. (193I). Endocrin. 15, 41. CARTER, G. S. (1932). Journ. Exp. Biol., 9, 264.
CLARK, A. J. (1913). Journ. Phytiol. 47, 66.
DWORKIN, S. D. and FINNEY, W. H. (1927). Amer. Journ. Phytiol. 80, 75. GELLHORN, E. (1924). Arch. get. Physiol. 203, 163.
GORER, P. A. (1930). Biol. Rev. 5, 213.
KENDALL, E. C. and SIMONSEN, D. G. (1928). Journ. Biol. Chem. 80, 357. MANN, F. C. (1916). Amer. Journ. Phytiol. 41, 173.
POCOCK, R. I. (1926). Article in 13th ed. Encyclopedia BriUmmca. RIDDLE, O. and FISHER, W. S. (1925). Amer. Journ. Phytiol. 72, 464. SCHENK, P. (1922). Arch. get. Phytiol. 197, 66.