Lateral Elements Inside Synaptonemal Complex-Like Polycomplexes in
ndt80
Mutants of Yeast Bind DNA
Hasanuzzaman Bhuiyan, Gunilla Dahlfors and Karin Schmekel
1Department of Molecular Biology and Functional Genomics, Stockholm University, SE-106 91 Stockholm, Sweden Manuscript received October 31, 2002
Accepted for publication November 14, 2002
ABSTRACT
The synaptonemal complex (SC) keeps the synapsed homologous chromosomes together during pachy-tene in meiotic prophase I. Structures that resemble stacks of SCs, polycomplexes, are sometimes found before or after pachytene. We have investigatedndt80mutants of yeast, which arrest in pachytene. SCs appear normal in spread chromosome preparations, but are only occasionally found in intact nuclei examined in the electron microscope. Instead, large polycomplexes occur in almost everyndt80mutant nucleus. Immunoelectron microscopy using DNA antibodies show strong preferential labeling to the lateral element parts of the polycomplexes.In situhybridization using chromosome-specific probes confirms that the chromosomes inndt80 mutants are paired and attached to the SCs. Our results suggest that polycomplexes can be involved in binding of chromosomes and possibly also in synapsis.
M
EIOSIS, the process that organizes proper segre- chromosomes. Although polycomplexes are often con-gation of homologous chromosomes into hap- sidered abnormal, they are seen in normal meiosis, as loid germ cells, depends strongly on chromosome pair- has been documented in several species, including yeast. ing and homologous recombination. These two processes The Ndt80 protein is a meiosis-specific transcription are essential for the homologous chromosomes to find factor in yeast that is expressed later than the earliest each other, connect, and remain attached until they recombination genes and that regulates genes during separate during the first meiotic division. The synapto- the middle phase of sporulation (Xuet al.1995). Ndt80p nemal complex (SC) assembles between the homolo- is required for exit from pachytene and for full meiotic gous chromosomes during synapsis and keeps them recombination.paired until crossovers have established physical connec- In the present investigation we confirm that the SCs tions between them. The SC is a protein structure that in spread preparations ofndt80mutants appear similar consists of several components. Two lateral elements to those in wild-type pachytene. Contrary to this impres-(LE), which are dense cores that run along the chromo- sion is the fact thatin situprepared mutant cells essen-somes, are directly attached to the chromosomes. The tially lack visible SCs as seen in electron microscopy two LEs are connected with transverse filaments, while (EM), but contain plenty of massive polycomplexes. The the central element is situated on the transverse fila- LE parts of the polycomplexes are preferentially stained ments and between the LEs (SchmekelandDaneholt by two independent DNA antibodies. Pairing of
homolo-1995). gous chromosomes is confirmed in the mutant strain.
Polycomplexes are aggregates of SC-related material We suggest that this is a situation in which SC entities that are observed before SC formation, during regular inside polycomplexes bind chromosomes.
SC formation, or most commonly, in postpachytene nu-clei (reviewed in Zickler and Kleckner 1999). Al-though the morphology of polycomplexes varies
be-MATERIALS AND METHODS tween species and between situations, polycomplexes
generally appear to be stacks of aligned SCs with the Yeast strains, growth, and spheroplast preparation: The yeast (Saccharomyces cerevisiae) strains used in this study are LEs of the individual SCs fused. Polycomplexes appear
as follows: NKY2310 (a/␣ho::LYS2/ho::LYS2 lys2/lys2 ura3/ frequently in many mutants of yeast that have
deficien-ura3 leu2::hisG/leu2::hisG ndt80⌬::LEU2/ndt80⌬::LEU2 his4X:: cies in chromosome synapsis, recombination, or both.
LEU2-MluI-URA3/his4B::LEU2-MluI), NKY1551 [a/␣ho::LYS2/ The general view is that the LEs of the individual SC- ho::LYS2 lys2/lys2 ura3/ura3 leu2::hisG/leu2::hisG arg4-Nsp/ like subunits in the polycomplex do not attach to the arg4-Bgl his4X::LEU2(Bam)-URA3/his4B::LEU2], and 1151 (a/␣ho::LYS2/ho::LYS2 lys2/lys2 ura3/ura3 leu2::hisG/? COM1:: Myc18::TRP1/COM1::Myc18::TRP1 trp1::hisG/trp1::hisG). The cells were cultured as described in Schmekel(2000). The cells
1Corresponding author:Department of Molecular Biology and
Func-were grown in sporulation medium at 30⬚for 5 hr (pachytene in
tional Genomics, Stockholm University, Svante Arrhenius va¨g 16–18,
the wild-type strains NKY1551 and 1151) or for 8 hr (thendt80
SE-106 91 Stockholm, Sweden.
E-mail: [email protected] mutant strain NKY2310).
EM preparation for serial sectioning: For conventional transmission, EM preparations of spheroplasts were embed-ded as described inSchmekel(2000, p. 111). The sections were collected on copper slot grids coated with formvar and carbon and stained with 5% uranyl acetate in water for 1 hr and then with lead citrate for 1 hr.
Preparation for immuno-EM: Embeddings for immuno-electron microscopy (immuno-EM) included preparation of nuclei according to Schmekel(2000, pp. 111–112), except that the cells were not DNase II treated. For embedding, the protocol in Schmekel (2000) was used with the following modifications: The isolated nuclei were fixed in 2% paraform-aldehyde in 0.1 m2-n-morpholino ethane sulfonic acid (at pH 6.4) containing 0.5msorbitol, for 2 hr at room tempera-ture (RT) and washed. For immunolabeling, the grids were
Figure 1.—Fluorescence images of spread nuclei of wild-incubated on drops of solutions: first for 10 min on 0.02%
type (NKY1551; a and c) andndt80mutant (NKY2310; b and glycine in PBS to inactivate free aldehyde groups, then on
d). (a and b) Immunostaining with the Zip1 antiserum. (c 1% bovine serum albumin (BSA; Sigma A2153) in PBS for
and d) DAPI staining showing the DNA distribution. (b) In blocking, and 1 hr at RT. The grids were then incubated with
most ndt80 mutant nuclei, a majority of the SCs appear as the primary antibody in PBS containing 1% BSA. The sections
separate entities; here all 16 SCs are distinguishable. The arrow were incubated with either of two monoclonal
anti-double-points at a Zip1-staining body, probably a polycomplex. Note stranded DNA antibodies (121-3 from BioGenex, San Ramon,
that this Zip1-staining body does not contain DNA. Bar, 2m. CA, and MAB1293 from Chemicon, Temecula, CA). The cells
were washed in PBS containing 1% BSA and then incubated with a gold-conjugated secondary antibody,
gold/goat-anti-lands). The images were recorded in a confocal microscope mouse (12 nm, Jackson ImmunoResearch Lab, West Grove,
(Zeiss LSM 510). PA) in PBS containing 1% BSA. After washing, the grids were
The antiserum against Zip1 was produced by overexpression fixed in 4% glutaraldehyde and stained with uranyl acetate.
of a part of the ZIP1 gene on a plasmid (the same construct The specimens were examined and photographed in a Zeiss
as that used for the production of the Zip1 antibody published EM 902.
in Sym et al. 1993). The glutathione S-transferase-tagged
Immunofluorescence andin situhybridization:The
immu-C-terminal part of the Zip1 protein was purified and injected nofluorescence experiments were performed with spread
into a rabbit. The serum (2460) stains SCs in spread prepara-preparations on glass slides (described byLoidlet al.1998).
tions of meiotic nuclei and SCs in EM preparations and gives The slides were incubated for 30 min with 3% BSA in PBS
a single band in Western blots. The immunofluorescence and at RT and then incubated with a Zip1 antibody (2460; see
confocal microscopic images were processed using Adobe Pho-description below) at a dilution of 1:100 in 1% BSA in PBS
toshop, version 6.0. for 1 hr. The slides were washed in PBS and, as a last step
before the secondary antibody incubation, in 1% BSA in PBS. Secondary antibody incubation was carried out with a
FITC-conjugated swine-anti-rabbit serum (Dako, Glostrup, Denmark) RESULTS for 30 min. DNA was detected by 4⬘,6-diamidino-2-phenylindole
Thendt80mutant SCs appear as wild type in spreads:
(DAPI) staining. After washing in PBS, the slides were inspected
in a fluorescence microscope (Zeiss Axioplan 2). As earlier documented, the ndt80 mutant yeast cells The fluorescencein situhybridization (FISH) experiments arrest in pachytene and appear more or less like wild were carried out according toWeinerandKleckner(1994) type, as seen in spread preparations for light microscopy with minor modifications. The spreading was carried out
ac-and EM (Xuet al.1995). We immunostained spreadings cording to Loidlet al.(1998). In brief, slides were treated
ofndt80mutant and wild-type cells with a Zip1 antiserum with RNase A instead of both RNase A and RNase T. After
for immunofluorescence microscopy (IF) and found that denaturation the probe was applied to the slide and incubated
for hybridization. Final washing was carried out in 1⫻SSC the SCs appeared similar in the two preparations. The and 0.1⫻SSC. To enhance the hybridization signal, hybridiza- ndt80mutant cells had been cultivated in sporulation tion was repeated on one occasion. The two probes were
medium for 8 hr and had thus been arrested in pachy-produced from large pieces of chromosomes inserted into
tene for 3 hr. In almost all nuclei, SCs were clearly cosmids that were amplified in bacteria. One probe was from
seen as individual entities (Figure 1, a and b), which a subtelomeric region of chromosome III [c9171; American
Type Culture Collection (ATCC) no. 70884], and the other colocalized with the labeling of DNA (Figure 1, c and was an interstitial part of chromosome VIII (c9315; ATCC no. d). In most nuclei of thendt80mutant we found bodies 71216). The probe was purified using a Wizard plus midiprep that were stained with the Zip1 antibody, which may be DNA purification kit (Promega, Madison, WI) and
digoxi-polycomplexes (Figure 1b, arrow). These Zip1 staining genin labeled by nick translation according to standard
proce-bodies were sometimes stained with DAPI, but usually dures (Boehringer Mannheim, Indianapolis). The probe was
detected using an antibody against digoxigenin (Boehringer that was not the case (Figure 2, a and b). Our overall Mannheim) as above and then by using a secondary antibody impression is that thendt80cells appear similar to wild-labeled with TRITC (Jackson ImmunoResearch Laboratories). type cells in their SC pattern.
The Zip1 protein was detected using the 2460 serum (see
Intact cells of the ndt80mutant lack visible SCs but
below) and a secondary antibody labeled with Cy5 ( Jackson
contain large polycomplexes: Spheroplasts that were ImmunoResearch Laboratories). DNA was detected using a
200). Eighteen percent of the nuclei contained poly-complexes, but since only 33% of the sections through a polycomplex-containing nucleus contain a piece of polycomplex, the actual number of nuclei that contain polycomplexes is 55%. A similar fraction of nuclei con-tained single pieces of SCs.
We have looked briefly at single sections of ndt80 mutant nuclei harvested after 5 and 6 hr in sporulation medium, but we have not done thorough investigations Figure 2.—Fluorescence images of spread nuclei of the
ndt80mutant (NKY2310) immunostained with (a) Zip1-antise- (i.e., serial sectioning) at these time points. At the earlier rum and (b) DAPI. Some nuclei show Zip1 staining that is time points we found both SCs and polycomplexes, and similar to wild type (top cell of the three), while others show
the impression is that free SCs are more frequent and a body that stains heavily with the Zip1 antibody (the two
polycomplexes less frequent in those nuclei than in cells nuclei in the bottom). In many cells, the body is negative for
harvested after 8 hr. DNA staining by DAPI (the nucleus to the left, solid
arrow-head), and in a few nuclei the body is stained by DAPI (the In conclusion, very few single SCs are visible in the nucleus to the right, open arrowhead). Bar, 6m. ndt80mutant nuclei that have been arrested in
pachy-tene for 3 hr, but most cells contain polycomplexes.
The SCs in the polycomplex are associated with DNA:
were embedded for conventional EM. Ultrathin sections
To further understand the chromosome pairing situa-of wild-type nuclei in pachytene revealed individual, short
tion and the nature of these polycomplexes, we wanted pieces of SCs at pachytene with chromatin attached
(Fig-to investigate the presence of DNA in the complexes. ure 3a). Surprisingly, thendt80mutant cells, on the other
Ultrathin sections embedded for immunolabeling were hand, showed massive polycomplexes in many nuclei
incubated with either of two commercial antibodies (Figure 3, b and c). In the wild-type cells, polycomplexes
(Chemicon and Biogenix) that bind to double-stranded containing two or three SCs were occasionally found in
DNA and then with a secondary antibody conjugated a single section (data not shown).
with 12-nm gold particles. Gold particles were located ndt80 mutant cells were serially sectioned,
docu-in the chromatdocu-in docu-in general, docu-in the nuclei and also docu-in mented in the EM, and reconstructed (the data are
polycomplexes (Figure 4a). The gold particles in the summarized in Table 1). To prevent biased selection,
polycomplexes were recorded and their precise location SC-containing nuclei, nuclei containing polycomplexes,
in the polycomplex was noted. Both antibodies showed and nuclei lacking any of the structures were chosen as
strong preference for LE labeling (see Table 2). We starting sections. We found that 14 of the 16 nuclei
noted that the outermost LE in a polycomplex appeared contained massive polycomplexes. Four of the
polycom-to have stronger labeling than the more centrally situ-plex-containing nuclei showed no free SCs, 8 nuclei had
ated LEs and wanted to eliminate the possibility that between one and five pieces of visible SCs in addition to
the high LE labeling was due to the presence of more the polycomplex, and 2 polycomplex-containing nuclei
DNA in those LEs. We also wanted to eliminate the had seven SCs. Of the 2 nuclei that did not contain
possibility that gold particles may represent the binding polycomplexes, 1 contained no SCs at all and the other
of the primary antibody to chromatin outside of the contained two pieces of SC. Polycomplex-containing
polycomplex. In the second counting, we therefore sections were counted and compared with the total
eliminated the gold particles in the outermost LEs and number of sections through a nucleus to determine the
the outermost 30 nm of the polycomplexes in the trans-fraction of sections that was occupied by polycomplexes.
verse direction (see dotted square in Figure 4a). The On average, 33% of the sections through a
polycomplex-limitation of 30 nm was chosen because this is the maxi-containing nucleus contained polycomplexes.
mum distance from the antigen to the gold particle The number of nuclei that contain polycomplexes
was counted in randomly selected single sections (N⫽ (BaschongandWrigley1990). Also under these
TABLE 1
Polycomplex and SC content in 16 serially sectioned nuclei of thendt80mutant
No. of Several No. of SC with No. of Nucleus Status in Completely No. of Amount PC-containing PC undetected PC sections identifier start sectiona sectioned? sections of PCs sections entities attachment lost
PC4A Contains PC Almostb 20 ⫹⫹⫹ 10 ⫹ — 1
PC2A — ⫹ 11 ⫹⫹⫹ 8 ⫺ —
PC1B — Almostb 11 ⫹⫹⫹ 4 ⫹ —
PC7B Contains PC ⫹ 13 ⫹⫹⫹ 5 ⫺ 1 2
PC8A — Almostb 23 ⫹⫹⫹ 9 ⫹ 2 3
PC6A Contains PC ⫹ 18 ⫹⫹⫹ 6 ⫺ 2 3
PC1D — ⫺ 13 ⫹⫹⫹ 10 ⫺ 2
PC1C Contains SC ⫹ 18 ⫹⫹⫹ 9 ⫺ 3
PC5A Contains PC Almostb 17 ⫹⫹⫹ 5 ⫺ 4 5
PC7A Contains SC ⫹ 22 ⫹⫹⫹ 8 ⫺ 5 2
PC3A Contains PC ⫹ 20 ⫹⫹⫹ 7 ⫺ 7
PC8B Contains PC Almostb 21 ⫹⫹ 5 ⫺ 4 4
PC4B Contains SC Almostb 17 ⫹ 2 ⫺ 7 2
PC5B — ⫹ 18 ⫹ 3 ⫹ — 4
PC3B — ⫹ 21 ⫺ ⫺ ⫺ 2
PC6B — ⫹ 21 ⫺ ⫺ ⫺ — 3
Summary of the serial sectioning ofndt80nuclei.
aFor practical reasons, a totally random choice of nuclei for serial sectioning and reconstruction is hardly possible. To get
nuclei that were representative of the population of cells, three different kinds of nuclei were selected on the basis of their features in the starting section, which is always more or less in the center of the nucleus. The nuclei in the starting section contained polycomplexes (PC), SC, or none of the two structures.
bSome nuclei were not completely sectioned because the series did not cover the complete nucleus. All nuclei in the series
except one (PC1D) were completely or almost completely sectioned. The number of sections that were missing, on the basis of the diameter of the nucleus in the last section, was estimated to be three or fewer per such nucleus.
ditions the labeling of the LEs was much stronger than LEs, was very low even with high antibody concentration. One of the antibodies, the Biogenix, was totally negative that of the CE and the space between.
Immuno-EM labeling of SCs in wild-type cells (the after DNase treatment (data not shown). Thus, the im-muno-EM investigation shows presence of DNA in the strain 1151) resulted in preferential labeling of
chroma-tin and LEs,i.e., less in the central element area (Figure LE part of the SCs inside the polycomplexes.
The chromosomes are paired inndt80nuclei:To see if 4b). The SC morphology is less well preserved in the
LR White-embedded cells than in the Agar 100 Resin- the SCs in thendt80mutant cells keep the chromosomes paired/synapsed, we performed FISH experiments us-embedded cells, which are fixed with glutaraldehyde
(Figure 3a), but the structure can be identified. Negative ing chromosome-specific DNA probes. If the homolo-gous chromosomes are paired, the signals should be controls in which preparations of embedded nuclei
were immunostained with the secondary antibody, in fused into one dot in the confocal microscope (i.e.,⬍0.7 m apart; Weiner and Kleckner 1994), and if the the absence of a first antibody, showed absence of
label-ing. In nuclei that had been DNase treated, the overall homologous chromosomes are not paired, two separate signals should appear. Hybridizations were done with nuclear antibody staining, including the staining of the
TABLE 3 TABLE 2
DNA antibody localization in different parts of the Chromosome pairing measured by FISH labeling with chromosomal markers
SC inside polycomplexes
Between bg
Pairing (0 dots/ Antiserum/ In LE LE and CE In CE
recorded area (%) (%) (%) N (one dot) ⬎2 dots) Pairing—
Strain/location (%) (%) bga(%) N
Chemicon/total PC 85 8 7 189
WT/chromosome III 47 7/9 56 180
Biogenix/total PC 77 12 11 57
WT/chromosome VIII 48 10/12 62 129
Chemicon/restricted
⌬ndt80/chromosome III 67 4/4 73 165
areaa 71 16 13 80
⌬ndt80/chromosome VIII 56 4/11 66 259 Biogenix/restricted
areaa 68 18 14 28
bg, background labeling. Strains: WT, NKY1551; ⌬ndt80, NKY2310; chromosome III, probe made from cosmid ATCC
aThe outermost LEs in the PC are eliminated as well as 30
no. 70884 on chromosome III; chromosome VIII, probe made nm from the end of the polycomplexes in transverse direction
from cosmid ATCC no. 71216 on chromosome VIII.Nis the (see Figure 4a).Nis the number of gold particles inside
poly-number of recorded nuclei. complex. For information about antisera, seematerials and
aThe number of nuclei with one dot plus the number of methods.
nuclei with two dots are set to 100%.
the ATCC no. 70884 probe (subtelomeric, chromosome
III) and with the ATCC no. 71216 probe (interstitial, tene and the accumulation of SC-containing cells (Xu chromosome VIII) on wild-type cells (NKY1551) and et al.1995). These observations have been done in hypo-⌬ndt80cells (NKY2310). Thendt80mutant cells showed tonically burst cells with the nuclei spread over a surface. a high level of pairing (the result is summarized in Table In the same kind of preparations of mutant cells, har-3). Considering the fact that the wild-type cells are less vested after 8 hr in sporulation medium, we also find synchronized than the arrestedndt80mutant cells, it is wild-type-like SC formation. In addition, larger entities, not surprising that the mutants have a slightly higher possibly polycomplexes, are stained by the Zip1-anti-degree of paired chromosomes. The SCs were well pre- body. We see that some, but not most, of the possible served in the FISH preparations and could be readily
polycomplexes contain DNA. identified as outlined by the anti-Zip1 antibody staining
Although thendt80mutants contain normal numbers (Figure 5). The localization of the probes was often
of apparently normal SCs and although their homolo-distinctly seen at the location of the SCs, indicating that
gous chromosomes are paired, we find almost no SCs the SCs are attached to the chromosomes. Thus, the
in the EM in these cells. The cells were fixed and embed-chromosome pairing in thendt80mutant appears like
ded according to a procedure that gives good SC mor-that in wild type.
phology in wild-type cells. Instead we find massive poly-complexes in most mutant nuclei. We consider the most DISCUSSION likely interpretation of our results to be that the SC entities in the polycomplexes seen in the EM are the The phenotype of the ndt80 null mutants has
pre-same as the SCs seen in spreads. The discrepancy be-viously been described, including the arrest in
pachy-tween the SC configuration in the two situations, solitary SCs in spreadsvs.polycomplexes in intact cells, may be due to differences in specimen preparation methods. The spreading method is harsher than that of the EM embedding and may cause disruption of the polycom-plexes, especially if the SCs are loosely connected.
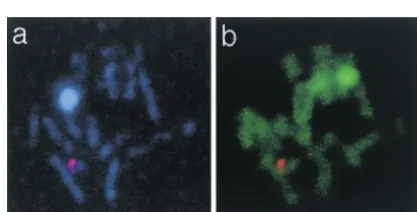
One may think that the lack of visible SCs in the EM preparations of thendt80mutant is due to loss of morphological integrity of the SCs. However, the SC proteins cannot have been shed off the chromosomes, since at least one major SC component, the Zip1 pro-tein, is still present in structures that appear as normal Figure 5.—FISH experiment of spread preparations of
ndt80mutant cells combined with DNA labeling (green) and SCs in spreads. It further seems unlikely that the SC Zip1 labeling (light blue). (a) A nucleus with SCs labeled proteins have adopted an alternative organization inside with the Zip1 antibody and probe 70884 labeling one end
the SC, because several examples ofndt80mutant SCs of chromosome III (red), showing the close location of the
are visible in EM and these look morphologically nor-homologous sites on the two chromosomes. (b) The same
filament-LE organization is seen in the polycomplexes that aggregation of SCs into chromosome-connected polycomplexes is part of a normal stage in the meiotic that reside inside the mutant nuclei.
chromosome metabolism. The polycomplexes, as they We find it likely that the SCs inside the polycomplexes
appear in thendt80mutant, may be transient and less bind chromosomes. The fact that the LEs inside the
pronounced in wild-type cells. The formation of poly-polycomplexes are preferentially stained with DNA
anti-complexes may be a transition from pachytene to later bodies strongly argues for the presence of chromatin
meiotic stages, perhaps involved in chromosome orien-inside the polycomplexes. Since the homologous
chro-tation. mosomes are closely situated in the arrestedndt80
nu-clei, as shown by FISH chromosomal labeling on the We thank Dr. Nancy Kleckner and Dr. Franz Klein for yeast strains and Dr. Harry Scherthan for advice on the FISH method. We are
SCs in spreads, we find it reasonable to believe that
grateful to Dr. Shirleen Roeder for giving us the ZIP1 construct from
chromosomes inside the polycomplexes are synapsed.
which we produced the antibody.Dr. Eva Bratt has generously given
The DNA-containing Zip1 complexes that are seen in technical advice on confocal microscopy. We thank Drs. George Far-spread preparations may be remnants of the larger rants and Christer Ho¨o¨g for constructive remarks on the manuscript. The project was supported by the Swedish Cancer Society, the
Nilsson-DNA-containing polycomplexes that we see in EM.
Ehle Foundation, and the Borgstro¨m Foundation.
The polycomplexes in the ndt80 mutant probably originate from previously normal, solitary SCs that have later aggregated into polycomplexes. Preliminary
obser-LITERATURE CITED vations that we have done at earlier time points indicate
Baschong, W., andN. G. Wrigley, 1990 Small colloidal gold
conju-that the ratio of polycomplexes to solitary SCs gradually
gated to Fab fragments or to immunoglobulin G as
high-resolu-increases with time during the pachytene arrest. tion labels for electron microscopy: a technical overview. J. Elec-tron Microsc. Tech.14:313–323.
Polycomplexes have usually been regarded either as
Goldstein, P., 1987 Multiple synaptonemal complexes
(polycom-aggregates of SC proteins under decay during stages plexes): origin, structure and function. Cell Biol. Int. Rep.11: when the SC disintegrates or as abnormal manifestations 759–796.
Loidl, J., F. KleinandJ. Engebrecht, 1998 Genetic and
morpho-of meiotic mutations (reviewed in Goldstein 1987;
logical approaches for the analysis of meiotic chromosomes in
ZicklerandKleckner1999). However, polycomplexes yeast. Methods Cell. Biol.53:257–285.
Schmekel, K., 2000 Methods for immuno-electron microscopic and
occur in wild-type cells, including yeast, during meiotic
fine analysis of synaptonemal complexes and nodules in yeast.
prophase I (Zickler and Olson 1975; Zickler and Chromosoma109:110–116.
Kleckner1999; our own observations). In intact wild- Schmekel, K., andB. Daneholt, 1995 The central region of the synaptonemal complex revealed in three dimensions. Trends Cell
type yeast cells of the SK1 background, we occasionally
Biol.5:239–242.
observed polycomplexes containing two or three SC Sym, M., J. A. EngebrechtandG. S. Roeder, 1993 ZIP1 is a synapto-components in the EM. nemal complex protein required for meiotic chromosome
synap-sis. Cell72:365–378.
Yeast cells go through meiotic prophase I rapidly.
Weiner, B. M., andN. Kleckner, 1994 Chromosome pairing via
The meiotic stages between pachytene and metaphase multiple interstitial interactions before and during meiosis in
yeast. Cell77:977–991.
(diplotene and diakinesis), in which the chromosomes
Xu, L., M. Ajimura, R. Padmore, C. KleinandN. Kleckner, 1995
gradually condense and organize into metaphase, are NDT80, a meiosis-specific gene required for exit from pachytene not very well defined in yeast as to chromosomal mor- in Saccharomyces cerevisiae. Mol. Cell. Biol.15:6572–6581.
Zickler, D., andN. Kleckner, 1999 Meiotic chromosomes:
integrat-phology. However, in a detailed EM study of wild-type
ing structure and function. Annu. Rev. Genet.33:603–754.
(tetraploid) yeast meiosis, diplotene nuclei are de- Zickler, D., andL. W. Olson, 1975 The synaptonemal complex
and the spindle plaque during meiosis in yeast. Chromosoma50:
scribed as lacking pachytene-like SCs and as having some
1–23.
SCs with thickened LEs and with polycomplexes in 30%