“ALTERNATE-2” DISJUNCTION DOES NOT EXIST
IAN A. BOUSSY
Department of Genetics University of California Davis, California 95616
Manuscript received October 19, 1981 Revised copy accepted December 8, 1981
ABSTRACT
JOHN and LEWIS (1965) proposed a model of disjunction of translocation heterozygotes that defined two types of alternate disjunction, alternate-I and alternate-2, differing from the classic view of only one type. ENDRIZZI
(1974), and subsequent workers citing him, claim to have observed two distinct alternate disjunction configurations, corresponding to JOHN and LEWIS’S alternate1 and alternate2 types, in meiotic preparations of several species. These observations are based on a two-dimensional interpretation of the three-dimensional phenomenon of disjunction, and are erroneous. I n each case the two supposed types are topologically identical. There is only one possible alternate disjunction configuration.
THE
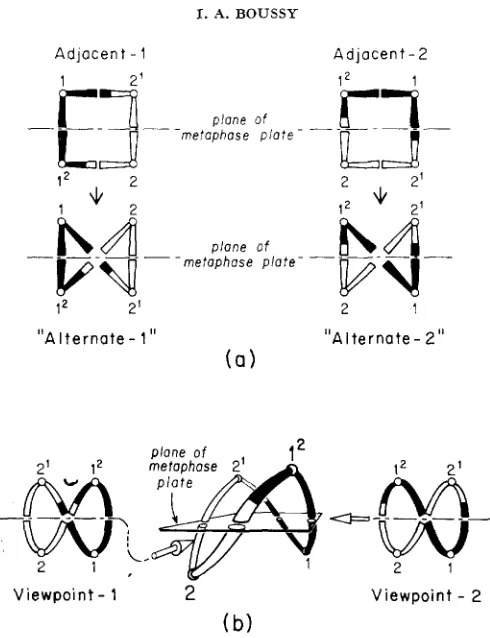
purpose of this note is to correct the misconception that two morpho- logically distinct types of alternate disjunction are discernible in meiosis of a chromosomal translocation heterozygote, and to discuss briefly the model upon which the misconception is based. The misconception has become in- creasingly prevalent in the cytogenetics literature. Three possible types of dis- junction (adjacent-I, adjacent-2, and alternate, named after the relative positions of centromeres in the quadrivalent ring at meiotic metaphase I) are clearly defined and discussed in many general cytogenetics texts (see, for ex- ample, BROWN 1972). The three types are the possible ways to group four ob- jects (the chromosomes involved) into two groups of two. In the absence of special relationships between the chromosomes, all three types are expected to occur with equal frequency.Rejecting this simple probabilistic analysis, JOHN and LEWIS (1965) argued that each type of adjacent disjunction should have a corresponding alternate disjunction. They reasoned that chromosomes do not assort completely at ran- dom from a multivalent. Rather, the primary event is co-orientation, in which two centromeres orient with respect to each other in order to separate to oppo- site poles. They thus proposed that there are four types of disjunction. Adja- cent-1 disjunction occurs when homologous centromeres form co-oriented pairs and disjoin from an “open-ring’’ configuration, so that nonhomologous centro- meres go to a pole together ( e . g . , in Figure la, 1 disjoins from l 2 and 2 dis- joins from 2l) ; and adjacent-2 occurs when nonhomologous centromeres form
Adjacent - 1
1 7‘
Adjacent - 2
1 2 1
metaphase p l a t e -
L A
L A
1‘ , 2 2 , 2’
“ A l t e r n a t e - 1 “ “Alternate- 2 ” ( a )
Viewpoint
-
1 2 Viewpoint - 2(b)
FIGURE 1.-a) The two types of adjacent disjunction and their purportedly corresponding alternate disjunctions, viewed at metaphase; after JOHN and LEWIS 1965; b) Alternate dis- junction and two-dimensional projections as viewed from two positions 90” apart on the plane of the metaphase plate; viewpoint-I (inverted) corresponds in all particulars to ‘Lalternate-l,” as does viewpoint-2 to “alternate-2.’’
co-oriented pairs and disjoin from an open ring configuration, so that homolo- gous centromeres go to a pole together (in Figure la, l 2 disjoins from 2 and 1 disjoins from 2l). Alternate disjunction is then the result of a subsequent “twist” in the ring, after the primary co-orientation pairing event, to form a “zig-zag” configuration. Thus
JOHN
and LEWIS derived alternate disjunctions from two different origins: “alternate-1,’’ in which the primary event is co- orientation for disjunction of homologous centromeres, with the “twist” involv- ing the translocated arms (in Figure la, 1 disjoins from 12, 2 disjoins from 2l); and “alternate-2,” in which the primary event is co-orientation and disjunction of nonhomologous centromeres, with the “twist” involving the nontranslocated arms (in Figure la, l 2 disjoins from 2, 1 from 2l). Since the “twist” is not a result of the primary co-orientation, JOHN and LEWIS considered ring and zig- zag configurations as equally likely, and predicted ratios of 1 adjacent-1 : 1ALTERNATE-2 DOES NOT EXIST 507
the frequencies of all four classes should be equal. SYBENGA (1972, 1975) de- veloped the same model of co-orientation as the primary event of meiosis.
However their model is not convincing. If the co-orientation and repulsion of centromeres is the primary event of disjunction, then JOHN and LEWIS’S model errs in not considering that each centromere in a quadrivalent ring is con- nected to two other centromeres, and thus that repulsion from both its neigh- bors must affect its disjunction, as suggested by GAIRDNER and DARLINGTON
(1931). Such repulsion should favor alternate disjunction, in which each cen- tromere disjoins from both its neighbors, over adjacent disjunction, in which two neighboring centromeres proceed together to a pole (this agrees with many reports of “directed disjunction?’ in the literature). But this then is a model of centromeres with “repulsion coefficients” between neighbors. If the coefficients are negligible, disjunction is random, with equal frequencies of alternate, adja- cent-1, and adjacent-2. Such repulsion coefficients should be related to the de- gree of synapsis, the lengths of arms and of translocated segments, and to the occurrence and behavior of chiasmata. Thus the process of meiosis of a trans- location heterozygote is complex, and simple models may be inadequate to de- scribe it. (Both JOHN and LEWIS (1965) and SYBENGA (1975) discuss the fac- tors which may be involved, and JOHN and LEWIS emphasize that “it is not homology per se [of centromeres] that matters but the morphology of associa- tion” (1965, p. 83) .)
Neither JOHN and LEWIS nor SYBENGA claimed that the two types of alter- nate disjunction produced physically different configurations, although their two-dimensional figures (as Figure l a ) seem to portray alternate-I and alter- nate-2 as different. In the cell, of course, the metaphase ring forms in three- dimensional space. If a single metaphase ring undergoing alternate disjunction is viewed from two different positions 90° apart on the plane of the metaphase plate, the two views will appear to be JOHN and LEWIS’S “alternate-1” and “alternate-2” disjunctions (see Figure l b )
.
In three dimensions, “alternate-1” and “alternate-2” are topologically identical, and the “twist” is seen to be an artifact of collapsing a three-dimensional phenomenon to two dimensions. Thus even if JOHN and LEWIS’S assertion of two pathways to the formation of alternate disjunction were true, it still would be impossible to differentiate between the two, based on viewing the metaphase disjunction.the “existence of alternate-I and alternate-2 types
. . .
beyond any reasonable doubt.. .”
(1976, p. 133) in the German cockroach [Blattella germanica (L.)]. H e felt that “it is clear that the long-held view that disjunction in translocation heterozygotes consists of [only] alternate, adjacent-1 and adjacent-2 types. . .
is no longer tenable” (1976, p. 133). LACADENA and CANDELA (1977) con- curred, and described the presence of the two types of alternate disjunction inrye (Secale cereale L.), although their data did not agree with the predicted
frequencies of the disjunction types, and they therefore questioned the con- ceptual basis for JOHN and LEWIS’S model. KODURU (1979) claimed to be able to distinguish the two types of alternate disjunction in pearl millet (Penni-
setum americanum Schum.)
,
and discussed at length the frequencies of dis-junction types that he observed. LAMM (1981) claimed to be able to discriminate between the two alternate disjunction types in the garden pea
(Pisum satiuum L.)
.
I n all the above papers, the observations of alternate-1and alternate-2 disjunctions must have involved observing alternate disjunctions from different angles. It follows that discussions based on these observations are erroneous.
LACADENA and CANDELA (1 977) unknowingly demonstrated quite clearly that the supposed two types of alternate disjunction were one and the same, when they noted that “it is possible to change (by squashing) an alternate-I configuration into a n alternate-I1 one” (1977, p. 187). Their squashing ap- parently rotated the configuration 90”.
The misconception outlined above is not confined to the primary literature, but has found its way into at least one recent cytogenetics text (SCHULZ- SCHAEFFER, 1980, p. 226, citing ENDRIZZI, 1974). I hope that this note will correct the situation. There is no basis for the claim that two types of alter- nate disjunction can be seen at meiotic metaphase in translocation heterozy- gotes.
I would like to thank JERRY COYNE and TIMOTHY PROUT and an anonymous reviewer for critical comment. This work was partially supported by National Institute of Health grant GM 22221 to the University of California a t Davis.
LITERATURE CITED
BROWN, M. S., M. Y. MENZEL, C. A. HASENKAMPF and S. NAQI, 1981 Chromosome configur- ations and orientations in 58 heterozygous translocations in Gossypium hirsutum. J. Hered.
Textbook of cytogenetics. C. V. Mosby, St. Louis, MO. 72: 161-168.
BROWN, W. V., 1972
COCHRAN, D. G., 1976 Disjunction types and their frequencies in two heterozygous reciprocal
COCHRAN, D. G., 1977 Patterns of disjunction frequencies in heterozygous reciprocal translo- cations from the German cockroach. Chromosoma 62: 191-198.
ENDRIZZI, J. E., 1974 Alternate-I and alternate-2 disjunctions in heterozygous reciprocal translocations. Genetics 77: 55-60.
GAIRDNER, A. E. and C. D. DARLINGTON, 1931 Ring-formation in diploid and polyploid
Campanula persicifolia. Genetica 13 : 113-150.
ALTERNATE-2 DOES NOT EXIST 509
JOHN, B. and K. R. LEWIS, 1965
KODURU, P. R. K., 1979 Metaphase I centromere co-orientation in interchange heterozygotes
LACADENA, J. R. and M. CANDELA, 1977 Centromere co-orientation at metaphase I in inter-
LAMM, R., 1981 Giemsa C-banding and silver-staining for cytological studies in Pisum.
SCHULZ-SCHAEFFER, J., 1980 SYBENGA, J., 1972
SYBENGA, J., 1975
The meiotic system. Protoplasmatologia 6: 1-335.
of pearl millet. Genet. Res., Camb. 34: 69-76.
change heterozygotes of rye, Secale cerale L. Chromosoma 64: 175-189.
Hereditas 94: 45-52.
Cytogenetics. Springer-Verlag, New York.
General cytogenetics. American Elsevier, New York.
Meiotic configurntiom. Springer-Verlag, New York.