Copyright2001 by the Genetics Society of America
Sequence and Chromosomal Context Effects on Variegated Expression of
Keratin 5/lacZ Constructs in Stratified Epithelia of Transgenic Mice
Angel Ramı´rez,* Eric Milot,
†Immaculada Ponsa,
‡Camelia Marcos-Gutie´rrez,
§Angustias Page,*
Mirentxu Santos,* Jose´ Jorcano* and Miguel Vidal
§*Cell and Molecular Biology, Centro Investigaciones Medio Ambientales y Energeticas (CIEMAT), 28040 Madrid, Spain,†Department of Cell Biology, Medical Genetics Center, Erasmus University, 3000 DR Rotterdam, The Netherlands,‡Departament de Biologia Cellular,
Fisiologia i Immunologia, Facultat de Medicina, Universitat Auto`noma de Barcelona, E-08193 Bellaterra, Spain and §Department of Developmental and Cell Biology, Centro Investigaciones Biolo´gicas, 28006 Madrid, Spain
Manuscript received June 1, 2000 Accepted for publication January 30, 2001
ABSTRACT
The expression of transgene loci in mammals often occurs in a heterocellular fashion resulting in variegated patterns of expression. We have examined the effect of chromosomal integration site, copy number, and transcriptionally activating sequences on the variegation of akeratin 5-lacZ(K5Z) construct in the stratified epithelia of transgenic mice.lacZexpression in these mice is always mosaic, and the-gal activity per cell is usually higher in the lines with a higher proportion of expressing cells. Similar constructs, in which cDNAs were exchanged bylacZsequences, showed no variegation. Also, when a strongly active, nonvariegating construct was coinjected withK5Z, most transgenic lines showed an almost homogeneous lacZexpression. The comparison of transgene arrays of different copies inserted at the same locus (obtained by using a lox/Cre system) showed that the reduction of copy number does not lead to an increase in the proportion of cells that express the transgene. Finally, in most of the variegating or nonexpressing lines the transgenes were located both at intermediate positions and at peritelomeric regions in the long chromosome arms. These findings suggest that the probability and efficiency of expression ofK5Zgenes depend on both long range chromosomal influences and on sequences in the transgene array.
G
ERMLINE transformation of mice has been invalu- et al.1993), although the mechanisms responsible for such silencing events are still poorly understood. able in understanding processes that control geneexpression and has led to the identification of tissue- Chromosomal context is important for variegation, shown by the mosaic expression patterns of genes lo-specific control elements (GrosveldandKollias1992).
cated near or at pericentromeric heterochromatin in However, a common feature of transgenic mice is the
Drosophila (Henikoff1990). No systematic analysis of wide variation in expression levels among individuals
the integration site of DNA constructs has been carried carrying the same transgene (Grosveld and Kollias
out in transgenic mice, although integration at pericen-1992). In addition, quite often DNA constructs express
tromeric regions has been reported with unexpected in a mosaic or variegated fashion, by which transgene
frequency (Dobieet al. 1996;Festenstein et al. 1996; expression occurs in some, but not all, the cells within
Milotet al. 1996). As observed before in Drosophila a population of equivalent transgene-bearing cells (
Tho-(DorerandHenikoff1994) and in plants (Assaadet
reyet al.1993;PorterandMeyer1994;Ramı´rezet al.
al.1993), variegation of DNA constructs in mice is re-1994;Elliottet al.1995;Dobieet al.1996;Festenstein
lated to the repeated nature of multicopy transgene
et al.1996;Milotet al.1996;Morleyet al.1996;
Grau-arrays (Garricket al.1998), although a direct relation
bertet al.1998). Thus, the transcriptional potential of
between copy number and proportion of expressing a given construct is controlled by more than just itscis
-cells is not always observed (Festenstein et al. 1996; control elements. Recent work by Whitelaw, Martin, and
Zhuma et al. 1999). Also, variegation correlates with colleagues suggest that the variability of expression
lev-chromatin accessibility but not with DNA methylation els among transgenic lines results from the differences
(Garricket al.1996). However, the presence in trans-in the proportion of cells that express the transgene
genic constructs of intact locus control regions (LCRs), (Robertsonet al.1995). The variegated expression also
which have a dominant activity, usually results in homo-occurs in Drosophila (Henikoff1990), yeasts (
Gottsch-geneous, nonmosaic expression patterns irrespective of
linget al.1990;Allshireet al.1994), and plants (Meyer
the integration site (Milot et al. 1996; Kioussis and
Festenstein 1997).
The above features of variegating transgenes in mice
Corresponding author:Miguel Vidal, Department of Developmental
derive mostly from constructs expressed in
hematopoi-and Cell Biology, Centro de Investigaciones Biolo´gicas, Vela´zquez
144, 28006 Madrid, Spain. E-mail: [email protected] etic tissues (Elliottet al.1995;Robertsonet al.1995;
DNA fluorescence in situ hybridization (FISH) analysis: Festensteinet al.1996;Garricket al.1998;Graubert
DNA FISH was performed as described (Milotet al.1996).
et al. 1998). In this report we have studied transgene variegation in stratified epithelia, such as those that line the surface of the skin and parts of the digestive tract
RESULTS
(FuchsandWeber1994). We have used thelacZgene
Variegated expression ofK5Zconstructs in transgenic
ofEscherichia colifused to sequences from the keratinK5
mice: The basicK5Z construct used consists of 5.3 kb gene, which encodes one of the intermediate filament
of 5⬘ flanking and promoter sequences of the bovine polypeptides specific to the basal layer of stratified
epi-K5gene fused to alacZgene ofE. coli(Figure 1). Only thelia. The remarkable tendency of theseK5-lacZ(K5Z)
three lines (B4, B7, and B38) showed a detectable ex-constructs to express in a mosaic fashion allowed us to
pression and were used to derive stable lines. Analysis ask whether copy number and chromosomal integration
of sections of adult tissues (4- to 8-week-old animals) site are major determinants of variegation and how such
showed restricted expression of the transgene to strati-variegation can be affected by the presence of
transcrip-fied epithelia and their appendages, like that of the tionally active sequences. The results show that
copy-endogenousK5gene. A discrepancy, however, was the number reductions do not affect variegation of K5Z
mosaic expression of the transgene in all epithelia of constructs and that position effects are imposed by
telo-every animal of lines B4, B38 (Figure 2A), and B7 (not meric regions. We also find a variability in
-galactosi-shown). Detection of-gal activity was done using vari-dase (-gal) activity levels per cell between different
ous conditions that allowed us to rule out that mosaic variegating lines and that the presence of strong
activat-expression was due to trivial reasons, such as differential ing sequences turns variegation into a homogeneous
accessibility of substrate or others. In addition, the pro-and efficient expression oflacZin almost all cells.
portion of expressing cells varied among the three lines. Thus, line B38 showed-gal activity in a larger number of cells in each epithelium than lines B4 or B7 did.
MATERIALS AND METHODS
There were exceptions, such as the esophagus, where
Transgenic mice:TheK5Zconstruct was obtained by liga- the proportion of expressing cells was consistently tion of sequences from the bovine K5 locus (⫺5300 bp to higher in line B4 than in B38. Moreover,-gal activity ⫹140 bp) to alacZgene, as previously described (Ramı´rezet al.
per cell was also higher in line B38 than in the other
1994). To constructK5ZMARthe 5⬘A element of the chicken
two lines, as indicated by the intensity of X-gal staining of
lysozymegene in plasmid pB-1-X1 (Stiefet al.1989) was ligated
to the 5⬘end ofK5Z. TheK5Zloxconstruct was obtained by -gal-positive cells after simultaneous, timed incubation
insertion of an oligonucleotide containing a single copy of with the substrate. Figure 2B shows the results for palate the 34-bp lox site at the 3⬘end ofK5Z. TheK5ZHK10construct epithelium, but similar results were observed in the contains sequences from the bovineK5locus (⫺5300 bp to
tongue and tailskin of these lines and in other lines
⫹50 bp) fused to a genomic fragment spanning the entire
described below (not shown).
coding region of the humanK10(HK10) locus (Riegerand
Such a variegated expression ofK5Ztransgenes
con-Franke 1988). Transgenic mice were generated and
geno-typed as described (Ramı´rezet al.1994). For Cre recombina- trasts with the usually nonmosaic expression from
simi-tion, circular Cre-expression vector pCAGGS-Cre (Arakiet al. lar constructs in which a number of cDNAs are driven 1995) was injected into mouse eggs at 5 ng/l.
by the sameK5regulatory sequences used in theK5Z
-Galactosidase activity and immunofluorescence:Tissues
construct. Figure 3 shows examples of the expression
were dissected and frozen in OCT medium. Histochemistry
of both wild-type and truncated cDNAs encoding
and immunodetection were done on 7-m cryosections loaded
on poly-l-lysine-coated slides as described (Ramı´rez et al. HEGFR. In the three lines analyzed, the protein product
1994). The mouse K5 was detected using a rabbit antipeptide of the transgenes is detected in an even, nonmosaic antiserum (BAbCo), HK10 with monoclonal antibody AE2
fashion in the basal layer of stratified epithelia. Similar
(ICN Biochemicals), and -gal with a rabbit antiserum
results have been obtained by other researchers who
(5Prime-3Prime, Boulder, CO). Human epidermal growth
fac-have used the K5regulatory sequences present in the
tor receptor (HEGFR) was detected with monoclonal antibody
EGFR528 (Santa Cruz Biotechnology). Secondary antibodies K5Zconstruct to drive expression of a variety of cDNAs.
were Texas-red-conjugated donkey IgG anti-rabbit IgG and We conclude that the variegated expression ofK5Zmice fluorescein-conjugated goat IgG anti-mouse IgG from Jackson is neither due to lack ofcis-acting sequences nor to a ImmunoResearch Labs (West Grove, PA).
consequence of a heterogeneity among epithelial cells.
Southern and Northern blot analyses: These were
per-Instead, it appears most likely that it is the consequence
formed as described (Casanova et al. 1995). To normalize
of a process in which transgene activity occurs only in
for loading differences in Southern blots, we used a Thy1
probe. Known amounts of plasmids were used to help deter- a subset of cells and in their descendants.
mine copy numbers of the various transgenes. HK10 and The reduction of transgene copies has no effect on mouseK5(mK5) mRNAs were detected in Northern blot using variegation of K5/lacZ constructs: To investigate the genomic fragments spanning the last exon of theHK10and
effect of copy number onK5Zvariegation we used the
of the mK5 genes, respectively. Hybridization signals were
lox/Cre system of specific recombination that allows
quantitated using a Phosphorimager and Imagequant software
Figure 1.—Structure of K5 constructs used to generate transgenic mice. KeratinK5denotes the 5⬘flanking and pro-moter sequences of the bovineK5locus. The boxes labeled lacZrepresent the protein coding region of thetrpS-lacZgene and pA, a SV40 polyadenylation signal. White boxes indicate exon 1 sequences of theK5gene, whereas black boxes denote the exons of theHK10 gene. The box labeled MAR corre-sponds to the 5⬘A element from the chickenlysozymelocus, and the box labeled loxP represents the site recognized by the Cre recombinase of bacteriophage P1. Arrows denote the transcription initiation site and the direction of transcription.
of the construct at the same chromosomal location (Arakiet al.1995). We usedK5Zlox, a construct identical toK5Zexcept for the addition of a single copy of the 34-bp recognition site (loxP) of the Cre recombinase of bacteriophage P1 (Figure 1). Since transgenes usually
integrate in multimeric arrays, whenever a head-to-tail Figure2.—Expression properties of K5Z mice. (A) Com-arrangement occurs Cre-mediated recombination be- parison of lacZ expression and endogenous K5 localization in adult tissues of K5Z mice. Tissue sections of transgenic lines
tween loxP sites in individual K5Zlox transgenes will
B4 and B38 were incubated overnight at 37⬚ with X-gal to
reduce copies in the array. We generated six stable
detect the-gal activity and counterstained briefly with eosin.
K5Zloxtransgenic lines. Two of them showed no-gal The mouse K5 was detected by indirect immunofluorescence activity (or only in a few cells) whereas the other four with an anti-K5 serum. (B) Comparison of-gal activity per showed variegated expression similar to that of K5Z cell. Palate tissue sections of lines B4 and B38 stained at 37⬚
with X-gal for the indicated periods of time.
constructs (Table 1). Cre-mediated recombination was carried out by microinjection of a Cre expression vector
in zygotes obtained from mating wild-type female mice cells in double K5Cre/K5Zlox transgenic mice derived with hemizygous K5Zlox males. We used line 4734 (7 from lines 4715 (mouse 6603) and 4736 (mice 6483 and copies) with a reduced proportion of nonexpressing 6484), compared with the absence of recombination in cells and line 4712 (14 copies) with many nonexpressing single K5Zlox transgenic mice (mice 6607 and 5419), cells. The offspring from microinjected oocytes that are shown in Figure 4B.
terns contrasted with that of mice bearing theK5Z con-struct alone. Thus, the-gal expression pattern in six of the lines (3578a, 3578b, 3668, 4460, 4472, and 4483) was almost indistinguishable from that of the endoge-nousK5gene (Figure 6A), except for very small clusters of cells lacking-gal activity (see below). Of the other two lines, one (4290) expressed-gal in most epithelial cells, although not as extensively as in the precedent lines, whereas the other (4464) showed few -gal-express-ing cells. The expression ofHK10was almost identical to that oflacZ (not shown). Table 1 summarizes these data.We conclude that the presence of sequences with LCR-type properties (K5HK10) enhances significantly the expression of variegating constructs (K5Z) in the same transgene array.
Heterogeneity of expression of individual copies within a transgene array:The detailed study of the few cases of mosaic expression in K5Z ⫹ K5ZHK10 lines,
Figure3.—Expression of K5 HEGFR transgenes. Immuno- which was done by double immunofluorescence with fluorescence analysis of sagital sections of tails from mouse
rabbit anti--gal antibodies and with a mouse anti-HK10
lines bearing wild-type (A) and truncated (B) cDNAs for the
monoclonal antibody, led to some unexpected
observa-HEGFR. The protein is detected continuously throughout the
basal layer of the epidermis and in the hair follicles that appear tions. Thus, although line 3668 expressed in a
nonvari-as circular structures due to the section angle. gated fashion in most epithelia, patches of cells in the
epithelium of the ventral tongue did not express lacZ
but expressedHK10(Figure 6B, III and IV). Conversely, Taken together, these results indicate that copy-number in areas of the tongue and tailskin epithelia of line reduction has little effect on the variegation of K5Z 3578b some cells expressed lacZ but not HK10 (not
transgenes. shown). The most perplexing pattern was seen in small
The presence of an LCR-type sequence inK5Ztrans- regions of various epithelia of line 4290. Here, some genic arrays overcomes variegation: The tendency of cells expressed -gal only, while other cells expressed
K5Z transgenes to express in a variegated fashion did HK10only and yet other cells expressed both or neither not change when the 5⬘A element of the chickenlysozyme of the two genes (Figure 6B, V and VI). In contrast, a
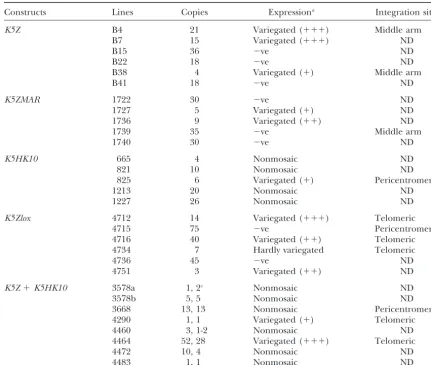
pat-TABLE 1
Characteristics of K5 transgenic lines
Constructs Lines Copies Expressiona Integration siteb
K5Z B4 21 Variegated (⫹⫹⫹) Middle arm
B7 15 Variegated (⫹⫹⫹) ND
B15 36 ⫺ve ND
B22 18 ⫺ve ND
B38 4 Variegated (⫹) Middle arm
B41 18 ⫺ve ND
K5ZMAR 1722 30 ⫺ve ND
1727 5 Variegated (⫹) ND
1736 9 Variegated (⫹⫹) ND
1739 35 ⫺ve Middle arm
1740 30 ⫺ve ND
K5HK10 665 4 Nonmosaic ND
821 10 Nonmosaic ND
825 6 Variegated (⫹) Pericentromeric
1213 20 Nonmosaic ND
1227 26 Nonmosaic ND
K5Zlox 4712 14 Variegated (⫹⫹⫹) Telomeric
4715 75 ⫺ve Pericentromeric
4716 40 Variegated (⫹⫹) Telomeric
4734 7 Hardly variegated Telomeric
4736 45 ⫺ve ND
4751 3 Variegated (⫹⫹) ND
K5Z⫹K5HK10 3578a 1, 2c Nonmosaic ND
3578b 5, 5 Nonmosaic ND
3668 13, 13 Nonmosaic Pericentromeric
4290 1, 1 Variegated (⫹) Telomeric
4460 3, 1-2 Nonmosaic ND
4464 52, 28 Variegated (⫹⫹⫹) Telomeric
4472 10, 4 Nonmosaic ND
4483 1, 1 Nonmosaic ND
aExpression in adult tissues as detected by X-gal staining. Lack of expression, or expression in very few cells,
is indicated as⫺ve. The degree of variegation is indicated from few nonexpressing cells (⫹) to a high proportion of nonexpressing cells (⫹⫹⫹).
bMiddle arm indicates lack of co-localization of transgene and centromeric or telomeric signals; telomeric
indicates co-localization of transgene and telomeric signals on the long chromosomal arm; pericentromeric indicates that the signal of the transgene is on the centromere or very close to the telomere of the short arm. ND, not done.
cCopies ofK5ZandK5HK10, respectively.
ingK5ZMAR line 1739 and for the nonmosaicK5Z⫹ are the two variegatingK5Z⫹K5ZHK10lines, suggesting that lack of expression in them could be related, at
K5HK10line 3668. The only variegatingK5HK10line
(825) and the nonexpressing K5Zlox line 4715 have least in part, to position effects imposed by telomere proximity.
their transgenes inserted at pericentromeric positions. Because of the acrocentric nature of mouse chromo-somes, we did not attempt to distinguish whether the
DISCUSSION
integration site is centromeric or telomeric on the short
arm in these two lines, because in our hands both signals The wide occurrence of variegated expression among transgenic lines has been noted recently. The mecha-do overlap (not shown). Among the lines with
peritel-omeric insertions were K5Z ⫹ K5HK10 with a high nisms of appearance of variegation are likely multiple. Some insight into these mechanisms has been obtained (4290) and low (4464) proportion of expressing cells.
Similarly,K5Zloxlines with a reduced (4734) or exten- from analysis of transgene expression in erythroid and lymphoid tissues. In this report we study transgene varie-sive (4712 and 4716) variegation showed insertions at
Figure4.—Effect of copy number onK5Zexpression. (A) Southern blot analysis ofK5Zloxmice with reduced copy number obtained by injecting a Cre-expressing plasmid in K5Zlox zygotes. Tail DNA was hybridized to alacZprobe and to aThy-1 probe to normalize for load-ing differences. Parental K5Zloxlines are shown above the lanes. A copy-number reduction, or the lack of it, is indicated by ⫹ and ⫺, respectively. The asterisks show truncated lacZgenes. (B) Southern blot analysis of genomic DNA isolated from liver (L), epidermis (E), and dermis (D) from offspring of crosses between K5Zlox mice and a mouse expressing Cre in the basal layer of stratified epithelia (K5Cre). Among the off-spring of line 4715 that carry K5Zlox, mouse 6603, but not 6607, is hemizygous for K5Cre.Similarly, among the K5Zloxoffspring of line 4736, mouse 6483 and mouse 6484, but not mouse 5419, are hemizygous forK5Cre. Blots were hybridized to a lacZ probe and to aThy-1probe.Specific copy-number reduction occurred in the epidermis of double transgenic mice. (C)-Gal activity of tissue sections from parental and reduced-copy-number mice from lines 4712 and 4712rec.c (I and II, respectively) and from lines 4734 (III, V, and VII) and 4734rec.a (IV, VI, and VIII).
DNA sequences and silencing: Some sequences can thelacZgene, because it could be argued that it is due, instead, to the LCR-like properties of HK10 alone or act as foci for gene silencing, as has been proposed to
explain the poor expression of ovine -lactoglobulin with K5 sequences. In any case, when the transgene arrays consisted of a mixture of K5Zand K5HK10 se-genes in transgenic mice (Clarket al.1997). Here, we
describe 17 K5Z mouse lines that either express in a quences most of the lines showed a nonmosaic expres-sion pattern. This is reminiscent of the lack of variega-variegated fashion or do not express at all. This adds
to the cumulative evidence of unreliable expression of tion often associated with transgenes harboring LCRs (KioussisandFestenstein 1997).
many lacZ-containing constructs, particularly in adult
tissues (Paldiet al. 1993;Thoreyet al.1993;Cuiet al. Using model (simplified) constructs, it has been shown that variegation varies depending on whether an 1994;Guyet al.1996). Interestingly, we show that when
cDNA fragments are exchanged by the lacZ gene, in enhancer is present or not. Thus, it has been proposed that enhancers increase the probability of expression equivalentK5constructs, they express more efficiently,
with little or no variegation. Other researchers have by preventing variegation alone, without affecting the transcription rate of the promoter (Walterset al.1996; used ourK5promoter sequences to drive expression of
a number of cDNAs, and in most cases they report Sutherlandet al.1997) or by affecting both ( Bouhas-sira et al.1997). Our data onK5Z, a construct with a nonmosaic expression (Robleset al.1996;Pierceet al.
1998;Wanget al.2000). This suggests that variegation complex set of regulatory sequences, support the latter model because the differences in-gal activity per cell (or even lack of expression) ofK5Zconstructs in mice
can be due, at least in part, tolacZ sequences, rather among ourK5Zlines, which are higher in the lines with a higher proportion of expressing cells, could indicate than to the absence ofcis-acting sequences required for
appropriate expression in the basal layer of stratified a variable transcription rate. An alternative possibility, however, takes into account possible differences in ac-epithelia. Nevertheless, the efficient, nonmosaic
Figure6.—Expression ofK5Z⫹K5HK10transgenes. (A) NonvariegatedlacZexpression in sections of tongue (I), tail-Figure5.—Expression ofK5HK10transgenes. (A)
Immu-skin (II), esophagus (III), palate (IV), and footsole (V) of a nofluorescence analysis of HK10 in tissue sections of a
repre-representative transgenic line. (B) Double immunofluores-sentativeK5HK10line. (B) Copy-number-dependent
expres-cence of HK10 (green) and-gal (red) in sections of dorsal sion ofK5HK10in transgenic mice. Ten micrograms of total
(B, I and II and V–VIII) and ventral tongue (B, III and IV) tailskin RNA isolated from a nontransgenic mouse (lane 1)
from the indicated lines. and fromK5HK10lines (lanes 2–6) was analyzed by Northern
blot using aHK10probe. To correct for loading differences, aK5probe was used. The normalized values ofK10mRNA are plotted against gene copy number.
gating lines are located at peritelomeric sites. While mosaic silencing of genes inserted at telomeric positions has been observed for both yeast (Gottschling et al.
not all copies in the transgene array are active within 1990) and Drosophila (Crydermanet al. 1999), well-the same cell. This is consistent with previous evidence documented evidence of such silencing was lacking in about chromatin heterogeneity within multicopy trans- mammals. Because of the potential silencing effects of gene arrays in mouse lines that showed either varie- lacZ sequences it could be argued that variegation of gated/position-dependent or efficient expression (Huber transgenes inserted into peritelomeric sites is not
neces-et al.1996;WhitelawandWebster1998). sarily only a reflection of a telomeric position effect.
Variegation and chromosomal position: The influ- However, the observation that, among the lines in which ence of the integration site for variegated expression of the effects due tolacZsequences can be overcome (by transgenes is well established for constitutively hetero- K5HK10sequences), the two variegating ones, but not chromatic regions at or near centromeres (Dobieet al. the one that expresses in a nonmosaic fashion, bear 1996;Festenstein et al.1996; Milot et al.1996). In- peritelomeric transgenes supports telomeric position deed, the two lines we find having pericentromeric in- effects in transgenic mice. This is consistent with recent sertions correspond to variegated (K5HK10 825 line) observations of telomeric position effects on replication or nonexpressing (K5Zlox4715 line) conditions. How- timing of human loci (Ofiret al. 1999) and indicates ever, in this study we provide evidence that transgene that both gene expression and DNA replication can be variegation does not require that the transgenes are affected by telomeric sequences in mammals.
varie-Figure7.—FISH of transgenic lines. (A–N) Bone marrow metaphase chromosomes were hybridized to a mixture of K5Z (pink) and mouse ␥-satellite probes (green) or to K5Z DNA (yellow) and a probe specific for mouse telomeric repeats (pink;Hanishet al.1994). Chromosomes were also stained with 4⬘ 6-dia-midino-2-phenylindole (blue). (O–R) Bone marrow metaphase chromosomes hybridized to theK5Zconstruct alone (pale green). For theK5ZMAR 1739 line, the transgene signal is yellow and chromosomes were stained with propidium iodide. Transgenic lines are indi-cated.
not at all. However, our analysis of the effect of gene tween variegation and transgene localization to hetero-chromatic regions of tissue culture cells (Francastel
repetition on variegation shows that a reduction in copy
number ofK5Ztransgenes does not increase the propor- et al. 1999). In transgenic mice, it has been reported that a silent transgene integrated into a peritelomeric tion of expressing cells, which suggests that repeated
induced gene silencing is unnecessary for transgene region co-localizes with centromeric satellite DNA on the nucleolus (Manuelidis 1991). It seems, however, repression. This is in contrast to results byGarrick et
al.(1998) who studied the expression properties of an that the size of the transgenic insert is critical for such a nuclear relocalization (Manuelidis1991). If this were
␣-globin/lacZ construct. The discrepancy could be
ex-plained by several differences between the two trans- the case, the␣-globin/lacZtransgene arrays, which seem larger than theK5Zloxtransgenes we have studied, may genic systems, such as the distinct cell types involved or
the possibility that subsets of chromosomal sites are find themselves in singular heterochromatic compart-ments within the interphasic nuclei from which they more favorable to repression than others. Thus, it is
conceivable that in three of the K5Zlox lines we used can be mobilized upon copy-number reduction. Any-how, our conclusion thatK5Zvariegation does not de-for copy-number reduction, the peritelomeric (two) or
pericentromeric (one) position of their transgenes may pend on the copies of the transgene is consistent with other reports of no apparent correlation between trans-impose a silencing effect regardless of copy number. An
alternative explanation lies in the compartmentalization gene copy number and degree of variegation (Zhuma
et al.1999). model of gene silencing. This model, put forward to
explain the silencing of transgenes located far away from In summary, we have shown that variegation ofK5Z
transgenes in stratified epithelia of transgenic mice is constitutive heterochromatin, proposes that the
varie-gating locus, after local heterochromatinization, is mis- not due to the lack of regulatory sequences. Instead, our data suggest that variegation can be related to pro-localized to a constitutive heterochromatic
be-Garrick, D., H. Sutherland, G. Robertsonand E. Whitelaw,
fects. This silencing effect, however, can be efficiently
1996 Variegated expression of a globin transgene correlates
overcome by strong activating sequences. Also, we have with chromatin accessibility but not methylation status. Nucleic
Acids Res.24:4902–4909.
shown that insertion of transgenes in pericentromeric
Garrick, D., S. Fiering, D. I. MartinandE. Whitelaw, 1998
Re-regions is not required for variegation and have
pro-peat-induced gene silencing in mammals. Nat. Genet.18:56–59.
vided evidence for telomeric position effects in trans- Gottschling, D. E., O. M. Aparicio, B. L. BillingtonandV. A.
Zakian, 1990 Position effect atS. cerevisiaetelomeres: reversible
genic mice. Finally, although we cannot rule out entirely
repression of Pol II transcription. Cell63:751–762.
the effect on variegation of the number of copies in a
Graubert, T. A., B. A. Hug, R. Wesselschmidt, C. L. Hsieh, T. M.
transgenic array, we have shown that, at least in some Ryanet al., 1998 Stochastic, stage-specific mechanisms account
for the variegation of a human globin transgene. Nucleic Acids
cases, both variegation and lack of expression are
inde-Res.26:2849–2858.
pendent of copy number.
Grosveld, F., andG. Kollias(Editors), 1992 Transgenic Animals, pp. 79–98. Academic Press, London.
We thank P. Vassalli for pCAGGS-Cre plasmid, A. Sippel for
pB-1-Guy, L. G., R. Kothary, Y. Derepentigny, N. Delvoye, J. Elliset
X1 plasmid, T. de Lange for pSXneo-1.6-T2AG3 plasmid, A. Langeveld
al., 1996 The beta-globin locus control region enhances tran-for technical assistance, J. Martı´nez, R. La´zaro, and E. Sole tran-for animal
scription of but does not confer position-independent expression care, and C. Cale´s for critical reading of the manuscript. This work
onto the lacZ gene in transgenic mice. EMBO J.15:3713–3721. was supported by Comision Interministerial de Cienca y Tecnologia
Hanish, J. P., J. L. Yanowitzand T. De Lange, 1994 Stringent (CICYT) grants PB94-1230 ( J.J.), PB94-0089, and PB97-1238 (M.V.). sequence requirements for the formation of human telomeres. I.P. was supported by a fellowship from the Universitat Auto`noma de Proc. Natl. Acad. Sci. USA91:8861–8865.
Barcelona. Henikoff, S., 1990 Position-effect variegation after 60 years. Trends
Genet.6:422–426.
Huber, M. C., G. KrugerandC. Bonifer, 1996 Genomic position effects lead to an inefficient reorganization of nucleosomes in the 5⬘-regulatory region of the chicken lysozyme locus in
trans-LITERATURE CITED
genic mice. Nucleic Acids Res.24:1443–1452.
Allshire, R. C., J. P. Javerzat, N. J. RedheadandG. Cranston, Kioussis, D., andR. Festenstein, 1997 Locus control regions: over-1994 Position effect variegation at fission yeast centromeres. coming heterochromatin-induced gene inactivation in mammals.
Cell76:157–169. Curr. Opin. Genet. Dev.7:614–619.
Araki, K., M. Araki, J. MiyazakiandP. Vassalli, 1995 Site-specific Manuelidis, L., 1991 Heterochromatic features of an 11-megabase recombination of a transgene in fertilized eggs by transient ex- transgene in brain cells. Proc. Natl. Acad. Sci. USA88:1049–1053. pression of Cre recombinase. Proc. Natl. Acad. Sci. USA 92: McKnight, R. A., A. Shamay, L. Sankaran, R. J. WallandL.
Hen-160–164. nighausen, 1992 Matrix-attachment regions can impart
posi-Assaad, F. F., K. L. Tucker andE. R. Signer, 1993 Epigenetic tion-independent regulation of a tissue-specific gene in trans-repeat-induced gene silencing (RIGS) inArabidopsis. Plant Mol. genic mice. Proc. Natl. Acad. Sci. USA89:6943–6947.
Biol.22:1067–1085. Meyer, P., I. HeidmannandI. Niedenhof, 1993 Differences in
Bouhassira, E. E., K. WestermanandP. Leboulch, 1997 Tran- DNA-methylation are associated with a paramutation phenome-scriptional behavior of LCR enhancer elements integrated at non in transgenic petunia. Plant J.4:89–100.
the same chromosomal locus by recombinase-mediated cassette Milot, E., J. Strouboulis, T. Trimborn, M. Wijgerde, E. De Boer
exchange. Blood90:3332–3344. et al., 1996 Heterochromatin effects on the frequency and
dura-Casanova, L., A. Bravo, F. Were, A. Ramirez, J. J. Jorcanoet al., tion of LCR-mediated gene transcription. Cell87:105–114. 1995 Tissue-specific and efficient expression of the human sim- Morley, S. D., I. Viard, B. C. Chung, Y. Ikeda, K. L. Parkeret al., ple epithelial keratin 8 gene in transgenic mice. J. Cell Sci.108: 1996 Variegated expression of a mouse steroid 21-hydroxylase/
811–820. beta-galactosidase transgene suggests centripetal migration of
Clark, A. J., G. HaroldandF. E. Yull, 1997 Mammalian cDNA adrenocortical cells. Mol. Endocrinol.10:585–598.
and prokaryotic reporter sequences silence adjacent transgenes Ofir, R., A. C. Wong, H. E. Mcdermid, K. L. SkoreckiandS. Selig, in transgenic mice. Nucleic Acids Res.25:1009–1014. 1999 Position effect of human telomeric repeats on replication Cryderman, D. E., E. J. Morris, H. Biessmann, S. C. Elginand timing. Proc. Natl. Acad. Sci. USA96:11434–11439.
L. L. Wallrath, 1999 Silencing atDrosophilatelomeres: nuclear Paldi, A., L. DeltourandJ. Jami, 1993 Cis effect of lacZ sequences organization and chromatin structure play critical roles. EMBO in transgenic mice. Transgenic Res.2:325–329.
J.18:3724–3735. Pierce, A. M., S. M. Fisher, C. J. ContiandD. G. Johnson, 1998
Cui, C., M. A. Wani, D. Wight, J. KopchickandP. J. Stambrook, 1994 Deregulated expression of E2F1 induces hyperplasia and cooper-Reporter genes in transgenic mice. Trends Genet.3:182–194. ates with ras in skin tumor development. Oncogene16:1267– Dobie, K. W., M. Lee, J. A. Fantes, E. Graham, A. J. Clarket al., 1276.
1996 Variegated transgene expression in mouse mammary Porter, S. D., andC. J. Meyer, 1994 A distal tyrosinase upstream gland is determined by the transgene integration locus. Proc. element stimulates gene expression in neural-crest-derived
mela-Natl. Acad. Sci. USA93:6659–6664. nocytes of transgenic mice: position-independent and mosaic
Dorer, D. R., andS. Henikoff, 1994 Expansions of transgene re- expression. Development120:2103–2111.
peats cause heterochromatin formation and gene silencing in Ramı´rez, A., A. Bravo, J. L. JorcanoandM. Vidal, 1994 Sequences
Drosophila. Cell77:993–1002. 5⬘of the bovine keratin 5 gene direct tissue- and cell-type-specific Elliott, J. I., R. Festenstein, M. TolainiandD. Kioussis, 1995 expression of a lacZ gene in the adult and during development.
Random activation of a transgene under the control of a hybrid Differentiation58:53–64.
hCD2 locus control region/Ig enhancer regulatory element. Rieger, M., andW. W. Franke, 1988 Identification of an
ortholo-EMBO J.14:575–584. gous mammalian cytokeratin gene. J. Mol. Biol.204:841–856.
Festenstein, R., M. Tolaini, P. Corbella, C. Mamalaki, J. Parring- Robertson, G., D. Garrick, W. Wu, M. Kearns, D. Martinet al., tonet al., 1996 Locus control region function and heterochro- 1995 Position-dependent variegation of globin transgene ex-matin-induced position effect variegation. Science 271: 1123– pression in mice. Proc. Natl. Acad. Sci. USA92:5371–5375.
1125. Robles, A. I., F. Larcher, R. B. Whalin, R. Murillas, E. Richieet
Francastel, C., M. C. Walters, M. GroudineandD. I. Martin, al., 1996 Expression of cyclin D1 in epithelial tissues of trans-1999 A functional enhancer suppresses silencing of a transgene genic mice results in epidermal hyperproliferation and severe and prevents its localization close to centrometric heterochroma- thymic hyperplasia. Proc. Natl. Acad. Sci. USA93:7634–7638.
tin. Cell99:259–269. Sabl, J. F., andS. Henikoff, 1996 Copy number and orientation
Stief, A., D. M. Winter, W. H. StratlingandA. E. Sippel, 1989 1996 Transcriptional enhancers act in cis to suppress position-effect variegation. Genes Dev.10:185–195.
A nuclear DNA attachment element mediates elevated and
posi-Wang, D., J. L. RussellandD. G. Johnson, 2000 E2F4 and E2F1 tion-independent gene activity. Nature341:343–345.
have similar proliferative properties but different apoptotic and Sutherland, H. G., D. I. MartinandE. Whitelaw, 1997 A globin
oncogenic properties in vivo. Mol. Cell. Biol.20:3417–3424. enhancer acts by increasing the proportion of erythrocytes
ex-Whitelaw, B., and J. Webster, 1998 Chromatin heterogeneity pressing a linked transgene. Mol. Cell. Biol.17:1607–1614.
within multicopy transgene arrays. Transgenic Res.7:401–402. Thorey, I. S., J. J. Meneses, N. Neznanov, D. A.Kulesh, R. A.
Zhuma, T., R.Tyrrell, B.Sekkali, G.Skavdis, A.Savelievet al., Pedersenet al., 1993 Embryonic expression of human keratin
1999 Human HMG box transcription factor HBP1: a role in 18 and K18--galactosidase fusion genes in transgenic mice. Dev.
hCD2 LCR function. EMBO J.18:6396–6406. Biol.160:519–534.