Copyright 0 1997 hy the Genetics Society of America
Conditions for Protected Inversion Polymorphism Under Supergene Selection
Gonzalo
Alvarezand Carlos Zapata
Departamento de Biologia Fundamental, Facultad de Biologia, Uniuersidad de Santiago de Compostela, Santiago de Compostela, Spain Manuscript received September 20, 1996
Accepted for publication February 27, 1997
ABSTRACT
Conditions for protected inversion polymorphism under the operation of both karyotype and super- gene selection in a viability model have been analytically determined. When supergene selection (the effect of recombination in homokaryotypes lowering the mean fitness of their offspring) is acting on gene arrangements and there is no karyotype selection, it is demonstrated that a polymorphic stable equilibrium is reached by the population, which is a function of only the recombination effects in
homokaryotypes. Under both supergene and karyotype selection the degree of dominance ( h ) of karyo-
type selection is critical to produce a protected inversion polymorphism. In general, the opportunity for protected polymorphism increases as the degree of dominance decreases. For small s values, the conditions for protected polymorphism are r > 2sh and c > 2s( h - 1 ) , where r and c are the average loss of viability for offspring of ST/ ST and I N / I N homokaryotypes, respectively. These findings suggest
that supergene selection may be an important balancing mechanism contributing to the maintenance of inversion polymorphism.
”
S
EVERAL types of balancing selection have been pro- posed to explain the maintenance of inversion polymorphism in natural populations of Drosophila. The superior fitness of heterokaryotypes or heterosis is the mechanism advanced by DOBZHANSKY in his classical investigations on the third chromosome inversion poly- morphism of D. pseudoobscuru (WRIGHT and DOBZHAN-SKY 1946; DOBZHANSKY 1947, 1970). According to DOBZHANSKY chromosomes carry supergenes, that is, complexes of linked genes that are favorable in some particular combinations but not in others, and the sup- pression of recombination in heterokaryotypes binds together these coadapted gene complexes. In a given population, the chromosomes with different gene ar- rangements are coadapted to yield highly fit heterokary- otypes ( DOBZHANSKY 1948,1970; DOBzHANsKYand PAv- LOVSKY 1953, 1958). Frequency-dependent selection is another type of balancing selection usually suggested to play an important role in the maintenance of inversion polymorphism in natural populations. In last decades, experimental evidence of balancing frequency-depen- dent selection, that is, selection favoring rare karyo- types, has been reported for fitness traits such as larval viability, sexual selection and fertility in Drosophila spe- cies ( KOJIMA and TOBARI 1969; NASSAR et al. 1973; AN- DERSON and WATANABE 1974; GROMKO and RICHMOND 1978; ANDERSON and BROWN 1984; ANDERSON et al.
1986). However, at present, it is not completely clear which could be the major balancing selection mecha-
Corresponding author: Gonzalo Alvarez, Departamento de Biologia Fundamental, Facultad de Biologia, Universidad de Santiago, Santi- ago de Compostela, Spain. E-mail: [email protected]
Genetics 146 717-722 (June, 1997)
nism responsible for the maintenance of inversion poly- morphism in Drosophila. In fact, several authors have suggested that a joint effect of different modes of selec- tion such as heterokaryotype advantage and frequency- dependent selection could be operating on the chro- mosomal polymorphism (KOJIMA and TOBARI 1969; GROMKO and RICHMOND 1978; ANDERSON et ul. 1986). WASSERMAN (1968, 1973, 1975) proposed another balancing selection mechanism for the inversion poly- morphism that is based on the effect of the recombina- tion between different supergenes. According to WAS
SEW (1968), several different supergenes could exist within one arrangement in a population. Recombi- nation between different supergenes in the homokaryo- types would destroy the coadapted combinations of genes and would thereby produce a frequency-depen- dent selection against the more common homokaryo- type. This supergene selection differs from DOBZHAN-
SKY’S viewpoint in that is based on the recombination
between different supergenes existing in each gene ar- rangement. Therefore, the mechanisms proposed by DOBZHANSKY and WASSERMAN, both based on the co- adapted supergene hypothesis, are not two mutually exclusive mechanisms, and they may be considered as cooperative balancing selection mechanisms. Recur- rence equations for karyotypic frequencies in a viability model with both supergene (recombination effect)
1968). In many instances, polymorphic stable equilibria were attained by the population and, in all these cases, heterokaryotypes were superior in total fitness at the equilibrium (denoted as “overall heterosis” by WASSER-
MAN 1968), However, the conditions with respect to
recombination effects and selection coefficients de- termining these equilibria could not be established.
In the present article, the conditions that determine what PROUT (1968) has called a “protected polymor- phism,’’ that is, conditions that bring about the increase of either allele when its frequency is close to 0, are analytically determined for an inversion polymorphism under the WASSEFWAN’S model ( WM ) with both super- gene and karyotype selection. On this basis, we deter- mine the regions of the parameter space of recombina- tion effects and selective values, where polymorphic stable equilibria occur. The analysis presented here shows that supergene selection may be a powerful bal- ancing selection mechanism and therefore could play an important role in the maintenance of inversion poly- morphism.
THE MODEL
A population with two gene arrangements, ST and IN, is considered. Selection is assumed to occur through viability differences. Karyotypic viability values are de- signed VI, V, and V3 corresponding to karyotypes ST/ ST, ST
/
IN and IN/
IN, respectively. Crossing over be- tween different epistatically balanced supergenes in ho- mokaryotypic females is assumed to produce unbal- anced recombinant chromosomes and therefore less fitoffspring. We denote the average loss of viability fitness for offspring of ST
/
ST and IN/
IN mothers as r and c , respectively. Discrete generations, sex-independent selection and random mating among selected adults are also assumed.Let us define the karyotypic frequencies of the repro- ducing adults after selection for ST/ ST, ST/ IN and IN/IN at generation t, X , Y and Z , respectively. The karyotypic frequencies after selection at t
+
1 genera- tion are as follows:X’ =
v,(p2
- r p X ) /Tq
Z ’ = &( 4 2 - c q Z ) /
R
p
=x
+
l/,Y,q
= Z + ’ / , Y ,Y’ = V2(2pq -
rqx
- c p Z ) /W,
( 1 )where
IT=
V,(P‘
-?X)
+
v,
x
( 2 p q -rqx
- c p 2 )+
v , ( 42 - c q Z ) .These expressions were obtained by WASSERMAN
( 1968) with the only difference being that in our case
a different recombination effect for each homokaryo- type is considered.
As it has been pointed out by WASSERMAN (1968), this equation system is very difficult to analyze directly to solve for equilibrium points. Therefore, we have in- vestigated the conditions that guarantee that both gene arrangements remain in the population, although the position of any interior equilibrium is not specified. The instability of the fixation points X = 1, Z = 0 and X = 0 and Z = 1 is investigated by means of the Jacobian matrix at ( X , Z )
.
This Jacobian matrix evaluated at the fixation point ( 1, 0 ) isVl( 1 - I/,r) -V,( 1 - c - l / , r )
1
vd1; r ) K ( 1 - r ),
0whose dominant eigenvalue is
A = V2(1 - ’ / z ~ ) / K ( l - r )
>
1,and the Jacobian matrix at ( 0 , 1 ) is
0 0
- v , ( 1 - r - l / , C ) v,(1 - ‘ / , c )
G ( 1
- c ) V3(1-
c)whose dominant eigenvalue is
A = V2( 1 - C)
/v3( 1
- C)>
1.Therefore, the necessary and sufficient conditions for instability of the trivial equilibria are as follows:
v,
( 1 - I/2r)>
VI ( 1 - T )V2(1 - l / , C )
>
G ( 1
- c ) . ( 2 )The biological meaning of these expressions can eas- ily be understood considering the total fitness values under the
WM.
Total fitnesses, defined as the ratios of genotypic frequencies after selection to genotypic frequencies before selection ( PROUT 1969; A L V ~ Z etal. 1984), are readily obtained from (1) as
w,
=K ( l
- r x / p ) ,w,
=v,
(1 - I / , r x / p - l/:!cz/ q ) ,w,
= V q ( 1 - c Z / q ) , ( 3 )where W,, W, and W3 are the total fitness values for ST
/
ST, ST/
IN and IN/
IN, respectively. Thus, under the WM total fitness splits into two independent compo- nents: karyotypic viability and supergene selection. This last component is responsible for the frequency-depen- dent fitnesses associated with the supergene selection. The ratios X / p and Z/ q , that is, the proportion ofgene arrangements in homozygous state, produce the frequency-dependent pattern.
Protected Inversion Polymorphism 719
limx+ Wl = V, ( 1
-
T )limx+l W, = V, ( 1 -1/2r)
lirnx+, W3 =
V,
for the fixation point ( 1, 0 ) , and as
limx+, Wl = V,
limx-" W , = V , ( 1 - 1/2 e )
limx+" W3 = r/l? ( 1 - c )
for the fixation point ( 0 , 1 )
.
Therefore, conditions for protected inversion polymorphism expressed by means of ( 2 ) are simply that the heterokaryotype total fitness must be higher than the fitness of the more frequent homokaryotype at each fixation point.Supergene selection with no karyotype selec- tion: When gene arrangements are under the opera- tion of supergene selection and there is no karyotype selection (V, = V , =
V,
= 1 ) , the necessary and suffi- cient conditions for protected inversion polymorphism from ( 2 ) are r > 0 and c>
0.In this case, the WM can be specified in terms of gene arrangement frequencies. Thus, the change in gene arrangement frequency from ( 1 ) becomes
aq=
[ q ( r X + CZ) - c Z ] / [ 2 ( 1 - r X - C Z ) ] .At equilibrium
Aq
= 0 , and thereforeq ( rX
+
CZ) -cz
= 0. ( 4 )Total fitnesses of karyotypes under supergene selec- tion, according to ( 3 )
,
are as follows:W, = 1 -
r x / p
W, = 1 - 1 / 2 r X / p -
'/,
c z /q
w,
= 1 - c z / q ( 5 )and so the selection coefficient of the heterokaryotype is the arithmetic mean of the selection coefficients of the two homokaryotypes. Given that at the equilibrium expression ( 4 ) must be satisfied, the total fitnesses at this point will be 6, =
z&
= zij3 = 1 - ( r X+
c Z ) , and therefore, in terms of relative fitness, 6, = = G3 =1. That is, fitness values converge to 1 at the equilibrium under supergene selection, and at this point there are no fitness differences among karyotypes. As a conse- quence of this fact, Hardy-Weinberg departures are not expected to occur at the equilibrium.
Consequently, under supergene selection it is possi- ble to assume that the karyotypes are in Hardy-Wein- berg proportions at the equilibrium, and then ( 4 ) re- duces to
q 2 ( r + e )
-
q ( 2 r + c)+
r = 0with a nontrivial equilibrium solution for
q =
r / ( r + c ) andp =
c / ( r + c ) . ( 6 )The formal similarity of expressions ( 6 ) with the equilibrium frequencies under heterozygote fitness ad- vantage does not mean that supergene selection and heterokaryotype advantage are similar selective mecha- nisms. Under supergene selection karyotype fitnesses are frequency-dependent according to
( 5 )
with the het- erokaryotype fitness always being intermediate between the two homokaryotypes. In addition, at the equilib- rium, supergene selection does not produce deviations from Hardy-Weinberg proportions ( F = 0 ) as shown above, while heterokaryotype advantage will produce a heterokaryotype excess, which can easily be demon- strated to be F = -st/ ( s+
t - s t ) , where s and t are the selection coefficients against the homokaryotypes (WORKMAN 1969).Supergene selection and different types of karyotype selection: To know the conditions for stable equilibria under the operation of both supergene selection and different types of karyotype selection, karyotypic viabili- ties are expressed in terms of selection coefficient (s)
and degree of dominance ( h )
,
so that V, = 1, V2 = 1- hs and
V,
= 1 - s. If it is done, ( 2 ) can be expressed as follows:r > s h ( 2 - r ) ,
c[l
+
s ( h - 2 ) ]>
2 s ( h - 1 ) .( 7 )
It immediately follows from these expressions that if s
is small enough,
r
>
2shand
c > 2 s ( h - 1 )
have to be satisfied to produce a polymorphic equilib- rium. This means that for given values of h and s, the minimum value of recombination effect required to produce a protected polymorphism will be higher in the favored homokaryotype than in the disfavored one. Only for large h do the two conditions of protected polymorphism tend to converge to the same value. In any case, if selection against the homokaryotypes due to recombination effect is stronger than twice selection against the heterokaryotype the occurrence of a pro- tected polymorphism is guaranteed.
Conditions for protected inversion polymorphism ac- cording to expressions (
7 )
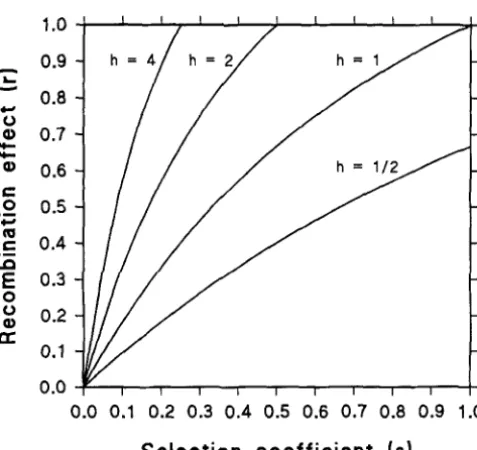
are given in Table 1 for particular cases of karyotype selection. It is observed that the degree of dominance of karyotype selection is critical to determine the conditions for protected polymorphism. Thus, the region of polymorphic equi- libria in the parameter space increases as the degree ofdominance decreases (see also Figure 1 )
.
720 C. Zapata
TABLE 1
Conditions for protected inversion polymorphism under supergene selection and different types of karyotype selection
Karyotype selection ( X , z) = ( 1 , 0) ( X , z) = (0, 1 )
General case r > sh(2 - r) c [ l
+
s ( h - 2 ) l > 2s(h - 1 )No karyotype selection (s = 0) r > O c > 0
Karyotype selection with h < 0 (superior heterokaryotype, h = - k ) r > - sk(2 - r) c > - s ( [ k ( 2 - c)
+
2(1 - c)] Karyotype selection with h zz 0 r > 2sh/(1+
sh) c [ l+ s ( h
- 211 > 2s(h - 1)Selection against recessive homokaryotype ( h = 0) r > 0 c > - 2s(l - c) Selection against semidominant homokaryotype ( h = 1/2) r > s / ( l
+
‘/ps) c > - s(1 - “/?C) Selection against dominant homokaryotype ( h = 1) r > 2s/(l+
s) c > 0Selection against heterokaryotype ( h > 1) r > 2sh/(1
+
sh) c [ l+
s(h - 2 ) ] > 2s(h - 1)Adaptive values corresponding to karyotype selection are expressed in terms of selection coefficient (s) and degree of domi-
nance ( h ) : & = 1, & = 1 - hs, V, = 1 - s.
tion effect ( T
>
0 ) in the favored homokaryotype guar-antees the maintenance of inversion polymorphism regardless of the magnitude of the selection coefficient. This means that although a strong viability selection is operating against a recessive homokaryotype, the exis- tence of a weak supergene selection will prevent the fixation of the favored arrangement and the mainte- nance of the inversion polymorphism will be assured. Under additive karyotype selection ( h = ‘ / 2 ) , the value of the recombination effect in the favored homokaryo- type ( T ) is, in comparison with the selection coefficient
( s ) , critical to determine a stable polymorphism. The equilibrium region for values of r and s is shown in Figure 1 as the area over the curve h = When the deleterious effect associated with karyotypic viability is completely dominant ( h = 1 ) , the occurrence of some recombination effect in both homokaryotypes is neces-
1
.o
0.9 0.8 0.7 0.6 0.5 0.4 0.3 0.2
0.1
0.0
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0
Selection coefficient
(SI
sary to assure the maintenance of polymorphism (see Figure 1 )
.
When selection is acting against the hetero- karyotype ( h>
1 ) , recombination effect in both homo- karyotypes must also be present to prevent the fixation of gene arrangements. In the particular case of h = 2 ,the conditions for polymorphism are r
>
4s/ ( 1+
2s)and c
>
2s. As can be seen in Figure 1, only a small proportion of the parameter space leads to stable equi- libria when the heterokaryotype is inferior in viability. To check the theoretical predictions of the WM here obtained, we performed several numerical examples by computer. From initial values of X , Yand Z , karyotypic frequencies were generated by means of ( 1 ) along suc- cesive generations until equilibrium points were reached. We use two started values identical to those used by WASSERMAN (1968) in his simulations: ( 1 )x,
= 0.999, y0 = 0.001, Z, = 0.000 and ( 2 ) = 0.000,
= 0.001, Z, = 0.999. Several combinations of T and
cvalues were considered for different hand s, including the case of no karyotype selection ( s = 0 ) . The numeri- cal computations performed confirmed all the theoreti- cal predictions and they also showed that the degree of dominance of karyotypic viabilities controls the fitness behavior at the equilibrium. Thus, a heterokaryotype advantage in total fitness is observed at the equilibrium when the degree of dominance is I ‘/2, but when the
degree of dominance exceeds stable equilibria with heterokaryotype inferiority in total fitness occur. Conse- quently, our analysis shows that heterosis is not a neces- sary prerequisite for the maintenance of inversion poly- morphism under supergene selection. In this sense, the term “overall heterosis,” used by WASSERMAN (1968)
to refer to the heterokaryotype advantage induced by recombination effect, is not appropriate to characterize the supergene selection mechanism since the hetero- karyotype advantage in total fitness at the equilibrium only occurs with some particular cases of karyotype se-
FIGURE 1.-Regions of the parameter space (above the lection (superior heterokaryotype, and additive and re-
curves) that yield polymorphic stable equilibria as a function cessive selection), which were precisely those studied
Protected Inversion Polymorphism 721
selection produces no differences in karyotypes at the equilibrium as shown
DISCUSSION
fitness among above.
The analyses presented here show that supergene selection, that is, the effect of recombination between different supergenes in homokaryotypes producing un- balanced recombinant chromosomes, can be a poten- tially important mechanism of balancing selection. When both supergene and karyotype selection are si- multaneously operating on two gene arrangements, the recombination effect will lead to a protected polymor- phism in many instances, even when karyotypic viability tends to produce the fixation of one of the two gene arrangements (Table 1 and Figure 1 )
.
The balancing effect of supergene selection overcomes in many cases the effect of directional karyotype selection, and the degree of dominance of karyotypic viabilities is critical to determine the occurrence of a stable inversion poly- morphism. In general, the oportunity for polymor- phism is increased as the degree of dominance de- creases. Even when selection operates against the heterokaryotype ( h>
1 ),
polymorphic stable equilibria can be attained by the population.Several lines of evidence support the occurrence of supergene selection in natural populations. First of all, it is commonly accepted that supergenes must be associ- ated with the existence of gametic disequilibrium be- tween alleles at these loci (see HEDRICK et ul. 1978). Although, disequilibrium studies between allozyme polymorphisms, carried out during the past two de- cades mainly with Drosophila species, seemed to indi- cate that nonrandom associations are rare in natural populations of outcrossing species, it has been recently shown that the statistical power of the standard tests used to detect associations were too low and that moder- ate gametic disequilibria are actually occurring in natu- ral populations of Drosophila ( ZAPATA and ALVAREZ
1992)
.
Second, theoretical studies on selection for newly arising inversions in multilocus systems at equilib- rium under recombination and selection in large popu- lations have shown that there will be appreciable selec- tion for the new inversion only if the population is at equilibrium with gametic disequilibrium (CHARLES-WORTH and CHARLESWORTH 1973). In addition, the ex- pected chance of survival of the new inversion depends on the loss of fitness due to recombination (recombina- tion genetic load) in the original population. Conse- quently, from a theoretical point of view, supergenes and recombination effects on fitness are both expected to occur associated with inversions. Finally, a direct ex- perimental evidence supporting supergene selection comes from the classical experiments of release of ge- netic variability through recombination in Drosophila. These experiments show that recombinant homozygous
chromosomes frequently present a consistent reduction in viability with respect to unrecombinant chromo- somes derived from natural populations (see SPIES 1989, pp. 548-551 )
.
However, this evidence refers to recombinant chromosomes in homozygous condition while the WM of supergene selection requires specifi- cally that the unbalanced chromosomes produced by the homokaryotype females have a dominant deleteri- ous effect on the nonrecombinant chromosomes yielded by the males. Obviously, this is due to the fact that the WM is primarily concerned with Drosophila and therefore crossing over limited to the female sex is assumed. Unfortunately, the experimental evidence on fitness effects of recombinant chromosomes in het- erozygous condition is rather scarce. WASSERMAN(1972) found a significant recombination effect in low- ering hatchability of eggs produced by homokaryotypic mothers by using chromosomal strains in D. subobscuru.
However, recombination had no detectable effect up- pon egg-to-adult viability of gene arrangements of D.
pseudoobscura, although a small number of indepen- dently derived chromosomes was used (WASSERMAN 1975). In
D.
mlunogaster, a large and significant effect of recombinant chromosomes on the female fecundity was detected by CHARLESWORTH and CHARLESWORTH( 1975), but only a small and nonsignificant effect of recombination on viability was observed. Therefore, the experimental evidence on fitness effects of recombi- nant chromosomes in heterozygous condition seems to be inconclusive and further experiments will be needed to evaluate the role of supergene selection in the main- tenance of the inversion polymorphism of Drosophila.
The authors are grateful to an anonymous reviewer who obtained the expressions for protected polymorphism for small s values.
LITERATURE CITED
~ V A R E Z , G., M. SANTOS and C. ZAPATA, 1984 Frequencydependent selection arisingfrorn inappropriate fitness estimation. Evolution
ANDERSON, W. W., and C. J. BROWN, 1984 A test for rare male mating advantage with Drosophila pseudoobscura karyotypes. Genetics 107:
ANDERSON, W. W., and T. K WATANABE, 1974 Selection by fertility in Drosophila pseudoobscura. Genetics 77: 559-564.
ANDERSON, W. W., J. ARNOLD, S. SAMMONS and D. G. YARDLEY, 1986
Frequencydependent viabilities ofDrosophilapsseudoobscura karyo- types. Heredity 56: 7-17.
CHARLESWORTH, B., and D. CHARLESWORTH, 1973 Selection of new inversions in multi-locus genetic systems. Genet. Res. Camb. 21:
CHARLESWORTH, B., and D. CHARLESWORTH, 1975 An experiment on recombination load in Drosophila melanagaster. Genet. Res. Camb. 25: 267-274.
DOBZHANSKY, T., 1947 Genetics of natural populations. XIV. A re- sponse of certain gene arrangements in the third chromosome of Drosvphilapseudoobscura to natural selection. Genetics 32: 142-
160.
DOBZHANSKY, T., 1948 Genetics of natural populations. XVIII. Ex- periments on chromosomes of Drosophila pseudoobscura from dif- ferent geographic regions. Genetics 33: 588-602.
DOBZHANSKY, T., 1970 Genetics of the Evolutionaly Process. Columbia University Press, New York.
38: 696-699.
577-589.
G. and C . Zapata
DOBZHANSKY, T., and 0. PAVLOVSKY, 1953 Indeterminate outcome
of certain experiments on Drosophila populations. Evolution 7:
DOBZHANSKY, T., and 0. PAVLOVSIW, 1958 Interracial hybridization and breakdown of coadapted gene complexes in Drosophila pau- listorum and Drosophila willistoni. Proc. Natl. Acad. Sci. USA 44:
GROMKO, M. H., and R. C. RICHMOND, 1978 Modes of selection maintaining an inversion polymorphism in Drosophila paulist- orum. Genetics 88: 357-366.
HEDRICK, P., S. JAIN and L. HOLDEN, 1978 Multilocus systems in evolution. Evol. Biol. 11: 101-184.
KOJIMA, K. I., and Y. N. TOBARI, 1969 Selective modes associated with karyotypes in Drosophila ananassae. 11. Heterosis and fre- quencydependent selection. Genetics 6 3 639-651.
NASSAR, R., H. J. MUHS and R. D. COOK, 1973 Frequencydependent selection at the Payne inversion in Drosophila melanagaster. Evolu- tion 27: 558-564.
PROur, T., 1968 Sufficient conditions for multiple niche polymor- phism. A m . Nat. 102: 493-496.
198-210.
622-629.
PROUT, T., 1969 The estimation of fitness from population data. Genetics 63: 949-957.
SPIESS, E. B., 1989 Genes in Populations. John Wiley & Sons, New York.
WASSERMAN, M., 1968 Recombination-induced chromosomal heter- osis. Genetics 58: 125-139.
WASSERMAN, M., 1972 Factors influencing fitness in chromosomal strains in Drosophila subobscura. Genetics 72: 691-708.
WASSEFNAN, M., and H. R. KOEPFER, 1975 Fitness of karyotypes in
Drosophila pseudoobscura. Genetics 7 9 113-126.
WORKMAN, P. L., 1969 The analysis of simple genetic polymorphism. Hum. Biol. 41: 97-114.
WRIGHT, S., and T. DOBZHANSW, 1946 Genetics of natural popula- tions. XII. Experimental reproduction of some of the changes caused by natural selection in certain populations of Drosophila pseudoobscura. Genetics 31: 125-156.
ZMATA, C., and G. ALVAREZ, 1992 The detection of gametic disequi- librium between allozyme loci in natural populations of Drosoph- ila. Evolution 46: 1900-1917.