ABSTRACT
CRAWFORD, BRIGITTE DIANE. Increasing Postharvest Quality and Propagation Success of Plant Cuttings. (Under the direction of John M. Dole.)
Unrooted cuttings are important to the floriculture industry, because many crop species are propagated primarily from stem and stem tip cuttings. The production of healthy rooted cuttings is affected by numerous factors, commencing with healthy stock plants and followed by proper harvest and postharvest handling, and optimal propagation conditions.
The effects of stock plant age and environment at time of cutting harvest and the application of rooting compound on Pelargonium x hortorum (geranium), Impatiens hawkeri [(New Guinea impatiens (NGI)], and Solenostemon scutellarioides (coleus) cuttings were investigated. Use of rooting compound increased root fresh and dry weights of geranium and root fresh weight of one NGI cultivar, but not of coleus. Cuttings harvested at the same time from stock plants of three distinct ages had similar fresh and dry weights, quality ratings, and number of abscised or yellow leaves. Coleus shoot and root fresh and dry weights and geranium shoot and root fresh weights were correlated with averaged day lengths and light intensities of the stock plant environment (averaged over up to 28 days prior to cutting harvest). When cuttings were propagated in a uniform growth chamber environment, the same correlations were lacking or inconsistent, suggesting that cutting propagation environment influenced performance more than stock plant environment or stock plant age.
cuttings, results varied by cultivar and storage conditions. Overall, callused cuttings rooted more consistently, and callusing greatly reduced the post-storage propagation time until roots formed, reducing the time spent in the propagation space.
The effects of propagation stage and anti-ethylene agent 1-MCP on sensitivity of geranium and NGI cuttings to ethylene and simulated shipment (0, 2 d storage at 21°C) were investigated. Stage 1 (freshly harvested) geranium ‘Tango Dark Red’ and impatiens ‘Super Sonic White’ and ‘Super Sonic Red’ cuttings placed in 2 d simulated shipment rooted at about the same time as cuttings placed directly into propagation. Since cuttings
apparently continued development during simulated shipment, short term storage at 21°C may be acceptable without increasing bench time required at the propagator. Ethylene resulted in reduced shoot fresh and dry weights of Stage 1 ‘Tango Dark Red’ cuttings. 1-MCP had negligible effects on cutting performance.
The effects of three types of modified atmosphere packaging (MAP) on quality of unrooted cuttings of eleven floriculture species and cultivars during shipment and four storage durations (0, 1, 2, 3 weeks) were investigated. All cultivars decreased in quality as storage duration increased, whether by increased Botrytis, abscised leaves, cutting
Increasing Postharvest Quality and Propagation Success of Plant Cuttings
by
Brigitte Diane Crawford
A thesis submitted to the Graduate Faculty of North Carolina State University
in partial fulfillment of the requirements for the degree of
Master of Science
Horticultural Science
Raleigh, North Carolina 2013
APPROVED BY:
_______________________________ Dr. John M. Dole
Committee Chair
ii DEDICATION
To Grandpa (Jack Walker) Crawford November 10th, 1919 – August 7th, 2012
While some people never even get to know their grandparents, I had the privilege of not only knowing Grandpa for 27 years, but growing up in the same town as all of my living grandparents. Grandpa’s service to his country, love and pride in his family, devotion to his 70 year marriage and passion for his hobbies will always be admired and remembered. After Grandpa passed, friends of his that I didn’t even know came up to me at his Celebration of Life, asking about my turkey hunting and giant pumpkin growing because Grandpa had told them about me. I know earning my Master of Science is something he would have been proud of and shared with them as well, so in the midst of mourning, months before my defense, I had to dig deep to refocus and find the strength and drive to keep going. Sad or happy, stressed or calm, “thesisizing” had to get done.
iii BIOGRAPHY
Brigitte Diane Crawford was born to Tim and Linda Crawford and older brother Nick on May 28th, 1985 in Chippewa Falls, Wisconsin. She grew up in the countryside of
iv ACKNOWLEDGMENTS
v TABLE OF CONTENTS
LIST OF TABLES ... xv
LIST OF FIGURES ... xxvii
CHAPTER 1. INTRODUCTION ... 1
Cutting production in the floriculture Industry ... 1
Stock plant age ... 3
Rooting compounds ... 4
Stage of propagation at shipment ... 4
Callus tissue ... 5
Storage temperature ... 5
Storage duration ... 6
Ethylene ... 7
Ethylene inhibitors ... 8
Negative effects of ethylene inhibitors ... 10
Oxygen and carbon dioxide ... 10
Modified atmosphere packaging ... 11
MAP application ... 12
Gas influence on MAP ... 13
Temperatures influence on MAP ... 14
vi
MAP selection ... 16
Literature Cited ... 17
CHAPTER 2. PROPAGATION SUCCESS OF UNROOTED PLANT CUTTINGS AS INFLUENCED BY STOCK PLANT AGE AND CROP SEASON ... 21
Abstract ... 22
Introduction ... 24
Stock plant age ... 24
Rooting compound ... 25
Materials and methods ... 26
Results ... 30
‘Stained Glass’ coleus ... 30
Effects of age and rooting compound in crop 1 ... 30
Effects of age and rooting compound in crop 2 ... 31
Effects of age and rooting compound in crop 3 ... 32
Age analysis of cuttings harvested on same dates ... 32
Harvest season analysis of cuttings of same age ... 32
Correlation with day length ... 33
Correlation with light intensity ... 34
‘Tango’ geranium ... 35
vii
Effects of age and rooting compound in crop 2 ... 36
Effects of age and rooting compound in crop 3 ... 36
Age analysis of cuttings harvested on same dates ... 37
Harvest season analysis of cuttings of same age ... 38
Correlation with day length ... 40
Correlation with light intensity ... 40
‘Kardino’ geranium ... 41
Effects of age and rooting compound in crop 1 ... 41
Effects of age and rooting compound in crop 2 ... 42
Effects of age and rooting compound in crop 3 ... 43
Age analysis of cuttings harvested on same dates ... 43
Harvest season analysis of cuttings of same age ... 43
Correlation with day length ... 44
Correlation with light intensity ... 45
‘Super Sonic White’ New Guinea impatiens ... 45
Effects of age and rooting compound ... 45
Correlation with day length ... 46
Correlation with light intensity ... 47
‘Hot Rose on Gold’ New Guinea impatiens ... 47
viii
Correlation with day length ... 48
Correlation with light intensity ... 48
‘Stained Glass’ coleus propagated in growth chamber ... 49
Effect of age and rooting compound ... 49
Correlation with day length ... 49
Correlation with light intensity ... 49
Correlation with greenhouse temperature ... 50
‘Tango Dark Red’ geranium propagated in growth chamber ... 50
Effect of age and rooting compound ... 50
Correlation with day length ... 50
Correlation with light intensity ... 50
Correlation with greenhouse temperature ... 50
‘Tango’ geranium propagated in growth chamber ... 51
Effect of age and rooting compound ... 51
Correlation with day length ... 51
Correlation with light intensity ... 51
Correlation with greenhouse temperature ... 51
‘Super Sonic White’ New Guinea impatiens propagated in growth chamber ... 51
Effect of age and rooting compound ... 51
ix
Correlation with light intensity ... 51
Correlation with greenhouse temperature ... 52
‘Super Sonic Red’ New Guinea impatiens propagated in growth chamber ... 52
Effect of age and rooting compound ... 52
Correlation with day length ... 53
Correlation with light intensity ... 53
Correlation with greenhouse temperature ... 53
Discussion... 54
Coleus ... 54
Geraniums ... 56
Impatiens ... 59
Conclusion ... 62
Literature Cited ... 63
CHAPTER 3. PROPAGATION SUCCESS OF CALLUSED PLANT CUTTINGS AS INFLUENCED BY STORAGE DURATION AND STORAGE TEMPERATURE ... 138
Abstract ... 139
Introduction ... 141
Callus tissue ... 141
Storage temperature ... 141
x
Materials and methods ... 144
Callusing cuttings ... 144
Callused experiment ... 145
Callused and uncallused experiment 1 ... 145
Callused and uncallused experiment 2 ... 146
Second propagation period ... 147
Results ... 149
Callused experiment ... 149
‘Rocky Mountain Violet’ geranium ... 149
‘Tango Dark Red’ geranium ... 150
‘Super Sonic White’ New Guinea impatiens ... 152
‘Hot Rose on Gold’ New Guinea impatiens ... 154
Callused and uncallused experiment 1 ... 156
‘Rocky Mountain Violet’ geranium ... 156
‘Tango Dark Red’ geranium ... 158
‘Super Sonic White’ New Guinea impatiens ... 159
‘Fanfare Orchid’ impatiens ... 161
Callused and uncallused experiment 2 ... 163
‘Summer Idols Orange’ geranium ... 163
xi
‘Super Sonic White’ New Guinea impatiens ... 164
‘Fanfare Orchid’ impatiens ... 165
Discussion... 167
Conclusion ... 182
Literature Cited ... 184
CHAPTER 4. PROPAGATION SUCCESS OF PLANT CUTTINGS AS INFLUENCED BY STAGE OF PROPAGATION AT TIME OF 1-MCP AND ETHYLENE EXPOSURE AND DURATION OF SUBSEQUENT STORAGE ... 200
Abstract ... 201
Introduction ... 203
Propagation Stage ... 203
Callus tissue ... 204
Ethylene ... 204
Ethylene Inhibitors ... 204
Negative effects of ethylene inhibitors ... 206
Materials and methods ... 207
Cutting production and preparation ... 207
Stage 3 cuttings ... 207
Stage 2 cuttings ... 208
xii
1-MCP and ethylene ... 208
Simulated shipment ... 209
Post-treatment propagation ... 210
Results ... 211
‘Fidelity Coral’ geranium ... 211
‘Tango Dark Red’ geranium ... 211
‘Super Sonic White’ New Guinea impatiens ... 213
‘Super Sonic Red’ New Guinea impatiens ... 214
Discussion... 216
Conclusion ... 220
Literature Cited ... 222
CHAPTER 5. CUTTING STORAGE AND SHIPMENT IN MODIFIED ATMOSPHERE PACKAGING ... ... 230
Abstract ... 231
Introduction ... 233
Storage and shipment trends ... 233
Gasses ... 235
Modified atmosphere packaging ... 236
MAP application ... 238
xiii
Temperature influence on MAP ... 239
Packaging type’s influence on MAP ... 240
MAP Selection ... 241
Materials and methods ... 242
Sourcing and storing cuttings ... 242
Supplier cuttings ... 242
NCSU cuttings ... 242
Post-storage ... 243
Results ... 247
Supplier 1 cuttings ... 247
‘Harmony Magenta’ New Guinea impatiens ... 247
Russian sage ... 248
Supplier 2 cuttings ... 250
‘Cabaret Hot Pink’ calibrachoa... 250
‘Sun Spun Silver’ petunia ... 251
‘Aztec Pink’ verbena ... 252
Supplier 3 cuttings ... 253
‘Comet Red’ argyranthemum ... 253
Stained Glassworks ‘Oompah’ coleus ... 254
xiv
NCSU cuttings ... 256
‘Fanfare Orchid’ impatiens ... 256
‘Super Sonic White’ New Guinea impatiens ... 257
‘Tango’ geranium ... 258
Discussion... 260
Supplier cuttings ... 260
NCSU cuttings ... 266
Conclusion ... 269
xv LIST OF TABLES
CHAPTER 2. PROPAGATION SUCCESS OF UNROOTED PLANT CUTTINGS AS INFLUENCED BY STOCK PLANT AGE AND CROP SEASON
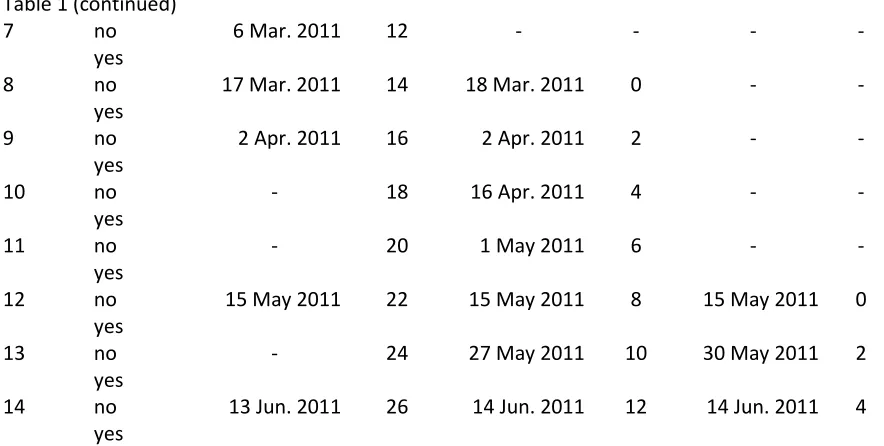
Table 1. Harvest dates of 'Stained Glass' coleus, ‘Tango’ geranium, and ‘Kardino’ geranium cuttings from three stock plant crops of each cultivar ... 65 Table 2. Harvest dates of 'Super Sonic White' and ‘Hot Rose on Gold’ New Guinea impatiens cuttings. ... 68 Table 3. Foliar tissue analysis from fully expanded leaves removed from base of cuttings before propagation ... 69 Table 4. Harvest dates of 'Stained Glass' coleus, ‘Tango Dark Red’ and ‘Tango’ geranium, and 'Super Sonic White' and ‘Super Sonic Red’ New Guinea impatiens cuttings ... 72 Table 5. Effect of age of three crops of 'Stained Glass' coleus stock plants on cuttings'
xvi Table 8. Correlation and correlation significance between 'Stained Glass' coleus crop 1 postharvest parameters and light intensity averaged over 1 - 28 days leading up to cutting harvest date ... 79 Table 9. Correlation and correlation significance between 'Stained Glass' coleus crop 2 postharvest parameters and light intensity averaged over 1 - 28 days leading up to cutting harvest date ... 80 Table 10. Correlation and correlation significance between 'Stained Glass' coleus crop 3 postharvest parameters and light intensity averaged over 1 - 28 days leading up to cutting harvest date ... 82 Table 11. Effect of age of three crops of 'Tango' geranium stock plants on cuttings'
performance when harvested at three similar harvest dates ... 84 Table 12. Effect of season of harvest on 'Tango' geranium cuttings harvested from three stock plant crops of the same number of weeks after transplant, but grown at different times of year ... 86 Table 13. Correlation and correlation significance between geranium 'Tango' postharvest parameters and day length averaged over the 7, 14, 21, and 28 days prior to cutting harvest day ... 88 Table 14. Correlation and correlation significance between 'Tango' geranium crop 1
xvii Table 15. Correlation and correlation significance between 'Tango' geranium crop 2
postharvest parameters and light intensity averaged over 1 - 28 days leading up to cutting harvest date ... 91 Table 16. Correlation and correlation significance between 'Tango' geranium crop 3
postharvest parameters and light intensity averaged over 1 - 28 days leading up to cutting harvest date ... 92 Table 17. Effect of age of three crops of 'Kardino' geranium stock plants on cuttings'
performance when harvested at three similar harvest dates ... 93 Table 18. Effect of season of harvest on 'Kardino' geranium cuttings harvested from three stock plant crops of the same number of weeks after transplant, but grown at different times of year ... 95 Table 19. Correlation and correlation significance between geranium 'Kardino' postharvest parameters and day length averaged over the 7, 14, 21, and 28 days prior to cutting harvest day ... 97 Table 20. Correlation and correlation significance between 'Kardino' geraniums crop 1 postharvest parameters and light intensity averaged over 1 - 28 days leading up to cutting harvest date ... 98 Table 21. Correlation and correlation significance between 'Kardino' geranium crop 2
xviii Table 22. Correlation and correlation significance between 'Kardino' geranium crop 3
postharvest parameters and light intensity averaged over 1 - 28 days leading up to cutting harvest date ... 101 Table 23. Correlation and correlation significance between New Guinea Impatiens 'Super Sonic White' postharvest parameters and day length averaged over 7, 14, 21, and 28 days prior to cutting harvest day ... 102 Table 24. Correlation and correlation significance between 'Supersonic White' New Guinea impatiens postharvest parameters and light intensity averaged over 1 - 28 days leading up to cutting harvest date... 103 Table 25. Correlation and correlation significance between New Guinea Impatiens 'Hot Rose on Gold' postharvest parameters and day length averaged over the 7, 14, 21, and
28 days prior to cutting harvest day ... 105 Table 26. Correlation and correlation significance between 'Hot Rose on Gold' New Guinea impatiens postharvest parameters and light intensity averaged over 1 - 28 days leading up to cutting harvest date... 106 Table 27. Correlation and correlation significance between 'Stained Glass' coleus
xix Table 28. Correlation and correlation significance between 'Stained Glass' coleus
postharvest parameters and light intensity averaged over 1 - 28 days leading up to cutting harvest date ... 109 Table 29. Correlation and correlation significance between 'Stained Glass' coleus
postharvest parameters and mean stock plant growing temperature averaged over 1 - 28 days leading up to cutting harvest date ... 111 Table 30. Correlation and correlation significance between 'Tango Dark Red' zonal geranium postharvest parameters and day length averaged over 7, 14, 21, and 28 days prior to cutting harvest day ... 112 Table 31. Correlation and correlation significance between 'Tango DarkRed' geranium
postharvest parameters and light intensity averaged over 1 - 28 days leading up to cutting harvest date ... 113 Table 32. Correlation and correlation significance between 'Tango Dark Red' geranium postharvest parameters and mean stock plant growing temperature averaged over 1 - 28 days leading up to cutting harvest date ... 114 Table 33. Correlation and correlation significance between 'Tango' zonal geranium
postharvest parameters and day length averaged over 7, 14, 21, and 28 days prior to
xxi Table 40. Correlation and correlation significance between 'Super Sonic Red' New Guinea impatiens postharvest parameters and light intensity averaged over 1 - 28 days leading up to cutting harvest date... 122 Table 41. Correlation and correlation significance between 'Super Sonic Red' New Guinea impatiens postharvest parameters and mean stock plant growing temperature averaged over 1 - 28 days leading up to cutting harvest date ... 123 CHAPTER 3. PROPAGATION SUCCESS OF CALLUSED PLANT CUTTINGS AS INFLUENCED BY STORAGE DURATION AND STORAGE TEMPERATURE
xxii Table 7. Effect of four storage durations and callus presence on ‘Tango Dark Red’ geranium cuttings' time until Stage 3 and quality ... 193 Table 8. Effect of four storage durations and callus presence on ‘Super Sonic White’
impatiens cuttings' time until Stage 3 and quality ... 194 Table 9. Effect of four storage durations and callus presence on ‘Fanfare Orchid’ impatiens cuttings' time until Stage and quality ... 195 Table 10. Effect of four storage durations and callus presence on ‘Summer Idols Orange’ geranium cuttings time until Stage 3 and quality ... 196 Table 11. Effect of four storage durations and callus presence on ‘Summer Idols Red’
geranium cuttings' time until Stage 3 and quality ... 197 Table 12. Effect of four storage durations and callus presence on ‘Super Sonic White’
impatiens cuttings' time until Stage 3 and quality ... 198 Table 13. Effect of four storage durations and callus presence on ‘Fanfare Orchid’
impatiens cuttings' time until Stage 3 quality ... 199 CHAPTER 4. PROPAGATION SUCCESS OF PLANT CUTTINGS AS INFLUENCED BY STAGE OF PROPAGATION AT TIME OF 1-MCP AND ETHYLENE EXPOSURE AND DURATION OF
SUBSEQUENT SIMULATED SHIPMENT
Table 1. Combinations of propagation stage, 1-MCP, ethylene, and storage durations administered to unrooted cuttings of ‘Tango Dark Red’ and ‘Fidelity Coral’ geranium
xxiii Table 2. Cutting harvest dates and propagation schedule. ... 225 Table 3. Effect of stage of propagation on sensitivity to ethylene exposure of 'Fidelity Coral' geranium cutting postharvest parameters after 28 total days of propagation ... 226 Table 4. Effect of stage of propagation on sensitivity to ethylene exposure of ‘Tango Dark Red’ geranium cutting postharvest parameters after 28 total days of propagation ... 227 Table 5. Effect of stage of propagation on sensitivity to ethylene exposure of ‘Super Sonic White’ New Guinea impatiens cutting postharvest parameters after 28 total days of
propagation ... 228 Table 6. Effect of stage of propagation on sensitivity to ethylene exposure of ‘Super Sonic Red’ New Guinea impatiens cutting postharvest parameters after 28 total days of
propagation ... 229 CHAPTER 5. CUTTING STORAGE AND SHIPMENT IN MODIFIED ATMOSPHERE PACKAGING Table 1. Shipping and storage details for each source of unrooted cuttings ... 273 Table 2. Effects of 3 types of MAP bags, supplier control bag, and 4 storage durations on gas concentrations within storage bag of unrooted cuttings of ‘Harmony Magenta’ New Guinea impatiens ... 274 Table 3. Effects of 3 types of MAP storage bags and supplier control bag and 4
xxiv Table 4. Effects of 3 types of MAP bags, supplier control bag, and 4 storage durations on gas concentrations within storage bag of unrooted cuttings of Russian sage ... 276 Table 5. Effects of 3 types of MAP storage bags and 4 pre-shipment storage durations on Russian sage unrooted cutting quality after storage (but before propagation) and post-propagation ... 277 Table 6. Effects of 3 types of MAP bags, supplier control bag, and 4 storage durations on gas concentrations within storage bag of unrooted cuttings of ‘Cabaret Hot Pink’
calibrachoa ... 278 Table 7. Effects of 3 types of MAP storage bags and 4 pre-shipment storage durations on 'Cabaret Hot Pink' calibrachoa unrooted cutting quality post-storage (but before
propagation) and after propagation ... 279 Table 8. Effects of 3 types of MAP bags, supplier control bag, and 4 storage durations on gas concentrations within storage bag of unrooted cuttings of ‘Sun Spun Silver’
xxv Table 11. Effects of 3 types of MAP storage bags and 4 pre-shipment storage durations on 'Aztec Pink' verbena unrooted cutting quality post-storage (but before propagation) and after propagation ... 283 Table 12. Effects of 3 types of MAP storage bags and 4 post-shipment storage durations on 'Comet Red' argyranthemum unrooted cutting quality post-storage (but before
propagation) and after propagation ... 284 Table 13. Effects of 3 types of MAP storage bags and 4 post-shipment storage durations on Stained Glassworks 'Oompah' coleus unrooted cutting quality post-storage (but before propagation) and after propagation ... 285 Table 14. Effects of 3 types of MAP storage bags and 4 post-shipment storage durations on impatiens SunPatiens 'Compact Coral' impatiens unrooted cutting quality post-storage (but before propagation) and after propagation ... 286 Table 15. Effects of 3 types of MAP storage bags and 4 storage durations on 'Fanfare
Orchid' impatiens unrooted cutting quality post-storage (but before propagation) and after propagation. Post-propagation evaluations are based on cuttings surviving at termination .... ... 287 Table 16. Effects of 3 types of MAP storage bags and 4 storage durations on 'Super Sonic White' New Guinea impatiens unrooted cutting quality post-storage (but before
xxvi Table 17. Effects of 3 types of MAP bags, a control bag, and 4 storage durations on gas concentrations within storage bag of unrooted ‘Tango’ geranium cuttings ... 289 Table 18. Effects of 3 types of MAP storage bags and 4 storage durations on geranium 'Tango' unrooted cutting quality post-storage (but before propagation) and after
xxvii LIST OF FIGURES
CHAPTER 2. PROPAGATION SUCCESS OF UNROOTED PLANT CUTTINGS AS INFLUENCED BY STOCK PLANT AGE AND CROP SEASON
Fig. 1. Effect of hormone and harvest time/age of stock plants on postharvest parameters of ‘Stained Glass’ coleus cuttings harvested from three crops, treated without or with
rooting compound, and propagated for 14 d ... 125 Fig. 2. Effect of hormone and harvest time/aging of stock plants on postharvest parameters of ‘Tango’ geranium cuttings harvested from three crops, treated without or with rooting compound, and propagated for 28 d ... 127 Fig. 3. Effect of harvest time on postharvest parameters of ‘Kardino’ geranium cuttings harvested from three crops, treated without or with rooting compound, and propagated for 28 d ... 129 Fig. 4. Effect of harvest time on postharvest parameters of ‘Super Sonic White’ New Guinea impatiens cuttings harvested and treated without or with rooting compound, and
propagated for 28 d ... 131 Fig. 5. Effect of harvest time on postharvest parameters of ‘Hot Rose on Gold’ New Guinea impatiens cuttings harvested and treated without or with rooting compound, and
xxviii Fig. 7. Effect of harvest time on postharvest parameters of ‘Tango Dark Red’ geranium cuttings harvested and treated without or with rooting compound, and propagated for 28 d . ... 134 Fig. 8. Effect of harvest time on postharvest parameters of ‘Tango’ geranium cuttings
harvested and treated without or with rooting compound, and propagated for 28 d ... 135 Fig. 9. Effect of harvest time on postharvest parameters of ‘Super Sonic White’ New Guinea impatiens cuttings harvested and treated without or with rooting compound, and
propagated for 14 d ... 136 Fig. 10. Effect of harvest time on postharvest parameters of ‘Super Sonic Red’ New Guinea impatiens cuttings harvested and treated without or with rooting compound, and
1 Chapter 1. Introduction
Unrooted cuttings are important to the floriculture industry, because many crop species are propagated primarily from stem and stem tip cuttings. The production of healthy rooted cuttings is affected by numerous factors, commencing with healthy stock plants and followed by proper harvest and postharvest handling, and optimal propagation conditions.
Cutting production in the floriculture industry
Production of stock plants typically takes place overseas due to favorable climate and less expensive labor. For example,Impatiens hawkeri Bull.(New Guinea impatiens) cuttings are mostly produced in Central America, Southern Europe, Northern Africa, and Eastern Asia (Lopez and Runkle, 2008). Stock plants are managed to maximize cutting propagation by selecting juvenile source material, rejuvenating source material,
manipulating environmental conditions and physiological status of the stock plant, and cutting selection, i.e. from certain parts of the plant or during certain seasons (Hartmann, et al., 2002). In addition, other factors such as degree-day systems, calendar days, and hours of light can be used to document and predict when optimum rooting may occur from cuttings (Hartmann et al., 2002).
2 To account for shipping time and the schedules of growers who propagate the cuttings after shipment, cutting harvest is not uniformly distributed during the week. Currently cutting suppliers need to harvest all cuttings early in the work week (Sunday to Wednesday), because cuttings harvested later in the week would remain in transit on Sunday or arrive at the customer's greenhouses during the weekend when there may not be sufficient staff to transplant them immediately.
Generally 2-3 days are required for unrooted cuttings to reach propagators after they are harvested (Lopez and Runkle, 2008). During this period, cuttings may encounter adverse conditions including lack of light, temperature extremes, lack of moisture,
pathogens, and ethylene build-up, which can all deteriorate quality of cuttings (Lopez and Runkle, 2008). Once cuttings are damaged, the chance of them regaining quality and resulting in a marketable finished crop is low (Arteca et al., 1996). Consequently, cuttings that arrive at the customers in poor condition may be refused and/or monetary claims may be filed (Lopez and Runkle, 2008). Unusable cuttings can disrupt production schedules, worker scheduling, grower/producer relationships, and reduce profits.
3 Within the storage unit, it is best to maintain nearly 100 percent humidity, and the
temperature should be as low as the hardiness of the given species can tolerate” (Hartmann et al., 2002).
If cuttings could be easily stored or shipped without a loss in quality, producers could harvest cuttings constantly during the week, even daily, and cuttings could be stored until shipped, or be stored upon arriving at customer's greenhouses during times when employees are not available to propagate.
The ability to store cuttings and maintain quality during shipping would help producers regulate the supply during periods of both surplus cutting production and peak demand by growers (Lopez and Runkle, 2008), thus, accommodating the gradual increase in cutting supply necessary to reach the sharp peaks in demand in grower’s production
schedules. Stock plant age
The age of the stock plant from which the cutting is taken can determine the ability of cuttings to readily form roots. Often, especially in woody species, it is not the
4 juvenile root stock, growing shoots from sphaeroblasts, and producing plants in
micropropagation (Hartmann et al., 2002).
When cuttings are taken from a stock plant in a juvenile stage, they root more readily. It is advantageous for cutting producers to utilize stock plants that are in or being maintained in a juvenile phase.
Using cuttings of the wrong age can cause problems such as slow rooting and lack of branching (Gibson, 2006). Plant material used for cuttings should contain actively growing shoots, but should lack fully developed flowers (Gibson, 2006).
Rooting compounds
Rooting compounds are considered a necessity to economically root some species, while for other species they are not required, but can be used to “accelerate root initiation, increase root uniformity, and increase the number and quality of roots produced” (Dole and Wilkins, 2005). Rooting compounds contain auxins such as indole-3-butyric acid and
naphthaleneacetic acid, and may also contain a fungicide (Dole and Wilkins, 2005). Stage of propagation at shipment
5 the same size for shipping as unrooted cuttings if shipped without substrate, but Faust et al. (2006) state that they are usually packed with fewer cuttings per box. The process of callusing cuttings requires labor and propagation time between harvest and shipping. Faust et al. (2006) note that rooted cuttings (Stage 3) are subject to physical damage and much concern must be taken with packing them in protective shipping materials.
Callus tissue. When cuttings are placed in a proper rooting environment, it is common for callus tissue to form at their basal end (Hartmann et al., 2002). Callus tissue is composed of parenchyma cells that grow adjacent to other cell types at the base of the cutting (Hartmann et al., 2002). With species that root easily, the formation of callus tissue and adventitious roots are independent processes (Dole and Hamrick, 2006; Hartmann et al., 2006), but both involve cell division and require similar environmental conditions (Hartmann et al., 2002).
Storage temperature
Reports have been made that reduced temperatures help maintain the quality of Pelargonium x hortorum L.H. Bailey (geranium) cuttings (Arteca et al., 1996). However, if temperatures are too cold, unrooted cuttings - especially of tropical and subtropical species - can exhibit chilling injury and necrosis (Lopez and Runkle, 2008). New Guinea impatiens, for example, have shown chilling sensitivity below 6°C (Erwin, 1995).
6 short periods of time during harvest and distribution (Lopez and Runkle, 2008). Declines in quality due to high temperatures have been observed in numerous studies. Arteca et al. (1996) stored ‘Snowmass’ geranium cuttings at temperatures ranging from 4 to 25°C and “a gradual decline in visual rating was observed with increasing temperatures.” In the same study, leaf weight and chlorophyll content also decreased as storage temperatures reached higher levels. Similar results occurred when cuttings were allowed to grow for 10 d after storage: as temperature increased, first a reduction in visual rating and root weight occurred, followed by reductions in leaf weight and chlorophyll content (Arteca et al., 1996). Both geraniums and New Guinea impatiens have been reported to lose a higher fresh weight percentage as storage temperature increases (Arteca et al., 1996; Lopez and Runkle, 2008).
Storage duration
7 Ninety seven percent or more of ‘Harmony White’ cuttings survived after 1 d of storage at temperatures between 0 and 30°C. Survival of cuttings stored for 5 d was lower: 4% survival was seen at 5°C and 70% at 30°C (Lopez and Runkle, 2008).
Storage duration also affects root development. Root initiation and development decrease when cuttings are exposed to extreme storage temperatures for extended periods of time. Lopez and Runkle reported that ‘Harmony White’ cuttings stored at 30°C for 5 d formed 86% less roots than the unstored control (2008). Similar effects of decreased root development have been reported in geraniums (Arteca et al., 1996; Mutui et al., 2005). Ethylene
8 Ethylene inhibitors
A number of compounds are known ethylene inhibitors, including
1-methylcyclopropene (1-MCP) and its analogs, "carbon dioxide, silver thiosulfate (STS), aminoethyoxyvinylglycine (AVG), 2,5-norbornadiene (2,5-NBD) and DACP
[diazocyclopentadiene]" (Blankenship and Dole, 2003).
Silver thiosulfate (STS) is an effective inhibitor of ethylene and has been shown to slow chlorophyll degradation of stored Pelargonium cuttings (Purer and Mayak, 1987) and limit leaf yellowing during storage of ‘Isabel’ geranium and Hibiscus rosa sinensis L.
(hibiscus) cuttings (Serek et al., 1998). It can be concluded that ethylene is involved in the loss of cutting quality of the untreated cuttings since the anti-ethylene agent overcame the leaf yellowing.
Another ethylene inhibitor, 1-methylcyclopropene (1-MCP), is thought to be effective by occupying ethylene receptors such that ethylene cannot bind to them
9 Serek et al. (1998) reported that 1-MCP was more effective in controlling ethylene damage when applied prior to ethylene exposure, as opposed to simultaneously with the exogenous ethylene application suggesting that 1-MCP should be applied to crops before ethylene exposure occurs.
While some crops such as Brassica oleracea var. italica L. (broccoli) (Able et al., 2002a) and Citrillus lanatus (Thunb.) Matsum. & Nakai (watermelon) (Blankenship and Dole, 2003) benefit from 1-MCP in the presence of endogenous or exogenous ethylene, others, such as Brassica rapa var. chinensis (pak choy), only benefit in the presence of exogenous ethylene (Able et al., 2002a). Able et al. (2002b) reported six Asian leafy vegetables had delayed leaf senescence when treated with both exogenous ethylene and 1-MCP, but only two of the six benefited from 1-MCP when no exogenous ethylene was present (Able et al., 2002b).
10 Negative effects of ethylene inhibitors
Ethylene inhibitors such as STS and 1-MCP reduce rooting of Pelargonium species, Dendranthema grandiflora (Ramat.) Kitamura, and (hibiscus) cuttings (Paton and Schwabe, 1987; Purer and Mayak, 1987; Serek et al., 1998). These studies suggest a role of ethylene in adventitious rooting, and ethylene has been reported to increase adventitious rooting in mung bean seedlings (Reid et al., 1985).
While possibly beneficial during shipping, anti-ethylene agents could inhibit the ability of cuttings to root once they are planted (Serek et al., 1998). Additionally, STS is toxic to plant tissues at levels not much higher than the levels effective at inhibiting ethylene action (Serek et al., 1998), and can make plants susceptible to Pythium (Arteca et al., 1996). Oxygen and carbon dioxide
Oxygen levels must remain within a range the plant can withstand, even during changes in temperature and humidity (Beaudry, 2000). If oxygen levels get too low, anaerobic activity can occur; if they get too high, less respiratory reduction will occur and the benefits of a low oxygen level storage will be negated (Beaudry, 2000). Low oxygen levels reduce ethylene biosynthesis, but there are conflicting reports on whether or not reduced oxygen concentrations lower the sensitivity of products to ethylene (Beaudry, 2000).
Responses to oxygen and carbon dioxide vary depending on the plant organ
11 carbon dioxide inhibits ethylene action as well as ACC synthase activity and ACC oxidase activity (Watkins, 2000).
Interactions of factors also affect the concentrations of carbon dioxide that can be tolerated. For example, excessively high or low levels of carbon dioxide combined with warm temperature and long durations can be detrimental to horticultural commodities, cause physiological disorders, and increase susceptibility to decay (Watkins, 2000). Additionally, some commodities such as Asparagus officinalis L. (asparagus) and Prunus persica L. (peach) can handle high levels of carbon dioxide, but only for short periods of time; asparagus and Capsicum annuum L. (chili pepper) can tolerate higher carbon dioxide levels at lower temperatures (Watkins, 2000).
Modified atmosphere packaging
Modified atmosphere packaging (MAP) was first evaluated for use in the 1940’s when packages were tested for their ability to slow apple ripening by lowering oxygen levels (Beaudry, 2000). Since then, its horticultural use has spread to various fruits, vegetables, and herbs. MAP can “benefit fruit and vegetable storage and handling by providing atmospheres (decreased oxygen and increased carbon dioxide) that can slow down the degenerative processes of senescence.” (Lange, 2000).
12 from contamination and spread of decay, (Kader and Watkins, 2000), respiration reduction, reduction in ethylene synthesis and perception, (Beaudry, 2000; Fonseca et al., 2002), and reductions in degradation of chlorophyll and cell walls (Beaudry, 2000).
One objective of MAP is to lower oxygen levels in the atmosphere around the plants (Beaudry, 2000; Fonseca et al., 2002) enough to prevent losses in quality, which in turn lengthens the possible storage time of the plant material (Beaudry, 2000). Benefits can be achieved when packaging with an appropriate oxygen transmission rate (OTR) is used (Lange, 2000). Respiration and senescence can be slowed due to lower oxygen levels inside the packaging, while aerobic respiration is maintained, and anaerobic respiration, which can cause microorganism growth and rapid product deterioration, can be prevented (Lange, 2000).
The films used in MAP may be microperforated or have a microporous membrane. Microperforated films have perforations applied by laser, heat, or pins, and microporous membrane films are composed of a highly permeable membrane attached to a film package (Lange, 2000). The films can also be used as carriers for chemicals such as ethylene
absorbers or fungicides, which can often be more beneficial than the modified atmosphere inside the package (Kader and Watkins, 2000).
13 added to “scavenge” oxygen, carbon dioxide, and/or ethylene (Kader and Watkins, 2000). This procedure, while adding cost, ensures that the desired atmosphere is established quickly (Kader and Watkins, 2000). Active MAP is typically used for fruit crops that have a short shelf life such as strawberries (Lange, 2000).
Passive modified atmospheres are created inside the packaging as the product consumes oxygen and produces carbon dioxide through respiration (Beaudry, 2000; Kader and Watkins, 2000). According to Fick’s Law, these changes will cause oxygen to enter and carbon dioxide to exit the package. However, constant oxygen levels will only result when the product consumes the same amount of oxygen as is entering the package; this can only occur if respiration is constant. Likewise, carbon dioxide inside the package will only be constant if the product produces the same amount of carbon dioxide that is exiting the package (Beaudry, 2000).
Ideally, passive MAP will cause the OTR of the packaging to match the respiratory requirements of the commodity contained within (Lange, 2000). Numerous factors influence the optimum OTR in passive MAP, and because of the difficulty in regulating atmospheres created passively, actively creating the modified atmosphere is often more preferred (Kader and Watkins, 2000).
14 a result of the low oxygen levels; these products are said to have no “safe working
atmosphere” and include Fragaria × ananassa Duch.(strawberry) fruits and asparagus spears (Beaudry, 2000). Non-damaging oxygen limits below which injury may occur have been determined for a variety of horticultural commodities and are compiled by Beaudry (2000). It should also be noted that responses differ among various plant organs. Whether or not reduced oxygen is beneficial ultimately depends on the balance of positive and negative reactions of the particular plant organ to low oxygen (Beaudry, 2000).
Carbon dioxide level is another important factor when using MAP. The effects of high carbon dioxide are similar to the effects of low oxygen, and a high carbon dioxide level by itself or in conjunction with a low oxygen level determines the overall beneficial or detrimental effects (Watkins, 2000).
15 change in proportion to the changes in product respiration as the temperature fluctuates (Lange, 2000).
The respiration of plant material responds to oxygen concentration, which is also temperature sensitive (Beaudry, 2000). If temperature increases and the respiratory demand for oxygen increases faster than the oxygen permeation increases, then target gas levels cannot be maintained (Beaudry, 2000). Several theoretical solutions have been investigated to solve this imbalance, including films that are sensitive to temperature and have higher oxygen permeability, packages that sense the status of the enclosed plant product or the environment inside the package and respond by altering the oxygen permeability, and packaging designed to work at higher temperatures than typically encountered during normal use (Beaudry, 2000).
Packaging type’s influence on MAP. MAP packaging can vary in the polymer
composition, film thickness, and layer ratio. The package-product relations such as product volume:film surface area and volume unoccupied by the commodity also come into play (Lange 2000). Mechanical problems with MAP packaging may occur because the material must be flexible and easy to handle, but also strong enough so it does not get damaged during normal handling procedures (Kader and Watkins, 2000). The films may also have negative effects such as slowing down the cooling of the product and promoting
16 MAP selection. The benefits of MAP must outweigh the associated extra costs including materials and labor. The cost of packaging and modifications in packaging procedures can reduce profit margins, usually for producers/processors (Kader and
Watkins, 2000). It is important for the reduction in respiration to add enough value to the product to offset the cost of using MAP technology (Beaudry, 2000).
Ideally when choosing a package, the product respiration rate at the desired gas concentrations and storage temperature and the film transmission rate of gasses at the storage temperature can be used to select a bag size for a given weight of product or a weight of product for a given bag size (Moyls et al., 1998). Films have an Arrhenius relationship in regards to their transmission characteristics, so OTR, carbon dioxide
17 LITERATURE CITED
Able, A.J., T.J. O'Hare, L.S. Wong, and A. Prasad. 2002a. 1-MCP is more effective on a floral brassica (Brassica oleracea var. italica L.) than a leafy brassica (Brassica rapa var. chinensis) Postharvest Biol. Technol. 26:147-155.
Able, A.J., L.S. Wong, A. Prasad, and T.J. O'Hare. 2002b. The effects of
1-methylcyclopropene on the shelf life of minimally processed leafy Asian vegetables. Postharvest Biol. Technol. 27:157-157-161.
Arteca, R.N., J.M. Arteca, T.-W. Wang, and C.D. Schlagnhaufer. 1996. Physiological,
biochemical, and molecular changes in Pelargonium cuttings subjected to short-term storage conditions. J. Amer. Soc. Hort. Sci. 121:1063-1068.
Beaudry, R.M. 2000. Responses of horticultural commodities to low oxygen: Limits to the expanded use of modified atmosphere packaging. HortTechnology 10:491-500. Blankenship, S.M. and J.M. Dole. 2003. 1-Methylcyclopropene: a review. Postharvest Biol.
Technol. 28:1-25.
Dole, J.M., J.L. Gibson and H.F. Wilkins, 2006. Crop-by-crop cutting propagation, p. 230-362. In: Dole, J.M. and J.L. Gibson (eds.). Cutting propagation. Ball Publishing, Batavia, IL. Dole, J.M. and H.F. Wilkins. 2005. Floriculture principles and species. Pearson Prentice
Hall, Upper Saddle River, NJ.
18 Erwin, J., 1995. Light and temperature, p. 41-54. In: Banner, W., and M. Klopmeyer (eds.).
New Guinea impatiens. Ball Publishing, Batavia, IL.
Faust, J.E., A.L. Enfield, S.M. Blankenship, and J.M. Dole, 2006. Postharvest, p. 145-154. In: Dole, J.M. and J.L. Gibson (eds.). Cutting propagation. Ball Publishing, Batavia, IL. Fonseca, S.C., Oliveira, A.R. and Brecht, J.K. 2002. Modeling respiration rate of fresh fruits
and vegetables for modified atmosphere packages: a review. J. Food Eng. 52:99-119.
Gibson, J.L., 2006. Stock plant management, p. 31-38. In: Dole, J.M. and J.L. Gibson (eds.). Cutting propagation. Ball Publishing, Batavia, IL.
Hartmann, H.T., D.E. Kester, F.T. Davies Jr., and R.L. Geneve. 2002. Plant propagation: principles and practices. 7th ed. Prentice Hall, Upper Saddle River, NJ.
Kader, A.A., and C.B. Watkins. 2000. Modified atmosphere packaging – toward 2000 and beyond. HortTechnology 10:483-486.
Lange, D.L. 2000. New film technologies for horticultural products. HortTechnology 10:487-490.
Lopez, R.G. and E.S. Runkle. 2008. Photosynthetic daily light integral during propagation influences rooting and growth of cuttings and subsequent development of New Guinea impatiens and petunia. HortScience 43:2052-2059.
19 polyethylene films for modified atmosphere packaging. Trans. Amer. Soc. Agr. Eng 41:1441-1446.
Muller, R., Serek, M., Sisler, E.C., Andersen, A.S., 1997. Poststorage quality and rooting ability of Epipremnum pinnatum cuttings after treatment with ethylene action inhibitors. J. Hort. Sci. 72:445-452.
Muller, R., Serek, M., Sisler, E.C., Anderson, A.S., 1998. Ethylene involvement in leaf
abscission, chlorosis, and rooting of Codiaeum variegatum var. pictum (Lodd.) Muell. 'Aucubaefolia'. Gartenbauwissenchaft 63:66-71.
Mutui, T.M., H. Mibus, and M. Serek. 2005. Effects of thidiazuron, ethylene, abscisic acid and dark storage on yellowing and rooting of Pelargonium cuttings. J. Hort. Sci. Biotechnol. 80:543-550.
Paton, F. and W.W. Schwabe. 1987. Storage of cuttings of Pelargonium x hortorum Bailey. J. Hort. Sci. 62:79-87.
Purer, O. and S. Mayak. 1987. Pelargonium cuttings – effect of growth regulators. Acta Hort. 261:347-354.
Rapaka, V.K., J.E. Faust, J.M.Dole and E.S. Runkle. 2008. Endogenous carbohydrate status affects postharvest ethylene sensitivity in relation to leaf senescence and
20 Reid, M.S., J.A. Robbins, J.L. Paul, and T.L. Rost. 1985. The effect of ethylene on adventitious
root formation in mung bean (Vigna radiata) cuttings. J. Plant Growth Regulation 4:147-157.
Serek, M., A. Prabucki, E.C. Sisler, and A.S. Andersen. 1998. Inhibitors of ethylene action affect final quality and rooting of cuttings before and after storage. HortScience 33: 153-155.
21 Chapter 2. Propagation Success of Unrooted Plant Cuttings as Influenced by Stock Plant Age and Crop Season
Brigitte D. Crawford1 and John M. Dole
Department of Horticultural Science, North Carolina State University, Raleigh, NC, 27695
Received for publication ________. Accepted for publication _______. We acknowledge the assistance of the Floriculture Research Alliance’s P3 Research Group, NCSU statisticians, floriculture research group’s technicians, graduate students, and student workers, and the NCSU Department of Horticultural Science.
1
22 Abstract. The production of healthy rooted cuttings is affected by numerous factors, including the age the stock plant from which the cutting is taken and environment for the stock plants and cuttings. The objectives of this study were to quantify the effect of stock plant age and production environment on Pelargonium x hortorum L.H. Bailey (geranium), Impatiens hawkeri Bull. (New Guinea impatiens, NGI), and Solenostemon scutellarioides (L.) Codd (coleus) cutting quality and propagation success, and to determine the interaction of stock plant age, production environment, and rooting compounds on propagation success. For coleus and one of two cultivars of NGI cuttings, shoot and root fresh and dry weights increased from first harvest to last harvest (34 weeks for coleus and 14 weeks for NGI). Rooting compounds increased root fresh weight of one of two NGI cultivars, but not of coleus cuttings. For geraniums, shoot and root fresh and dry weights initially increased then decreased from first harvest to last harvest (28 weeks), and use of rooting compound increased root fresh and dry weights. For coleus and geraniums, cuttings harvested at the same time from stock plants of three distinct ages had similar fresh and dry weights, quality ratings, and number of abscised or yellow leaves. Variations in postharvest quality
characteristics may be due to environmental parameters such as day length and light
24 The production of healthy rooted cuttings is affected by numerous factors,
commencing with healthy stock plants and followed by proper harvest and postharvest handling, and optimal propagation conditions. Stock plants are managed to maximize cutting propagation by selecting juvenile source material, rejuvenating source material, manipulating environmental conditions and physiological status of the stock plant, and cutting selection, i.e. from certain parts of the plant or during certain seasons (Hartmann et al., 2002). In addition, other factors such as degree-day systems, calendar days, and hours of light can be used to document and predict when optimum rooting may occur from cuttings (Hartmann et al., 2002).
Stock plant age. The age of the stock plant from which the cutting is taken can determine the ability of cuttings to readily form roots. Often, especially in woody species, it is not the chronological age of the stock plant, but rather the ontogenetic age, or what phase the plant is in (embryonic, juvenile, transitional, or mature) (Hartmann et al., 2002). In these species, various methods are utilized to rejuvenate plants to keep them “young” including inducing adventitious buds to grow from roots, chemical manipulation, grafting onto juvenile root stock, growing shoots from sphaeroblasts, and producing plants in micropropagation (Hartmann et al., 2002)
25 Using cuttings of the wrong age can cause problems such as slow rooting and lack of branching (Gibson, 2006). Plant material used for cuttings should contain actively growing shoots, without fully developed flowers (Gibson, 2006).
Rooting compound. Rooting compounds are considered a necessity to economically root some species, while for other species they are not required, but can be used to
“accelerate root initiation, increase root uniformity, and increase the number and quality of roots produced” (Dole and Wilkins, 2005). Rooting compounds contain auxins such as indole-3-butyric acid and naphthaleneacetic acid, and may also contain a fungicide (Dole and Wilkins, 2005).
26 Materials and Methods
Stock plants of ‘Stained Glass’ coleus, ‘Tango’ and ‘Kardino’ zonal geraniums, and ‘Super Sonic White’ and ‘Hot Rose on Gold’ New Guinea impatiens were established from rooted cuttings received from commercial suppliers. Stock plants for this study were rooted from cuttings harvested from the original stock plants. Coleus plants were grown in 20 cm plastic pots and geraniums and impatiens were grown in 15 cm plastic pots. Stock plants were grown in ambient air in a glass glazed greenhouse at 24°C days and 18°C nights. Shade cloth (Revolux XLS 15 Fire Retardant 50% shade cloth, Ludvig Svensson, Kinna, Sweden) covered the growing area near glass height during the night from late fall through mid-spring and during the middle of the day from late mid-spring to mid fall to reduce greenhouse temperatures. Geranium and coleus were fertigated via drip irrigation with200 mgL-1 N from a water soluble fertilizer (Peters Professional 20N-4.4P-16.6K, Scotts-Sierra
Horticultural Products Co., Marysville, OH) and impatiens were fertigated via drip irrigation with 150 mgL-1 N from the same fertilizer. Substrate was treated with flowable lime (Cleary Limestone F Liquid Flowable Limestone, Cleary, Dayton, NJ) to increase substrate pH as necessary.
27 Cuttings were harvested approximately every two weeks (Tables 1 and 2). Cuttings were sorted into two equal groups of 20 cuttings (5 repetitions of 4) by stem caliper, and treated without or with rooting compound powder (0.20% 1-naphthaleneacetamide; 4.04% thiram – tetramethyl thiuramdisulfide, Green Charm Rootone Brand F, Gro Tec, Inc.,
Madison, GA). For cuttings treated with rooting compound, the bottom 0.6 cm of stem was dipped into the powder. For geraniums and coleus, two additional stock plant crops were transplanted from rooted cuttings and cuttings were harvested as above, starting 4 and 6 months after the first crop’s harvests began. Harvests continued from original stock plants until three harvests were made from the third set of stock plants.
Cuttings were propagated for 14 d (coleus) or 28 d (geraniums and impatiens) in 1206 packs in a peat based commercial substrate (Fafard 4P, Conrad Fafard, Inc., Agawam, MA). Cutting propagation conditions were the same as the stock plants’, with the addition of mist applied at a frequency of 6 s every 30 min.
28 3=1-5 roots at least 2.5 cm, or 6-10 roots less than 2.5 cm; 2=1-5 roots less than 2.5 cm; 1=no roots). If less than five cuttings were surviving at termination, roots of all surviving cuttings were rated. Five cuttings in each treatment were used to record shoot and root weights. Substrate was removed from cuttings’ roots by submerging in and/or spraying with water. After rinsing roots, excess water was blotted off the cuttings with felt fabric or paper tissues. Roots were separated from shoots, and fresh weights of shoots and roots were taken. Shoots and roots were then dried in paper envelopes or bags in a Precision Thelco laboratory oven (Thermo Fisher Scientific Inc., Waltham, MA) at 70°C for a minimum of 48 hours and dry weights were recorded.
Stock plant substrates were tested for pH and EC every two weeks (data not presented) and foliar tissue samples were collected from stock plants and dried for tissue analysis with every cutting harvest. Foliar tissue samples were collected from fully
expanded leaves removed from base of cuttings before propagation, then dried and ground to pass a ≤ 0.5 mm sieve prior to being sent to a commercial tissue analysis lab (Table 3).
29 Cuttings were harvested as described above approximately every 4 weeks (Table 4), sorted into two equal groups based on stem caliper, and treated with or without rooting compound powder (0.1% indole-3-butyric acid, Hormodin 1, OHP, Inc., Mainland, PA), as described above. Foliar tissue samples were not collected.
Cuttings were propagated for 14 d (coleus and impatiens) or 28 d (geraniums) in a peat based commercial substrate (Fafard 4P, Conrad Fafard, Inc., Agawam, MA) in 1206 packs. Cuttings were propagated in a growth chamber set at 21°C under 12 h per day of cool white fluorescent lighting emitted from fixtures with two 1.2 m 34 watt bulbs each. Packs were placed in single rows on crates such that the surface of each pack was 15 cm beneath bulbs.
The chamber floor was wet down daily to maintain high humidity. Cuttings were hand misted daily until turgidity could be maintained without misting; thereafter cuttings were misted every 2 d or as needed to keep substrate moist. After propagation, data was collected as above.
30 Results
‘Stained Glass’ coleus
Effects of age and rooting compound in crop 1. The number of yellow/necrotic leaves per cutting was affected by an interaction between rooting compound and time (Fig. 1A). Cuttings from harvests 3, 4, and 11-14 had more yellow/necrotic leaves than all other harvests, which had 0.00 to 0.20 yellow/necrotic leaves. Cuttings from harvests 11-13 had lower average shoot quality ratings than most other harvests, 4.63, 4.45, and 4.35,
31 weights increasing over time. Weights reached a maximum for both without and with rooting compound in harvest 15, 0.038 and 0.055 g, respectively, and then declined again with the last two harvests.
The number of abscised leaves per cutting was not significantly affected by rooting compound or time; survival for coleus cuttings was 100% for all treatments (data not presented).
Effects of age and rooting compound in crop 2. In earlier harvests, hormone treated cuttings tended to have higher numbers of yellow/necrotic leaves and lower or the same shoot quality ratings as untreated cuttings (Fig. 1A). For example, cuttings from the second harvest had 0.15 and 1.10 yellow/necrotic leaves for without and with hormone,
respectively. In later harvests cuttings treated with hormone had lower numbers of yellow/necrotic leaves and higher or the same shoot quality ratings as untreated cuttings. Shoot quality ratings in the last few harvests were all near 5.00 (Fig. 1B).
32 Abscised leaves were not significantly affected by hormone or age (data not
presented). Survival was 100% for all treatments (data not presented).
Effects of age and rooting compound in crop 3. Shoot fresh and dry weights decreased with age (Fig. 1D and 1F). Yellow/necrotic leaves, shoot quality, and root fresh and dry weights and quality were not affected by hormone or age (Fig. 1A, 1B, 1E, 1G, 1C). Abscised leaves were not affected by hormone or age; survival was 100% for all treatments (data not presented).
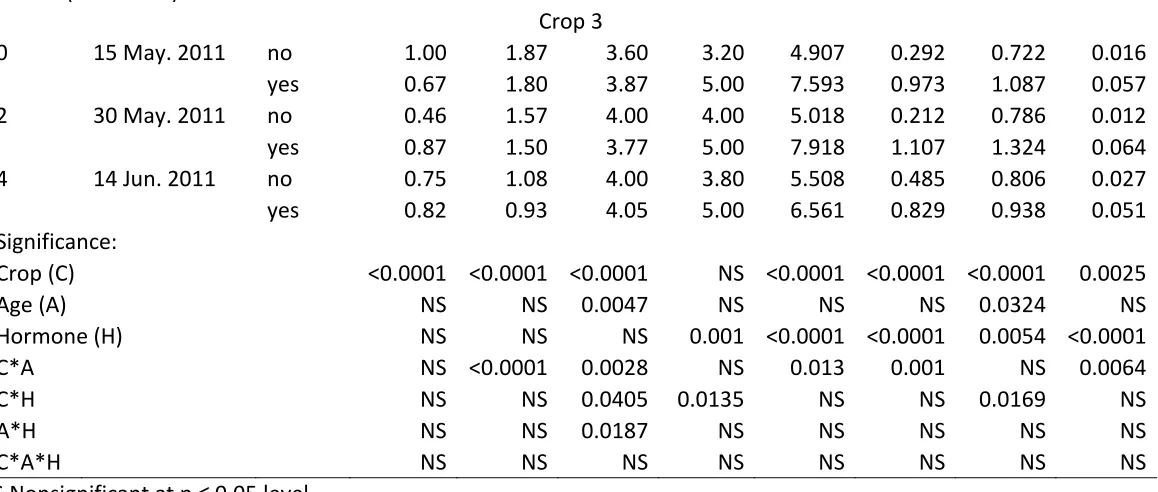
Age analysis of cuttings harvested on same dates. Crop age did not significantly affect any postharvest parameters analyzed (Table 5; data not presented for abscised leaves or survival). Over the three harvests analyzed, yellow/necrotic leaves decreased, shoot quality increased, shoot dry weight decreased and root fresh and dry weights decreased. Hormone increased root dry weights. All three crop ages showed decreased shoot fresh weights over the three harvest dates.
Harvest season analysis of cuttings of same age. The crop whose harvests started in May 2011 had the least amount of yellow/necrotic leaves (Table 6). For November 2010 and March 2011 crops, yellow/necrotic leaves increased from first to third harvest, while for May 2011 crop, there were more yellow/necrotic leaves in cuttings from initial harvests and less in later harvests.
33 decreased over harvests of the November 2010 and March 2011 crops, while in the May crop shoot quality increased to 5.00 and root quality was 5.00 for all harvests.
Crop season interacted with age to affect shoot fresh and dry weights. The November 2010 crop had the lowest shoot fresh and dry weights and the May 2011 crop had the highest. Weights decreased from the first to third harvest in all three crops.
Root dry weights decreased over harvests in the November 2010 and May 2011 crops, but increased in the March 2011 crop (Table 6). Root dry and fresh weights were highest in the May 2011 crop and lowest in the November 2010 crop.
A three-way interaction between crop season, stock plant age at harvest date and hormone affected yellow/necrotic leaves and shoot quality; however, effects appear to be inconsistent, and could be due to cuttings from the second harvest of crop 2 having an unusually high number of yellow/necrotic leaves and consequently a low shoot quality rating. If ignoring this treatment, yellow/necrotic leaves were generally low (0.55 and lower) and shoot qualities high (4.60 and higher). Abscised leaves and survival were not significantly affected (data not presented).
34 over up to at least 28 DPH. Shoot quality was positively correlated to day length averaged over 21 and 28 DPH. For crop 3, yellow/necrotic leaves, shoot fresh and dry weights, and root dry weight were negatively correlated with day length averaged over 28 days DPH. Shoot quality was positively correlated with day length averaged over 28 DPH.
Correlation with light intensity. For crop 1, root quality, and shoot and root fresh and dry weights were positively correlated with light intensity averaged over at least the first 28 DPH (Table 8).
For crop 2, yellow/necrotic leaves were negatively correlated with light intensity averaged over up to at least 28 DPH (Table 9). Shoot quality was positively correlated with average light intensity over up to at least 28 DPH. Root quality was positively correlated with light intensity averaged over 3 up to at least 28 DPH. Root fresh and dry weights, and shoot dry weights were positively correlated with light intensity averaged over at least the first 28 DPH, with the exception of light intensity averaged over 2 and 3 DPH correlating with shoot dry weight.
35 Root dry weight was positively correlated with light intensity averaged over 1-7 DPH, and negatively correlated with light intensity averaged over 13-28 DPH.
‘Tango’ geranium
Effects of age and rooting compound in crop 1. Cuttings from the initial harvests had few abscised and yellow/necrotic leaves and numbers increased then decreased as harvests continued (Fig. 2A and 2B). The highest numbers of abscised and yellow/necrotic leaves 0.97 and 2.50 leaves respectively, were observed in cuttings from harvest 11 treated with rooting compound.
Shoot quality ratings decreased concurrently as damaged leaves increased (Fig. 2C). Shoot fresh weights were generally higher when hormone was applied (Fig. 2E). For example, cuttings from the first harvest had shoot fresh weights of 6.342 and 7.706 g for without and with hormone, respectively. Shoot fresh weights increased then decreased over time and cuttings from the last harvest had similar weights to those of the first harvest, 7.096 and 8.058 g for without and with hormone, respectively.
Root fresh weights were generally higher when hormone was used (Fig. 2F). Root fresh weights also increased then decreased over time; weights were at their maximum in cuttings from harvest 4 (3.233 and 3.868 g for without and with hormone, respectively) and weights from the last harvest were close to weights from the first harvest.
36 almost all harvests and trends over time were similar to those of root fresh weights (Fig. 2H). Root quality ratings were not significantly affected by the use of rooting compound or by the age of stock plants increasing, and ranged from 3.80-5.00 (Fig. 2D). ‘Tango’ geranium cuttings had 100% survival in all treatments except for cuttings from harvest 12 not treated with rooting compound, which had 90% survival (data not presented).
Effects of age and rooting compound in crop 2. Abscised leaves generally increased as age increased (Fig. 2A). Cuttings from the first two and the last harvest had less
yellow/necrotic leaves, while cuttings from the middle harvests, harvests 3-6, tended to have more (Fig. 2B). Shoot quality ratings ranged from 3.4-4.1 for all treatments (Fig. 2C). Shoot fresh weights were generally higher in earlier harvests than in later harvests (Fig. 2E). Root fresh weights were usually higher in cuttings treated with rooting compound and decreased as time went on; however, cuttings from the last harvest had root weights that were higher than the previous 4 harvests (Fig. 2F). Cuttings without hormone had lower root dry weight than cuttings with hormone, 0.055 and 0.084 g, respectively (Fig. 2 H).
Root quality and shoot dry weight were not significantly affected by hormone or age (Fig. 2D and 2G). Survival was 100% for the majority of treatments; all treatments had 80% or higher survival (data not presented)
37 cuttings treated with hormone (Fig. 2D, 2E, 2G, 2F, 2H, respectively). Abscised leaves were not significantly affected by hormone or age (Fig. 2A). Survival was 100% in all treatments (data not presented).
Age analysis of cuttings harvested on same dates. Crop age affected shoot fresh weight. The youngest crop had the lowest shoot fresh weight, 6.251 g, while the oldest crop had the highest shoot fresh weights 7.470 g (Table 11). Hormone increased root quality, root fresh and dry weights, and shoot fresh and dry weights. Crop age interacted with the date of harvest to affect the number of abscised leaves, yellow/necrotic leaves, root fresh and dry weights, and cutting survival.
The crop with the most leaf damage varied by harvest date (Table 11). The youngest crop had the most abscised leaves at the first and third harvest dates, while the oldest crop had the most abscised leaves for the second harvest date and the most yellow/necrotic leaves for the first harvest date. The middle aged crop had the most yellow/necrotic leaves for the second and third harvest dates. Despite the significant effects, the number of damaged leaves did not vary a lot between crops – for abscised leaves, a range of 0.63 (oldest crop) to 0.76 (youngest crop) was observed, and for yellow/necrotic leaves, a range of 1.46 (oldest crop) to 1.81 (middle aged crop) was observed.
38 weights for the third harvest date. Overall, the youngest crop also had the lowest weights, but the middle aged crop had the highest. Survival was 100% for most treatments and above 80% in all treatments (Table 11).
Shoot fresh weights were higher in cuttings with hormone, but the difference between with and without hormone diminished from the first to third harvest (Table 11). Shoot dry weights followed a similar trend.
In the first harvest, cuttings from the oldest crop had a lower shoot quality rating if treated with hormone, while cuttings from the younger crops had higher ratings if treated with hormone (Table 11). In the second harvest, cuttings from the oldest crop had a higher rating if treated with hormone, while the younger crops had lower ratings if treated. In the third harvest, cuttings from the two older crops had lower shoot ratings if treated with hormone, while cuttings in the youngest crop had higher shoot ratings if treated with hormone.
Harvest season analysis of cuttings of same age. Cuttings from the first harvests had lower shoot dry weights, while cuttings from the second and third harvests had higher average shoot dry weights (Table 12). Hormone increased shoot fresh weights, root fresh weights, and root dry weights. Cuttings from the December crop had 0.00 to 0.05 abscised leaves (Table 14), while the March and May crops averaged 0.25 and 0.76 leaves,
39 Crop season interacted with age at the three harvests to affect yellow/necrotic leaves and shoot quality. In cuttings from the December and May crop harvests, yellow/necrotic leaves decreased from the first to third harvest, while shoot quality
increased. In the March crop, the second harvest had the lowest number of yellow/necrotic leaves and the highest shoot quality and yellow/necrotic leaves were highest and shoot quality lowest in the third harvest.
Crop season interacted with stock plant age at the three harvest dates to affect shoot fresh weight and root fresh and dry weights. For shoot fresh weight in the December crop harvests, weights increased from first to third harvest. In the March and May crop harvests, weights were higher for the second harvest and lower for the first and third harvests. For root fresh and dry weights, the December crop had highest weights in the third harvest. Cuttings from the March and May harvests had higher weights in the first two harvests, then lower in the third.