content of guajillo
5
0
0
Full text
(2)
(3)
(4)
(5)
Figure
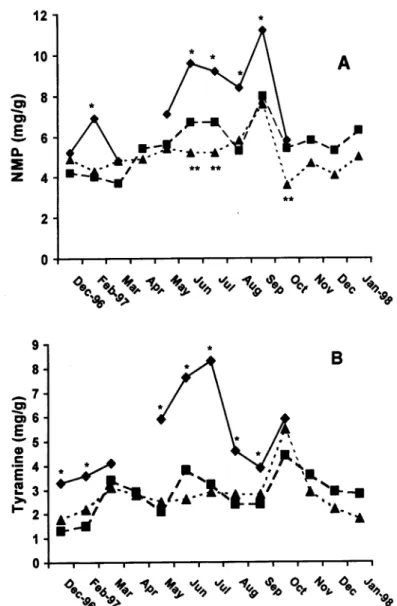
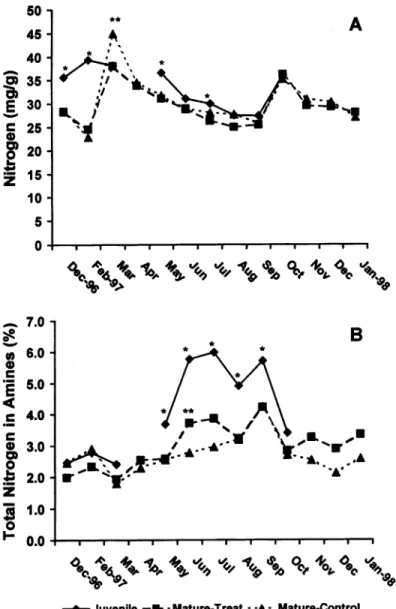
Related documents
Moreover, our study showed that the simulated acid rain significantly increased the soil organic C content and C:N ratio, lowered the hydrolysable N and the total P concen- tration,
Mean baseline fasting plasma NPY concentrations were significantly increased in BN patients compared to the controls Plasma NPY levels increased significantly after the
In vivo, in the rat mesentery, thrombin (0.5 U/ml) or N(G)-nitro-L- arginine-methyl ester (50 microM) significantly increased venular leukocyte rolling and adherence, which were
Physical solvents such as DEPG (Selexol™ or Coastal AGR ® ), NMP or N-Methyl-2- Pyrrolidone (Purisol ® ), Methanol (Rectisol ® ), and Propylene Carbonate (Fluor Solvent™) are
Compared to stagnant treatments, aerated treatments significantly increased arsenate. concentrations, but reduced arsenite concentrations in roots (p <
Fertilization did not change the N/S ratio of grain compared to the control, but the N/S ratio of straw significantly increased with increasing soil fertilizer
Concentrations of nutrients (N, P, K, Ca and Mg) and total carbohydrates, crude protein and mycorrhizal dependency of some yield parameters were significantly increased in
the terminal methyl group in fatty acid methyl esters provides similar prediction results with n -SVR and PLS; however, the n - SVR model, due to a better fit throughout the
