Dynamics of 5 carboxylcytosine during hepatic differentiation: potential general role for active demethylation by DNA repair in lineage specification
Full text
Figure
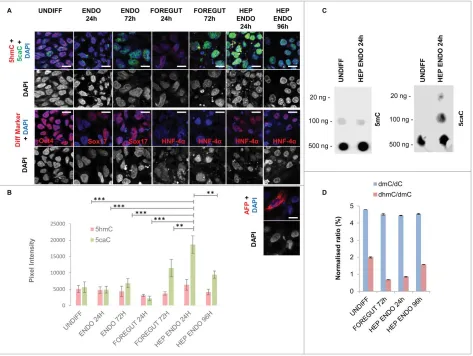
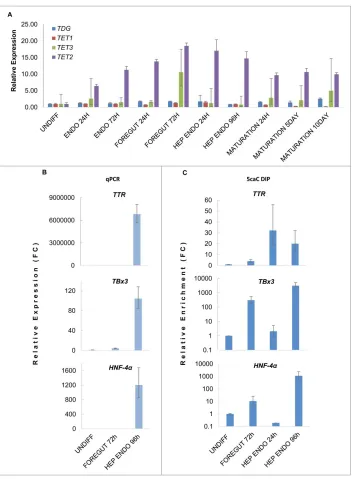
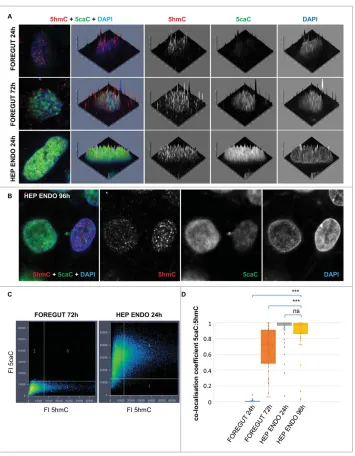
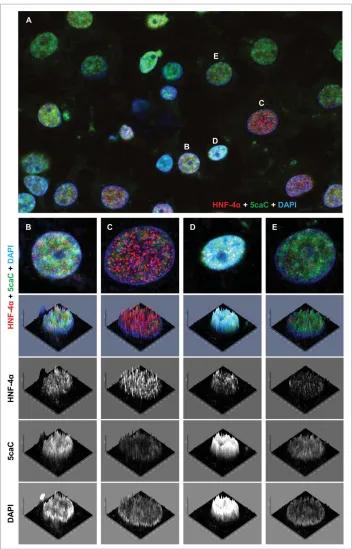
Related documents
High School Gold Region 5 Jackson Center FCCLA Gold Region 5 Miami Valley Career Technology Center 2 Gold Region 5 Newton High School Silver Region 5 Piqua
Extraction and classification of cells in histology images is an important problem that has many applications in practical medicine. The analysis of cell images can be considered
The Universal Declaration of Human Rights (UDHR, 1948), recognises, amongst other things, the right to education, the right of everyone to a standard of living adequate to the
Abbreviations: AMNR, acute macular neuroretinopathy; APMPPE, acute posterior multifocal placoid pigment epitheliopathy; CRA, chorioretinal atrophy; F, female;
From a theoretical point of view, Nicolas Marquis will benefit from a) the expertise of the scientist in charge and more globally b) of the other CERMES3 members. a)
Comparison of susceptibility testing methods with mecA gene analysis for determining oxacillin (methicillin) resistance in clinical isolates of Staphylococcus aureus and
Were Patterson to en- gage that work, quintessentially in Kant's moral and aesthetic thinking and in what we might call the issues of framing posited within
A STUDY OF CONSUMER BUYING BEHAVIOUR TOWARDS THE BRANDED APPARELS IN LUDHIANA
