Neurotrophic support by traumatized muscle-derived multipotent progenitor cells: Role of endothelial cells and Vascular Endothelial Growth Factor-A
11
0
0
Full text
Figure
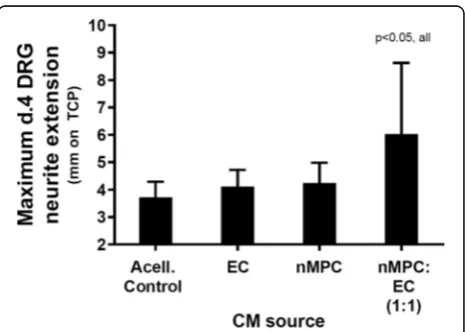
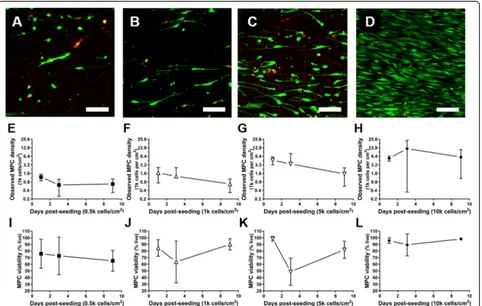
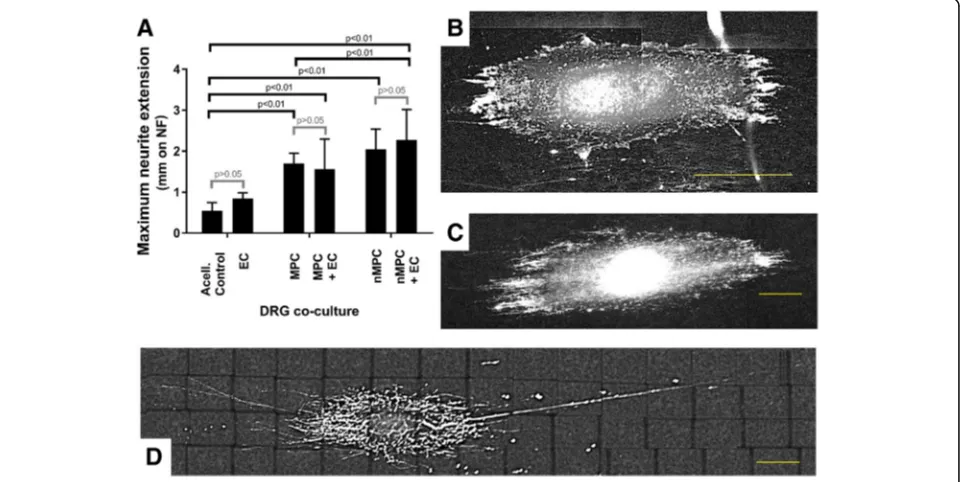
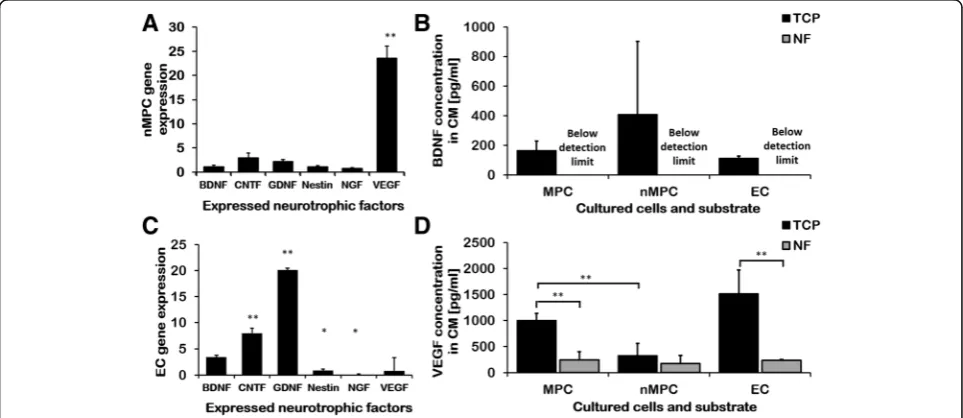
+2
Related documents