Activation of the SCAR/WAVE Complex
Andrew J. Davidson, Seiji Ura, Peter A. Thomason, Gabriela Kalna, Robert H. Insall
Beatson Institute for Cancer Research, Bearsden, Glasgow, United Kingdom
The SCAR/WAVE complex drives actin-based protrusion, cell migration, and cell separation during cytokinesis. However, the contribution of the individual complex members to the activity of the whole remains a mystery. This is primarily because com-plex members depend on one another for stability, which limits the scope for experimental manipulation. Several studies suggest that Abi, a relatively small complex member, connects signaling to SCAR/WAVE complex localization and activation through its polyproline C-terminal tail. We generated a deletion series of theDictyostelium discoideumAbi to investigate its exact role in regulation of the SCAR complex and identified a minimal fragment that would stabilize the complex. Surprisingly, loss of either the N terminus of Abi or the C-terminal polyproline tail conferred no detectable defect in complex recruitment to the leading edge or the formation of pseudopods. A fragment containing approximately 20% Abi—and none of the sites that couple to known signaling pathways—allowed the SCAR complex to function with normal localization and kinetics. However, expression of N-terminal Abi deletions exacerbated the cytokinesis defect of theDictyostelium abimutant, which was earlier shown to be caused by the inappropriate activation of SCAR. This demonstrates, unexpectedly, that Abi does not mediate the SCAR com-plex’s ability to make pseudopods, beyond its role in complex stability. Instead, we propose that Abi has a modulatory role when the SCAR complex is activated through other mechanisms.
C
ell movement derives from the extension or protrusion ofregions at the front of the cell followed by the coordinated retraction of the rear. When this behavior is coupled to the per-ception of their situation and surroundings, cells gain the ability to migrate with persistence and direction. For instance, during che-motaxis cells display an ability to detect and move along chemical gradients. Another example is during cytokinesis, where dividing cells establish opposing polarity and migrate apart from each other in a precisely coordinated manner so as to aid in the
sepa-ration of the newly formed daughter cells (1). Directed migration
requires the integrated activities of a multitude of proteins, rang-ing from sensory receptors and mediators of intracellular signal-ing to contractile actomyosin filaments, inducers of membrane curvature, and actin nucleators and their activators.
Dictyostelium discoideumhas long been the model of choice in the study of eukaryotic chemotaxis, as its movement is very com-parable to that of higher eukaryotic cells, such as neutrophils, and
it is extremely amenable to genetic manipulation (2). Such traits
makeDictyostelium discoideuma powerful model for dissecting the fundamental principles underlying chemotaxis.
InDictyostelium, as in all motile eukaryotic cells, coordinated cycles of actin polymerization and depolymerization underlie cell motility. As an actin nucleator, the Arp2/Arp3 complex induces actin polymerization that promotes the formation of cellular
pro-trusions (or pseudopodia [3, 4]). This culminates in a dense
branched network of actin capable of supporting a growing pseu-dopod. The Arp2/Arp3 complex alone has a low basal level of
activity (3). Several activators of the Arp2/Arp3 complex have now
been identified, including the WASP family of proteins. Members of the WASP family include WASP, SCAR (also named WAVE),
and the recently discovered WASH (5,6). Interactions with the
Arp2/Arp3 complex are mediated through a highly conserved C-terminal VCA domain, which is capable of promoting Arp2/Arp3 nucleation activity. The N-terminal regions of WASP family pro-teins are more varied and comprise the regulatory regions that
connect the Arp2/Arp3-activating C termini to intracellular
sig-naling events (7).
SCAR has been clearly demonstrated to be important for pseudopod extension and localizes to the extreme front of such
protrusions (8).In vivo, SCAR exists as part of a large
hetero-pentameric complex containing PIR121, Nap1, Abi, SCAR, and
HSPC300, which are collectively known as the SCAR complex (9).
A single, well-conserved homolog of each complex member is
found in Dictyostelium discoideum (10). Recently, the crystal
structure of the human SCAR/WAVE1 complex has been solved,
revealing the complex architecture (11). Sra-1 (also named
PIR121) and Nap1 form a platform upon which nestles a trimer composed of the other three complex members. The C-terminal VCA of SCAR/WAVE1 is sequestered within the complex, sug-gesting a means by which it is inhibited. However, the crystal structure yields little other information as to the purpose of the remaining bulk of the SCAR complex. Attempts to dissect the contributions of the other members to the complex as a whole have been hindered by the inherent instability of the complex to
manipulation (12,13).
As revealed by the crystal structure, SCAR/WAVE1 complex member Abi is situated at the entrance to the PIR121/Nap1 cradle wherein SCAR/WAVE1 is found. It was known that Abi interacted strongly with Nap1, and now that the crystal structure is available, it is notable that of the members of the Abi/SCAR/HSPC300
Received15 May 2013Accepted5 September 2013
Published ahead of print13 September 2013
Address correspondence to Robert H. Insall, [email protected].
Supplemental material for this article may be found athttp://dx.doi.org/10.1128 /EC.00116-13.
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/EC.00116-13
on September 8, 2020 by guest
http://ec.asm.org/
trimer, Abi is the only one to make any substantial contact with
Nap1 (14,15). The long C-terminal proline-rich tail of Abi had to
be removed to aid crystallization, presumably because of its in-trinsic disorder. However, it is most probable that it protrudes out from the main body of the complex, where it would be free to interact with other factors, including the extended C terminus of
SCAR, following its release from the complex (16).
In addition to the above-described structural considerations, it has been reported that Abi directly recruits the SCAR complex to signaling complexes containing its well-known activator, Rac1
(17). Numerous phosphorylation sites have also been identified in
Abi, and some of these have been found to be responsive to stim-uli, such as epidermal growth factor treatment or serum
starva-tion, in mammalian cells (18). Also, it has been suggested that Abi
localizes and activates the complex through its interaction with
ABL kinases, from which it originally derived its name (19).
Over-all, the literature describes Abi to be a fundamental regulator of the SCAR complex.
Previously, our lab reported the phenotype ofDictyostelium
cells lacking Abi (abiA-null cells) (20). Unlike knockouts in any of
the other complex members, in which SCAR is no longer stable
and barely detectable by Western blotting,Dictyostelium abiA
-null cells still retain appreciable levels of SCAR protein. It has been demonstrated that cells lacking Abi possess a cytokinesis defect unique to it among the null cells of the SCAR complex members.
Since cells devoid of Abi and SCAR (abiA scrAdouble-null cells)
have a phenotype equivalent to that of cells lacking SCAR (scrA
-null cells), this defect has been attributed to the residual SCAR that
is still present inabiA-null cells and its possible mislocalization or
inappropriate activity during cytokinesis.
All of the facts mentioned above strongly imply that Abi is a crucial component of the SCAR complex responsible for its local-ization and subsequent activation during chemotaxis and cytoki-nesis. For these reasons, we have undertaken the creation of a
comprehensive deletion series of theDictyosteliumAbi gene. The
resulting truncated proteins were examined for their ability to stabilize the SCAR complex and to rescue different aspects of the
abiA-null phenotype. We have thus identified the minimal
func-tional fragment of Abi required to restore SCAR complex levels
and activity inabiA-null cells.
MATERIALS AND METHODS
Cell culture.Dictyosteliumcells were cultured axenically in HL-5 medium (Formedium) at 22°C in petri dishes, and plasmid selection was main-tained with 10g/ml of G418. For growth curves, 5⫻104cells were plated
in 6-well petri dishes, with cells of an individual well harvested and a cell count performed for each 12-h time point. Alternatively, for growth in shaking culture, flasks were inoculated with 5⫻104cells/ml and a cell
count was again performed every 12 h. Cell counts were performed with a CASY cell counter and analyzer system, model TT (Innovates AG). Dou-bling times were derived, and statistical significance and two-tailedP val-ues were calculated through the use of unpairedttests.
Immunoblotting.For Western blotting, protein lysates were prepared by boiling cells in LDS buffer (Invitrogen) and were separated by electro-phoresis on NuPAGE 4 to 12% bis-Tris gels (Invitrogen). Proteins were transferred onto nitrocellulose membranes (Hybond-C-extra; Amersham Biosciences) and then blocked with a solution containing 5% nonfat dried skimmed milk dissolved in Tris-buffered saline. The membranes were then probed with antibodies specific to SCAR, PIR121, or green fluores-cent protein (GFP), followed by the use of fluorescence secondary anti-bodies that were detected using an Odyssey infrared imaging system (LI-COR Biosciences). Quantification of SCAR levels was achieved by
normalization of the SCAR band intensity to the PIR121 band intensity for each sample over four blots. For quantification, unpaired, two-tailed Studentttests were used to determine statistical significance.
For native PAGE, lysates were separated by electrophoresis on Native-PAGE Novex bis-Tris 4 to 16% gels (Invitrogen) according to the manu-facturer’s protocol. Proteins were transferred onto polyvinylidene difluo-ride (PVDF) membranes, blocked, probed with anti-SCAR antibody, and detected by chemiluminescence (Hybond-P PVDF membrane [Amer-sham], horseradish peroxidase [HRP]-conjugated secondary antibodies and Immobilon Western chemiluminescent HRP substrate [Millipore]).
TIRF microscopy.The HL-5 medium from confluent plates was re-placed with development buffer (10 mM KNaPO4, 2 mM MgCl2, 1 mM
CaCl2, pH 6.5), and cells were starved for 7 h. The cells were then
har-vested from the plates, seeded in glass-bottom dishes (Iwaki), and then slightly compressed under 0.4% agarose slabs. SCAR complex localization and cell morphology were imaged simultaneously using total internal fluo-rescence (TIRF) microscopy and differential interference contrast (DIC) microscopy, respectively, on a modified Nikon Eclipse TE 2000-U micro-scope using a⫻100 Nikon TIRF objective (numerical aperture [NA], 1.45). Images were recorded every 2 s. Pseudopodia were counted manu-ally with the aid of ImageJ software (National Institutes of Health). Pseu-dopodia were defined as large convex, semicircular protrusions devoid of granular cytosol. This definition excluded all but the largest runs of blebs. HSPC300-GFP recruitment to these protrusions was then retrospectively determined. Ten cells (collectively amounting to an hour of cell migra-tion) were analyzed for each of the transformants, and statistical signifi-cance and two-tailedPvalues were calculated through the use of unpaired Studentttests.
DAPI-phalloidin staining.Doubling times were derived from the growth curves prepared as described above, and cells were seeded at a low density on glass coverslips in a staggered fashion so that the transformants underwent approximately 10 divisions. The coverslips were removed, and cells were then fixed for 5 min with a solution containing 6% (wt/vol) formaldehyde, 15% (vol/vol) picric acid, and 10 mM PIPES [piperazine-N,N=-bis(2-ethanesulfonic acid)] adjusted to pH 6.5. Cells were then washed with phosphate-buffered saline (PBS) before permeabilization with 70% ethanol for 2 min. The fixed cells were washed with PBS and then stained with 33 nM Texas Red phalloidin (Invitrogen) for 30 min. Coverslips were washed again in PBS and finally mounted on glass slides with antifade reagent containing DAPI (4=,6-diamidino-2-phenylindole; Prolong Gold; Invitrogen). Cells were imaged using an inverted wide-field microscope (IX81; Olympus) and a⫻60 objective (NA, 1.42) or a Nikon A1 confocal microscope with a⫻60 objective (NA, 1.4). The number of nuclei per cell was counted for⬎100 cells for each of the transformants, and the cells were then grouped into those with either 1 orⱖ2 nuclei. The chi-square test was used for investigation of the differences between the observed frequencies of nuclei per cell. When the expected counts fell below 5, Yates’ correction was applied.
RESULTS
Only a small fragment of Abi is required to stabilize the SCAR complex.From the crystal structure available from the Protein
Data Bank (accession number3P8C;Fig. 1a), it is apparent that
Abi2 is incorporated into the mammalian SCAR/WAVE1
com-plex through the second of its two N-terminal alpha helices (␣2;
Fig. 1a). This domain was identified in theDictyosteliumAbi by
protein alignment (Fig. 1b). We reasoned that this alpha helix
alone could be sufficient to fully stabilize the complex in
Dictyo-stelium abiA-null cells, and so we proceeded to incrementally
de-lete all peripheral sequence on either side of it (Fig. 1c). Deletion of
the N terminus of Abi resulted in the loss of the anti-Abi epitope, which confounded detection (see Fig. S1 in the supplemental ma-terial). We were reluctant to directly tag Abi because the different relative positions of GFP could interfere differentially with Abi’s
on September 8, 2020 by guest
http://ec.asm.org/
normal activity. However, in order to compare the expression of all our constructs, we additionally N-terminally fused our deletion series to GFP, allowing each member of the series to be detected
with anti-GFP.Figure 2a demonstrates that all of the Abi
con-structs were expressed and there were no gross differences in pro-tein levels. Unless otherwise stated, in all subsequent work we coexpressed our untagged deletion series with HSPC300-GFP, an
established marker of the SCAR complex (8). The untagged,
trun-cated Abi proteins were then tested for their ability to stabilize the
SCAR complexin vivowhen expressed in theabiA-null cells. The
SCAR protein level, as assayed by Western blotting, was initially
utilized as a readout of complex integrity (Fig. 2a). We have
pre-viously shown that SCAR complex member PIR121 is stable in
abiA- andscrA-null cells, and so we utilized it as a loading control
(12). The domains of Abi that proved dispensable for SCAR
sta-bilization included the first alpha helix (N-terminally truncated
Abi [⌬Nt Abi]) and—as has been shown previously in
mamma-lian cells (14,21)—the entire C-terminal polyproline tail
(C-ter-FIG 1Design and implementation of Abi deletion series. (a) Illustration of human SCAR/WAVE1 complex derived from the recently resolved crystal structure available from the Protein Data Bank (accession number3P8C). The sequence corresponding to that deleted in theDictyosteliumAbi is highlighted in red. Red ribbon, the Abi2 polyproline region that is absent from the crystal structure (not to scale). Labels designate different domains/features of Abi2:␣1, 1st alpha helix; ␣2, 2nd alpha helix; CL, conserved loop. (b) Protein alignment between human Abi2 (above), represented in panel a, andDictyosteliumAbi (below) with a color scheme identical to that described for panel a. The sequence corresponding to the C-terminal SH3 domain of human Abi2 was removed to aid alignment, as DictyosteliumAbi does not possess this domain. Asterisks, amino acid identities; underlined sequence, amino acids corresponding to the 2nd alpha helix in panel a. (c) Diagrammatic representation of theDictyosteliumAbi domain structure and the subsequent deletion series undertaken. WT, full-length Abi;⌬Nt, N-terminally truncated Abi;⌬Ct1and⌬Ct2, C-terminally truncated Abi as shown;⌬Abi⌬, combination of both N-terminally and C-terminally truncated Abi.
on September 8, 2020 by guest
http://ec.asm.org/
minally truncated Abi1 [⌬Ct1Abi]). The most distal C-terminal
Abi2 sequence for which there are crystal structure data available (i.e., immediately before the start of the polyproline tail) was
strik-ingly conserved (Fig. 1b). Following its deletion in combination
with the polyproline tail (⌬Ct2Abi), it, too, surprisingly, proved
unnecessary for SCAR stabilization (Fig. 2a). Further truncation
of the C terminus of Abi yielded fragments that were expressed but did not contribute to SCAR stability (data not shown). Finally, a minimal Abi fragment which had deletions at both termini and
consisted of little more than the␣2 helix (Abi with a combination
of both N-terminal and C-terminal truncations [⌬Abi⌬])
re-stored SCAR protein levels in abiA-null cells, if only partially.
Quantification of SCAR levels in these cells demonstrated that
⌬Abi⌬restored SCAR significantly less than wild-type (WT) Abi
(unpairedttest,P⬍0.05) but restored it to a level significantly
more than the basal level found in the null strain expressing
HSPC300-GFP alone (unpairedttest,P⬍0.001;Fig. 2b). The Abi
constructs tagged with GFP also restored SCAR levels inabiA-null
cells to a comparable extent (Fig. 2a).
The stabilization of the intact complex was confirmed by native
gel electrophoresis, as demonstrated inFig. 2c. Under these
con-ditionsDictyosteliumSCAR was found in only one
high-molecu-lar-weight complex, the presence of which depended on Abi. Both PIR121 and the GFP-tagged Abi deletion series comigrated with this high-molecular-weight complex, confirming that it was the
intact SCAR complex (data not shown; see Fig. S2 in the supple-mental material). Neither monomeric nor complexed SCAR was
detected inabiA-null strains in native PAGE, which is
unsurpris-ing, considering the low sensitivity of the method and the small
amount of residual SCAR that is present whenabiA is deleted.
Also of note was the significant band shift in the intact complexes that had lost the C-terminal polyproline domain of Abi. Since Abi does not contribute a large amount to the total mass of the SCAR complex, it is likely that truncation of the disordered polyproline tail allows more efficient passage of the complex through the gel during electrophoresis.
In summary, we were successfully able to delete 239 of the 332
total amino acids comprisingDictyostelium Abi and still retain
SCAR complex integrity. For the first time, we were in a position to interrogate the contribution of an individual complex member to the activity of the whole. With this in mind, we sought to de-termine if our Abi constructs restored every aspect of the
wild-type SCAR complex inabiA-null cells with the aim of identifying
functions that could be specifically attributed to Abi.
Abi deletion series rescues the growth defect ofabiA-null cells in proportion to SCAR stabilization.The SCAR complex is
required for the optimal growth ofDictyostelium discoideumin
liquid medium, presumably due to its involvement in
macropi-nocytosis (22). Therefore, having restored the SCAR protein to
wild-type levels, we next sought to determine if the truncated Abi
FIG 2Identification of minimal Abi fragment. (a) Western blot revealing stabilization of SCAR by the untagged/tagged Abi deletion series transformed into an abiA-null strain. (Top) Lysates probed with antibody against PIR121, a stable member of the complex that was used as a loading control; (second panel) lysates probed with anti-SCAR antibody demonstrating stabilization of SCAR by different untagged Abi fragments; (third panel) expression of truncated GFP-tagged Abi constructs confirmed by anti-GFP Western blotting; (bottom) lysates probed with anti-SCAR antibody demonstrating stabilization of SCAR by different GFP-tagged Abi fragments. The dashed line divides blots into those derived from cells that possessed untagged Abi deletion series (above the dashed line) and those that possessed GFP-tagged Abi deletion series (below the dashed line). (b) Quantification of SCAR levels was achieved by normalization of the SCAR band intensity to the PIR121 band intensity for each untagged transformant over four blots. *, significantly different from WT Abi SCAR levels (unpairedttest,P⬍ 0.05); **, significantly different from⌬Abi⌬SCAR levels (unpairedttest,P⬍0.0001). Error bars indicate SEMs. (c) Native PAGE demonstrating the stabilization of the entire SCAR complex by the untagged Abi fragments. Intact protein complexes were separated by native PAGE and probed with anti-SCAR antibody following Western blotting. WT, full-length wild-type Abi;⌬Nt, N-terminally truncated Abi;⌬Ct1and⌬Ct2, C-terminally truncated Abi as shown inFig. 1c; ⌬Abi⌬, combination of both N-terminally and C-terminally truncated Abi; Null, HSPC300-GFP-only vector/empty GFP vector.
on September 8, 2020 by guest
http://ec.asm.org/
constructs were also capable of rescuing the growth impairment of
these cells. As can be seen fromFig. 3a,⌬Nt Abi,⌬Ct1Abi, and
⌬Ct2Abi all restored growth on substrate to a rate comparable to
that ofabiA-null cells rescued with full-length Abi.⌬Abi⌬did not
significantly improve the growth ofabiA-null cells in petri dishes.
However, partial rescue was difficult to observe under these
con-ditions, as the defect is relatively small. As shown in Fig. 3b,
growth in suspension exacerbated the growth defect of theabiA
-null cells and revealed an intermediate phenotype of cells rescued
with⌬Abi⌬. Under these conditions, cells rescued with ⌬Abi⌬
doubled significantly more slowly than cells rescued with WT Abi
(by 13.9⫾3.9 h; unpairedttest,P⬍0.05). The severe growth
defect of theabiA-null cells in suspension led to uneven growth
and made it impractical to calculate a doubling time and perform statistical comparisons. However, it is clear that transformation
with⌬Abi⌬consistently improved the growth ofabiA-null cells in
suspension.
We have thus demonstrated that loss of either terminus of Abi has no detrimental effect on cell growth. Furthermore, since
⌬Abi⌬ partially restored SCAR levels and partially restored
growth, the extent to which the growth defect of theabiA-null cells
was rescued appeared to simply correlate with the SCAR protein levels in these cells. We infer that Abi contributes little to cell growth beyond stabilization of the SCAR complex. This result implies that even when lacking a large proportion of the Abi se-quence, the SCAR complex still retains sufficient functionality to support optimal growth and so argues against a central role for Abi in complex recruitment and activation.
This prompted us to investigate the subcellular localization and dynamics of our mutant SCAR complexes, with particular interest in their ability to drive pseudopod extension.
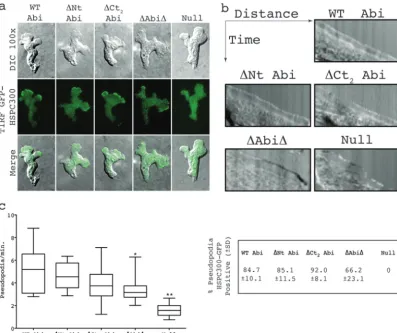
SCAR complex containing truncated Abi localizes normally in migrating cells.Coexpression of GFP-tagged complex member HSPC300 alongside the Abi fragments identified as described
above in anabiA-null background allowed the localization of the
respective mutant complexes to be determined. When visualized
by TIRF microscopy, which illuminates only the bottom⬃100 nm
of a cell, HSPC300-GFP was seen to localize strongly to the leading edge of migrating wild-type cells, correlating with pseudopod ex-tension. No obvious differences in SCAR complex dynamics were observed between any of the Abi fragments and full-length Abi, which all showed robust concentrations of the complex at the
leading edge of randomly migrating cells (Fig. 4a; see Movie S1 in
the supplemental material). This was accompanied by the smooth extension of cellular protrusions, as illustrated by the kymographs inFig. 4b. The HSPC300-GFP recruitment was weaker in cells
expressing⌬Abi⌬, and they were also more prone to outbreaks of
blebbing. Both observations are consistent with there being
re-duced levels of SCAR in these cells (23). However, SCAR
complex-driven pseudopodia were also clearly evident, although the rate of
protrusion was seen to be significantly decreased (unpairedttest,
P⬍0.05) compared to that of WT Abi-rescued cells (Fig. 4bandc;
see Movie S1 in the supplemental material). HSPC300-GFP was
essentially completely delocalized inabiA-null cells transformed
with HSPC300-GFP alone. These cells almost exclusively moved
by means of blebbing (Fig. 4b) and rarely extended anything that
could be classified as a pseudopod. This was reflected in a
signifi-cant reduction in the protrusion rate of⬃30% compared to that
of WT Abi-expressing cells (unpairedttest,P⬍0.001;Fig. 4b).
The few pseudopodia that were generated were devoid of
concen-trated HSPC300-GFP (Fig. 4c) and were likely driven by WASP, as
we have previously shown in SCAR complex-deficient cells,
in-cludingabiA-null cells (8). All of this implies that not only is the
majority of the Abi sequence unnecessary for SCAR complex sta-bilization but also it is not essential for normal complex localiza-tion. Furthermore, since all the transformants were generating the normal diverse range of pseudopodia, which are suppressed in the
abiA-null cells, it would appear that the majority of Abi is not
required for SCAR complex activation or function either.
Deletion of the␣1 helix alone confers a multinucleate phe-notype.Our lab has previously shown thatDictyostelium abiA -null cells have a tendency to become multinucleate due to a failure
in cytokinesis (20). Many Dictyostelium mutants accumulate
multinucleate cells during growth in shaking culture but can re-solve such cells when introduced to a substrate by myosin-inde-pendent cytokinesis or by the mitosis-indemyosin-inde-pendent process of
traction-mediated cytofission (24). However, even in the presence
of a substrate,abiA-null cells often become multinucleate. The
abiAconstructs generated here were tested to determine which
deletions could rescue this phenotype. Doubling times were
de-FIG 3Truncated Abi proteins rescue the growth defect ofabiA-null cells. (a, b) Growth curves corresponding to transformants cultured on a petri dish (a) or in shaking suspension (b). A cell count was performed every 12 h, and the fold increase was calculated and plotted. Error bars indicate SEMs. WT, full-length Abi;⌬Nt, N-terminally truncated Abi;⌬Ct1and⌬Ct2, C-terminally truncated Abi as shown inFig. 1c;⌬Abi⌬, combination of both N-terminally and C-terminally truncated Abi; Null, HSPC300-GFP-only vector.
on September 8, 2020 by guest
http://ec.asm.org/
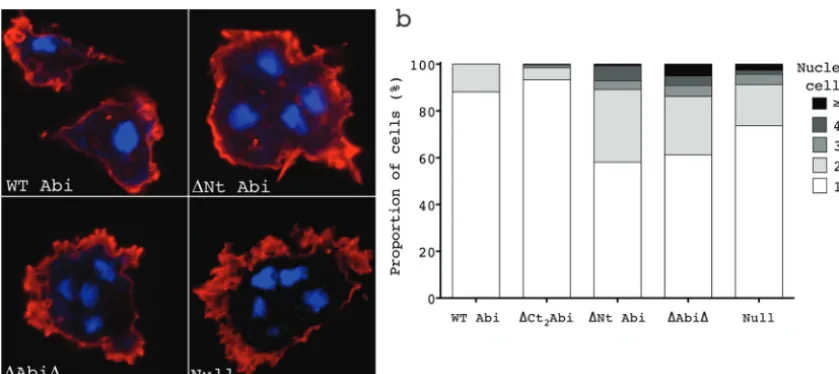
rived from the growth curves shown inFig. 3a. Cells were plated at a low density and cultured for 10 divisions. Following this, the cells were then fixed and stained with DAPI to visualize the nuclei and with Texas Red phalloidin for actin so as to clearly define the cell
boundaries (Fig. 5a). The number of nuclei per cell was then
counted, and the proportion of multinucleate cells for each of the
transformants is shown inFig. 5b. Cells withⱖ2 nuclei were rarely
seen inabiA-null cells rescued with full-length Abi or Abi lacking
the C-terminal polyproline tail. The vast majority of these cells were mononucleate, with only a small population of cells
hav-ingⱖ2 nuclei (12 and 7% of the total, respectively). As has been
previously documented, the abiA-null cells expressing only
HSPC300-GFP accumulated multinucleate cells, with a
signifi-cant increase in the number of cells withⱖ2 nuclei compared to
that for the WT Abi controls (26%; chi-squared test,P⬍0.0001)
being detected. Furthermore, there were also more severely multi-nucleate cells present, including some with as many as 10 nuclei
per cell (Fig. 5b). Surprisingly, deletion of the N-terminal␣1 helix
resulted in a significant increase in the number of cells withⱖ2
nuclei (42%) even compared to the number ofabiA-null cells
(chi-square test,P⬍0.0001). This was also true of⌬Abi⌬
-ex-pressing cells (39% of cells withⱖ2 nuclei). Again, this proved to
be a significant increase compared to the results for both the WT
Abi controls and theabiA-null cells (chi-square test,P⬍0.0001
andP⬍0.01, respectively), although there was no statistically
significant difference compared to the results for the⌬Nt
Abi-expressing cells (chi-square test,P⫽0.36). The different severity
in the phenotype observed between theabiA-null cells and the
⌬Abi⌬- and⌬Nt Abi-expressing cells correlates with the
increas-ing SCAR levels in these cells (Fig. 2aandb) and, therefore,
pre-sumably, an increasing amount of deregulated SCAR complex. These data are consistent with our lab’s previous conclusion
that it is the remaining SCAR in theabiA-null cells that is
respon-sible for its multinuclearity. The data presented here identify the
␣1 helix of Abi as being important for the regulation of the SCAR
complex during cytokinesis.
DISCUSSION
Abi is considered a key component of the SCAR complex that is thought to control its activation by acting as a target for
phosphor-FIG 4SCAR complexes containing truncated Abi proteins localize normally in migrating cells. (a) Localization of HSPC300-GFP inabiA-null cells expressing different Abi fragments. (Top row) DIC images of migrating cells coexpressing Abi-truncated constructs and the SCAR complex marker HSPC300-GFP; (middle row) TIRF images of the same cells revealing localization of mutant SCAR complexes; (bottom row) merged images of the upper two rows. (b) Representative kymographs highlighting smooth progression of advancing protrusions in cells expressing Abi truncations. Null cells present with a ragged slope indicative of blebbing. (c) All truncated Abi proteins rescue the suppressed pseudopod formation ofabiA-null cells. *, significantly reduced rate of pseudopod formation compared to that for WT Abi-rescued cells (unpairedttest,P⬍0.05); **, significantly reduced rate of pseudopod formation compared to that for⌬Abi⌬-rescued cells (unpairedttest,P⬍0.001). The percentages in the box on the right indicate the proportion of pseudopodia with robust HSPC300-GFP localization⫾SD. WT Abi, full-length Abi;⌬Nt, N-terminally truncated Abi;⌬Ct2, C-terminally truncated Abi;⌬Abi⌬, combination of both N-terminally and C-terminally truncated Abi; Null, HSPC300-GFP-only vector.
on September 8, 2020 by guest
http://ec.asm.org/
ylation and through its interaction with various regulatory pro-teins, such as receptor tyrosine kinases. However, here we deleted the majority of the Abi sequence, including all the conserved phosphorylation sites identified, and still observed robust local-ization of the SCAR complex at the leading edge of migrating cells. The loss of Abi in its entirety resulted in a loss of complex integrity, and so we cannot rule out the possibility that the remaining alpha helix required for complex stability is involved in activation. How-ever, structural considerations suggest that this is unlikely, as the minimal Abi fragment that still stabilized the complex consisted of little more than a single alpha helix that incorporated Abi into the complex. Furthermore, on the basis of the model that emerged from the resolution of the crystal structure, this helix would be oriented away from the plasma membrane, where activation is presumed to take place. Most surprisingly, deletion of the entire C-terminal polyproline tail of Abi, which accounts for almost half of the entire sequence, yielded no detectable phenotype. Instead, it was deletion of the first 44 amino acids of Abi that was sufficient to reproduce and apparently exacerbate the multinucleate defect of
theabiA-null cells.
Dictyosteliumpossesses multiple robust mechanisms to ensure successful cell division. These include the substrate-independent action of myosin at the cleavage furrow of a dividing cell, which acts to pinch the cell in two. This is coupled to adhesion-depen-dent directed migration, which acts to drive the newly forming daughter cells apart. These two modes of cytokinesis almost cer-tainly work together in concert to ensure efficient cell division; however, they are also sufficient to compensate in the absence of one or the other. As a final safeguard, multinucleate cells on a substrate can randomly tear themselves apart independently of mitosis by traction-mediated cytofission. We have previously shown that these two substrate-dependent means of cell division
require the SCAR complex (1). Thescar-null cells themselves have
a low level of multinuclearity, as myosin is sufficient to drive
effi-cient cytokinesis. The fact that theabiA-null cells become
multi-nucleate has been attributed to a misregulation or localization of
their residual SCAR protein, with the resulting aberrant activity disrupting cell division. Despite restoring SCAR to wild-type lev-els, expression of the N-terminally deleted Abi merely aggravated the cytokinesis defect, supporting our previous conclusions. It is notable that the multinucleate phenotype documented here is not strong compared with the phenotype derived from mutations that specifically block cytokinesis. Given that the cells were cultured for 10 divisions, the vast majority of even the most severely affected
⌬Nt Abi- or⌬Abi⌬-expressing cells were no more than
binucle-ate. This is well below what the particularly robustDictyostelium
can tolerate and possibly explains why⌬Nt Abi confers no
signif-icant adverse effect on growth. By stabilizing the SCAR complex with our N-terminally truncated Abi proteins, we were also likely restoring traction-mediated cytofission in these cells, which works to counteract any specific defect in cytokinesis.
From all this we conclude that Abi is not required for recruit-ment or activation of the SCAR complex even during cell migra-tion and cytokinesis. Instead, we suggest here that Abi’s role is to modulate the activity of the SCAR complex and act as a signal integrator. During events such as cytokinesis, where the SCAR complex appears to be hyperactivated, the small imbalance in
ac-tivity conferred by the absence of the␣1 helix of Abi would have
the greatest effect and could possibly accumulate to ultimately
contest and disrupt normal cell division (1). As to how exactly the
␣1 helix of Abi is regulating SCAR complex activity and what it is
interacting with remain unknown. Nevertheless, this work has identified one functional domain on what still remains a mostly impenetrable complex.
There exists strong conservation between the Dictyostelium
SCAR complex members and those found in higher eukaryotes, in terms of both sequence and function. However, in higher eu-karyotes, Abi is evidently more complicated, possessing an SH3
domain and interactors not present inDictyostelium. These
pre-sumably accommodate the increasingly complex layers of regula-tion required to control SCAR activity in multicellular organisms. As a modulator of SCAR complex activity, Abi would appear to be
FIG 5Deletion of the 1st alpha helix is sufficient to reproduce and to exacerbate the multinucleate phenotype ofabiA-null cells. Cells were cultured for the time taken for 10 divisions on glass coverslips before fixation and stained with Texas Red phalloidin and DAPI. The number and extent of multinucleate cells were then counted. (a) Examples of normal and multinucleate cells found inabiA-null cells expressing WT Abi,⌬Nt Abi,⌬Abi⌬, and the HSPC300-GFP vector only (Null). (b) Percentages of cells with 1, 2, 3, 4, orⱖ5 nuclei cells rescued with the different Abi constructs. WT Abi, full-length Abi;⌬Nt, N-terminally truncated Abi;⌬Ct2, C-terminally truncated Abi;⌬Abi⌬, combination of both N-terminally and C-terminally truncated Abi; Null, HSPC300-GFP-only vector.
on September 8, 2020 by guest
http://ec.asm.org/
a natural target through which to exert these additional layers of regulation. This observation, together with the findings presented in this paper, lead us to propose that Abi’s role in all motile cells is as a modulator. Abi is unlikely to directly mediate either SCAR complex recruitment to the membrane or activation in any cell type, including cells of higher animals. Instead, it most likely tunes the activity of the SCAR complex during or after activation.
ACKNOWLEDGMENTS
We are thankful for the funding provided by Cancer Research UK. We also thank Margaret O’Prey of the Beatson Advanced Imaging Resource for technical help with the microscopy and Douwe Veltman for providing various plasmids. Finally, we voice our appreciation for the Dictyosteliumcommunity resource,dictybase.org.
REFERENCES
1.King JS, Veltman DM, Georgiou M, Baum B, Insall RH.2010. SCAR/ WAVE is activated at mitosis and drives myosin-independent cytokinesis. J. Cell Sci.123:2246 –2255.
2.Andrew N, Insall RH.2007. Chemotaxis in shallow gradients is mediated independently of PtdIns 3-kinase by biased choices between random pro-trusions. Nat. Cell Biol.9:193–200.
3.Mullins RD, Heuser JA, Pollard TD.1998. The interaction of Arp2/3 complex with actin: nucleation, high affinity pointed end capping, and formation of branching networks of filaments. Proc. Natl. Acad. Sci. U. S. A.95:6181– 6186.
4.Svitkina TM, Borisy GG.1999. Arp2/3 complex and actin depolymeriz-ing factor/cofilin in dendritic organization and treadmilldepolymeriz-ing of actin fila-ment array in lamellipodia. J. Cell Biol.145:1009 –1026.
5.Machesky LM, Insall RH.1998. Scar1 and the related Wiskott-Aldrich syndrome protein, WASP, regulate the actin cytoskeleton through the Arp2/3 complex. Curr. Biol.8:1347–1356.
6.Linardopoulou EV, Parghi SS, Friedman C, Osborn GE, Parkhurst SM, Trask BJ.2007. Human subtelomeric WASH genes encode a new subclass of the WASP family. PLoS Genet. 3:e237. doi:10.1371/journal.pgen .0030237.
7.Pollitt AY, Insall RH.2009. WASP and SCAR/WAVE proteins: the driv-ers of actin assembly. J. Cell Sci.122:2575–2578.
8.Veltman DM, King JS, Machesky LM, Insall RH.2012. SCAR knockouts in Dictyostelium: WASP assumes SCAR’s position and upstream regula-tors in pseudopods. J. Cell Biol.198:501–508.
9.Eden S, Rohatgi R, Podtelejnikov AV, Mann M, Kirschner MW.2002. Mechanism of regulation of WAVE1-induced actin nucleation by Rac1 and Nck. Nature418:790 –793.
10. Caracino D, Jones C, Compton M, Saxe CL, III.2007. The N-terminus of Dictyostelium Scar interacts with Abi and HSPC300 and is essential for proper regulation and function. Mol. Biol. Cell18:1609 –1620.
11. Chen Z, Borek D, Padrick SB, Gomez TS, Metlagel Z, Ismail AM, Umetani J, Billadeau DD, Otwinowski Z, Rosen MK.2010. Structure and control of the actin regulatory WAVE complex. Nature468:533–538. 12. Ibarra N, Blagg SL, Vazquez F, Insall RH.2006. Nap1 regulates Dictyo-stelium cell motility and adhesion through SCAR-dependent and -inde-pendent pathways. Curr. Biol.16:717–722.
13. Kunda P, Craig G, Dominguez V, Baum B.2003. Abi, Sra1, and Kette control the stability and localization of SCAR/WAVE to regulate the for-mation of actin-based protrusions. Curr. Biol.13:1867–1875.
14. Innocenti M, Zucconi A, Disanza A, Frittoli E, Areces LB, Steffen A, Stradal TE, Di Fiore PP, Carlier MF, Scita G.2004. Abi1 is essential for the formation and activation of a WAVE2 signalling complex. Nat. Cell Biol.6:319 –327.
15. Gautreau A, Ho HY, Li J, Steen H, Gygi SP, Kirschner MW. 2004. Purification and architecture of the ubiquitous Wave complex. Proc. Natl. Acad. Sci. U. S. A.101:4379 – 4383.
16. Davidson AJ, Insall RH.2011. Actin-based motility: WAVE regulatory complex structure reopens old SCARs. Curr. Biol.21:R66 –R68. doi:10 .1016/j.cub.2010.12.001.
17. Innocenti M, Frittoli E, Ponzanelli I, Falck JR, Brachmann SM, Di Fiore PP, Scita G.2003. Phosphoinositide 3-kinase activates Rac by entering in a complex with Eps8, Abi1, and Sos-1. J. Cell Biol.160:17–23.
18. Lebensohn AM, Kirschner MW.2009. Activation of the WAVE complex by coincident signals controls actin assembly. Mol. Cell36:512–524. 19. Leng Y, Zhang J, Badour K, Arpaia E, Freeman S, Cheung P, Siu M,
Siminovitch K.2005. Abelson-interactor-1 promotes WAVE2 membrane translocation and Abelson-mediated tyrosine phosphorylation required for WAVE2 activation. Proc. Natl. Acad. Sci. U. S. A.102:1098 –1103. 20. Pollitt AY, Insall RH.2008. Abi mutants in Dictyostelium reveal specific
roles for the SCAR/WAVE complex in cytokinesis. Curr. Biol.18:203–210. 21. Stradal T, Courtney KD, Rottner K, Hahne P, Small JV, Pendergast AM.2001. The Abl interactor proteins localize to sites of actin polymer-ization at the tips of lamellipodia and filopodia. Curr. Biol.11:891– 895. 22. Seastone DJ, Harris E, Temesvari LA, Bear JE, Saxe CL, Cardelli J.2001.
The WASp-like protein scar regulates macropinocytosis, phagocytosis and endosomal membrane flow in Dictyostelium. J. Cell Sci.114:2673–2683. 23. Ura S, Pollitt AY, Veltman DM, Morrice NA, Machesky LM, Insall RH. 2012. Pseudopod growth and evolution during cell movement is con-trolled through SCAR/WAVE dephosphorylation. Curr. Biol.22:553–561. 24. Nagasaki A, de Hostos EL, Uyeda TQ.2002. Genetic and morphological evidence for two parallel pathways of cell-cycle-coupled cytokinesis in Dictyostelium. J. Cell Sci.115:2241–2251.