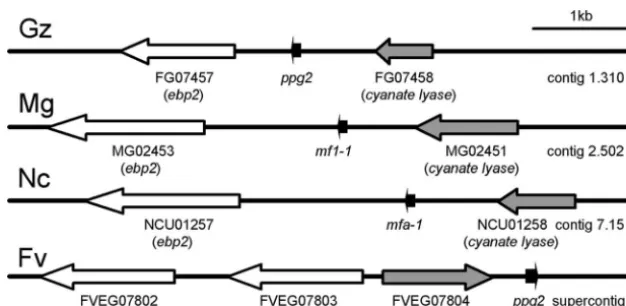
Expression and Function of Sex Pheromones and Receptors in the Homothallic Ascomycete Gibberella zeae
Full text
Figure




Related documents
The total coliform count from this study range between 25cfu/100ml in Joju and too numerous to count (TNTC) in Oju-Ore, Sango, Okede and Ijamido HH water samples as
So, the aim of the present was to screen this endemic plant extracts has been earlier to have antimicrobial activity against the pathogens causing
(A) Type I IFN production by SV40T MEFs from matched wild-type and Sting -deficient mice stimulated with 0.1 M CPT for 48 h, measured by bioassay on LL171 cells (data shown as
6 4 The plaintiff elected to recover damages for bad faith practices prescribed under the Texas Insurance Code in addition to attorneys' fees under the DTPA.1 65 The
However, much research has shown that the develop- ment of sound clinical reasoning skills and decision- making abilities is inextricably linked to experience. More
Who came first larvae or adults? Origins of bilaterian metazoan larvae BELINDA J SLY, MARGARET S SNOKE and RUDOLF A RAFF* Department of Biology, Indiana University, Bloomington, Indiana,
This R 2 value indicates that the weighted quadratic spline model with two point of knot is an appropriate model for describe the relationship between age
Sialolithiasis refers to the formation of calcareous concentration or sialoliths in the salivary duct leading to obstruction of the salivary flow, resulting in salivary ectasia,
