http://dx.doi.org/10.4236/jbm.2015.33004
How to cite this paper:
Abdel-Hameed, U.K., Tantawy, M.E., Salim, M.A., Mourad, M.M. and Ishak, I.F. (2015) Phenetic
Analysis of Morphological and Molecular Traits in Acanthaceae Juss. Journal of Biosciences and Medicines, 3, 18-34.
http://dx.doi.org/10.4236/jbm.2015.33004
Phenetic Analysis of Morphological and
Molecular Traits in Acanthaceae Juss
Usama K. Abdel-Hameed
*, Mohamed E. Tantawy, Mohamed A. Salim, Magdy M. Mourad,
Ishak F. Ishak
Botany Department, Faculty of Science, Ain Shams University, Cairo, Egypt
Email:
*[email protected]
Received November 2014
Abstract
Acanthaceae has received considerable taxonomic attention at the familial, subfamilial, tribal and
subtribal levels. Several different infra-familial classifications have been proposed for the
Acan-thaceae, but no taxonomic consensus has yet been reached. The main objective of the present
study is to throw light on the phenetic relationships and to explore the contribution of
morpho-logical and molecular characters in systematics of Acanthaceae. The morphomorpho-logical data viz.
ma-cromorphology, stomatography, lamina architecture and ISSR profiles of 30 Egyptian
acantha-ceous taxa were investigated. The phenetic analysis using NTSYS-PC version 2.02 software based
on 55 potentially informative morphological and molecular characters indicated that the used
morphological and ISSR criteria is likely to be useful and valuable taxonomic traits. The
morpho-logical characters and ISSR aspects of all the studied species produced a phenogram that showed
two series; one of them had two subseries, the first one comprised only three taxa while the
second divided into two clusters, each contained two groups. The delimitation and the
member-ship of the studied taxa clearly merit additional study using more criteria. The phenetic analysis of
both morphological and molecular attributes clarified the segregation of genus Avicennia as a
dis-tinct identity away from Acanthaceae. Acanthus mollis & A. montanus are isolated in its own series
that comparable to tribe Acantheae of the current taxonomic systems. The studied species of
Thunbergia are gathered its own subseries that comparable to tribe Thunbergiae and Ruellia in its
own group that comparable to tribe Ruellieae.
Keywords
Phenetic Analysis, Numerical Taxonomy, Morphology, ISSR, Acanthaceae
1. Introduction
Acanthaceae has received considerable taxonomic attention at the familial, subfamilial, tribal and subtribal
le-vels by various workers, the earlier system of infra-familial classifications of Acanthaceae is that of [1], he
sub-divided the family on the basis of two to four stamens and recognized either genera viz.
Acanthus
,
Barleria
,
Blepharis
,
Dianthera
,
Dilvaria
,
Justicia
,
Ruellia
and
Thunbergia
. Since then, the family has been subjected to
various taxonomic treatments and has received attention from many authors; [2] recognized four subfamilies viz.
Nelsonioideae, Mendoncioideae, Thunbergioideae and Acanthoideae on the basis of types of fruits, number of
ovules and presence or absence of retinacula and their shape. He further subdivides Acanthoideae into two
sec-tions (Contortae and Imbricatae).
Several different infra-familial classifications have been proposed for the Acanthaceae, but no taxonomic
consensus has yet been reached [3]. On the basis of morphology it has been suggested that the family is not
“natural” [4] [5]. Molecular data has helped botanists move towards a more clearly circumscribed family, by
supporting the inclusion of
Avicennia
[6],
Thunbergia
and others that often receiving their own family status [7].
This has, however, led to a situation where the family cannot be definitively and distinctively constrained by
morphological characters [8]. Recent work on the evolution and the diversification of the Acanthaceae provides
a phylogenetic context for assessing the taxonomic significance of possible characters (including micro-mor-
phological structures) within the family [9].
Two subfamilies were recognized by [10]; Anechmatacantheae (without retincula) and Echmatacantheae
(with retinacula). The former comprises two tribes
viz
. Thunbergieae and Nelsonieae, which together include
[2]’s Thunbergioideae, Nelsonioideae and Mendoncioideae. The latter has nine tribes (Hygrophileae, Ruellieae,
Barlerieae, Acantheae, Aphelandreae, Gendarusseae, Eranthemeae, Dicliptereae and Andrographideae)
com-prising [2]’s Acanthoideae.
The family was split by [11] and [12] into three suborders viz.; Thunbergideae (with
Thunbergia
), Ruellideae
(with all the genera having contorted corolla lobes in bud) and Acanthideae (genera with imbricate corolla
lobes). Tribes and sub-tribes were recognized within last two suborders in this system; Nelsonieae is treated as a
tribe of Ruellideae, tribes Gendarussae and Dicliptcreae are amalgamated to form the tribe Justicieae with a
number of sub-tribes. The present family was classified by [13] into five tribes; Thunbergieae, Nelsonieae,
Ru-ellieae, Acantheae and Justicieae. The tribe Thunbergieae and Nesonieae correspond to [2]’s Thunbergioideae
and Nelsonioideae respectively while Acantheae, Ruellieae and Justicieae together constitute his Acanthoideae.
The number of tribes was reduced by [14] to a total of five with Thunbergieae being reduced to tribal level; the
Ruellieae, Nelsonieae and Acantheae remaining mostly unchanged. However in this system the tribe Justicieae
was expanded, comprising the previously recognized tribes of Barlerieae, Andrographideae and Asystaceae at
the sub-tribal level. Apart from the reduction in the rank of some tribes to sub-tribes, this classification is similar
to [11] [12]’s treatment.
For the first time Thunbergiaceae was raised as a separate family by [15] accommodating the first three
sub-families of Acanthaceae sensu [2], [13]’s Acanthaceae comprising [2]’s subfamily Acanthoideae only. Two
subfamilies were formulated by [16]; Thunbergioideae and Acanthoideae that correspond to [13]’s
Thunbergia-ceae and AcanthaThunbergia-ceae and [10]’s Anechmatacantheae and Echmatacantheae. [14]’s Thunbergioideae also
in-cludes the first three sub-tribes of [2]. [17] comprised five tribes and sixteen sub-tribes, this classification has
been accepted by many taxonomists and is in use even today [18]-[24]. In other works [25]-[28], it is well
ap-preciated, though with some modifications.
The extensive work of [5] was thrown much light on the classification of the family. According to him [2]’s
Thunbergioideae and Mendoncioideae show greater affinity to Bignoniaceae and Pedaliaceae than to latter’s
Acanthoideae; and his Nelsonioideae is very much related to the tribe Rhinantheae of Scrophulariaceae. In this
consideration Bremecamp suggested to place Nelsonioideae near Rhinantheae as long as the latter is placed in
Scrophulariaceae. But this reshuffling has received varying response. He was also recognized two subfamilies
viz
. Acanthoideae and Ruellioideae, the former with five tribes and the latter with seven tribes. Along with
pol-len characters [5] did use other supporting characters such as nature of corolla, fruits and seeds. But within
Jus-ticeae there are a number of genera with uncertain positions [5]. He proposed many changes in the generic
lim-its.
There have been some molecular systematic studies specifically addressing the higher level systematics of
Acanthaceae [3] [32]-[34]. The genes used for these studies were rbcL, ndhF, trnL-trnF and trnL-trnF combined
with ITS, respectively. The rbcL data of [32] was re-analyzed by [3] but provided limited resolution within
Acanthaceae. The strict consensus tree of [3] and [34] resolved Acanthaceae sensu [2] as a group. These
analys-es demonstrated that there is no support for separating
Thunbergia
from Acanthaceae as proposed by [5]. The
strict consensus tree of [33] resolved Acanthaceae sensu [2] with the exception of
Elytraria
(Nelsonioideae)
which remained unresolved. [35] showed that
Thunbergia
, are a natural group based upon the shared possession
of woody bristles on the anthers, lack of an endothecial cell layer, elongated connective tips and poricidal
open-ing of the thecae.
The main objective of the present study is to throw light on the phenetic relationships and to explore the
con-tribution of morphological and molecular characters in systematics of Acanthaceae.
2. Materials and Methods
2.1. Sampling
The present study was conducted on 30 species of Acanthaceae in Egypt belonging to 17 genera that were
col-lected from some Egyptian botanical gardens.
Blepharis edulis
and
Avicennia marina
were collected from
natu-ral habitats in Sinai Peninsula (
Table 1
). The identification of wild taxa was taken by the aid of [36] and [37],
while the horticultural taxa with the help of [38] and [39]. The taxa were further matched against the dried
spe-cimens in the Herbaria at Ain Shams University, Faculty of Science (CAIA), Cairo University, Faculty of
Science (CAI), Flora and Phytotaxonomy Research Department (CAIM) and Orman Botanical Garden (Geiza).
Voucher specimens of the studied species were kept in CAIA.
2.2. Morpholoical Investigation
Macromorphological characters were examined directly from the investigated specimens. Stomatography (LM
and SEM) was carried on the bases of traditional method of [40]. A Reichert Microstar IV microscope was used
for microphotographs documentation at the Plant Taxonomy Research Laboratory, Botany Department, Faculty
of Science, Ain Shams University, Cairo, Egypt. Lamina vein architecture was done according to [41].
Descrip-tive terminology of epidermal characteristics based on [41]-[44] while leaf architectural terminology follows
[41].
2.3. Molecular Investigation
Genomic DNA extraction was performed as suggested by Qigene multisource Genomic DNA Mini-Prep Kit
(USA, cat. No. Ap-MN-MS-GDNA-50). DNA samples of each plant were analyzed individually to detect intra-
specific variations and bulked to detect inter-specific variations. An initial screening of 20 ISSR primers
(suc-cessfully utilized in other plant species, was performed in order to test their readability and amplification
pro-files for polymorphism). After this screening procedure, five ISSR primers were selected (
Table 2
). Polymerase
chain reactions (PCR) were carried out according to [45]. The products of ISSR based PCR were detected by
electrophoresis on agarose gel (1.2% in 1× TBE buffer), then stained with ethidium bromide (0.3 ug/ml).
Am-plicon sizes were estimated using 1 Kb DNA standard (Bioron, Germany). Reproducible bands visualized on the
gels were scored using a binary code (1/0) for their presence or absence based on the UVP gel documentation
system (Gel Works ID advanced software, UVP).
2.4. Phenetic Analysis
Unweighted Pair-Group Method using Arithmetic Averages with SAHN function [46] was used to estimate
states of characters variation among the species, each taxa was considered as operational taxonomic unit (OTU)
and states of characters analysed as binary characteristics. The formation of groups was depending on the values
of similarity. All computations were carried out by the aid of the NTSYS-PC version 2.02 [47].
3. Results
Table 1.
The studied taxa of acanthaceae and avicennia marina.
1 Acanthus mollis L.-Sp. Pl. 2: 639. 1753 [1 May 1753] (IK)
2 A. montanus T. Anderson-J. Proc. Linn. Soc., Bot. 7: 37. 1863 [1864 publ. 1863] (IK)
3 Anisacanthus virgularis Nees-Prodr. [A. P. de Candolle] 11: 445. 1847 [25 Nov 1847] (IK)
4 Aphelandra squarrosa Nees-Fl. Bras. (Martius) 9: 89. (IK)
5 Barleria cristata L.-Species Plantarum 2 1753 (APNI)
6 B. prionitis L.-Sp. Pl. 2: 636. 1753 [1 May 1753] (IK)
7 Blepharis edulis (Forssk.) Pers.
8 Eranthemum nervosum R. Br. ex Roem. & Schult.-Syst. Veg., ed. 15 bis [Roemer & Schultes] 1: 174. 1817 [Jan-Jun 1817] (IK)
9 Fittonia argyroneura E. Coem.-Fl. des Serres xvi. (1865-67) 103. (IK)
10 F. verschaffeltii (Lem.) Van Houtte-Fl. des Serres 15: 186. 1865; cf. R.K.Brummitt in Curtis’s Bot. Mag., 182(4): 167. 1979 (IK)
11 Hypoestes sanguinolenta Hook-Bot. Mag. 91: t. 5511. 1865 (IK)
12 Jacobinia carnea (Lindl.) G. Nicholson-Ill. Dict. Gard. ii. 206 (1885) (IK)
13 Justicia adhatoda L.-Sp. Pl. 1: 15. 1753 [1 May 1753] (IK)
14 J. betonica L.-Sp. Pl. 1: 15. 1753 [1 May 1753] (IK)
15 J. brandegeeana Wassh. & L. B. Sm.-Fl. Ilustr. Catarin. Pt. 1, Acantac., 102 (1969) (IK)
16 J. gendarussa Burm.f.-Fl. Ind. (N. L. Burman) 10. 1768 [1 Mar-6 Apr 1768] (IK)
17 J. spicigera Schltdl.-Linnaea 7: 395. 1832 (IK)
18 Odontonema cuspidatum (Nees) Kuntze-Revis. Gen. Pl. 2: 494. 1891 [5 Nov 1891] (GCI)
19 Pachystachys lutea Nees-Prodr. [A. P. de Candolle] 11: 320. 1847 [25 Nov 1847] (IK)
20 Pseuderanthemum atropurpureum Radlk.-Sitzungsber. Math.-Phys. Cl. Königl. Bayer. Akad. Wiss. München 13: 286. 1884 ; nom. inval. (IK)
21 P. bicolor Radlk.-Sitzungsber. Math.-Phys. Cl. Königl. Bayer. Akad. Wiss. München 13: 286. 1884; nom. inval. (IK)
22 Ruellia alba Nees-Fl. Bras. (Martius) 9: 55 (IK)
23 R. brittoniana Leonard-J. Wash. Acad. Sci. xxxi. 96 (1941); Fernald in Rhodora, xlvii. 7 (1945),cum descr. ampl. et emend (IK)
24 R. glabra B.Heyne ex Roth-Nov. Pl. Sp. 312. 1821 [Apr 1821] (IK)
25 R. humilis Nutt.-Trans. Amer. Philos. Soc. ser. 2, 5: 182. 1835 [late 1835] (IK)
26 Sanchezia nobilis Hook. f.-Bot. Mag. 92: t. 5594. 1866 (IK)
27 Thunbergia alata Bojerex Sims-Bot. Mag. 52: t. 2591. 1825 (IK)
28 T. erecta T. Anderson-J. Proc. Linn. Soc., Bot. 7: 18. 1863 [1864 publ. 1863] (IK)
29 T. grandiflora Roxb.-Edwards’ Botanical Register 6 1820 (APNI)
30 Avicennia marina (Forssk.) Vierh.-Denkschr. Kaiserl. Akad. Wiss., Wien. Math.-Naturwiss. Kl. lxxi. 435 (1907) (IK)
Table 2.
ISSR primer names, sequence, and annealing temperature (Ta).
No. Primer Name Sequence Ta (˚C)
1 890 ACG (GT)7 50˚C
2 17898b (CA)6 GT 40˚C
3 835 (AG)8 CC 55˚C
4 851 (GT)8 CG 55˚C
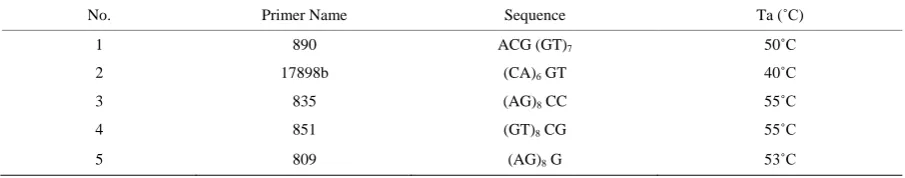
[image:4.595.87.540.631.720.2]all the studied species were summarized in
Tables A1-A4
. The profiles of ISSR that illustrated in
Figure 1
cla-rified that primer 890 produced 15 total bands (three unique and three monomorphic), the remainder nine
poly-morphic bands reaching the polymorphism 80%, primer 1789 b generated 14 bands (one unique and four
mo-nomorphic) with 71.4% polymorphism ratio where the polymorphic bands were nine, 12 polymorphic bands
produced by primer 835 in addithion to one monomorphic and two unique bands the polymorphism reached
93.3%, no unique bands were generated by both primers 851 and 809 the former produced six monomorphic
bands and six polymorphic ones giving 60% polymorphism while the latter produced three monomorphic bands
and 14 polymorphic ones increasing the polymorphism to 82.3%. Coding of character states was produced in
Table 3
. The obtained cladogram (
Figure 2
) showed two series, the second one had two subseries, one of them
divided into two clusters, each one divided into two groups.
(a)
(b)
(c)
(d)
(e)
Figure 1.
ISSR profiles of the studied taxa of Acanthaceae senso lato generated by
Table 3.
Morphological & molecular characters (55), their states (172) and codes of taxa under investigation.
No. Character Character State and its (Code)
1 Duration Perennial (1), Annual (0)
2 Gross morphology Shrub: present (1) Absent (0), Subshrub: present (1) Absent (0), Herb: present (1) Absent (0)
3 habit Climbing (1), Erect (0)
4 Leaf composition Lobed (1), Simple (0)
5 Leaf arrangement Decussate (1), Superposed (0)
6 Leaf shape Lanceolate: present (1) Absent (0), Ovate: present (1) Absent (0), Obovate: present (1) Absent (0), Oblonovate: present (1) Absent (0), Cordate: present (1) Absent (0)
7 Leaf margin Entire: present (1) Absent (0), Spiny: present (1) Absent (0), Sinuate: present (1) Absent (0), Crenate: present (1) Absent (0)
8 Leaf apex Acute: present (1) Absent (0), Acuminate: present (1) Absent (0), Obtuse: present (1) Absent (0)
9 Leaf texture Glabrous: present (1) Absent (0), Hairy: present (1) Absent (0), Spiny: present (1) Absent (0)
10 Leaf base Cuneate: present (1) Absent (0), Rounded: present (1) Absent (0), Cordate: present (1) Absent (0), Truncate: present (1) Absent (0)
11 Bract Present (1) Absent (0)
12 Sepals number Five: present (1) Absent (0), Four: present (1) Absent (0), Numerous: present (1) Absent (0)
13 Sepals union United (1) Free (0)
14 Corolla shape 1-lipped: present (1) Absent (0), Bilabiate: present (1) Absent (0), Funnel: present (1) Absent (0)
15 Petals texture Hairy (1) Glabrous (0)
16 Corolla tube length Long (1) Short (0)
17 Stamens number two (1) Four (0)
18 Stamens fertility Sterile (1) Fertile (0)
19 Stamens length Exerted (1) Included (0)
20 Stamens symmetry Unequal (0) Equal (1)
21 Stamens texture Hairy (1) Glabrous (0)
22 Ovary texture Hairy (1) Glabrous (0)
23 Stigma shape Forked: present (1) Absent (0), Single: present (1) Absent (0), lobed: present (1) Absent (0)
24 Basal nectary disc present (1) Absent (0)
25 1ry vein category Basal actinodromus (1) Pinnate (0)
26 2ry vein category
Festooned semicraspedoromum: present (1) Absent (0), Semicraspidodromus: present (1) Absent (0), Festooned brochidodromus: present (1) Absent (0), Brochidodromus: present (1) Absent (0), Eucamptodromous: present (1) Absent (0), Weak brochidodromus: present (1) Absent (0)
27 2ry vein spacing Uniform: present (1) Absent (0), Irregular: present (1) Absent (0),
Decreasing toward base: present (1) Absent (0), Increase toward base: present (1) Absent (0)
28 2ry vein angle Abruptdly increase to base: present (1) Absent (0), Uniform: present (1) Absent (0), Smoothly increase toward base: present (1) Absent (0), Smoothly decrease toward base: present (1) Absent (0)
29 Intersecondary veins Absent: present (1) Absent (0), Weak: present (1) Absent (0), strong: present (1) Absent (0)
30 3ry vein category
Alternate percurrent: present (1) Absent (0), Random reticulate: present (1) Absent (0), Opposite percureent: present (1) Absent (0), Mixed opp/alte: present (1) Absent (0),
Regular polygonal reticulate: present (1) Absent (0)
Continued
32 3ry vein angle Obtuse: present (1) Absent (0), Acute: present (1) Absent (0), Perpendicular: present (1) Absent (0)
33 4ry vein category Alternate percurrent: present (1) Absent (0), Dichotomizing: present (1) Absent (0), Regular polygonal reticulate: present (1) Absent (0), Ill-developed: present (1) Absent (0)
34 5ry vein category Dichotomizing: present (1) Absent (0), Regular polygonal reticulate: present (1) Absent (0), Ill-developed: present (1) Absent (0)
35 Aereolation Well developed: present (1) Absent (0), Moderately developed: present (1) Absent (0), lacking: present (1) Absent (0)
36 Free ending ultimate vein
Absent: present (1) Absent (0), Unbranched: present (1) Absent (0), 1-branched: present (1) Absent (0), 2-or more branched: present (1) Absent (0)
37 Marginal
ultimate venation Incomplete loop(1) Looped(0)
38 Abaxial Lamina
epidermal cell shape Irregular (1)Polygonal (0)
39 Abaxial Epidermal cell anticlinal wall
Straight: present (1) Absent (0), Undulate: present (1) Absent (0), Sinuous: present (1) Absent (0), Curved: present (1) Absent (0)
40 Abaxial Stomata type Diacytic: present (1) Absent (0), Amphidiacytic: present (1) Absent (0), Polocytic : present (1) Absent (0), Amphipericytic: present (1) Absent (0)
41 Abaxial Abnormal stomata Present (1) Absent (0)
42 Trichome type Absent: present (1) Absent (0), Non glandular uniseriate: present (1) Absent (0), Non glandular unicellular: present (1) Absent (0), Glandular: present (1) Absent (0)
43 Abaxial Trichome
foot cell Present (1) Absent (0)
44 Abaxial Crystals Present (1) Absent (0)
45 Leaf type Amphistomatic (1) Hypostomatic (0)
46 Lamina surface sculpture
Colliculate: present (1) Absent (0), Ruminate: present (1) Absent (0), Reticulate: present (1) Absent (0), Rugose: present (1) Absent (0).
47 Anticlinal wall width Broad (1) Narrow (0)
48 Anticlinal wall elevation Depressed (1) Raised (0)
49 Periclinal wall elevation Raised (1) Depressed (0)
50 Periclinal wall texture Striate (1) Smooth (0)
51 Primer 890 Bands 1 - 12: Present (1) Absent (0)
52 Primer 17898b Bands 1 - 10: Present (1) Absent (0)
53 Primer 835 Bands 1 - 14: Present (1) Absent (0)
54 Primer 851 Bands 1 - 6: Present (1) Absent (0)
55 Primer 809 Band 1 - 14: Present (1) Absent (0)
4. Discussion
The phenetic analysis of both morphological and molecular attributes generated a dendrogram that clarifies the
segregation of genus
Avicennia
as a distinct identity owing to three characters
viz
. the shrub habit, the presence
of moderately developed areolation and the ocellate adaxial surface sculpture. This is in accord with [6] who
used data from both the chloroplast and the nuclear genome, that implied the black mangrove genus
Avicennia
,
usually treated as a separate family in Lamiales or as a genus within Verbenaceae, but more recently has been
placed by some botanists in the monogeneric family Avicenniaceae and recent phylogenetic study [48] has
sug-gested that
Avicennia
is derived from Acanthaceae but is included in that family according to [49].
Figure 2.
UPGMA dendrogram based on 173 morphological and molecular attributes of the studied taxa of Acanthacea
sen-so lato.
Inside Series II,
Blepharis edulis
segregated as a distinct identity at a taxonomic distance less than 1.176
ow-ing to the incomplete looped marginal ultimate vein. The core of the present series distributed into two subseries
(A & B) at a taxonomic distance 1.176. In taxonomic context
Blepharis
and
Acanthus
are joined in the same
tribe Acantheae of most current taxonomic systems [2] [5] [10] [13] [14] [50] [51] the present study did not
support this combination, while the segregation of
Blepharis
from Series II at taxonomic distance near
Acanthus
group may maintain the combination of
Blepharis
with
Acanthus
in a single tribe as the previous taxonomic
systems.
Subseries A that marked with valuable taxonomic traits viz. the climbing habit and the numerous sepals,
in-cluded
Thunbergia alata
,
T. grandiflora
&
T. erecta
, this is in agreement with [10] [13] [14] who recorded
Thunbergia
in a tribe Thunbergiae while [2] [50] [51] recorded the same genus in a subfamily Thunbergioideae.
[52] reported that the Thunbergioideae comprised five genera,
Thunbergia
is the largest of them, and the
subfa-mily is characterized by a predominantly twining habit, enlarged bracteoles, and a reduced calyx. Furthermore
Thunbergioideae lack the retinaculate fruits found in core Acanthaceae.
Group II includes
Pseuderanthemum atropurpureum
,
P. bicolor
&
Sanchizia nobilis
,
Aphelandra squarossa
,
Odontonema cuspidatum
,
Anisacanthus virgularis
,
Justicia gendraussa
,
jacobinia carnea
,
J. specigera
,
Pa-chystachys lutea
,
J. adhatoda
,
J. betonica
&
J. brandegeeana
.
Pseuderanthemum atropurpureum
,
P. bicolor
&
Sanchizia nobilis
separated in a Subgroup A at a taxonomic
distance less than 1.008, owing to the presence of sterile stamens. This is in accord with [50], while in contrast
with [4] [5] that placed
Pseuderanthemum
and
Sanchizia
in two different tribes; Trichanthereae and Justicieae
respectively.
Genus
Odontonema
and
Aphelandra
separated away from the remaining members in a subgroup B in clan 1 at
a taxonomic distance less than 0.868 owing to the reticulate adaxial surface sculpture.
Justicia
species with
Ja-cobinia
and
Pachystachys
srparated in a Clan 2 at a taxonomic distance 1.036. The inclusion of
Pachystachys
with
Justicia
and
Jacobinia
have been recorded in tribe Gendarusseae according to [10], and in a tribe Justicieae
according to [4] [5], while [2] segregated
Pachystachys
in a single tribe named Graptophylleae. [50] combined
Eranthemum
,
Ruellia
&
Sanchizia
(subtribe Ruelliinae),
Anisacanthus
,
Fittonia
,
Hypoestes
,
Justicia
,
Odonto-nema
&
Pseuderanthemum
(Subtribe Justiciinae) and
Barleria
(Subtribe Barleriinae) all in a single tribe
Ruel-lieae.
5. Conclusion
Finally it is recommended that all of these hypotheses clearly merit additional study and need to be tested
against more data, including both more characters and more taxa. For precise delimitation and the membership
of them. In conclusion, the phenetic analysis of both morphological and molecular attributes clarified the
segre-gation of genus
Avicennia
as a distinct identity away from Acanthaceae.
Acanthus mollis
&
A. montanus
are
isolated in its own series that comparable to tribe Acantheae of the current taxonomic systems as the same as the
studied species of
Thunbergia
in its own subseries that comparable to tribe Thunbergiae and
Ruellia
in its own
group that comparable to tribe Ruellieae.
References
[1]
De Jussieu, A.L. (1789) Genara Plantarum. Secundum, Ordines Naturales, Disposita. Parisiis, 307.
[2]
Lindau, G. (1895) Acanthacaeae. In: Engler, A. and Prantel, K.,
Die Naturalichen Pflanzenfamilien
, Vol. 4, 274-354.
[3]
Scotland, R.W., Sweere, J.A., Reeves, P.A. and Olmstead, R.G. (1995) Higher-Level Systematics of Acanthaceae
De-termined by Chloroplast DNA Sequences.
American Journal of Botany
,
82
, 266-275.
http://dx.doi.org/10.2307/2445533
[4]
Bremekamp, C.E.B. (1955) Notes on Some Acanthaceous Genera of Controversial Position.
Acta Botanica
Neerlandi-ca
,
4
, 644-655.
http://dx.doi.org/10.1111/j.1438-8677.1955.tb00358.x
[5]
Bremekamp, C.E.B. (1965) Delimitation and Subdivision of the Acanthaceae.
Bulletin of the Botanical Survey of India
,
7
, 21-30.
[6]
Schwarzbach, A.E. and McDade, L.A. (2002) Phylogenetic Relationships of the Mangrove Family Avicenniaceae
Based on Chloroplast and Nuclear Ribosomal DNA Sequences.
Systematic Botany
,
27
, 84-98.
[7]
Wortley, A.H., Harris, D.J. and Scotland, R.W. (2007) On the Taxonomy and Phylogenetic Position of Thomandersia.
Systematic Botany
,
32
, 415-444.
http://dx.doi.org/10.1600/036364407781179716
[8]
Judd, W.S., Campbell, C.S., Kellogg, E.A., Stevens, P.F. and Donoghue, M.J. (2008) Lamiids (Euasterids I). In: Plant
Systematics: A Phylogenetic Approach. 3rd Edition, Sinauer Associates, Sunderland, 459-494.
[9]
McDade, L.A., Daniel, T.F. and Kiel, C.A. (2008) Toward a Comprehensive Understanding of Phylogenetic
Relation-ships among Acanthaceae s.l. (Lamiales).
American Journal of Botany
,
95
, 1136-1152.
http://dx.doi.org/10.3732/ajb.0800096
[10]
Nees Von Esenbeck, C.G. (1847) Acanthaceae. In: Candolle, A., Ed.,
Prodomus Systema Naturalis
, Vol. 11, Masson,
Paris, 46-519.
[11]
Anderson, T. (1864) An Enumeration of the Species of Acanthaceae from the Continent of Africa and Adjacent Islands.
Journal of the Linnean Society. Botany
,
7
, 13-54.
[12]
Anderson, T. (1867) An Enumeration of the Indian Species of Acanthaceae.
Journal of the Linnean Society. Botany
,
9
,
425-526.
[15]
Van Tieghem, M.P. (1908) Structure du pistil et de l’ovule du fruit et de la graine des Acanthacées.
Annales des
Science Naturelles
,
Serie
9,
Botanique
,
7
, 1-24.
[16]
Wettstein, R.V. (1935) Handbuch der Systematischen Botanik, Vierte Aufl. Franz Deuticke. Leipzig and Wien.
[17]
Leonard, E.C. (1951) The Acanthaceae of Colombia I.
Contributions from the United States National Herbarium
,
31
,
1-115.
[18]
Melchior, H. (1964) Engler’s syllabus der Pflanzenfamilien. Gerbruder Borntraeger Berlin.
[19]
Wasshaussen, D.C. (1966) Acanthaceae in Lundel, C.L. of Texas. Vol. 1, Renner, Texas, 276-282.
[20]
Heine, H. (1962) Notes on Some West African Acanthaceae: The Reduction of the Genus Asteracantha Nees to
Hy-grophila R.Br.
Kew Bulletin
,
16
, 171-173.
[21]
Heine, H. (1966) Acanthaceae in Flora due Gabon, Paris.
[22]
Hutchinson, J. (1973) The Families of Flowering Plants. 3rd Edition, The Clarendon Press, Oxford, 519-524.
[23]
Gibson, D.N. (1974) Acanthaceae in Flora of Guatemela. Fieldiana.
Botany
,
24
, 328-461.
[24]
Thorne, R.F. (1992) Classification and Geography of the Flowering Plants.
Botanical Review
,
58
, 225-348.
http://dx.doi.org/10.1007/BF02858611
[25]
Barker, R. (1986) Revision of Australian Acanthaceae.
Journal of the Adelaide Botanic Gardens
,
9
, 1-292.
[26]
Valsaladevi, G. (1987) Cytological and Palynological Studies on the South Indian Acanthaceae. Ph.D. Thesis,
Univer-sity of Kerala, Trivandrum.
[27]
Balkwill, K. and Norris, F. (1988) Classification of the Acanthaceae. A Southern African Perspective.
Monographs of
Systematic Botany of the Missouri Botanical Garden
,
25
, 503-516.
[28]
Scotland, R.W. (1993) Pollen Morphology of Contortae (Acanthaceae).
Botanical Journal of the Linnean Society
,
111
,
471-504.
http://ora.ox.ac.uk/objects/uuid:cd07aee0-b24c-4ef3-a5b8-816c0ae9e06c
http://dx.doi.org/10.1111/j.1095-8339.1993.tb01916.x
[29]
Gopal, S.C. and Jayantilal, A.I. (1984) Leaf Architecture of Some Acanthaceae.
Botanical Magazine Tokyo
,
97
, 469-
481.
http://dx.doi.org/10.1007/BF02489579
[30]
Shendage, S.M. and Yadav, S.R. (2009) Cuticular Studies in Genus Barleria L. (Acanthaceae) from India.
Journal of
the Indian Botanical Society
,
88
, 176-183.
[31]
Patil, A.M. and Patil, D.A. (2011) Investigations on Foliar Epidermal Characteristics in Some Acanthaceae.
Current
Botany
,
2
, 1-8.
[32]
Hedren, M., Chase, M.W. and Olmstead, R.G. (1995) Relationshipsin the Acanthaceae and Related Families as
Sug-gested by Cladistics Analysis of rbcL Nucleotide Sequences.
Plant Systematics and Evolution
,
194
, 93-109.
http://dx.doi.org/10.1007/BF00983219
[33]
McDade, L.A. and Moody, M.L. (1999) Phylogenetic Relationships among Acanthaceae: Evidence from Non-Coding
trnL-trnF Chloroplast DNA Sequences.
American Journal of Botany
,
86
, 70-80.
http://dx.doi.org/10.2307/2656956
[34]
McDade, T.F., Daniel, S.E. and Riley, K.M. (2000) Phylogenetic Relationships within the Tribe Justicieae
(Acantha-ceae): Evidence from Molecular Sequences, Morphology, and Cytology.
Annals of the Missouri Botanical Garden
,
87
,
435-458.
http://dx.doi.org/10.2307/2666140
[35]
Schönenberger, J. and Endress, P.K. (1998) Structure and Development of the Flowers in Mendoncia, Pseudocalyx,
and Thunbergia (Acanthaceae) and Their Systematic Implications.
International Journal of Plant Sciences
,
159
, 446-
465.
http://dx.doi.org/10.1086/297563
[36]
Täckholm, V. (1974) Student’s Flora of Egypt (ed. 2), Cairo University, 501-503.
[37]
Boulos, L. (2002) Flora of Egypt. Vol. 3 (Verbenaceae-Compositae). Al Hadara Publishing, Cairo, 97-104.
[38]
Bailey, L.H. (1949) Manual of Cultivated Plants. The Macmillan Company, New York.
[39]
Bailey, L.H. and Bailey, E.Z. (1976) A Concise Dictionary of Plants Cultivated in the U.S. and Canada. Hortus Third
“Revised by Staff of the L. H. Bailey Hortium”. The Macmillan Publishing Company. New York.
[40]
Stace, C.A. (1965) Cuticular Studies as an Aid to Plant Taxonomy.
Bulletin of the British Museum
(
Natural History
)
Botany
,
4
, 3-78.
[41]
LAWG (1999) Manual of Leaf Architecture: Morphological Description and Categorization of Dicotyledonous and
Net-Veined Monocotyledonous Angiosperms. Smithsonian Institution, Washington DC.
[42]
Metcalfe, C.R. and Chalk, L. (1950) Anatomy of Dicotyledons. Vol. 2, Clarendon Press, Oxford.
[43]
Murley, M.R. (1951) Seeds of the Cruciferae of Northeastern North America.
American Midland Naturalist
,
46
, 1-81.
[44]
Prabhakar, M. (2004) Structure, Delimitation, Nomenclature and Classification of Stomata.
Acta Botanica Sinica
,
46
,
242-252.
[45]
Whitty, P.W., Powell, W. and Sprent, J.I. (1994) Molecular Separation of Genera in Cassiinae (Leguminosae), and
Analysis of Variation in the Noulating Species of Chamaecrista.
Molecular Ecology
,
3
, 507-515.
http://dx.doi.org/10.1111/j.1365-294X.1994.tb00129.x
[46]
Sneath, P.H. and Sokal, R.R. (1973) Numerical Taxonomy. Freeman, San Fransisco, 573.
[47]
Rohlf, P.J. (2000) NTSYS-PC. Numerical Taxonomy and Multivariate Analysis Systems Exeter Publishing, New
York.
[48]
Abdel-Hameed, U.K., Salim, M.A., Mourad, M.M., Ishak, F.I. and Tantawy, M.E. (2014) Phylogenetic Analysis of
Acanthaceae in Egypt Based on Morphological Criteria.
Vegetos
,
27
, 29-39.
[49]
A.P.G. (2009) An Update of the Angiosperm Phylogeny Group Classification for the Orders and Families of Flowering
Plants: APG III.
Botanical Journal of the Linnean Society
,
161
, 105-121.
http://dx.doi.org/10.1111/j.1095-8339.2009.00996.x
[50]
Scotland, R.W. and Vollesen, K. (2000) Classification of Acanthaceae.
Kew Bulletin
,
55
, 513-589.
http://dx.doi.org/10.2307/4118776
[51]
Engler, A. and Prantl, K. (1897) Die Natürlichen Pflanzenfamilien nebst ihren Gattungen und wichtigeren Arten,
in-sbesondere den Nutzpflanzen, unter Mitwirkung zahlreicher hervorragender Fachgelehrten.
Leipzig
:
W. Engelmann
,
4
,
274-354.
Appendix
Table A1.
Macromorphological characters of the studied taxa.
C
T Duration
Gross
morphology Habit
Leaf composition
Leaf arrangement
Lamina shape
Lamina margin
Lamina apex
Lamina texture
Lamina base
1 Annual Herb Erect Lobed Decussate Lanceolate Sinuate Acute Glabrous Cuneate
2 Perennial ≈ ≈ ≈ ≈ ≈ Spiny Acuminate Spiny ≈
3 ≈ Sub-shrub ≈ Simple ≈ ≈ Entire ≈ Glabrous ≈
4 Annual Herb ≈ ≈ ≈ Obovate ≈ ≈ ≈ ≈
5 Perennial Sub-shrub ≈ ≈ ≈ Ovate ≈ ≈ Hairy ≈
6 ≈ ≈ ≈ ≈ ≈ ≈ ≈ acute ≈ ≈
7 ≈ Herb ≈ ≈ Superposed Lanceolate Spiny ≈ ≈ ≈
8 ≈ Sub-shrub ≈ ≈ Decussate Ovate Entire Acuminate Glabrous ≈
9 Annual Herb ≈ ≈ ≈ ≈ ≈ Acute Hairy Rounded
10 ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈
11 ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ Glabrous ≈
12 Perennial Sub-shrub ≈ ≈ ≈ Lanceolate ≈ ≈ ≈ Cuneate
13 ≈ ≈ ≈ ≈ ≈ Ovate ≈ Acuminate ≈ ≈
14 ≈ ≈ ≈ ≈ ≈ ≈ ≈ Acute ≈ ≈
15 ≈ Herb ≈ ≈ ≈ ≈ ≈ ≈ Hairy ≈
16 ≈ Sub-shrub ≈ ≈ ≈ Lanceolate ≈ ≈ Glabrous ≈
17 ≈ ≈ ≈ ≈ ≈ Oblongovate ≈ ≈ ≈ ≈
18 ≈ ≈ ≈ ≈ ≈ Ovate ≈ Acuminate ≈ ≈
19 ≈ ≈ ≈ ≈ ≈ Lanceolate ≈ ≈ ≈ ≈
20 ≈ ≈ ≈ ≈ ≈ Ovate ≈ Obtuse ≈ Rounded
21 ≈ ≈ ≈ ≈ ≈ Ovate ≈ Acute ≈ Cuneate
22 Annual Herb ≈ ≈ ≈ Ovate ≈ ≈ Hairy ≈
23 ≈ ≈ ≈ ≈ ≈ Lanceolate ≈ ≈ Glabrous ≈
24 ≈ ≈ ≈ ≈ ≈ Ovate ≈ ≈ Glabrous ≈
25 ≈ ≈ ≈ ≈ ≈ Ovate ≈ ≈ Hairy ≈
26 Perennial Sub-shrub ≈ ≈ ≈ Ovate Crenate Acuminate Glabrous ≈
27 ≈ ≈ Twining ≈ Superposed Cordate Sinuate Acute Hairy Cordate
28 ≈ ≈ ≈ ≈ ≈ Ovate Entire Acuminate Glabrous ≈
29 ≈ ≈ ≈ ≈ ≈ Ovate Sinuate ≈ Glabrous Truncate
C
T Bract
Sepal No.
Sepal union
Corolla shape
Corolla tex.
Corolla tube
No. of
stamens Staminodes L.R.C.
Stamens symmetry
Stamens tex.
Gynoecium tex.
Stigma
shape Disc
1 + 5 United 1-lipped G. Short 4 Absent Exerted Equal G. G. Forked -
2 ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈
3 ≈ ≈ ≈ Funnel ≈ Long 2 ≈ ≈ ≈ ≈ ≈ Single +
4 ≈ ≈ Free Bilabiate ≈ ≈ 4 ≈ Included ≈ ≈ ≈ Forked -
5 ≈ 4 United Funnel ≈ ≈ ≈ ≈ ≈ Unequal ≈ ≈ Single +
6 ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈
7 ≈ ≈ Free 1-lipped ≈ Short ≈ ≈ ≈ Equal ≈ ≈ ≈ ≈
8 ≈ 5 United Funnel ≈ Long ≈ ≈ Exerted ≈ ≈ ≈ Lobed -
9 ≈ ≈ ≈ ≈ ≈ Short 2 ≈ Included ≈ ≈ ≈ Single ≈
10 ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈
11 ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ +
12 ≈ ≈ ≈ Bilabiate ≈. Long ≈ ≈ Exerted ≈ ≈ ≈ ≈ ≈
13 ≈ ≈ ≈ ≈ H. Short ≈ ≈ ≈ ≈ ≈ H. Forked ≈
14 ≈ ≈ ≈ ≈ G. ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈
15 ≈ ≈ ≈ ≈ H. Long ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈
16 ≈ ≈ ≈ ≈ G. Short ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈
17 ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ Single ≈
18 ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ Forked ≈
19 ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈
20 ≈ ≈ ≈ Funnel ≈ Long ≈ 2 Included Unequal ≈ ≈ ≈ -
21 ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈
22 - ≈ ≈ ≈ ≈ ≈ 4 Absent ≈ Equal H. G. Single +
23 ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈
24 ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈
25 ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈
26 + ≈ ≈ Bilabiate ≈ Short 2 2 Exerted Unequal G. H. ≈ -
27 - ∞ Free Funnel ≈ Long 4 Absent ≈ ≈ H. G. ≈ +
28 ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈
29 ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈ ≈
Table A2.
Lamina epidermal characteristics of the studied taxa (LM).
C T Cells shape (Ab/Adaxial) Anticlinal wall (Ab/Adaxial) Stomata type (Ab/Adaxial) Crystals (Ab/Adaxial)Foot cell of Trichomes (Ab/Adaxial) Trichome type Abnormal stomata (Ab/Adaxial) Leaf type
Irregular/Same Sinuous/Curved Diacytic &
Amphidiacytic/- +/+ −/−
NG.
uniseriate +/− Hypostomatic ≈ Curved/Sinuous Diacytic & Polocytic/− −/− +/+ ≈ −/− ≈ ≈ Sinuous/Same Diacytic & polocytic/- +/+ −/− − ≈ ≈
Polygonal/Same Straight/Same Diacytic/- −/− ≈ NG.
uniseriate ≈ ≈
Irregular/Same Sinuous/Same Diacytic& polocytic/- ≈ ≈ ≈ ≈ ≈
≈ Undulate/Same Diacytic/- +/+ +/+ NG.
unicellular ≈ ≈
Polygonal/Same Straight/Sinuous Diacytic/Diacytic, Amphipericytic +/− −/−
NG. unicellular,
glandular
≈ Amphistomatic
≈ Straight/Same Amphipericytic/- +/+ +/+ NG.
uniseriate ≈ Hypostomatic
≈ Straight/undulate Diacytic/- ≈ ≈ ≈ ≈ ≈
Irregular/Same Undulate/Sinuous ≈ −/+ ≈ ≈ ≈ ≈
≈ Undulate/Same ≈ +/+ −/−
NG. unicellular
glandular
+/− ≈
≈ Sinuous Same Diacytic, polocytic/- −/− ≈ NG.
uniseriate −/− ≈
≈ ≈ Diacytic/- ≈ +/− ≈ ≈ ≈
≈ Curved/Same Diacytic/Same ≈ −/− − ≈ Amphistomatic
≈ Undulate/Sinuous Diacytic/- +/− ≈ NG.
uniseriate ≈ Hypostomatic
≈ Undulate/Curved ≈ −/+ +/+ − ≈ ≈
≈ Sinuous/Same Diacytic& polocytic/- −/− −/+ NG.
uniseriate ≈ ≈
Polygonal/Same Straight/Same Diacytic &
amphidiacytic/- ≈ −/− ≈ ≈ ≈
Irregular/Same Undulate/Curved Diacytic/Same +/+ +/+
NG. uniseriate,
glandular
≈ Amphistomatic
Polygonal/Same Straight/Same Diacytic/- −/+ −/+ − ≈ Hypostomatic
≈ ≈ ≈ −/− +/− ≈ ≈ ≈
Irregular/Same Curved/Undulate ≈ ≈ −/−
NG. uniseriate,
glandular
≈ ≈
Polygonal/Same Straight/Same Diacytic,
Amphipericytic/Diacytic ≈ ≈ − ≈ Amphistomatic
≈ Curved/Same Diacytic/Same ≈ ≈ ≈ ≈ Amphistomatic
Irregular/Same ≈ ≈ +/+ ≈ ≈ +/− Amphistomatic
Polygonal/Same Straight/Same Amphipericytic/same +/− +/+ ≈ −/− ≈
Irregular/Same Curved/Sinuous Diacytic/Same −/− −/−
NG. uniseriate,
NG. unicellular
≈ ≈
≈ Sinuous/Same Diacytic/- ≈ ≈ − ≈ Hypostomatic
≈ Curved/Same ≈ +/− ≈ NG.
unicellular ≈ ≈
Polygonal/Same Curved/Straight ≈ −/− ≈ − ≈ ≈
Table A3.
Lamina epidermal characteristics of the studied taxa (abaxial surface, SEM).
C
T Sculpture
Anticlinal wall width
Anticlinal wall elevation
Anticlinal wall surface
Periclinal wall elevation
Periclinal wall surface
Ruminate Broad Depressed Smooth Raised Smooth
Colliculate Narrow ≈ ≈ ≈ ≈
Ruminate ≈ ≈ ≈ ≈ ≈
Reticulate ≈ Raised ≈ Depressed ≈
≈ ≈ ≈ ≈ ≈ Striate
Colliculate ≈ Depressed ≈ Raised Smooth
Ruminate ≈ ≈ ≈ ≈ Striate
Reticulate ≈ Raised ≈ Depressed ≈
Rugose ≈ ≈ ≈ ≈ ≈
≈ ≈ ≈ ≈ ≈ ≈
Reticulate ≈ ≈ ≈ ≈ ≈
Rugose ≈ ≈ ≈ ≈ Smooth
Ruminate Broad Depressed ≈ Raised ≈
≈ ≈ ≈ ≈ ≈ ≈
≈ ≈ ≈ ≈ ≈ Striate
≈ ≈ ≈ ≈ ≈ Smooth
Rugose Narrow Raised ≈ Depressed ≈
Reticulate ≈ ≈ ≈ ≈ ≈
Rugose ≈ ≈ ≈ ≈ ≈
Ruminate ≈ Depressed ≈ Raised ≈
≈ ≈ ≈ ≈ ≈ ≈
Rugose ≈ Raised ≈ Depressed ≈
Reticulate ≈ ≈ ≈ ≈ ≈
Rugose ≈ ≈ ≈ ≈ ≈
Ruminate ≈ Depressed ≈ Raised ≈
≈ ≈ ≈ ≈ ≈ ≈
≈ ≈ ≈ ≈ ≈ ≈
≈ ≈ ≈ ≈ ≈ ≈
≈ ≈ ≈ ≈ ≈ ≈
Table A4.
Lamina vein architecture of the studied taxa.
C T 1 vein category 2 vein category 2 veinspacing 2 vein angle
Inter 2 veins 3 vein category 3 vein course 3 vein angle to 1
Pinnate Festooned
semicraspedoromus Irregular
Abruptdly
increase to base Weak
Alternate percurrent
Exmedially
ramified Acute
≈ Semicraspidodromus Uniform ≈ ≈
Regular polygonal
reticulate
≈ Obtuse
≈ brochidodromus Festooned toward base Decrease Uniform ≈ Random reticulate
Admedially
ramified ≈
≈ ≈ Uniform Smoothly
increase to base Absent Mixed ≈ ≈
≈ ≈ toward base Decrease Uniform Strong ≈ ≈ ≈
≈ ≈ ≈ ≈ Absent ≈ ≈ ≈
≈ Semicraspidodroms Irregular ≈ ≈ Random
reticulate ≈ Acute
≈ Brochidodromus Decrease
toward base Uniform ≈ Mixed Straight Obtuse
≈ ≈ ≈ Smoothly increase toward base
≈ percurent Opposite Sinuous ≈
≈ brochidodromus Festooned ≈ ≈ ≈ Mixed Admedially
ramified ≈
≈ ≈ Irregular ≈ ≈ Alternate
percurrent ≈ ≈
≈ ≈ toward base Decrease Uniform ≈ opposite
percurrent Straight Perpendicular
≈ ≈ ≈ ≈ ≈ Mixed Sinuous Obtuse
≈ ≈ ≈ Smoothly decrease
toward base
Weak Alternate percurrent
Admedially
ramified ≈
≈ ≈ ≈ Uniform Absent Random
reticulate ≈ Acute
≈ Eucamptodromous ≈ ≈ Weak ≈ ≈ ≈
≈ brochidodromus Festooned Uniform ≈ Absent Mixed Straight Obtuse
≈ ≈ toward base Decrease Smoothly increase toward base
Strong reticulate Random Admedially ramified ≈
≈ brochidodromus Weak ≈ Uniform Weak opposite
percurrent Straight ≈
≈ brochidodromus Festooned Uniform
Smoothly increase toward base
Absent Mixed Sinuous ≈
≈ ≈ ≈ Uniform Weak Random
reticulate
Admedially
ramified ≈
≈ Weak brochidodromus Decrease
toward base ≈ Absent ≈ ≈ ≈
≈ ≈ ≈ ≈ ≈ polygonal Regular
reticulate
≈ Perpendicular
Continued
≈ brochidodromus Weak toward base Increase ≈ ≈ ≈ ≈ ≈
≈ brochidodromus Festooned ≈ ≈ ≈ ≈ ≈ Acute
Basal
actiondromus ≈ ≈
Abruptdly
increase to base Weak ≈ ≈ Obtuse
Pinnate ≈ Uniform Uniform Absent ≈ ≈ ≈
Basal
actinodromus Brochidodromus
increase toward base
Abruptdly
increase to base Weak Mixed ≈ Perpendicular
Pinnate Brochidodromus ≈ Uniform ≈ ≈ ≈ Obtuse
C
T 4 vein category 5 vein category Areolation F. E. V. S Mar. veins
Regular polygonal reticulate Dichotomizing Well developed 2 or more branched Looped
Dichotomizing Ill developed ≈ Absent ≈
Alternate percurrent Dichotomizing ≈ 2 or more branched ≈
Regular polygonal reticulate Ill developed Lacking Absent ≈
Alternate percurrent ≈ Well developed ≈ ≈
≈ Dichotomizing ≈ 2 or more branched ≈
Regular polygonal reticulate Ill developed ≈ ≈ Incomplete loop
≈ ≈ ≈ Unbranched Looped
Ill developed ≈ ≈ Absent ≈
Alternate percurrent ≈ ≈ 2 or more branched ≈
≈ ≈ ≈ ≈ ≈
≈ Regular polygonal reticulate ≈ ≈ ≈
≈ ≈ ≈ ≈ ≈
Regular polygonal reticulate Ill developed ≈ ≈ ≈
≈ ≈ ≈ ≈ ≈
≈ Ill developed ≈ 1-branched ≈
Alternate percurrent Dichotomizing ≈ 2 or more branched ≈
Regular polygonal reticulate Ill developed ≈ Absent ≈
≈ Dichotomizing ≈ 2 or more branched ≈
≈ Ill developed ≈ Unbranched ≈
≈ Dichotomizing ≈ 2 or more branched ≈
≈ Regular polygonal reticulate ≈ Absent ≈
≈ Ill developed ≈ 2 or more branched ≈
≈ Dichotomizing ≈ Unbranched ≈
≈ Ill developed ≈ Absent ≈
≈ Dichotomizing ≈ 2 or more branched ≈
≈ Ill developed ≈ Absent ≈
≈ Dichotomizing ≈ 2 or more branched ≈
≈ ≈ ≈ ≈ ≈