Mirroring cannot account for understanding action
Full text
Figure
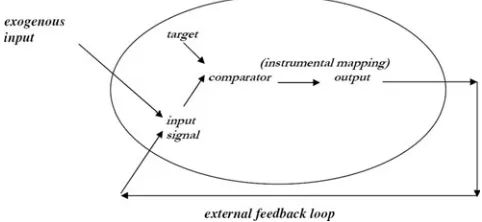
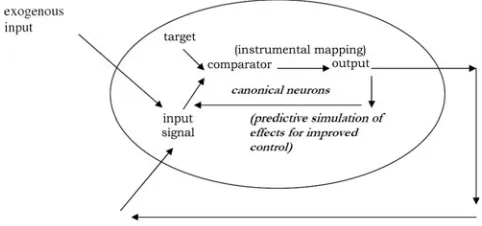
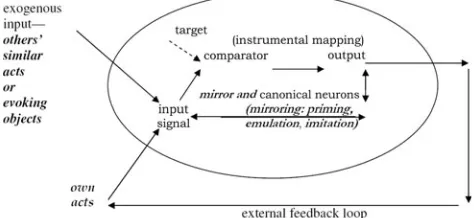
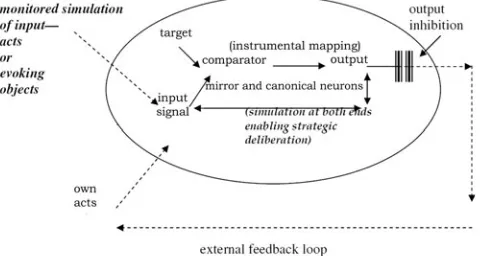
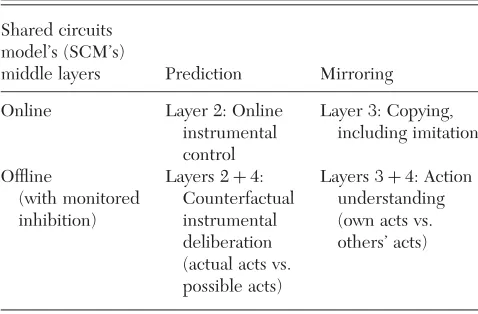
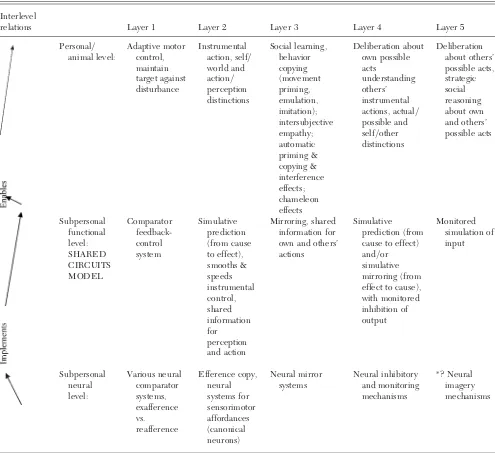
Outline
Related documents
All the banks surveyed reported having continuity and recovery strategies for a wide range of potential terrorist activities and man-made disasters and crises, such as
Also, the positive impact of the effectiveness of containment measures in preventing COVID-19 decease cases seems to fade and even turns to be good for growth potentials:
(You must make a request in writing to obtain access to your healtll intormation. You may obtain a form to request access by using the contact information listed at the end of
Local resolution is the first stage of the NHS complaints process. Local resolution gives you the opportunity to explain what it is you are unhappy about and what you
Effect of routine isoniazid preventive therapy on tuberculosis incidence among hiv-infected men in south africa: A novel randomized incremental recruitment study.. Golub JE,
An analysis of the economic contribution of the software industry examined the effect of software activity on the Lebanese economy by measuring it in terms of output and value
Class Work and Homework Policy:
