0022-538X/79/11-0692/05$02.00/0
Evolution of
Defective-Interfering Double-Stranded
RNAs of
the Yeast Killer
Virus
WILLIAM P. KANE,* DENNIS F.PIETRAS,t AND JEREMY A. BRUENN
Division of Cell and Molecular Biology, State University of New YorkatBuffalo,Buffalo, New York14260 Received for publication 26 June 1979
We havecharacterized by
T,
fingerprint analysisseveral defectiveinterfering(DI) double-stranded RNAs of thesimpleyeastvirusScV.Acommonsequence
ofabout 0.5 to 0.6 kilobasepairs, including both 3' termini of the parental RNA, was presentin eachDIRNA.Several DI RNAs had novel
T,
oligonucleotidesnotpresentin theirparental RNA.
Saccharomyces~cerevisiae has asimple
dou-ble-stranded (ds) RNA virus (ScV) with two,
separately encapsidated
dsRNA's(4,
10). Likeother fungal viruses (17) ScV exhibits only a
latent infection. The larger viral dsRNA (L) encodes the major capsid protein (14). The
smaller (M) encodes a toxin
("killer factor")
lethal to strains without ScV-M (S. cerevisiae
virus-M-containing) particles (1, 16; K. A. Bos-tian et al., Cell, in press). L and M have no
detectable sequencehomology (2), and ScV-M particles are dependent onScV-L
particles
for the synthesis of their capsidprotein
(8, 11). Deletion mutations of M result in defective-in-terfering (DI) particles containing fragments ofM(ScV-S particles) (2,7;thiswork). ScV-M and ScV-S
particles
areequallydependent
on prod-uctsof thenucleargenomefor theirreplication,
products not necessary for the replication of ScV-L particles (19, 20). ScV-S RNAs are de-rived from M by internal deletion, sometimes followed by tandemduplication (2, 7), andthey diverge in sequence from their parental RNA
(2),asdothe RNAs of animal virusesreplicated inpersistent infections(13).
In the present work we utilize two-dimen-sionalgel electrophoresis of
T,
oligonucleotidestocharacterizeseveral S RNAs, including three independently derived S RNAs from parental
strains with differentgeneticbackgrounds. The dsRNA's werephenolextracted andpurifiedby
CF11chromatography and agarose gel electro-phoresis (2, 3). The sizes of dsRNA's were
de-termined by electron microscopy and by
CH3HgOH agarose denaturing gel electropho-resis (2). The two new S dsRNA's examined wereS14,of0.78kb(kilobase pairs), from strain L014 (M. Vodkin); and S733, of 0.77 kb, from
strain K733(R.Wickner). The S dsRNA'swere present inScVparticleswith a density inCsCl
tPresent address:Departmentof ViralOncology,Roswell Park Memorial Institute, Buffalo,NY 14263.
of 1.36 g/ml. ScV-Lparticles had a density of
1.42, and ScV-M particles hadadensity of1.37
g/ml.
We havefingerprinted four of these dsRNA's: Sl(a) from strain T132B NK3; Sl(b) from strain T132B NK1;S14, and S733. Thesearecompared with Mfrom strain T158D SK and witha
pre-viously published
T,
fingerprint of S1 from strain T132B NK3, Sl(c). Uniformly32P04-3-labeled dsRNA waspurified and digested with RNase
T,
aspreviously described (2, 3), and the products were separated by two-dimensional polyacrylamide gel electrophoresis (5, 6). The resolution of the two-dimensional gelsystem isgreater thanthat of the cellulose acetate-hom-ochromatography system previously used (2). We have adoptedanew numbering systemfor both S and M. Figures 1 and2depict only the resolvable large
T,
oligonucleotides,
and Table1gives the pancreatic digestion products of those large
T,
oligonucleotides
ofM presentinone or moreS dsRNA's.All four S dsRNA'sare clearly derived from M. Sl(a), -(b), and -(c) havea minimum of19
large
T,
oligonucleotides of M; S14 and S733 have 17. Each dsRNA has both of the 3'T,
oligonucleotides
of M(oligonucleotides5and13or54of S and43and13 or 44ofM). Therefore, allappear to be theresultof internal delection
ofM,followedbytandem
duplication
in the case ofS1
(2, 7). Threeof the fourSdsRNA's
havelarge
T, oligonucleotides
notpresent in M. Ofthose novel
T,
oligonucleotides previously ob-served in S1 [2, shown in Sl(c)], Sl(b) haspreservedallthree(no. 3, 8, and 20) and added twomore(thetwo
T,
oligonucleotidesinspot4).S1(a)
has lostall
three.S1(b)
haslostoneof the21
T,
oligonucleotides
originally derivedfrom M[no.44ofS1(a) or
Sl(c)].
S14 is
slightly
largerthan S3 (50 to 100 basepairs), but it clearlyarose from M in an event very similar to that whichgeneratedS3. S14has
692
on November 10, 2019 by guest
http://jvi.asm.org/
NOTES 693
3 , s
5?
24 54
23-2 0
5.3
3 40
9 44 3
9>
g
M
exc
5,
24 C,o D5 2'
Kb "'-\46 t4 5
31022K~)
Q014
S()89 K7j---
4h
13 S aJ5Q~-16 2
43\ 556
22:~
C2
iC C
2'.
.%--n 34
,-' 2-; -3 -,- \2 2
I 3BBP /S|r,
-,11 I !,
435 5 _,
.-k2
./, Xc
44
7...
-
S1
(b)
2nd
1st --t
FIG. 1. Two-dimensional gel electrophoresis ofT, digests of32PO4.3-labeleddsRNA's.RefertoTable1for
thepancreaticanalysis oftheT, oligonucleotides. Thepositions ofthebromophenolblue(BB)andthexylene cyanol(XC)markerdyesareshownforeachgeL T1 oligonucleotides ofMthatarepresent inone or moreS dsRNA'sareshaded.(Sl(a)is Sl dsRNAfromstrainT132B NK3.Sl(b)isSl dsRNAfromstrain T132BNKI.
17 ofthe 21 large
T,
oligonucleotides from Mpresentin Sl(c). It has both of the 3'T1 oligo-nucleotides of M. In contradistinction to Si, it hasnodetectable large
T,
oligonucleotides notderivedfrom M.S733,ontheotherhand,hasat
least threelarge
T,
oligonucleotidesnotpresentinM (no. 56, 62, and 65). S733 is similar in size
toS3,preservesboth 3' ends ofM, and contains
16 of the 21 large
T,
oligonucleotides derived from M by S3. S733 has also been observedtoform tandem duplicates, that is, some clonal
isolates of strain K733 produce an S RNA of
twice the usual sizewiththesame
T,
fingerprint.S733hasoneadditionallarge
T,
oligonucleotide (no. 63) derived from M (no. 32) that is not present in any other S dsRNA so far finger-printed.The existenceofthesame3'terminionM, S1,
S33, and S14 has been confirmed by 3' end
labelingandsequenceanalysis (V. Brennanand
J. Bruenn, unpublished data). The T1 finger-prints of in vivo labeled RNA have
demon-strated theheterogeneityof atleast one3'end
ofM;
T,
oligonucleotides 13 and 44 of M havedifferent mobilities but apparently have the
same pancreaticdigestion products (Fig. 1 and
2and Table1). This would be consistentwitha
change in the 3' terminal nucleoside, which is not detected in the pancreatic digestion
prod-ucts.Most of the S dsRNA's have the 3' terminal
T,
oligonucleotide with the mobilityof13, butS733hasonewith themobility of 44(no. 54 of
S733). Sl(a) has both, like M. The other 3' terminus istentatively assignedtoT1 oligonucle-otide no. 5 of S (no. 43 of M). Thepancreatic products andmobilityof thisoligonucleotideare
26
)5
.-,.2 5 2' -.6 ,
2C -.,-) -
1-"I 1 4
2 .- "-/
A
'l-'-/
r-1; 4
,--/ -7,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.507.60.452.71.417.2]694 NOTES
.
0
...-.
S
0
,.
.
0 lbP
FIG. 2. T1fingerprints ofS733and S14arecomparedwiththesameMT,fingerprintsasthose inFig. 1. Notation isasexplainedinFig.1.
consistentwiththe3'sequenceanalysis
(Bren-nan and Bruenn, unpublished data). S733 ap-pearstohavetwopossible3'
T,
oligonucleotidesatthis end (no. 5 and 61) with closely related
sequences.
Among the fourDIRNAsanalyzed, ofwhich threeareindependentlyderived,there isa core
ofsome12large T1 oligonucleotidesfrom Mthat are present ineach S (Table 1),includingboth
3'
T,
oligonucleotides of M. Each of the S dsRNA's thus appears to be the result ofthe internal deletionofM,as suggested for S3 (7).Furthermore, thisdeletion seemsto occurin a
roughly similar place in eachcase.The
complex-ity of theoriginalS dsRNAderived from Mcan, in eachcase,beapproximatedby the expression
C=
N/(0.24)(1-0.24)n-1,
whereNis thenumberof T1 oligonucleotides of sizen orlarger and 0.24
isthefrequency of G residues (2). This figure is
about0.65 to 0.80 kb in eachcase,whichagrees well with the size ofS3,S14,andS733(0.7to 0.8
kb). Thecoresequencecorrespondstoabout 0.5 to 0.6 kb. Therealso appearsto beasize
con-straintin the formation of DIRNAs,since there
areonlythe two size classes: that of S3 and that
ofSi, the latter beingatandem duplicationof
the former. At leastsomeanimal virus DI RNAs
arealsogeneratedbydeletion(15).
Asimple modelcanaccountfor both deletion
andtandem duplication. ScV RNA replication appearstobe similar to that of reovirus (9, 12, 18; J. D. Welschand M.J. Leibowitz, personal communication). Ifa transcriptase copying M
dissociated from its template, reassociated at
some internal sequence, and then completed synthesis, a deletionwould result. If the
tran-scriptase completed the synthesis of the viral
plus strand and dissociated from its template
-.0
--. 1-
on November 10, 2019 by guest
http://jvi.asm.org/
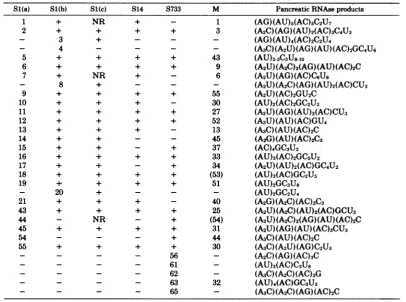
[image:3.507.64.458.74.434.2]TABLE 1. T,oligonucleotides of M and defective interfering dsRNA 's S1(C) NR + + S14 S733 + _ + +
+ + + 43
+ + + 9
NR + - 6
+ + + 55
+ + - 30
+ + + 27
+ + + 52
+ + - 13
+ - - 45
+ - + 37
+ + + 33
+ - + 34
+ + + (53)
+ + + 51
+ + _ 40
+ + + 25
NR - + (54)
+ + + 31
- - + 44
+ + + 30
- - 56
-- - 61
-- - 62
-- - 63 32
- - 65
-Pancreatic RNAse products (AG)(AU)5(AC)5C2U7 (A2C)(AG)(AU)2(AC)3C4U3 (AG)(AU)4(AC)2C2U4 (A3C)(A2U)(AG)(AU)(AC)2GC4U6 (AU)2-3C3U810 (A2U)(A2C)2(AG)(AU)(AC)2C (A2U)(AG)(AC)C6U6 (A2U)(A2C)(AG)(AU)2(AC)CU3 (A2U)(AC)2GU2C (AU)2(AC)2GC2U3 (A2U)(AG)(AU)2(AC)CU3 (A3U)(AU)(AC)GU4 (A3C)(AU)(AC)2C (A2G)(AU)(AC)2C3 (AC)4GC3U2 (AU)3(AC)2GC5U2 (A2U)(AU)2(AC)GC4U2 (AU)3(AC)GC2U5 (AU)2GC3U8 (AU)2GC2U4 (A2G)(A2C)(AC)3C3 (A2U)(A2C)(AU)2(AC)GCU3 (A2U)(A2C)2(AG)(AU)(AC)2C (A2U)(AG)(AU)(AC)3CU3 (A3C)(AU)(AC)2C (A3C)(A2U)(AG)C2U3 (A2C)(AG)(AC)3C (AU)3(AC)C3U8 (A3C)(A2C)(AC)3G (AU)4(AC)GC3U3 (A3C)(A2C)(AG)(AC)2C
aT1 oligonucleotides are numbered asinFig. 1 and2. Symbols: +, present; -, absent;NR,notresolved;
( ), pancreatic RNAse productsnottested.Sl(c) is Si dsRNA analyzedon10April1977from T132BNK3 (2).
Sl(b) is S1 dsRNA analyzedon1June1978from T132B NK1.Sl(a)isSi dsRNAanalyzedon1December 1978
from strain T132B NK3.S14isS14 dsRNA from strain L014. S733 is S733 dsRNA from strainK733,andMis M dsRNA fromstrainT158D SK.
withoutreleasing the plus strand, the
transcrip-tasecould then associate withasecond molecule
of S present in the particle (or with the same
molecule ofS)and continueplus-strand synthe-sis, making atandemly duplicated plus strand.
In both deletion and duplication, a replicase
wouldsynthesize the virus minus strand. Since tandem duplication of L-, M-, and Sl-sized DI RNAsis notobserved,thepresenceoftwo mol-ecules of a dsRNA in one particle may be a
prerequisite for tandem duplication. Two mole-cules of L (4.5 kb)-, M (1.8 kb)-, orSl-sized DI
RNAs (1.5 kb) may not be packaged easily, whereas two molecules of S3-sized DI RNAs (0.75 kb) may.
Theappearance of novel T1 oligonucleotides
in the S dsRNA's and the disappearance of M
T,
oligonucleotides originallypresentareindic-ative of the accumulation ofsequencevariants ofthe DI RNAs. All of the DI RNAsexamined (including S14)areheterogeneousatthe 3' ends (Brennan and Bruenn, unpublished data), in
supportof thisproposition. Similar changes have beenobserved in DI RNAs of vesicular
stoma-titis virus(13). Constantsequencechange is only
tobeexpected in the absence of selective
pres-sure, and the S RNAs do not code for any
proteinsnecessaryformaintenance of the virus.
We thank S. Kahn andJ. Campagna forexcellenttechnical assistance,Loren Fieldand Herb Thomas for electron
micros-copy, and G. Fink, M. Vodkin, and R. Wickner foryeast
strains. This research wassupported byNIH grant 5 RO 1
GM22200toJ. Bruenn.
LITERATURE CTED
1. Bevan,E.A.,and M.Makower. 1963. Thephysiological basis of the killercharacterin yeast,p.202-203.In S.J. Geerts (ed.), Geneticstoday,vol. 1. Macmillan,The Hague.
2. Bruenn, J., and W. Kane. 1978. Relatedness of the double-stranded RNAs present in yeast virus-like
par-ticles.J. Virol. 26:762-772.
3. Bruenn, J.,and B. Keitz. 1976. The 5' ends of yeast killer factor RNAsarepppGp. Nucleic Acids Res. 3: 2427-2436.
4. Buck,K.W.,P.Lhoas,and B. K. Street. 1973. Virus S1(a) 1 2 5 6 7 9 10 11 12 13 14 15 16 17 18 19 21 43 44 45 54 55 Sl(b) 3 4 + + 8 + + + + + +
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.507.53.460.83.385.2]particles in yeast. Biochem. Soc. Trans. 1:1141-1142. 5. DeWachter,R., and W. Fiers. 1972.Preparative two-dimensional polyacrylamide gel electrophoresisof 32P-labeledRNA. Anal. Biochem.49:184-197.
6. Faller,D.V., andN.Hopkins.1977.RNase TI-resistant oligonucleotides of B-tropic murine leukemia virus from BALB/cand five of itsNB-tropicderivatives. J. Virol. 23:188-195.
7.Fried, H. M., andG. Fink. 9978.Electronmicroscopic heteroduplex analysisof"killer"double-strandedRNA speciesfromyeast. Proc. Natl. Acad. Sci. U.S.A. 75: 4224-4228.
8.Harris, M. S. 1978. Virus-like particles and double-stranded RNA from killer and non-killer strains of Saccharomycescerevisiae. Microbios 21:161-176. 9. Hastie, N., V. Brennan, and J. Bruenn. 1978. No
homology between double-stranded RNA and nuclear DNA ofyeast. J.Virol.28:1002-1005.
10. Herring, A. J.,and E. A.Bevan.1974.Virus-like par-ticlesassociated with the double-stranded RNAspecies foundinkillerand sensitive strains oftheyeast Saccha-romycescerevisiae.J. Gen. Virol.22:387-394. 11. Herring, A.J., and E. A. Bevan.1975.Double stranded
RNA containingparticles from the yeast Saccharomy-ces cerevisiae and their relationship to the "killer" character,p. 149-154. InS. Puiseux-Dao (ed.), Molec-ularbiology of nuclear cytoplasmic relationships. Elsev-ierScientific, Amsterdam.
12. Herring,A. J. and E. A. Bevan.1977.Yeast virus-like particles possess a capsid-associated single-stranded RNApolymerase.Nature(London) 268:464-466.
13. Holland,J.J.,E.A.Grabau, C.L.Jones, and B.L. Semler. 1979. Evolution ofmultiplegenomemutations during long-term persistent infection byvesicular
sto-matitis virus.Cell 16:495-504.
14. Hopper, J. E.,K. A.Bostian, L.B.Rowe, andD. J. Tipper. 1977.Translationof theL-speciesdsRNA ge-nome of thekiller-associatedvirus-likeparticlesof Sac-charomycescerevisiae. J. Biol. Chem.252:9010-9017. 15. Nomoto, A., A. Jacobson,Y.F. Lee, J.Dunn, andE. Wimmer. 1979. Defective interfering particles ofpolio virus: mapping of the deletion and evidence that the deletions in the genomesDI(1),(2)and (3) are located in the same region. J. Mol. Biol.128:179-196. 16. Palfree,G.E.,and H.Bussey.1979.Yeast killer toxin:
purification andcharacterization of the protein toxin from Saccharomyces cerevisiae. Eur. J. Biochem.93: 487-493.
17. Saksena,K.N.,and P. A. Lemke. 1978.Virusesinfungi, p. 103-143.H. Frankel-Conratand R. R. Wagner (ed.), Comprehensive virology, vol. 12. Plenum Press, New York.
18. Silverstein,S. C., J. K.Christman,and G. Acs.1976. The reovirus replicative cycle.Ann. Rev.Biochem.45: 375-408.
19. Somers,J. M. 1973. Isolation of suppressive sensitive mutantsfrom killer and neutralstrains of Saccharo-mycescerevisiae. Genetics 74:571-579.
20. Wickner, R. B. 1978. Twenty-six chromosomal genes needed to maintain the killer double-stranded RNA plasmidofSaccharomyces cerevisiae. Genetics 88:419-425.