0022-538X/83/060737-12$02.00/0
Copyright © 1983, AmericanSocietyforMicrobiology
Identification of Adenovirus Genes
That
Require
Template
Replication for
Expression
LYLE D.CROSSLANDtANDHESCHEL J. RASKAS*
Department of Pathology, Washington University School of Medicine, St. Louis, Missouri 63110
Received 15 November 1982/Accepted 1 March 1983
Therelationshipbetween adenovirus type 2 DNA replication and expression of
intermediate stage viral genes was investigated. The 1.03-kilobase mRNA from
early region lb (Elb)and the mRNAscoding for proteins IX and IVa2 were first
detectedbetween 6and8 hpostinfection. Inhibition of viral DNAreplication with
hydroxyurea prevented expressionof the IX andIVa2 mRNAs, but not of the Elb
mRNA. Pulse-labelingexperiments demonstrated that the block ofIX and IVa2
expression in hydroxyurea-treated cells was at the level oftranscription. By a
series of superinfectionexperiments, it was determined that the viral and cellular
factors present during the late stage of adenovirus infection are insufficient to
activate IX geneexpression. The viral DNAtemplate must firstreplicatebefore
IXtranscription canbegin.
Lytic infection by humanadenoviruses is
di-vided into four temporal stages of gene
expres-sion: pre-early, early, intermediate, and late (5,
19, 34).Theassignmentofgenes to aparticular
stagehas been basedonthetime ofappearance
of the gene product. mRNAs that appear
be-tween 6 and 8 h after infection have been
as-signedtotheintermediate class(34, 38).
Includ-edin this classarethe0.52-kilobase (kb)mRNA
from region Ela, the 1.03-kbmRNAfrom Elb,
and themRNAsforprotein IVa2 and proteinIX
(Fig. 1). Previous work has indicated that the
appearance at intermediate times ofthe0.52-kb
and 1.03-kb mRNAs is due to increased
cyto-plasmic stability ofmRNA species producedin
the nucleus at early times (41). The IX gene,
however, is regulated atthetranscription level,
andits promoterisnotactive untilintermediate
times (42). The mode ofregulation of the IVa2
gene has notbeen determined.
Thepromoterfor themajor late transcription
unit (Fig. 1) is active at early times, although
premature termination of the transcripts pre-vents formationof the mRNAs for late regions L2 through L5 (2, 29, 35). The onset of viral
DNA replication has traditionally marked the
beginning ofthe late stage of adenovirus
infec-tion (8). Thomas and Mathews (39)
demonstrat-ed that the gene products of late regions L2
throughL5 areexpressedonlyfrom a replicated
DNAtemplate;eventhough themajor late
pro-moterisactiveonparental genomes,replication
tPresent address: The Biological Laboratories, Harvard University,Cambridge,MA 02138.
and some coincident physical change in the
template are required for expression of the
downstream late regions. The relationship of
intermediate gene expression to DNA
replica-tion is lesscertain. Although intermediate gene
expression begins between6and8 hafter
infec-tion (about thesametime thatviral DNA
repli-cation begins), evidence of IX and IVa2 gene
expressionhas been foundincells infected inthe
presenceofcytosine arabinoside, aninhibitorof
DNAreplication (24, 30). Itiscertain, however,
that theexpression ofIXandIVa2mRNAis at
itsmaximum levelatlate timesafter viral DNA
replication has begun (24, 38, 42). Because all
fiveearly transcription units (28) and the major
latetranscription unit (2, 29, 35)areactiveby3 h
postinfection, while the IX transcription unit is
inactive atthis time (42), the IXpromoter may
be the last viral promoter to be activated. The
exact times of activation of the other known
viral promoters, IVa2 and the alternative E2
promoter at map position 72, are notknown.
The goal of this work was to determine the
relationshipbetween viral DNAreplication and
intermediate gene expression, focusing on the
geneforproteinIX. Thefirstsetofexperiments
was designed to determine how hydroxyurea
(HU) blockade ofDNA synthesis affects
inter-mediate gene expression. HU was chosen
be-cause at10 mM itblocks detectable DNA
syn-thesis,does notbreak downduringthe courseof
the experiment (39), and does not significantly
affect RNAsynthesis. Thesecondsetof experi-mentsexamined thebasis ofthedependence of
IXgeneexpressionupon viral DNAreplication.
737
on November 10, 2019 by guest
http://jvi.asm.org/
738 CROSSLAND AND RASKAS
E1 B EIA
1.02 kbi-' A M,l
.88 kbi 'A .4
1.03kkbbIV
1 k.52kb_,' "
*.49kbl
-rIZ2
''. 161kbiQ
Ad2
dl313
Hpa E Hind m G
C2 cl
---4C1
HindM C
Sma F
FIG. 1. Map positions of Ad2 left-end mRNAs (6, 9, 21) and DNA fragments cloned foruse as probes. Spliced-out regions of mRNAsareindicatedby dashed lines, and theapproximatesize in kbof each mRNA is given. ML is the major late promoter (13, 44). The hashed box indicates theregiondeleted indl313 (18). All DNA fragments were inserted into pBR322, with the exception of Sma F whichwasinserted into M13 phage, andwere
used asrecombinant DNAs. The M13-Sma F recombinant used released the left strand of Sma F in phage particles.
MATERIALSAND METHODS
Cells and virus. Cells were grown as monolayer
cultures in Dulbeccomodified Eaglemedium (DME)
containing10%calforhorseserum(KC Biologicals)
for KB or 293 cells, respectively. Cells were split 2 days before theexperiments,tobeapproximately 70%
confluent when infected. Virus stockswereprepared
asdescribed (37). tsl25 virus wasobtained from Jim Williams, d1313 was obtained from TomShenk, and Ad2+ ND5wasobtained from Andrew Lewis.
Infectionconditions. Cells for infectionweregrown in 150-mm dishes. Themediumwasaspirated off,and the monolayers were washed with serum-free DME. Virus was added in 5 ml of DME per plate, and absorptionwascontinued for1 hwithgentleagitation every 15 min.Theinoculumwasremoved, and 30 ml of DMEcontaining10%serumand, insomecases, 10 mMHUwasaddedtoeachplate.
Thesuperinfection protocolwasidenticaltothatfor
infection, except thata45-min absorptionwasused. Greatcarewasneededtoavoiddetachingthe 293 cells whenadding thesuperinfection inoculum and overlay medium.
Recombinant DNAprobes. The left strandof the Ad2
SmaF fragment (map units 11.5-18.5) cloned in the
single-strandedbacteriophageM13 wasobtained from MarkBoguski. The HpaE (map units 0-4.4), HindIII C (7.9-17.0), EcoRI C (89.7-100), HindIll H (72.8-79.9), andEcoRIFl (72.8-75.9)fragments of
adenovi-rustype2(Ad2) cloned in pBR322wereobtained from Beth Ladin, Elizabeth Slattery, Michael Tigges, D.
Huang, and Richard Rosenthal, respectively. The
HindIIl Cl (11.5-17.0) and C2 (7.9-11.1) fragments
were obtained by digesting the original HindIII C clone with SmaI, which cuts at 11.1 and 11.5. The linear DNAwasthendigested withasecond enzyme
that cutstheplasmid sequenceneartheinsert.EcoRI
digestionwasusedtoexcise the11.5-17.0region,and
BamI digestion was used to excise 7.9-11.1. The
DNAs werethen recircularized with T4 DNAligase and usedtotransform Escherichiacoli HB101.
Pulse-labeling RNA. Topulse-label cells, the
medi-umwasaspirated off and replaced with 8 ml per plate of DME with serum,containing300,uCi of13H]uridine
permland, whereappropriate,10mMHU.After 5or 10minof incubation, the plateswereplacedonice, the labeling solution was removed, and the cells were scraped into coldphosphate-buffered saline.
RNA isolation. Cytoplasmic, polyadenylic acid
[poly(A)]-containing RNAwas isolatedby Nonidet
P-40leakage in isotonic buffer, extraction of the post-nuclear supernatant with phenol-chloroform-isoamyl
alcohol, andoligodeoxythymidylate-cellulose chroma-tography, allasdescribedpreviously (25).
Totalcell RNAwasisolatedbyanadaption of the procedure of Sauerbrier and Brautigam (32). Harvested cells were pelleted from coldphosphate-buffered
sa-line and taken up in1 mloflysisbuffer (1% sodium dodecyl sulfate, 1 mM EDTA, 0.1 M NaCl, 0.1% diethyloxydiformate, and 10 mM Tris, pH 7.5). The mixturewasvortexedvigorously,heatedat85°C for 30
sand cooledonice.An8-mlvolumeof8.5molal CsCl inlysis buffercontaining 0.1% sodium dodecyl sulfate
was added, and the solution was pipetted several times. Thelysed cell mixturewaslayeredover2.5ml of 9.5 molal CsCl inanSW41 tube andcentrifugedat
26,000 rpm for 22 h at 8°C in an SW41 rotor. The
supernatantwas aspirated, and the RNA pellet was
suspended inwaterandprecipitated bytheaddition of sodiumacetate to0.2Mand 2.5volumes of ethanol.
RNA blots. RNA samples for Northern analysis were incubatedwith 1 Mglyoxal in 10 mMNa2PO4 (pH6.8)at50°C for1 h(26) and thenelectrophoresed
in 1.1 or1.4%agarosegels.Inall cases,RNAfroman
equivalentnumberof cellswasloadedoneachlaneof
agivengel.The RNAsweretransferredto nitrocellu-lose as described by Thomas (40) and hybridized to
nick-translatedDNAprobes.
RNAgradients. Pulse-labeledRNAforsucrose gra-dientfractionationwastakenupin 250,ulofasolution containing80%formamide,2 mMEDTA,and40mM
PIPES [piperazine-N,N'-bis(2-ethanesulfonic acid)]
(pH7.4). The mixturewasheatedat68°C for7.5min
i
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.488.109.399.74.222.2]and layered on a preformed gradient of 5 to 20%
sucrose in the same buffer solution. The gradients werecentrifuged in an SW41 rotor at 30,000 rpm for 48 hat 4°C, and350-,ul fractions were collected from the bottom of the tube. The RNA in each fraction was precipitated with ethanol, taken up in buffer, and partiallyhydrolyzed with alkali as described by Nev-ins (27). Selected fractions were then hybridized to DNA bound tonitrocellulose filters as described previ-ously (25).
DNAblots. To parepare DNA forSouthernanalysis, nuclei were isolated as described (16) andlysedby the addition of DNA extraction buffer (0.1 M NaCl, 10 mMEDTA,0.5% sodium dodecyl sulfate, 50 mM Tris, pH 8.5). The mixture was digested with 1 mg of pronase B per mlovernight at 37°C, extracted with phenol-chloroform (1:1) three times, and precipitated withethanol. The DNA was taken up in anappropriate buffer, digested with the restriction enzyme KpnI, and electrophoresedon a1% agarosegel. The DNA bands were transferred to nitrocellulose by the method of Southern (36) and hybridizedto nick-translated Ad2 DNA.
RESULTS
Time course ofintermediate gene expression.
Asa firststeptoward studying their regulation,
the time of appearance of the intermediate
mRNAs was examined. To determine the
steady-state concentration of the mRNAsinthe
cytoplasmasinfectionprogressed,
poly(A)-con-tainingcytoplasmicRNAwasisolated from2 x
107 KB cells infected for4, 6, 8, or 10h. The
RNA was subjected to electrophoresis in an
agarose gel and then transferred to
nitrocellu-lose. The Ad2 HindlIl C fragment (7.9 to 17.0
mapunits), cloned in pBR322,was
nick-translat-ed and hybridized to the RNA blot. The three
intermediate mRNAs that hybridize to this
probe, the 1.03-kb Elb mRNA and the IXand
IVa2 mRNAs,werefirstdetected by this
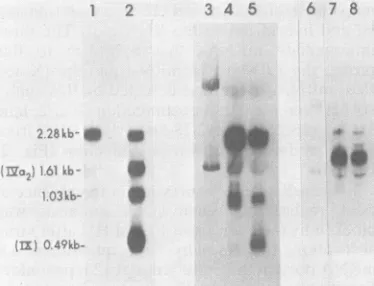
analy-sis between6and 8hpostinfection(Fig. 2, lane
3). As expected, the 2.28-kb early mRNA from
Elb was detected at 4 h postinfection (Fig. 2,
lane 1).
Intermediategene expression in theabsenceof DNA replication. When DNA synthesis was
blockedby theaddition of 10mM HUafter virus
absorption, the normally large quantity of IX
mRNApresentin 2 x
107
cellsat12hpostinfec-tionwasundetectable(Fig. 2, lanes 8 and 9).In
contrast, the 2.28-kb and 1.03-kb Elb mRNAs werebothpresent in HU-treated cells,although
the1.03-kbspecieswasreduced inquantity.The
HindIlI C2probe (7.9-11.1)used inthis
experi-mentdoesnothybridize IVa2 mRNA. When the
infectionwascontinued for15h and theHindIII
Cprobe (7.9-17.0)wasusedin thehybridization
analysis,neither the IX nor theIVa2 mRNA was
detected inHU-blockedcells(Fig.2, lanes 5and
6). Similarexperiments(not shown) suggest that
the 0.52-kb Ela mRNA has the same time
course and resistance to HU blockage as the
1.03-kb Elb mRNA. Although the time course
of appearance of the four intermediate mRNAs was the same and thelevels ofthe0.52-kb Ela
and 1.03-kbElb mRNAs were reduced by HU,
only the IX and IVa2 mRNAs were dependent
uponviralDNAreplication for expressionin the
cytoplasm.
Activation of IX and IVa2expression is inde-pendent of template number. At 12 h
postinfec-tion, cells infected in thepresence ofHU
con-tained fewer templates for viral transcription
than control cells infected without drugs. To
ensure that this difference in template number was notresponsible forourinabilitytodetectIX
mRNAinHU-treated cells, infectionconditions
weredetermined that would place
approximate-ly thesamenumber oftemplates in the nuclei of
thecontrol andHU-treated cells. Fourplates of
KBcellswereinfectedat amultiplicityof
infec-tion (MOI) of 10, and at various times after
infection nuclear DNA was isolated. A fifth
plate was infected at an MOI of 500 in the
presence of HU, and at 10 h postinfection its
nuclear DNA was isolated. Equal amounts of
these DNA samples were bound to
nitrocellu-lose filters andhybridized tosaturatingamounts
3
L 3 5 6
S
FIG. 2. Time course of intermediate mRNA
expression. Cytoplasmic, poly(A)-containing RNA was isolated from cells infected for 4, 6, 8, or 10 h
(lanes1through4), for 12 h(lanes8 and9),orfor 15 h (lanes5and6). Lanes5and9arefrom cells infected in
the presence of10 mM HU. Lane 7is a 14C-labeled
rRNA marker. The mRNAswere electrophoresedin
an agarose gel, transferredto nitrocellulose, and hy-bridizedto32P-labeledHindIII C(lanes1through 7)or
HindIII C2(lanes 8 and 9)DNA. Theadditional band
comigrating with the 18SrRNAmarker inlanes3, 4,
and 5 is occasionally seen with several Ad2 DNA
probes.Presumably, it results fromRNAaggregation
due to rRNA contamination of poly(A)-containing
RNAandsubsequent overloading of thisregion ofthe
gel. VOL 46,1983
-3, 61 -r
i
;-I .k,
a
06.
-OW.
a
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.488.250.441.376.527.2]- 21
o 18
z
0
0 .
HOURS POST INFECTION
FIG. 3. Quantitation of Ad2 DNA in infected cells. The amount of Ad2 DNA per cell was determined by binding nuclear DNA from 3.3 x104infected KB cells tonitrocellulose andhybridizingto saturatingamounts of3H-labeled Ad2 DNA. Under identical conditions, 3.9x 10-3jigoffilter-bound Ad2 DNA (the equivalent of 9.4 x 10'Ad2 DNA molecules) hybridized 24,400 cpm.Itwasconcluded that each 8.6 cpm hybridized to the experimental filters represented one Ad2 DNA molecule per cell. Symbols: 0,DNAfrom cells infect-edatanMOIof 10;0,DNAfrom cellsinfected at an MOI of 500 in the presence of HU.
of3H-labeled Ad2 DNA (Fig. 3). Theopencircle
inthe center ofFig. 3 indicates the number of
adenovirus DNA moleculespercellat10 hafter
aninfection at 500 MOI in the presence of HU. Theamountof intranuclear Ad2DNApresent at
12 h after a10 MOI infection without drugs is
approximatelyequaltotheamountpresentafter
infection at an MOI of500 in the presence of
HU.
Basedontheresults described above,
experi-ments were performed to determine the
influ-enceoftemplatenumberonIXandIVa2
expres-sion. KB cells were infected at an MOI of10
withnodrugor at anMOI of500in thepresence
of HU.At12 hpostinfection, RNAwasisolated
for blot hybridization analysis. When ablot of
cytoplasmic,poly(A)-containingRNAfrom2 x
107cellswas hybridized to the HindlIl Cprobe
(7.9-17.0), the IX and IVa2 mRNAs were only
detected indrug-free control cells, whereas the
Elb 2.28-kb and1.03-kbmRNAsweredetected
in control andHU-treated cells (Fig. 4, lanes 1
and2) (the 1.03-kb bandwastoofaint to
repro-duce in the
1photograph).
When total cell RNA from 2 x 10 cellswashybridizedtotheHindIIIfragment C2 (7.9-11.1) (Fig. 4, lanes 4 and5) a
similar resultwasobserved,except that the level
ofthe 1.03-kb mRNAwas relatively greater in
total cell RNA. Hybridization ofthe total cell
RNA blots to the EcoRI C (89.7-100) (Fig. 4,
lanes 7 and 8) probe demonstrated that the
predominant earlymRNAsfromregionE4were
present in equal or greater amounts in the HU-treated cells.
Newlysynthesized IX and IVa2 mRNA can only be detected when viral DNA replication occurs.
Regulation ofmRNAlevels in the cytoplasmcan
occur atseveral levels (11).Todetermine
wheth-erthe absence of IX and IVa2 mRNA in
HU-treatedcellswasbecause of lack of transcription
of these genes, newly synthesized molecules
were quantitated after a briefpulse label.
Be-causethe entiresequenceofIX mRNAisshared
with the Elb transcript, no DNA probe can
specifically hybridize onlyIX mRNA. To
quan-titate IXtranscription it was therefore necessary
to size-fractionate IX mRNA and the Elb
mRNAs before hybridization. KB cells were
again infected at10 MOI without drugs or 500
MOI in thepresence of HU. At12 h
postinfec-tion, when the numberof viral DNAtemplates
in thetwo setsof cellswasequal,thecellswere
pulse-labeled with [3H]uridine for10min. Total
cell RNA was isolated and fractionated by su-crosegradient centrifugation.Tolocatethe frac-tions containing mRNAs of interest, samples of
fractions 9 through 25 were electrophoresed in
anagarosegel, transferredtonitrocellulose, and
hybridized to 32P-labeled HindIII CDNA
(7.9-17.0). Transcription activity was assayed by
hybridization of gradient fractionsto
nitrocellu-lose-bound HindlIl C2DNA(7.9-11.1)to assay
Elb and IX, or SmaF left-strand DNA
(11.5-1 2 3 4 5 6 7 8
L;')5kb *
03D S
FIG. 4. Effect of HU on early and intermediate
mRNA accumulation. KB cells were infected at an
MOI of500 inthe presenceof HU(lanes 1, 4, and 7)or at an MOI of 10 without drugs (lanes 2, 5, and 8).
Twelve hours postinfection, cytoplasmic, poly(A)-containing RNA (lanes 1 and 2) or total cell RNA (lanes 4-8)wasisolated,electrophoresedinanagarose gel, and transferredtonitrocellulose. TheRNAblots
werehybridizedto32P-labeled HindlllC(lanes1and 2), HindIII C2(lanes3through5),orEcoRI E(lanes7
and 8) DNA. Lanes 3 and 6 are 14C-labeled rRNA markers.
VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.488.75.221.61.206.2] [image:4.488.260.447.418.561.2]18.2) to assay IVa2 (Fig. 5). In both RNA
gradients,asignificant peak of [3H]RNA, about
22Sin size, hybridizedto the HindIlI C2 probe
(7.9-11.1). This peak represents either the Elb precursor (2.36 kb) or the 2.28-kb message, or both. The absence of apeak corresponding to
the 1.03-kb mRNA in either gradient suggests
that 10min is insufficient time for labeling and
splicing ofasignificantamountof theprecursor
to occur.Transcriptionfrom the IX gene,
repre-sentedby the 9S peak (see Fig. 5), and the IVa2
gene, represented by the 17S peak, was not
detectable after a 10-min pulse unless DNA
replicationwasallowed. Similarly, aftera5-min
[3H]uridine pulse, transcription from the IX
gene was not detected unless DNA replication
was allowed (Fig. 6). Other pulse-labeling
ex-periments (not shown) havedemonstrated that
IX mRNAcontinuestobe transcribed if HU is
added after viral DNA replication has begun.
Weconclude that, when viral DNAreplication is
blocked with HU,transcriptionofIXandIVa2 is
reduced atleast90% and isverylikely absent.
Development of asuperinfection protocol.
Su-perinfection experiments were used to
deter-mine how IX transcription is related to viral
DNAreplication. Activation ofthe IX promoter
might be dependent upon trans-acting, viralor
cellular, regulatory factors that are produced
onlyasthevirusbeginsDNAreplication. If this
is thecase, anunreplicated viralgenome
enter-ingapreviously infected, late-stage cell should
begin transcribing the IX gene. Alternatively,
the IX promoter might require activation by a cis-acting change in the template itself during
replication. In this case, a superinfecting viral
genomewould first haveto bereplicatedbefore
it could act as a template forIX transcription,
even if it had entered a late-stage cell. To
distinguish between these possibilities, we
per-formed a series ofsuperinfection experiments
utilizingadenovirus deletion and insertion
mu-tants.
Inall of thesuperinfection experiments, d1313
wasused as the preinfecting virus. d1313 is an
AdSdeletion mutantlackingpartofEl,
includ-ing the IXpromoter(18). The AdS-transformed
human cell line 293 contains integrated AdS
DNAfromregion El (1, 17); d1313 canlytically
infect 293 cells because themissing El functions
areprovided bythe hostcell.However, because
theviralIXpromoterisdeleted and the
integrat-ed IX gene cannot beactivatedbyinfection, no IX mRNA or protein is produced during the
infection(37).
It wasessentialthatd1313-infectedcells be in thelatestageof infectionand thus beproducing
any factors that might be necessary for IX
transcription before superinfection. To
deter-minewhen the late stage begins,d1313-infected
A
--2.28kb
am -1,61blib2)
0 - 1 03kb
* -0.49kb1(E)
8
'-4
x U I
2
28s 18s
I 4s
20 FRACTION NUMBER
30
B
s -2.28kb
-103 kb
T0
x
C-I.
18s
10
4S
FRACTION NUMBER
FIG. 5. Newly synthesized IX and IVa2 mRNA. Cellsinfectedat anMOIof 10 withoutdrugs(A)or at anMOIof 500 in the presence of HU (B)were
pulse-labeled for 10 min with [3H]uridine. Total cell RNA was isolated and fractionated on sucrose gradients, andfractionswerehybridizedtonitrocellulose-bound HindIII C2 DNA (0)or SmaFleft-strand DNA(0).
Thelocations ofthe28S, 18S, and 4S RNA markers in the gradient are indicated. The insets show agarose gels of fractions9through 25, transferredto nitrocellu-lose andhybridizedto32P-labeledHindIIl CDNA.
293 cells were pulse-labeledwith
[3H]thymidine
at various times after infection. DNA, isolated from cellsfromeach timepoint, washybridized
tonitrocellulose-boundAdSDNA(Fig.7). Viral
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.488.247.440.72.502.2]A
F
B
20 21 22 23 2425 26I
03kb-20 21 22 23 24 25 26
1.61kb- so
i
03kb-3
49kb--'A,,R
..t... ..rs
il_
FIG. 6. Newly synthesized IX mRNA. Cells were infected at 500 MOI in the presence of HU (upper panels)
or at10MOI in the absence ofdrugs(lower panels) and were pulse-labeled for 5 min with[3H]uridine.Total cell RNA wasfractionated on sucrose gradients, and fractions were hybridized to nitrocellulose-boundHindlllC2 DNA(A). RNAfromfractions 20 through 26 was electrophoresed in an agarose gel, transferred to nitrocellulose, andhybridized to32P-labeled HindIlI C DNA (B).
DNAreplicationin cells infected withdl313and
the Ela deletion mutant d1312 began at 12 h
postinfection, 6 h later than in wild-type
Ad5-infected cells.
We next demonstrated that prior infection
with d1313 can provide functions needed for a
superinfection toproceed. For this experiment
we utilizedd1313and, as asuperinfectingvirus,
the temperature-sensitive AdS mutant ts125.
ts125 codes for a temperature-sensitive
DNA-binding protein and cannotreplicate viralDNA
at40°C(12,43). Mock-infectedand
d1313-infect-ed293 cells wereshifted from37 to40°Cat13 h
postinfection and superinfected with ts125.
Southern blot analysis (36) ofthe viral DNA in
these cells revealed that mock-infected,
ts125-superinfected cells contained only about as
muchts125 DNAafter 12 h at40°Cas asimilar
preparation blocked with HU (Fig. 8, lanes b
and e). Little viral DNA replication had
oc-curred, and the bands observed were
presum-ably due mostly to parental ts125 DNA.
dl313-infected, tsl25-superinfected cells containDNA
from both viruses.Toobserve thereplicationof
only the superinfecting virus by Southern blot
hybridization, amarker specific for ts125 DNA
wasrequired. Because of the deletion in d1313,
the HindIII G fragment present in Ad5 (and
tsl25) is missing. Prior infection with d1313
allowed significant productionof ts125 DNA in
12h at40°C,asevidencedby thedensity of band
G in lane a versus lane b ofFig. 8. A similar
resultwasobtained whents125wasallowed24 h
toreplicate(Fig. 8,lanes candd).These results
prove that the superinfectingts125 DNAenters
the nucleus, where an early protein coded by
d1313 cancomplement its missingfunction.
Only replicated templates can express IX
mRNA.Thesuperinfection protocolwasutilized to assay IX expression from unreplicated and
replicated templates in a late environment. In
addition toIX, the superinfecting virus should
code foran early RNA (one that is expressed
prior to viral DNA replication) that can be
J. VIROL.
.1 .4
-4.
ell"---k
)6".,
a,4,
cl;
E 'i... r, I t-I
-L,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.488.75.429.60.370.2]ment.Through analysis of the Ad2-SV40 hybrid mRNAs, one can independently confirm that the superinfecting template is transcriptionally ac-tive for anearly promoter. Ad2+ND5 replication was monitored in d1313-infected,
Ad2+ND5-su-perinfected cells by [3H]thymidine
pulse-label-ing and hybridization of cellular DNA to SV40
DNA. Replication of the superinfecting viral
DNA was just detectable at 5 to 7 h
post-superinfection (datanotshown).It wasexpected
that asuperinfectionallowed toproceed for 5 to
7 h before the addition of HU would produce
Ad2+ND5-coded mRNAs whose expression
was dependent upon template replication.
To perform the superinfection experiment,
fourplates of 293 cellswereinfected withd1313,
5
3 6 9 12 15 18
HOURS POST INFECTION
FIG. 7. Viral DNAreplicationin deletion
mutant-infected cells. Infected cells were pulse-labeled with [3H]thymidine for 1 h atthe times indicated. Their
DNA was isolated and hybridized to
nitrocellulose-bound AdS DNA, and the quantity ofhybridizable [3H]DNAwasdetermined. WT, Wild-type virus.
detectedagainstabackground of d1313 mRNAs. Ad2+ND5 is a nondefective derivative of Ad2 carrying a 1.3-kb insertion of simian virus 40
(SV40)DNA in early region 3 (E3) (15, 20, 23).
Early in Ad2+ND5 infection, mRNAs of 1.5 and 2.3 kbweredetectedby RNA blot hybridization withSV40 DNAas aprobe (Fig. 9, lane a). The 2.3-kb mRNA hybridized to HindIl H DNA (72.8-79.9) (Fig. 9, lane e) but notto EcoRI Ft DNA(72.8-75.9) (lane d). Thissuggeststhatthe 2.3-kb mRNA originates at theE3 promoter at
76mapunits. Thestructureof the 1.5-kb mRNA is unknown. Late in AD2+ND5 infection, mRNAsof3.1, 3.5, 4.0, and 6.8 kb hybridizedto
SV40 DNA (Fig. 9, lane b). These species also hybridized to the EcoRI Ft DNA (72.8-75.9) (lane c), suggesting that they result from fusion of late region L4 mRNA bodies with SV40 sequences.
The superinfection protocol consists of infec-tion of 293 cells withd1313, followed by superin-fection with AD2+ND5 in thepresence of HU. Thisprocedure allows observation of the ability
of the unreplicated, superinfecting template to
transcribe the IXgene while in a late
environ-__I A
fe
J.
;,
3
H.Ciz!R S
m2 C4 Xr b c de f ' ' c
i.
Lm a
- - -
-37
-A
-B
-D
--Fp
G
H
me 4 go -I
FIG. 8. d1313 preinfectioncan complement
defec-tive early function ofsuperinfecting ts125. Line 293 cells were preinfectedwithd1313 (aandc) or
mock-preinfected (bandd).After 13 hat37°C,the cellswere
shiftedto40°C, superinfectedwithts125,and incubat-ed for 12 h(aandb)or24 h(candd)more.For(e),
cellswereinfected with ts125at40°Cin thepresence
of HU and incubated for 12 h. For (f), cells were
infected with ts125 at 33°C and incubated for 24 h. DNA from each sample was digested with KpnI, electrophoresedina1%agarosegel,and transferredto nitrocellulose. KpnI-digested d1313, Ad2, and Ad5 DNAswereincludedasmarkers. The DNA blotwas
hybridizedto32P-labeled Ad5 DNA.
0
x
U
I
(V)
15r
dl 313 10j
x
220
I 15
en,
dl 312
10 46,
r,
L
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.488.55.214.56.341.2] [image:7.488.245.445.264.547.2]65 70 5 79 86
___> 23kb 3kb
abcd e~~~~~~~O
E->oS;RI
6.84b
43Okb235kb
-
1.5kb-expression detected (Fig. 10, lanes c and d). The
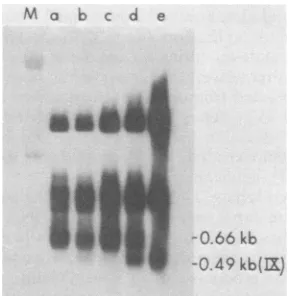
0.66-kbmRNAhybridizingtheHindIII Cprobe
is transcribed from d1313 and is a fusion product of Ela and Elb sequences(37).
Hybridization with the SV40 probe detected low levels of the 2.3- and 1.5-kb early hybrid mRNAs from the superinfecting virus, even
when DNA replication was blockedimmediately aftersuperinfection (Fig. 10, lane a). A low level of the early hybrid mRNAs is to be expected after superinfection into late-stage cells, as E3 promoteractivity declineslatein infection(28). The 3.1-, 3.5-, 4.0-, and 6.8-kb late hybrid mRNAs were detected only when more than 3 h wasallowed for the superinfecting virus to enter andreplicate (Fig. 10, lanes c andd).
The results presented above demonstrated that IX mRNA was expressed only in cells in which the superinfecting viral DNA had
replicat-FIG. 9. Ad2+ND5 hybrid mRNAs. The hatched boxdepicts the SV40 insert in Ad2+ND5. Numbers above the lineareAd2 mapunits;those below the line areSV40mapunits.Proposedstructuresof thehybrid
mRNAs(except 1.5kb)aregiven, alongwiththe map locations of the Ad2 DNAfragmentsclonedforuse as
probes. The unbroken lines represent Ad2 sequences, the dashed lines indicate spliced-out sequences, and the open boxes represent SV40 sequences. The four
largermRNAbodies areproposedtobejoinedtothe Ad2 late tripartite leader (4, 10, 22). Line 293 cells wereinfected withAd2+ND5 foi 4 h (a,d, e)or15h (b, c). Their cytoplasmic, poly(A)-containing RNA was isolated, electrophoresed in anagarosegel, and transferred to nitrocellulose. The RNA blots were
hybridizedto32P-labeledSV40DNA(a,b),EcoRI Fl
DNA(c,d),orHindlll H DNA (e).
and four were mock-infected. At 13 h
postin-fection, the cells were superinfected with
AD2+ND5. One plate was changed to
HU-containing mediumat0h,anotherat3 h,anda
thirdat6hpost-superinfection;all were
harvest-ed at 9 h. Cytoplasmic,poly(A)-containingRNA wasisolatedfromeachplate, electrophoresed in anagarosegel,andtransferredtonitrocellulose.
Duplicate RNA blots were hybridized to
nick-translated HindIII C (7.9-17.0) DNA or SV40
DNA (Fig. 10 and 11). The0.49-kb IX mRNA
was notdetectedby theHindlIl Cprobe when
DNAreplicationwasblockedimmediately after
superinfection (Fig. 10, lane a) or when it was
blocked 3 h post-superinfection (lane b). Only
when more than 3 hwasallowed forthe
superin-fecting virus to enter and replicate was IX
I_.
..~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.
__.... ...7
ricurs
h o le, -2
*U
:...4
"A o b t-- d
.s ,kE
....
_ 4i-k b
SF _ 3 kb 3 i kb
-x13-rE - 5.Kb
[image:8.488.65.236.76.321.2]-R:_,!. ' De.v-NE PR
FIG. 10. Expression ofIXmRNArequires
replica-tion. Line 293 cellswereinfected withd1313(athrough d)and13hlaterwere superinfected withAd2+ND5. The cellswerechangedtomediumcontaining10 mM HU at thetimes indicated (dashed lines indicate the presence ofHU) andwereharvested 9hafter
superin-fection. Cytoplasmic, poly(A)-containing RNA was
isolated, electrophoresed inanagarosegel,and
trans-ferredtonitrocellulose. TheRNAblotswere
hybrid-izedto32P-labeledHindIll C DNAorSV40 DNAas
indicated. Autoradiogram of the hybridized blot. M
denotes14C-labeledrRNAmarkers.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.488.260.447.295.563.2]A
nJ
0-d
fll
B D D
a1
b 1\
ARV
post-superinfection, the poly(A)-containing RNA was isolated from
107
cellsfor analysis as before. Hybridization with HindIII C probe (7.9-17.0) revealedIX mRNAfromthe 10-MOIand 100-MOI, no-drug superinfections (Fig. 12,
lanes a and b), but no IX mRNA from the 10-MOI and 100-10-MOI plus HU superinfections (lanes a and c).Despitea10-fold greater input of superinfecting virus, expression of IX mRNA could not be detected in the absence of DNA replication.
DISCUSSION
This paperdescribesexperiments designedto test the effect oftemplate replication upon the
expression of Ad2 intermediategenes. All four
intermediate mRNAsassayed appearedat6 to 8
hpostinfection, a timecourse ofexpression in
agreementwithprevious work(30, 38).
We have compared the steady-state levels of
the IX andIVa2mRNAs in cellsreplicatingviral
DNA and cells blocked in DNAsynthesis.IX or
IVa2 mRNA was detectable by RNA blot
hy-bridizationonlywhen viral DNAreplicationwas
allowed. Densitometer scans of the
autoradio-graphs from these experiments were used to
quantitate the reductionin IX and IVa2 mRNA
levels. Inthree experiments in which both sets
of cells were infected at the same MOI, HU
treatment reduced the steady-state level of IX
IM a b c d e
c
d
FIG. 11. Densitometer scansof lanes athrough d ofFig.10,hybridizedtoHindIlI C DNA (A) andSV40
DNA(B).
ed. Duetoreplication,those cells also contained
a larger number ofpotential templates for IX
transcription. To demonstrate that the
replica-tion-dependent regulation of IX transcription is
independent oftemplatenumber, the amount of
superinfecting virus was varied 10-fold.
d1313-infected 293 cells were superinfected with
Ad2'ND5at10 or 100MOI. Half of each set of
cells wasincubatedin the presence of HU, and half was incubated without drugs. Nine hours
I
-0.66kb
[image:9.488.88.184.71.512.2]-0.49kb(2)
FIG. 12. Expression ofIX mRNA is independent of MOI.Line 293 cellswere infected withd1313 (MOI,
20; lanes athrough d)ormock-infected (e) and 13 h
later were superinfected with Ad2+ND5 as follows: lane a, MOIof10plus HU; laneb, MOI of 10 without HU;lanec, MOIof100plus HU; lanes d and e, MOI of100without HU. Nine hours aftersuperinfection, cytoplasmic, poly(A)-containing RNA was isolated, electrophoresed inanagarosegel, and transferred to
nitrocellulose. Each lane represents the material from
107cells. TheRNAblotwashybridizedto32P-labeled HindIIICDNA.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.488.269.413.415.563.2]mRNA an average of 800-fold and that of IVa2 mRNA an average of 350-fold. Underinfection
conditions that resulted in equal numbers of
viral templates in both sets of cells, HU
treat-ment reduced the steady-state level of IX
mRNA500-fold and that ofIVa2 mRNA50-fold.
Thesefiguresrepresent minimum values, as the
films wereprobably exposed beyond thelinear
response range in the no-drug lanes inan effort
to revealfaint bands intheHU-treated lanes.
We havealsocomparedthe rateof
transcrip-tion ofIX andIVa2 mRNAs in cells replicating
viralDNAand cellsblocked inDNA synthesis.
When equal numbers of viral templates were
presentin both sets of cells, newly synthesized IX and IVa2 mRNA could only be detected
when viral DNA replication was allowed.
Be-cause of the high backgrounds inherent in this
pulse-labeling experiment,we can saywith
cer-tainty only that the rate of transcription of IX
and IVa2 mRNAs is reduced 10-fold in the
absenceof viralDNAreplication. The
accumu-lated evidence suggests, however, thatno IX or
IVa2 mRNAis produced when viral DNA
repli-cation is blocked.
Other investigators have observed some IX
protein (30)orIVa2 mRNA (24) in cellsinfected
inthe presence of the DNA synthesis inhibitor
cytosine arabinoside, although the quantity of
these products present was not directly
com-pared tothat present in control cells. Cytosine
arabinoside has been reported to break down
during prolonged incubation (39). The IX and
IVa2 geneactivity observed by these
investiga-torscould haveresulted fromasmallamountof
viral DNAreplication due to cytosine
arabino-side breakdownduringthecourseof the
experi-ment. Alternatively, the observed activity could
have resulted frominefficient expressionofthe
IX and IVa2 genes in cells in which no viral
DNA replication had taken place. Our results
have demonstrated, however, that this
expres-sion isinsignificantly low.
Bacteriophage T4lategenes, like the Ad2 IX
gene, areexpressed only after viral DNA repli-cation hasbegun. Investigations of T4late-gene
regulation suggest that bothphage-encoded
sol-uble geneproducts and aphysical changein the
template fortranscriptionarerequiredforphage
late-gene expression (31). The superinfection
experiments presented inthis paper demonstrate
thattheAd2 IX promoter cannot beactivatedon
aninfecting, parental template. Diffusible,
trans-acting factors produced late in infection with
d1313are notsufficienttoactivatethe IX gene on
aparentaltemplate.The possible objectionthat
d1313 does not produce the needed regulatory
factorsbecause of mutation is dismissed by the
observation thatd1312, whichwasderived from
the same parent virus and thus shoulddifferonly
in region El (18), produces IX mRNA and
protein wheninfecting 293 cells. BesidesIX,the
onlyEl mRNAmissing from d1313-infected293 cellsisthe0.52-kb mRNA from Ela (37), which is also absent fromd1312-infected 293cells.
Activation of theIX gene promoteris
proba-bly dependent upon a change in the physical
structureof the template during replication. An
alteration in the distribution or composition of
nucleosomes (33), exposing the IXpromoter to
the action of RNA polymerases, could be
re-sponsible for this change. Another possibility is
that soluble regulatoryfactors necessaryfor IX
promoter activity cannot bind to the parental
DNAbecause of the nucleosomestructure.
Dur-ingorafter replication, accessibilitytothe
bind-ing site of these regulatory factorson the
tem-plate is afforded by the perturbation of the
deoxyribonucleoproteinduringreplication. This
model of IX regulation resembles the stable,
active and inactivetranscription complexes
ob-servedby Bogenhagen etal. in theXenopus5S
RNA gene (7). A third possibility is that the
replicatedgenomes enter aspecific nuclear
com-partment,andonly in thatcompartment arethe
factorsnecessaryforIXtranscription available.
Further work will be needed to determine
which, ifany, of these models is correct.
Although IX and lateregions L2 through L5
are activated by the same event, viral DNA
replication,IXprotein is observed earlier during
infection than proteins encoded by L2 through
L5(14, 30).Thistime differencemayresult from
morerapidtranscription, processing, transport,
and translation ofthe small, unspliced IX
mes-sage(3).Also, the different mechanisms of
regu-lation-promoter activation for the IX gene,
versusantitermination and poly(A) site selection
for L2 through L5-may be responsible for
some of the time difference. The observation
that fiber protein from L5 isnot seen until2 h
after the appearance of hexon protein protein
fromL3(30) indicates thatlate-geneactivation is
complex andmayinvolvefactorsor eventsother
than DNAreplication.
Several characteristics of the IX and IVa2 genesdifferentiatethemfromthe 0.52-and 1.03-kb intermediate genesfrom El:(i) IXandIVa2
aretheonlyAd2transcriptionunits toproducea
single transcript and polypeptide, whereas the
other intermediates are membersof families of
mRNAspecies originating at the same
promot-er;(ii)IX andIVa2,andpossiblythe E2
promot-eratmapposition72,arethelast Ad2promoters
to be activated during lytic infection, whereas
the other intermediates derive frompromoters
active at early times; (iii) IX and IVa2 are
regulated by promoter activation, whereas the
other intermediates are regulated by mRNA
stability.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
ACKNOWLEDGMENTS
We thank thecolleagues who kindly provided recombinant DNAs for thisstudy. We alsothank EllenJohnson and Scott Hauser for technicalassistance,andBetsy Klein for typing the manuscript.
This work was supported by research grants from the American CancerSociety (MV-31H) and the National Cancer Institute (CA-16007). Cell culture mediawereprepared in a Cancer Centerfacility funded bythe NationalCancer Institute (CA-16217). This studywasalsosupported byBrown
William-son Tobacco Corp., Philip Morris, Inc., R. J. Reynolds TobaccoCo.,and the United StatesTobacco Co.
LITERATURE CITED
1. Aieio,L.R.,R.Guifoyle,K.Huebnpr,andR. Weinmann. 1979. Adenovirus 5 DNA sequencespresent and RNA
sequencestranscribed in transformed human embryonic kidney cells(HEK-Ad5or293). Virology 94:460-469. 2.Akusjarvi, G., and H. Persson. 1981. Controls of RNA
splicing and termination in the major late adenovirus transcription unit. Nature (London) 292:420-426. 3. Alestrom, P., G.Akusjarvi,M.Perricaudet,M. B.
Math-ews,D.F.Kiessig,and U. Pettersson. 1980. Thegenefor polypeptide IX ofadenovirus type 2 and its unspliced
messengerRNA.Cell 19:671-681.
4. Berget,S.M.,C.Moore,andP. A.Sharp. 1977. Spliced
segments atthe 5'terminusof adenovirus 2late mRNA. Proc. Natl.Acad. Sci. U.S.A.74:3171-3175.
5. Berk, A. J., F. Lee,T. Harrison, J.Williams, and P. Sharp. 1979. Pre-early adenovirus 5gene product
regu-lates synthesis of early viral messenger RNAs. Cell 17:935-944.
6. Berk, A. J., and P. A. Sharp. 1978. Structure of the adenovirus 2early mRNAs. Cell 14:695-711.
7.Bogenhagen,D.F.,W. M.Wormington, andD.D.Brown. 1982.Stabletranscription complexes of Xenopus 5S RNA
genes: a means to maintain the differential state. Cell 28:413-421.
8. Carter,T.H.,and H. S.Ginsberg.1976. Viral transcrip-tion in KB cells infectedby temperature-sensitive "early"
mutantsof adenovirustype5.J.Virol. 18:156-166.
9. Chow, L. T., T. R. Broker, and J. B. Lewis. 1979. Complex splicingpatterns ofRNAs from theearly regions of adenovirus 2. J. Mol.Biol. 134:265-303.
10. Chow, L. T.,R. E. Gelinas, T. R. Broker, and R. J. Roberts. 1977.Anamazingsequencearrangementatthe 5' ends of adenovirus 2messengerRNA.Cell 12:1-8. 11. DarneUl,J.E. 1982.Varietyin the levelofgenecontrol in
eukaryoticcells. Nature(London)297:365-371. 12. Ensinger,M.J.,and H. S.Ginsberg.1972.Selectionand
preliminary characterizationoftemperature-sensitive
mu-tantsoftype5adenovirus.J. Virol.10:328-339. 13. Evans, R., J. Weber, E. Ziff, andJ. E. Darnell. 1979.
Premature termination during adenovirus transcription. Nature(London)278:367-370.
14. Everitt, E.,and L.Philipson.1974.Structuralproteinsof adenovirus. XI. Purification of three low molecular weight virion proteins of adenovirus type 2 and their synthesis during productive infection. Virology 62:253-269.
15. Flint,S.J.1977. Thetopographyandtranscriptionof the adenovirusgenome.Cell10:153-166.
16. Goldenberg, C. J., and H.. J. Raskas. 1979. Splicing patternsof nuclearprecursorstothemRNAfor adenovi-rus2DNAbinding protein.Cell16:131-138.
17. Graham,F. L.,J. Smiley, W.C. Russell,andR. Nairn. 1977. Characteristics ofahumancell linetransformedby DNA from human adenovirus type 5. J. Gen. Virol. 36:59-72.
18.Jones, N., and T. Shenk. 1979. Isolationofadenovirus type5 hostrangedeletionmutantsdefective for transfor-mation ofratembryocells. Cell17:683-689.
19. Jones, N.,and T.Shenk. 1979. Anadenovirustype5early
gene function regulates expression of otherearly viral genes.Proc.Natl. Acad. Sci.U.S.A.76:3665-3669. 20. Kelly, T. J., andA.M.Lewis.1973. UseQf nondefective
adenovirus-simian virus 40hybridsformappingthesimian virus 40 genome.J.Virol. 12:643-652.
21. Kitchingman, G. R., S.-P. Lai,and H. Westphal. 1977. LoopstructuresinhybridsofearlyRNAand the separat-ed strands of adenovirus DNA. Proc. NatI. Acad. Sci. U.S.A. 74:4392-4395.
22. Klessig, D. F. 1977. Two adenovirus mRNAs have a common5' terminal leader sequence encodedatleast 10 kb upstreamfrom their maincoding regions.Cell 12:9-21. 23. Lebowitz, P.,T.J.Kelly,D.Nathans,T. N.H.Lee,and A. M. Lewis. 1974. Acolinear maprelating the simian virus 40(SV40) DNA segmentsof sixadenovirus-SV40 hybrids to theDNA fragments produced byrestriction endonucleasecleavage of SV40. Proc. NatI. Acad. Sci. U.S.A. 71:441-445.
24. Lewis,J. B.,H.Esche, J.E.Smart,B. W.Stillman,M. L. Harter, and M. B. Mathews. 1980. Organization and expressionof the left third of the genome of adenovirus. ColdSpringHarborSymp. Quant. Biol. 44:493-508. 25. McGrogan, M.,andH.J.Raskas.1977.Species
identifica-tion and genomemapping ofcytoplasmicadenovirus type 2RNAssynthesizedlateininfection. J. Virol. 23:240-249. 26. McMaster, G. K.,andG.C. Carmichael. 1977.Analysisof
single- and double-stranded nucleic acidson polyacryl-amide and agarose gels by using glyoxal and acridine orange.Proc. Natl. Acad.Sci. U.S.A.74:4835-4838. 27. Nevins,J.R.1980.Definition andmappingof adenovirus 2
nucleartranscription. MethodsEnzymol.65:768-785. 28. Nevins,J. R., H. S.Ginsberg, J.-M. Blanchard, M.C.
Wilson,andJ.E.Darnell.1979.Regulationof theprimary expressionof theearlyadenovirustranscriptionunits. J. Virol. 32:727-733.
29. Nevins, J. R., and M. C. Wilson. 1981. Regulation of adenovirus 2 geneexpressionatthelevel of transcription-al termination and RNA processing. Nature (London) 290:113-118.
30. Persson, H., U. Pettersson, and M. B. Mathews. 1978. Synthesis ofa structural adenovirus polypeptidein the absence of viral DNAreplication.Virology90:67-79. 31. Rabussay,D.P.,and E. P.Geiduschek. 1977.Regulation
of gene action in thedevelopmentoflytic bacteriophages, p. 1-196. In H. Fraenke-Conrat and R. Wagner (ed.), Comprehensive virology, vol. 8. Plenum Publishing Corp.,New York.
32. Sauerbrier,and A.R.Brautigam. 1970.Asimplemethod forisolatingRNAfrom bacteria. Biochim.Biophys.Acta
199:36-40.
33. Sergeant, A., M. A. Tigges, and H. J. Raskas. 1979. Nucleosome-like structural subunits of intranuclear pa-rentaladenovirus type 2 DNA. J. Virol. 29:888-898. 34. Sharp, P.A., J. Manley, A. Fire,and M. Gefter. 1980.
Regulation of adenovirusmRNA synthesis. Ann.N. Y. Acad.Sci. 354:1-15.
35. Shaw,A. R., and E.B. Ziff. 1980. Transcriptsfrom the adenovirus-2 major late promoter yield a single early familyof 3' coterminal mRNAs and five late families.Cell 22:905-916.
36. Southern, E. M. 1975. Detection of specific sequences amongDNAfragments separatedby gel electrophoresis. J.Mol.Biol. 98:503-517.
37. Spector, D. J., D. N. Halbert, and H. J. Raskas. 1980. Regulation of integrated adenovirus sequences during adenovirus infection of transformed cells. J. Virol. 36:860-871.
38. Spector, D. J., M. McGrogan, and H. J. Raskas. 1978. Regulation of the appearance ofcytoplasmic RNAsfrom region 1 of the adenovirus 2 genome. J. Mol. Biol. 126:395-414.
39. Thomas,G. P.,and M.B. Mathews. 1980. DNA replica-tion and theearlyto late transition inadenovirus infec-tion.Cell22:523-533.
40. Thomas,P.S. 1980. Hybridizationof denaturedRNA and
on November 10, 2019 by guest
http://jvi.asm.org/
748 AND RASKAS
small DNAfragments transferredtonitrocellulose. Proc. Natl. Acad.Sci. U.S.A. 77:5201-5205.
41. van derViet, P. C., A. J. Levine, M. J. Ensinger, and H.S. Ginsberg. 1975. Thermolabile DNA binding proteins from cells infected withatemperature-sensitivemutantof adenovirus defective in viral DNA synthesis. J. Virol. 15:348-354.
42. Wilson, M. C., and J. E. Darnel. 1981. Control of
messengerRNAconcentration by differential cytoplasmic half-life. AdenovirusmessengerRNAs fromtranscription
units IA and 1B. J. Mol. Biol. 148:231-251.
43. Wilson,M. C., N. W. Fraser, and J. E.DarneDl. 1979. Mapping of RNA initiation sites by high doses ofuv irradiation: evidence for three independent promoters
within theleft 11% of the Ad2genome.Virology 94:175-184.
44. Ziff, E. B., and R. M. Evans. 1978. Coincidence of the
promoter and capped 5' terminus of RNA from the adenovirus 2 major latetranscription unit. Cell 15:1463-1475.
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/


![FIG. 5.gelsTheCellsandantheloselabeledwasHindIII Newly synthesized IX and IVa2 mRNA. infected at an MOI of 10 without drugs (A) or at MOI of 500 in the presence of HU (B) were pulse- for 10 min with [3H]uridine](https://thumb-us.123doks.com/thumbv2/123dok_us/1445913.97017/5.488.247.440.72.502/gelsthecellsandantheloselabeledwashindiii-newly-synthesized-infected-drugs-presence-pulse-uridine.webp)
![FIG. 6.orandRNADNA at Newly synthesized IX mRNA. Cells were infected at 500 MOI in the presence of HU (upper panels) 10 MOI in the absence of drugs (lower panels) and were pulse-labeled for 5 min with [3H]uridine](https://thumb-us.123doks.com/thumbv2/123dok_us/1445913.97017/6.488.75.429.60.370/orandrnadna-newly-synthesized-infected-presence-absence-labeled-uridine.webp)
![FIG. 7.boundinfectedDNA[3H]DNA[3H]thymidine Viral DNA replication in deletion mutant- cells](https://thumb-us.123doks.com/thumbv2/123dok_us/1445913.97017/7.488.245.445.264.547/fig-boundinfecteddna-thymidine-viral-replication-deletion-mutant-cells.webp)