JOURNAL OFVIROLOGY,Dec. 1988,
p.
4499-4509 0022-538X/88/124499-11$02.00/0Copyright
C1988,AmericanSociety
forMicrobiologyVol. 62, No. 12
Characterization
of Cellular Factors That Interact with the Human
T-Cell Leukemia
Virus Type I
p4OX-Responsive
21-Base-Pair Sequencet
KUAN-TEH
JEANG,4*
IMRE BOROS, JOHN BRADY, MICHAEL RADONOVICH, AND GEORGE KHOURYLaboratoryof Molecular Virology, National Cancer Institute, Bethesda, Maryland 20892
Received 9 June 1988/Accepted 19 August 1988
Transcriptionalactivation of the humanT-cellleukemia virus type I(HTLV-I)long terminal repeat (LTR) by viral protein p40xrequires a 21-base-pair (bp) sequence which is repeated three times within the LTR. This sequence contains a core octanucleotide (TGACGTCT) which has been attributed to be a cyclic-AMP (cAMP)-responsive element. We demonstrate here that the HTLV-I LTR can be specifically stimulated by cAMP regulatorsand haveidentified four proteins in HeLa cells that bind to theHTLV-I21-bpsequence. We correlated the in vitro bindingandtranscriptional activity of one of these cellular factors
(Mr,
180,000) to the trans-activation oftheHTLV-ILTR byp40".Point mutations were generated within the cAMP octanucleotide of the HTLV-I 21-bp sequencethatsimultaneously abolished biological responsiveness to trans-activation byp4oX and to stimulation by cAMP. We found that these mutations also eliminated the binding of the 180-kilodalton HeLa factor tothe HTLV-I 21-bp element. In the absence of a demonstrable DNA-binding property for p40X, we hypothesize thatcellular proteins areinvolved, possibly through signal transduction pathways, in its trans-activation ofresponsivepromoters.
HumanT-cell leukemia virus type I (HTLV-I) has been
establishedastheetiologicalagent for one particular form of
adultT-cell leukemia (forareview, seereference19). At the 3' end oftheHTLV-Igenome is an open reading frame that
encodes, through a double-splicing event, a single
2.1-kilobase (kb)mRNAwhichcan bealternately translatedinto
three distinct proteins (p40x, p27X and p21x) (33, 47). A
similar open reading frame has also been described for HTLV-II (27) andbovine leukemiavirus (9, 42). InHTLV-I, thep4Ox protein,a40-kilodalton(kDa)polypeptide,has been
directly implicatedastheviralgene productresponsible for
positive
transcriptional trans-regulation of the promoterelement found within the HTLV-I long terminal repeat
(LTR)(14, 17, 53, 58).
TheHTLV-ILTRcontainsthreecopiesof animperfectly conserved 21-base-pair (bp) sequence (32, 51). Deletion analysis of the HTLV-I LTR and chimeric constructions
utilizing
LTR sequenceslinked toreporter genesillustratedthat two or morecopies of this 21-bpsequence,regardless of orientation, can serve as a sufficient target for p4OX
trans-activation (4, 50, 54). It has also been shown that some
cellulargenesare
transcriptionally
stimulatedby
p40xbutdo notcontain this21-bp element(8, 29).Thesefindings
suggest anabsence ofonespecific
sequenceexclusively
required
forp40X-mediated
trans-activation.Thistheme ofalack ofexclusivetarget sequence
require-ment has beensimilarly
described for other viraltrans-activators(for a
review,
seereference 34). The adenovirus Ela, the herpesviruses immediate-earlyproteins,
and theSV40large-Tantigenalltrans-activatemany differentgenes.
* Correspondingauthor.
tDedicatedtothe memory ofGeorgeKhoury.
tPresentaddress: Laboratory of Molecular
Microbiology,
Na-tionalInstitute ofAllergyand Infectious Diseases, Bethesda, MD 20892.Ela, for example, can transcriptionally induce genes as diverse as ratpreproinsulin(21), human
P-globin
(60),
simianvirus 40 (SV40) early promoter (61), and other adenovirus
promoters (3, 31). An equallylarge recitation can be
com-piled for the herpes and SV40 trans-activators. Since the
stimulated genes do notsharecommontarget sequencesfor induction,it isunlikelythattheseviralproteins
mediate
their transcriptional effects through a direct sequence-specific binding mechanism. Thus generalized cellular factors and pathways areprobablyinvolved in many viraltrans-activa-tion events.
It is possible, however, that
p40x
acts through a directinteraction withthe upstreamsequencesof
responsive
genes (19). Wetestedthispossibility and failedtodetectsignificant
binding of
p40x
to the HTLV-I LTR or evengeneralized
binding to neutralDNAs. Hence, weundertookto
identify
cellular
proteins
that mayinteractwith theHTLV-I LTRas a step toward a betterunderstanding
of theregulatory
properties of
p40x.
Thepresent study utilized aUV-cross-linking procedure to probe for
proteins
in HeLa cells thatbindto the
p40X-responsive 21-bp
element ofHTLV-I. We found four groups ofproteins
with molecularweights
of 76,000, 120,000, 180,000, and >200,000, as determinedby
sodium dodecyl
sulfate-polyacrylamide gel
electrophoresis
(SDS-PAGE), thatpreferentially
boundtotheHTLV-I21-bp
sequence.Results from
point
mutationsgenerated
within theHTLV-I 21-bp sequence suggest that the
binding
ofone ofthesecellular
proteins,
the 180-kDaspecies,
correlated with thetrans-activation function of
p40X.
These mutations alsore-vealed the importance of a
highly
conserved octamericsequence
(5'-TGACGTCT-3'),
shownin othersystemstobe a cyclic-AMP(cAMP)-responsive
element,
forp40x-medi-ated
regulation
ofthe HTLV-I LTR. We present evidencesuggesting
that similar target sequences may be shared within the HTLV-I LTR for activationby
bothp40x
and cAMP.4499
on November 10, 2019 by guest
http://jvi.asm.org/
MATERIALS AND METHODS
UV cross-linking of proteins to DNA probes. Synthetic
oligonucleotide probes were generated using an Applied Biosystems model 380A synthesizer. The DNAwaspurified accordingtothe manufacturer's instructions. A1-,ug portion ofoligonucleotide was nick translatedat 17°Cfor 90minin 10 mM Tris hydrochloride (pH7.5)-10mM MgCl2-100mM NaCl-1 mMdithiothreitol-250 puMdGTP-250,uMdATP-1.5 xuM [32P]dCTP (specific activity, 400 Ci/mmol; Amersham
Corp.)-250 ,M bromodeoxyuridine (BUdR)-triphosphate (Sigma Chemical Co.) by using5UofKornbergpolymerase (Boehringer Mannheim Biochemicals). Where necessary,
DNA was preincubatedwith 10-8g (finalconcentration) of DNase Iatroom temperature for 1 minto generate nicks. Afterradiolabeling, the oligonucleotides were analyzed by
gel electrophoresis and visualized by autoradiography to
verify that theyremained intact.
In theUV-cross-linking reactions, 50,ugofHeLa nuclear extract (11) was first incubated on ice in a 10-,ul reaction volume in buffer (20 mM HEPES [N-2-hydroxyethylpipera-zine-N'-2-ethanesulfonic acid], pH 7.9; 1.5 mM MgCl2; 40 mM KCl; 1 mMspermidine) containing2to4,ug of poly(dI-dC) (Pharmacia Fine Chemicals) for 15 min. Radiolabeled DNA probe (10 ng) wasadded, and incubation was
contin-ued foranadditional 15min. The entire reaction mixturewas
thentransferred ontoapiece of parafilm and irradiated with
a handheld UV lamp (Mineral Light model R-51) at a
distance of2 cm for 90 s. After irradiation, 10 pI of SDS
samplebuffer(50mM Trishydrochloride, pH 7.0; 2%SDS; 1.5 M 3-mercaptoethanol; 5% glycerol; 0.01% bromophenol blue) wasadded. The mixturewas heatedat68°C for5 min and thenelectrophoresed ina10%SDS-polyacrylamide gel.
Gel mobility shift assay. Gel mobility shift assays were
performed aspreviously described (16, 20). Inthe competi-tionexperiments, 1,ug ofunlabeled competitor oligonucleo-tide or 2 to 4 ,g of poly(dI-dC) was added to the HeLa proteinextract and incubated on ice for 15 min before the
addition of labeled probe. Electrophoresis of protein-DNA complexes was conducted in 4, 6, or 8%acrylamide gel in
0.25x TBE (lx TBE is 90 mM Tris hydrochloride plus 90 mM boric acid plus 1 mM EDTA [pH 8.3]) at a constant
currentof11 mA. In some experiments, the protein-DNA
mixturewas subjected toUVirradiation before electropho-resis. Relevant bands from these experiments were
visual-izedbyautoradiography ofwetgel, followed byrazorblade
excisionof thegelslices. Aftersaturationat68°C for15to30 mininSDS sample buffer, the gel sliceswere"stuffed" into
thestacking wells ofan SDS-polyacrylamide gel. This
sec-ond gel electrophoretically resolved the covalently linked protein-DNA complexes, as determinedfrom the migration
properties of theproteincomponent.
Oligonucleotide sequences and plasmid constructions. The
oligonucleotides used in the experimental protocols in-cluded:
(i) wild-type (WT)HTLV-I 21-bp oligonucleotide (32)
5'-GATCAAGGCTCTGACGTCTCCCCCC TTCCGAGACTGCAGAGGGGGGCTAG (The seven mutated versions of this 21-bp sequence are
listed inFig. 2.)
(ii)lac operatoroligonucleotide (57)
5'-GATCAATTATGAGCAGATAATAATT TTAATACTCGTCTATTATTAACTAG
(iii) ori-P oligonucleotide (66)
5'-GATCCAGATTAGGATAGCATATGCTACCCA
GTCTAATCCTATCGTATACGATGGGTCTAG
(iv)H23 oligonucleotide
5'-GATCAGAGCCTCCCAGTGAAAAACA
TCTCGGAGGGTCACTTTTTGTAAAG
(v) rat somatostatinoligonucleotide (46) 5'-GATCCCTTGGCTGACGTCAGAGAGA
GGAACCGACTGCAGTCTCTCTCTAG
For in vivo chloramphenicol acetyltransferase (CAT) as-says, tandem
synthetic
copies of WT(pWTA1OCAT2),
mu-tant HTLV-I 21-bp (e.g.,pH19A1OCAT2),
or ratsomato-statin
(pRSA1OCAT2)
oligonucleotides were inserted in the sense orientation into the unique BglII site of plasmidA1OCAT2
(35). ThisBglII restriction site is located approxi-mately 102 bp upstream of the SV40 AT-rich promoter element.Columnchromatography. A HeLa cell nuclearextract(11, 64) was made from 10 liters of
suspension
culture. The extractequilibratedin buffer(10mMTrishydrochloride,
pH7.9; 100mM KCl; 10% [wt/vol] glycerol) was fractionated
with afast protein liquid chromatography (FPLC) Mono-S
column
(Pharmacia).
A 60-ml linear salt gradient (starting buffer:10 mMTrishydrochloride, pH 7.9;100 mMKCl;10% [wt/vol]glycerol;end buffer: 10 mMTrishydrochloride, pH7.9; 1.0 M KCl; 10% [wt/vol] glycerol)wasused forprotein elution.
DNaseI footprinting. DNase I footprinting (18) was
per-formed withend-labeled DNA
fragments.
Singleormultiple
copiesof the WTorH19
synthetic
HTLV-I21-bpwasplaced
in the sense orientation into the unique BglII site of CATvector
pA1OCAT2
(35). The resulting plasmids werere-stricted with AccI and labeled with
[32P]dCTP
(3,000 Ci/ mmol;Amersham) and Klenow enzyme (NewEngland Bio-Labs, Inc.). Redigestion withSfiI
generated uniquely end-labeledfragments whichwere then gelpurified.In atypical experiment, 5,000cpmofprobe and 2 ,ug of poly(dI-dC)were incubatedon ice with variousamounts of proteinin buffer (20 mM HEPES, pH 7.9; 1.5 mM MgCl2;
100 mM KCl) for15 min. The reactionwasthenbroughtto roomtemperatureanddigestedwith DNase 1 (50,ug/ml,final
concentration) for 60 s, followed by phenol-chloroform extraction, ethanol precipitation, and analysis in a
urea-polyacrylamide gel.
In vitro transcription and RNA analysis. Cell-free
tran-scription was performed with whole-cell extract (44). A
typicalassaycontained0.5 p,gof supercoiledtestDNA,0.5 ,ugofaninternal control
P-globin
(ITSVHPBA128)
(58)DNA, and 1.5jig
ofHeLawhole-cell extract in 15 ,ul of buffer (20 mMHEPES,pH 7.9; 100 mMKCl;12.5mM MgCl2; 0.1 mMEDTA; 2mMDTT; 17%glycerol; 1 mM each of UTP, ATP,
CTP, and GTP). Incubations were for 1 h at 300C. Equal volumesof buffer or 100 ng of protein column fractions were addedto somereactions.
RNAtranscribed in vitro was isolated by phenol-chloro-formextraction, followed byethanolprecipitation, and then
analyzed by usingaprimerextension assay. A 5'-TTGGGA TATATCAACGGTGG-3' oligonucleotide primer was syn-thesized thatwas complementary tothe 5' endof the CAT gene.Details of thisanalysishavebeen described previously
(51).The M13probe and conditions for
S1
nucleaseanalysison November 10, 2019 by guest
http://jvi.asm.org/
HTLV-I 21-bp BINDING PROTEIN 4501
B 21 bp lacO
r-- -% I 21bP I{- lacO
---:u
120-
k
~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.
Mss
90-!§iw-to
M 1 2 3 4 5 M 312 34 56 1 23 4 5 6
FIG. 1. Identificationof HeLa cell factorsthatbindtothe 21-bp repeatedsequenceof theHTLV-I LTR. (A)Protection of the HTLV-I
21-bpsequencefromnuclease digestion.A 10-ngportionofa32P-labeled DNA fragment containingthree tandem copies of the HTLV-I 21-bp
sequencewasincubatedonice for15minwith (i) buffer alone(lanes 1 and 5), (ii)buffer with 10pugof HeLanuclearextract(lane 2),(iii)buffer
with50 jigof HeLa nuclearextract(lane 3),or(iv) buffer with 70jigofHeLa nuclearextract(lane 4). After incubation, each reaction mixture
wasdigestedwith 50 jig ofDNase Iperml and electrophoresed ina6% urea-acrylamide gel. Theregion of protectionoverthe HTLV-I 21-bp sequencesis labeled 3 x 21bp. The arrowhead pointstoagenerated DNase I-hypersensitive region. Molecular size markersareend-labeled
pBR322HpaIIfragments(lane M). (B) A WT 21-bpHTLV-I probeor alac-O probe(seeMaterials and Methods)wasused inUV-cross-linking
experiments with 50 ,ug ofHeLacell nuclear proteinextract.Reactionconditionswere asfollows:UVirradiation for 90s(lanes 1and4);UV
irradiationfor 90sfollowed byDNase I"trimming" at1 ,ug/ml(final concentration) for 5minatroomtemperature(lanes2and 5);andno
UV-irradiation (lanes 3 and 6). (Left) Coomassie blue-stainedgel. (Right) Autoradiographic exposureof the samegel. The fourgroupsof
bandsidentified in lanes1and 2oftheautoradiogram migrated with approximatemolecularweightsof >200,000, 180,000, 120,000, and 76,000
(toptobottom).
of,-globin-specific transcriptswere asdescribed by Laimins
etal. (36).
cAMPregulator experiments. K562 erythroleukemia cells (107cellspertransfection)weretreatedwith 5jigof eithera
human immunodeficiency virus (HIV) LTR-CAT (pBENN CAT)(22)or anHTLV-ILTR-CATplasmid (pU3RCAT) (58) bytheDEAE-dextranprocedure (8).At24 h aftertreatment the cells were either mock induced or induced with cAMP
regulators (25 ,uM forskolin, 1 mM 8-bromo-cAMP, and 0.5 mM3-isobutyl-1-methylxanthine; Sigma).Afteranadditional
24 h(i.e.,48 h afterDNAintroduction), cellular CAT activi-ties were assayed. The same experiments were also
per-formed withplasmidspWTA10CAT2and
pH19Aj0CAT2.
Other techniques. Standard approaches for recombinant DNA manipulations, gel electrophoresis (43), and CATassays(24)wereused. CATassayquantitations werebased
upon the average percentage ofincorporation from three separateexperiments, as assessedby scintillationcounts.
RESULTS
Identification of cellular factors that bind to the HTLV-I LTR. The LTRs of several retroviruses are important in
determiningthepathogenicity and tissuespecificityof these viruses (5, 10, 39; for a review, see reference 13). The HTLV-I LTRcontains apromoter element that is strongly and efficiently trans-activated by a virally encoded gene
product, p40x,which is also known astat-I (14, 17, 53, 58).
Becausebiologically active HTLV-I p40x produced in large
amountsbyeitherprocaryotic (22)oreucaryotic (29)
expres-sion systems shows no detectable DNA-binding property
(unpublished observations),webelieve thatcellular factor(s) probably play an important role in this trans-activation process.
Threerepeated 21-bpsequencesarefound in the HTLV-I LTR andareconservedatthesequencelevel in theHTLV-II LTR (55). This 21-bp sequence,when present inmore than
onecopy, isa sufficient target forp4Ox-mediated
transcrip-tional trans-activation (4, 50, 54). We reasoned that the 21-bp repeated sequence would be a likely binding site for putative cellular factors. This reasoning was tested by DNase I footprinting and a UV-cross-linking procedure to
identifyfactors withineucaryotic cells that specifically
rec-ognize thissequence.
We inserted three synthetic copies ofthe HTLV-I 21-bp repeat in tandem(see Materials and Methods) upstream of
an enhancerless SV40 promoter. This construction permit-ted a biochemical analysis ofthe 21-bp repeats that were
independent of other HTLV-I LTR sequences. A uniquely end-labeledDNAfragmentcontainingthe threecopiesofthe
HTLV-I 21-bp repeat was gel isolated and incubated with
HeLa cell nuclear proteins. Subsequent DNaseI treatment revealed a stronglevel ofprotection of the three copies of
HTLV-I 21-bp sequence from nuclease digestion by HeLa
cellularprotein(s)(Fig.1A,lanes 2 to4).Thisin vitrofinding
A
309- *
242-
160-VOL.62, 1988
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.71.560.72.325.2]suggested
that host cellfactor(s)
can indeedspecifically
recognize
theHTLV-I
viral21-bp
element.We next used one
synthetic
copy of the HTLV-I21-bp
sequence toidentify specific binding protein(s)
inHeLacell nuclear extract. AnHTLV-I
21-bp
oligonucleotide probe
wasfirstuniformly
incorporated
withBUdR and[32P]dCTP
and then incubated with HeLa cell nuclear
proteins.
This mixture was irradiated with UVlight
togenerate
covalentlinkages
between DNA andproximally
attachedproteins.
SDS-PAGE ofthe treatedsamples
delineatedfourproteins
(some
migrating
asdoublets)
with molecularweights
ofapproximately
76,000,
120,000, 180,000,
and>200,000
(Fig.
1B,
right)
thatwere visualizedby
autoradiography
by
theircoupling
toradiolabeledDNA.Parallelexperiments using
anoligonucleotide probe
containing
onecopyof theprocaryotic
lac
operator
sequence(see
Materials andMethods)
demon-stratedno
significant
binding
toeucaryotic
nuclearproteins
(Fig.
1B,
lanes 4to6).
These results illustrated theprefer-ential
recognition
of the HTLV-I21-bp
sequenceby
atleast four nuclearpolypeptides
over asimilarly
sized butunre-latedDNA
fragment.
Correlationof in vitro
protein binding
withinvivo function. Wemutagenized
the HTLV-I21-bp
sequencetoinvestigate
the nucleotide sequences
required
forprotein binding
andbiological activity.
One deletion mutation(H19)
and sixtransition mutations
(H21, H29, H31, H33, H45,
andH47)
spanning
the21baseswereindependently
generated
(Fig.
2,
top).
Twocopies
of each wereinserted,
in a senseorienta-tion,
upstream
ofanSV40
early promoter-CAT hybrid
gene(pA10CAT2
[36];
Fig.
2,
middle).
In theresulting plasmids
the HTLV-I21-bp
sequences werepositioned
roughly
100bases from the
SV40
early
TATA element. Eachplasmid
was cotransfected into CV-1 cells with a
p40x-producing
plasmid
(48).
CAT activitieswereassayed
todetermine their relativeresponsiveness
top40X.
Wefoundthatonly
the H19 and H21mutations,
whencompared
withWT,
wereinactiveto trans-activation
by
p40x
(Fig.
2,
bottom).
The otherfive mutations(H29, H31, H33, H45,
andH47)
did not differsignificantly
fromWTinbiological activity (Fig.
2,
bottom).
WT
oligonucleotide
andoligonucleotides
of each ofthe seven mutationswere radiolabeled andusedingel
mobility
shift
experiments
(Fig.
3).
The WTprobe,
when incubatedwith HeLa nuclear extract,
produced
fourmigration-re-tarded
DNA-protein complexes (Fig.
3,
lane1,
bandsA, B,
C,
andD).
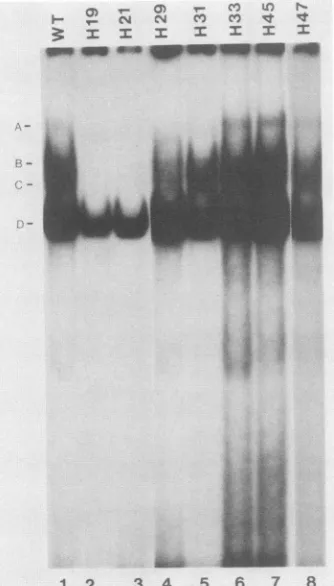
All of the mutantprobes
shared theDcomplex
(Fig.
3,
lanes 2 to8); however,
the two mutations thatinactivated
biological activity
(H19
andH21)
did notform either the B or the Ccomplexes
(Fig.
3,
lanes2 and3).
Incontrast,
both the B and C bands were seenwhen the fivebiologically
intactmutants(H29, H31, H33, H45,
andH47;
Fig.
3,
lanes 4 to8)
were usedasprobes.
The A band wasinconsistently
present
(Fig.
3,
lanes 1 and 6to8)
andoften variable inintensity.
These results correlated theformationofthe B and C
protein-DNA
complexes
in vitro with theirresponsiveness
top40X
trans-activationin vivo.Identificationof
polypeptides
contained within the protein-DNAcomplexes.
Thegel
shiftexperiment
described abovecorrelatedthe invitro
binding
by
thecellularfactorsrespon-sible fortheformationof the B and C
complexes
with the invivo
biological activity
of theHTLV-I21-bp
sequence. Weperformed
an additional set ofexperiments using
unrelatedoligonucleotides
ascompetitors
to furtherverify
the speci-ficitiesofthesetwocomplexes (Fig.
4).
Wefoundthat whenfour different
oligonucleotides
were used incompetition
studies,
theBand Ccomplexes
wereerasedonly by
the WTsequence
(Fig.
4A,
lane5),
whereastheAandDcomplexes
both showed variable amounts ofcompetition
by nonspecific
oligonucleotides(Fig. 4A, lanes 2 to4).
We next usedaradiolabeled,
BUdR-incorporated
probe
to identify thepolypeptidecomponentof the fourDNA-protein
complexes. A WT
probe,
incubated with nuclear extract,was UV irradiated before electrophoresis,
generating
anelectrophoreticprofileidentical to thatseenin
Fig.
4A(lane 1). In this instance, because of UVinduction,
the protein-DNAcouplingswerecovalent in nature.Thus,
the attached radiolabeled DNAprobepermittedthe visualization ofinter-acting proteinsby autoradiography after
electrophoresis
of the complexesby SDS-PAGE.Since the B and Ccomplexes
migrated
closetogether
in themobility shift analysis,they
were excisedtogether
in asinglegelslice. Thisgelslicewassaturated with SDS
sample
buffer, and theprotein-DNAcomplexeswereelectrophore-ticallyeluted and then resolved
by
SDS-PAGE. An autora-diogram of this polyacrylamide gel identifiedpolypeptides
with molecularweightsof180,000and120,000
(Fig.
4B,
lane 1). A similar analysis of the D complex demonstrated a76-kDaprotein(Fig. 4B,lane2). The A
complex
waspoorly
UV cross-linked.Aweaklylabeledproteinmigratingwitha
molecular weight in excess of
200,000
was sometimes ob-served (K.-T. Jeang, results notshown).
Analysis of the 180-kDaproteinin nucleaseprotection and cell-free transcription. The UV-cross-linkingprocedure can
be used to visualize relevant polypeptides
during
purifica-tion. This approach is sensitive enough to detect
relatively
small amounts of DNA-binding activity against alarge
background of total cellularproteins. This is howwe moni-tored the chromatographic separation of the four
HTLV-I
21-bp bindingproteins.Apreparation of HeLa nuclear extract was applied to a
FPLC Mono-S resin column (see Materials and
Methods)
and eluted with a linear (0.1 to 1.0 M KCl) salt
gradient.
Sequential column fractions were collected, and a smallportion ofevery second fraction was tested by UV
cross-linking to a radiolabeled BUdR-incorporated WT
oligonucle-otideprobe. These samples, after treatment, wereresolved bySDS-PAGE, stainedwithsilver, andexposedfor autora-diography. Although the silver-stainedprofilesof the column fractions were unremarkablefor any particular
polypeptide
(Fig. SA, left), relatively clear patterns identifying the76-,
120-, 180-, and >200-kDa proteins were seen in the
autora-diogramof the samegel (Fig. SA, right). This
autoradiogram
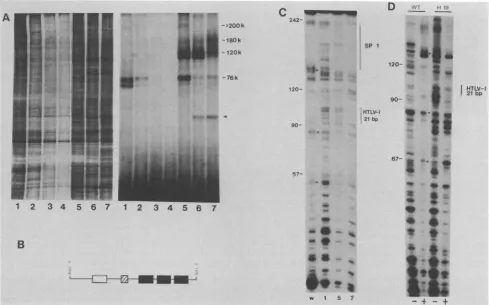
also delineated a partial separation of the four groups of DNA-bindingactivitiesthatcould nototherwise be appreci-ated in the stained gel.Thisseparation of the four DNA-bindingproteinsallowed ustoassesseachproteinfor its individual ability to protect the HTLV-I 21-bp sequence from DNase I digestion. A DNA probe containing one copy of the HTLV-I 21-bp
sequence was uniquely radiolabeled at an AccI restriction
site (Fig. SB). This 249-bp fragment, following incubation
with whole HeLa nuclear extract (Fig. SC, lane w) orwith
selective elution fractions from the FPLC Mono-S column (Fig. 5C, lanes 1, 5, and 7), was treated with DNase I. As
expected,
weobserved aprotectingactivity for theHTLV-I21-bp
sequence in the whole unfractionatednuclear extract(Fig. SC, lanew).Thisactivityintheunfractionated nuclear extract was specific for binding to the WT HTLV-I 21-bp
element, since it showed poor DNase I protection for a
biologically
inactive mutated 21-bp sequence (Fig. SD).In serialassays of column fractions, we found that fraction
1,
which contained the 76-kDa protein (Fig. SA, lane 1), showednoprotectionovertheHTLV-I 21-bp sequence(Fig.on November 10, 2019 by guest
http://jvi.asm.org/
HTLV-I 21-bp BINDING PROTEIN 4503
5'
H29 H21 H31 H33
AAG G CT CTG ACG TCT CCC CCC TTC C GA GAC TGC AGA GGG GGG
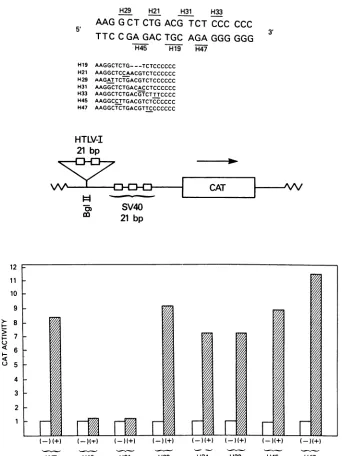
H45 H_
3'
H19 AAGGCTCTG- --TCTCCCCCC H21 AAGGCTCCAACGTCTCCCCCC H29 AAGATTCTGACGTCTCCCCCC H31 AAGGCTCTGACACCTCCCCCC H33 AAGGCTCTGACGTCTTTCCCC H45 AAGGCCTTGACGTCTCCCCCC
H47 AAGGCTCTGACGTTCCCCCCC
HTLV-I 21 bp
SV40
co 21
bp
12 11 10 9
>-8
> 7
<6 0 5 4 3
WT H 19 H21 H29 H31 H33 H45 H47
FIG. 2. Mutationalanalysis of the HTLV-I 21-bp sequence. (Top) The WT HTLV-I 21-bpsequenceispresentedinduplexform. The relativepositions ofsevenindividual mutationsareindicatedbyover- orunderlining.The actual sequences of the mutatedoligonucleotides
arepresented in smaller lettersshowingthesensestrandonly.(Middle)SchematicrepresentationofaSV40-CATvector
(pAjOCAT2)
(35)that containstwotandem copies of HTLV-I 21-bp repeat. (Bottom) Relative CAT activities in arbitraryunits for the various HTLV-I 21-bpAjOCAT2
constructions transfected into CV1 cells with (+)orwithout(-)ap40x-producing plasmid (48).Thegraphrepresentsthe average values from three separate experiments.5C, lane 1). Column fraction 5, which contained primarily the76-and 120-kDaproteins, alongwith smallcontaminating amountsof the 180- and >200-kDaproteins (Fig. 5A,lane5), also did not protect the HTLV-I
21-bp
sequence(Fig.
5C, lane 5). Fraction 7, however, which was enriched withthe180-kDa
protein
(Fig.
5A, lane7),
did protect the 21-bpsequencefrom nuclease
digestion (Fig. 5C,
lane7). Since the DNAprobe used forfootprinting
also contained threecopies of theSV40 GCbox,
aprominent
SP1activity
overtheSV4021-bp repeats (12) could also be seen in theunfractionated HeLanuclearextract
(Fig. 5C,
lanew),
aswellasin column fraction 7(Fig. 5C,
lane7).The in vitro
footprinting
results are consistent with the180-kDafactor (complexB, Fig. 3 and4) playingarole in the biology of p40x-mediated trans-activation. To verify the function of this factor in cell-free transcription, fractions from the FPLC Mono-S column containingpartiallypurified
120-kDa(Fig. 6A,lane5)or180-kDa(Fig. 6A, lane7) factors were separately added to Manley transcriptionlysates pre-paredfrom HeLa cells. Twotemplateswerecomparedinthe in vitro transcription assay. The first was plasmid pHl9
A1OCAT2,
which contained two synthetic copies of the inactive H19 mutations placed upstream ofanSV40 early-promoter-driven CATgene (Fig. 6B).The second plasmid,pWTA1OCAT2,
contained tandemcopiesof the WTHTLV-I VOL.62, 19881
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.141.483.68.524.2]CF -N )
T-C
CY) Lr P"" CM4 N M m q* II
'3 I :r I I ]: I
_ _
_i z ii [image:6.612.320.553.74.265.2]12 3 4 5 6 7 8
FIG. 3. Bindingofcellular factors toWT and mutated HTLV-I
21-bpoligonucleotides. 32P-labeled WTormutantoligonucleotides (10 ng)and 4,ug ofpoly(dI-dC)wereincubatedwith 50,ugof HeLa
cell nuclearprotein. Theprotein-DNA complexeswereresolved in
a6%gel. The free probe hasmigrated offthegel. Theindividual
probesused inthedifferentincubationsarelabeledatthetop.The fourprotein-DNA complexes describedin thetextaredesignated A,
B, C,and D.
21-bp repeat placed upstream of the SV40 early promoter
(Fig. 6B).
The templates were separately added with a P-globin
controlplasmid (61)toManleyHeLacellextract. Incubation wasconductedat30°C for 60mininthepresenceorabsence
ofsupplementation with FPLC column fractions enriched for either the 120- or 180-kDa protein. Half of the in vitro-generated RNA from each reactionwasused forassay
by primer extension with a CAT oligonucleotide (Fig. 6C,
top).Theotherhalfwasquantified for 3-globin-specific RNA
by Si nuclease protection by using a single-stranded M13
probe (Fig. 6C, bottom). These results showed an
approxi-mately 10-fold stimulation by the 180-kDa-protein-con-taining fraction of
pWTA1OCAT2
when compared with the mutantpH19CAT2 template (Fig. 6C; compare lane 4 withlane 8). In contrast, addition of the 120-kDa-protein-con-tainingfraction to the incubation mixtures failed to cause
significant differences in the relative transcriptional rates
between the two templates (Fig. 6C; compare lane 3 with
lane 7). Transcription from the internal control f-globin promoter remained relatively constant in each of the dif-ferent reactions (Fig. 6C, bottom). Although we cannot
exclude eitherpositive or negative contributions from
con-taminating cofractionated proteins, one interpretation of
thesefindings is thatspecific binding by the 180-kDaprotein to the HTLV-I 21-bp repeats in plasmid
pWTA1OCAT2
positivelyaugments transcription.1 2 3 4 5
FIG. 4. Characterization of HTLV-I 21-bp-sequence-binding proteins bysubstratecompetition. (A)A10-ng sampleof WT21-bp probe incorporated with [32P]dCTPand BUdRwasincubated with
50jigof HeLa nuclearextractin thepresenceof 2,ugofpoly(dI-dC) (lane 1), 1 ,ugof H23 oligonucleotide (see Materials andMethods; lane 2), 1 pLg of ori-P oligonucleotide (lane 3), 1 p.g of lac-O oligonucleotide (lane 4), or 1 ,ugof unlabeled WToligonucleotide (lane 5). Sampleswereelectrophoresed ina4%polyacrylamide gel followedby autoradiography. Complexes A, B,C,and D(described previously)arelabeled.(B)A10-ngsampleofWT21-bpprobewas
incubated with HeLanuclearextractasin lane 1 (panel A) except that the reactionwasUV irradiatedbeforegel electrophoresis and
autoradiography. Regionsof thegel containing complexesBand C
togetherandcomplexD alonewereseparatelyexcised andanalyzed
by SDS-PAGE. Lanes: 1, proteins eluted from the B and C
complexes;2,proteinfrom the Dcomplex; 3,14C-labeledmolecular
weightmarkers. The three arrowheadspointtoproteinsof molecu-lar weights 180,000, 120,000, and 76,000 (top to bottom,
respec-tively).
Control of the HTLV-I LTRthrougha signaltransduction pathway. The identification of cellular proteins that can interact with the HTLV-I 21-bp sequence agrees with a
molecular model whereby p40x-mediated trans-activation
occurs via a cellularpathway. One possible route for the actionsofp4Ox was suggested byGoodman andco-workers
(46). These investigators found, by comparing nucleotide sequences,thepresenceofanoctamericsequencewithin the HTLV21-bpelement that is highly conservedingenes that
are transcriptionally regulated by cAMP (Fig. 7A). This finding, ifsignificant,arguesthatHTLV-I could have
incor-porated an element ofcellular metabolism in its
transcrip-tional repertoire. We therefore experimentally determined whetherthe HTLV-I LTRpromoter could be regulated by changesinintracellular cAMPlevels.
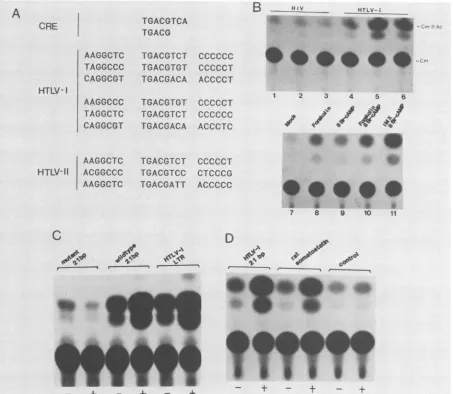
Initial screening experiments showed that the HTLV-I LTR was inducible by cAMP in a variety ofdifferent cell
types. Erythroleukemia K562 cells, however, consistently gave the highest level ofinduction. We transfected K562 cells with eitheranHIVLTR-CAT(pBENNCAT) (22) (Fig. 7B, samples 1 to 3) or an HTLV-I LTR-CAT (pU3RCAT)
(58) (Fig. 7B, samples4to6) plasmid. After24h,cellswere
mock treated (Fig. 7B, samples 1 and 4), treated with adenyl-cyclase stimulator(forskolin; Fig. 7B, samples2and 5), ortreated with cAMP analog (8 bromo-cAMP; Fig. 7B, samples 3and6). cAMPregulatorssignificantly induced the HTLV-I LTR-CAT plasmid but notthe control HIV
LTR-A
B
P
4
1 2 3
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.97.264.80.373.2]HTLV-I 21-bp BINDING PROTEIN 4505
C200k24-D WT H19
1l-
,1ll1|
~~~~~~~-180k
||. 1k0 , 120- a : bp
267- H7;V,1
5.
-separation of HTLV-I 21-bp-sequence-bindng proteins
d
1 2 3 4 5 6 7 1 2 3 4 5
67-w1 5 7
FIG. 5. Partial separation of HTLV-1 21-bp-sequence-binding proteins and
analysis by
nuclease protection. (A)Sequential
column fractions (lane 1, low salt; lane 7, high salt) of HeLa nuclear extract from an FPLC Mono-S column were assayed by UV cross-linkingto WT probe.(Left)Silver-stainedSDS-PAGE of the assayed fractions.(Right)Autoradiogram of thesamegelshowing the presence of the 76-, 120-, 180-, and>200-kDa proteins. The arrowhead pointsto a46-kDaprotein that is observed in differentHeLanuclearpreparations (lanes6and 7)andmay representaproteolytic product. (B)Schematicrepresentationof the DNAfragmentusedfor DNase Ifootprinting.Therelative positionsof asingle copyof the HTLV-I21-bp sequencein the sense orientation(O),a one-thirdcopyof theSV4072-bp repeat(u),
and three copiesof theSV4021-bp repeat(U)
areshown. The latter two elements are as described for theplasmidA,oCAT2
(35).(C)DNase Iprotection of HTLV-I21-bp sequencewithwhole HeLa nuclear extract or selective column fractions. Lanes: w,unfractionated wholeHeLaextract; 1,fraction 1 frompanel A; 5,fraction 5 frompanel A; 7,fraction 7 frompanelA. Nucleotidesarenumberedbytheir distance fromtheAccIsite. Protectedregionsover the SV4021-bp repeat (SP 1)and theHTLV-I 21-bp sequencearelabeled.(D)DNase IassayofWTorH19
sequenceswithwhole HeLa nuclear extract. WTprobe fragmentis asdescribed forpanelB.TheH19probeisidenticalto WTexceptthat asinglecopy of the H19 oligonucleotidein the sense orientation is substituted inplaceof theWTsequence.DNase Idigestion
patterns
in theabsence(-)orpresence(+)of HeLaproteinextract are shown. EquivalentpositionsinpanelsC andD areindicatedbyfilledsquares.CAT
plasmid.
When thesecellsweretreatedsimultaneously
withtwo separate cAMPmodulators (e.g., 8bromo-cAMP and
isobutyl-methylxanthine),
apositive
induction ofanup to 25-fold increase over the basal strength of the LTR promoterwas observed(Fig.
7B, comparelane 7with lane11). This increase is
comparable
to that seen forp40X-mediatedtrans-activationofthe HTLV-I LTR in K562 cells
(results notshown).
In other
experiments,
we usedpA1OCAT2
plasmid
con-structions which contained tandem
copies
of eithermutated(pH19A10CAT2;
Fig. 7C)
or WT(pWTA1OCAT2;
Fig. 7C) HTLV-I 21-bpoligonucleotides.
We introducedpH19A1O
CAT2,
pWTA1OCAT2,
orpU3RCAT
(HTLV-I
LTR;
Fig. 7C)
into K562cells andtested theinducibility
ofeachby
cAMPregulators. CATassaysshowedthat theH19
plasmid,
whichwas not trans-activated
by p40x
(Fig. 2),
was also notstimulated
by
increased levels of intracellular cAMP. In contrast, cells transfected with eitherpWTA1OCAT2
orpU3RCAT
showedelevatedacetylase
activities after treat-mentwith 8bromo-cAMP andisobutyl-methylxanthine).
The results described above indicated that the HTLV-I
21-bpsequence,under the
appropriate
conditions,
can serveas a
cAMP-responsive
element. Thisfinding
is of interest since itsuggestedthecross-utilization oftargetsitesby
bothp40"
andcAMP.Wetested thispossibility
withtheconstruc-tion ofpRSA10CAT2,aplasmidthatcontainedtwo
synthetic
copies
of the rat somatostatincAMP-responsive
element (see Materials andMethods)upstreamofanSV40promoter-driven CATgene. Wefoundthat while
pA10CAT2
(control;
Fig. 7D) did not respond top40x
induction,
pRSA10CAT2
(ratsomatostatin; Fig.
7D) in the presence ofp40x
protein
did show an increase in CAT
activity comparable
to theincreaseseenfor
pWTA10CAT2
(HTLV-I
21bp;
Fig. 7D).
DISCUSSION
Inthisstudywehave usedaUV-induced indirect
labeling
techniquetoidentify
cellular factors thatbindtothe HTLV-I21-bp
p40X-responsive
sequence. Thisapproach
issimilartothatreported
by
Sharp
and co-workers(6).
We and others(65) have utilized this method in
conjunction
withmobility
shift assaystofurthercharacterize
protein
factors responsi-bleforgel-retarded
complexes
(Fig. 4).
Wehave
identified,
using
thisUV-labeling
approach,
fourcellular
polypeptides
of76, 120,
180,
and>200 kDainHeLa VOL.62, 1988on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.65.554.75.380.2]B
1f...
-C
Mutant Wildtype
.~.s
160I
12G _
Vt
527
,w40
M 1 2 3 4 5 6 7 8 9
FIG. 6. Stimulation of in vitrotranscription bycolumnfractioncontainingthe 180-kDaprotein. (A) Sequentialcolumn fractions froman
FPLC Mono-S columnwereassayed by UV cross-linkingtoWT probe (lanes1 to 11). Anautoradiographshowing the separationofthe 76-kDa (lanes1and2),120-kDa(lanes 5 and6),and 180-kDa(lanes7 and8) proteinsispresented. For in vitrotranscriptionassayseither fraction 5 (lane 5)orfraction7(lane 7)wasused forsupplementation.LaneM, '4C-labeledproteinmolecularweightmarkers.(B)Schematic representations ofthe analysis of in vitro-generated transcripts. RNA synthesized from the HTLV-I pWTAjOCAT2 or pH19A1OCAT2 templateswereanalyzed withaCAT-specific primer.Asingle-strandedM13probe (426 bp)wasused toanalyzeP-globin-specific transcript generatedfromaninternal controlplasmid by S1nuclease protection. (C) Analysesof RNA fromin vitrotranscription reactions.(Top) A 32P-labeled20-bp primer complementary tothe CAT mRNA(59)was usedtoquantitateRNA transcribedfromapA1OCAT2template (35)
containingtwocopiesofsynthetic WTormutantHTLV-I21-bpsequence(see panel B).Eitherfraction 5 orfraction7(panel A)wasused
tosupplementaManleyextracttitratedtogivealow level of basalactivity. (Bottom) AnalysisoftranscriptsfromaP-globininternalcontrol plasmid with a uniformly radiolabeled M13 single-stranded probe. Lanes: 1, basal transcription of pH19AjOAT2; 2, transcription of
pH19AjOCAT2
in thepresenceof 1,ugof a-amanitinperml; 3, transcriptionofpH19AjOCAT2
supplementedwithfraction5; 4, transcription ofpH19A1OCAT2 supplemented with fraction 7; 5, transcription ofpWTA1OCAT2 in Manley extract without supplementation; 6, sametemplate transcribedin thepresenceof1 ,ugofa-amanitinperml; 7, transcriptionof
pWTAjOCAT2
supplementedwith fraction5; 8,sametemplate transcriptionallysupplementedwith fraction7; 9,noDNA addedtotranscriptionreaction. Abbreviations:CAT,a140-nucleotide band,theexpectedsize for RNA initiated from theSV40 earlypromoter;globin, a426-band transcript; M,molecularsizemarkers.
nuclearextractsthat bindtotheHTLV-I 21-bprepeat(Fig. 1). Of these four proteins,two(120and 180kDa)werehighly
specific for bindingtothe WT21-bpsequenceand resistant tocompetition with heterologousDNAs(Fig. 4).Inmobility shiftassays,sevenoligonucleotidesthat mutatedatdifferent positions within the HTLV-I 21-bp element demonstrated that the invitro binding of the 120- and 180-kDa proteins (but
notthe 76-or>200-kDaproteins) correlated well with the in
vivo responsiveness of these mutants totrans-activation by p40x (Fig. 2). Comparing the 120- and 180-kDa factors, we
found that column fractions enriched with the latter
con-tained an HTLV-I 21-bp DNase I protecting activity and were active in the transcriptional stimulation of HTLV-I 21-bp-sequence-containing templates in vitro (Fig. 5 and 6). Preliminary experiments suggested that the 120- and 180-kDaproteins shared some commonpartial proteolytic
frag-ments(K.-T. Jeang, unpublished data). Given the possibility ofdegradation during the protein extract preparation, the 120-kDamoleculemayrepresentaproteolytic product of the
larger, biologically active 180-kDa protein.
Weconsidered that the180-kDa proteinmayplayarole in the trans-activation of the HTLV-ILTRby p40x. While the exactmechanism of this regulation remainstobe elucidated,
onesuggestion has been that p40" mayinteract directly with
sequences upstreamofresponsive promoters (19). We have usedbiologically active protein fromoverproducing
procar-yotic cells (23), from overproducing eucaryotic cells (30), and froman HTLV-I-transformed T-cell line (37) and have beenunabletoattributeanyinvitro DNA-binding properties
top40" (unpublished observations). Hence, itappears
prob-able that p4Ox utilizes cellular protein intermediaries in interactions with target promoterelements. Sincethe
com-pletion ofour study, other investigators (1, 49) have also
demonstrated binding of host cell factor(s) to the HTLV-I 21-bp element and have suggested an indirect pathway in
p40X-mediated activation of the HTLV-I LTR.
The underlying mechanistic interplay between p40x and cellular factors is not obvious. We have noted that the
presence ofp4Ox protein, added exogenously, in DNase I footprinting assays or in cell-free transcription reactions does not appreciably alter either the pattern of nuclease protection or the level of template-specific transcriptional
stimulation (unpublished observations). Thus the effects of p40x may not be easily reproducible in vitro by simple protein-protein interactions. These observations suggest a complexschema ofp40x-mediated regulationwhich is com-patiblewithpublishedreportsof thetrans-activationby p40" ofheterologous promoters that donotshareovertsequence homologywiththe HTLV-I LTRorwitheach other(8, 29).
How might p40x then activate a number of unrelated genes? One possibility is that this proteinutilizes a
ubiqui-toussignaltransductionpathway suchas acAMP-dependent
cascade. The cAMP kinase cascade isoperativeinvirtually all eucaryotic cells (for a review, see reference 25) and probablyrepresentsanextension ofaprocaryotic regulatory
mechanism(63). Hence,it isperhaps morethan coincidence
that theHTLV-I, HTLV-II (Fig. 7A), and bovine leukemia virus(9) 21-bp repeatedsequences all conserve, withslight
variations, theconsensusoctanucleotide commonly found in cAMP-responsivegenes(46). Inthepresentstudy,we have
demonstrated that intracellular cAMPlevelscanspecifically
coordinate theexpressionofareportergenethat is linkedto the HTLV-I LTR (Fig. 7B). Three pieces of evidence suggest that this cAMP-mediated regulation is relevant to
p4ox trans-activation. First, the HTLV-I 21-bp element, in multimers,canserve asasufficienttargetfor both p40x (Fig. 2)andcAMP(Fig.7C).Second,amutation (H19) within the
cAMP 5'-TGACGTCA-3' consensus (46) of the 21-bp
se-quence inactivated both p40x (Fig. 2) and cAMP (Fig. 7C) responsiveness. Third, an unrelated cAMP-responsive ele-mentfrom the ratsomatostatin gene canapparently
substi-tutefor the HTLV-I21-bpsequencein p40x-mediated
trans-activation (Fig. 7D). These findings suggest that p40x and cAMPmayshareaconvergent,butnotnecessarily identical, pathway in the regulation ofsome genes. Similar observa-tionscorrelating aninteraction betweenthe adenovirus Ela
A
160 120
1 2 3 4 5 6 7 S 9 l1 m
-CAT
-Gobin
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.63.550.71.196.2]HTLV-I 21-bp BINDING PROTEIN 4507
B
H I Y ..w. -fu*
S.-~~~~~~~~~~~~~~~~~~~~~~~~~~~
HTLV-I
fi_ _ _
_oI
|; -i---A,,-*e
*@(A
1 2 3 4 5 6
0
4~
7 8 9 10 11
C
opoeo I -& , I r
D
4<
--% I& 04.
_; X
+ - - tI
- +
-FIG. 7. The HTLV-ILTR containsacAMP-responsive element. (A) The octameric (46) and pentameric (62) cAMP-responsive element
(CRE)arecompared against the three21-bpsequencesfound inanAmerican HTLV-Iisolate (upperrows[32]),aJapanese HTLV-I isolate
(lowerrows[52]), andaJapanese HTLV-II isolate (55). (B) An HIV LTR-CAT plasmid (pBENNCAT) (22)or anHTLV-ILTR-CATplasmid
(pU3RCAT) (58) wasseparately introduced into K562 cells. CAT activities wereassayed 48 h after transfection and 24 h after regulator
treatmentfrommock-induced cells (samples 1 and 4), cells induced with 25 ,uM forskolin (samples 2 and 5), and cells induced with 1 mM 8 bromo-cAMP(samples 3and 6). Samples 7 through 11comparethe relative induction values of the HTLV-I LTRby single (forskolin, sample
8;8bromo-cAMP, sample9)ordouble(forskolin plus 8 bromo-cAMP, sample10; isobutyl-methylxanthine plus 8 bromo-cAMP, sample 11) cAMP regulators. Abbreviations: Cm, chloramphenicol; Cm-3-Ac, acetylated chloramphenicol. (C) CAT activities from K562 cells
transfectedc with plasmidspH19A1OCAT2 (mutant 21 bp), pWTA1OCAT2 (wildtype 21 bp),or pU3RCAT(HTLV-I LTR) that were mock
induced(-)orinduced (+) simultaneously with1mM 8bromo-cAMP and0.5mMisobutyl-methylxanthine. (D)K562 cellsweretransfected
separately with plasmid DNAspWTA1OCAT2(HTLV-I21bp),pRSA1OCAT2(rat somatostatin),andpA1oCAT2(control).-,Samplesthatare
plasmid DNAs transfected alone; +,samples thatareDNAscotransfectedwithanHTLV-I p40x-producing plasmid (48).
andcAMP-responsiveelementshave also beenreported (26, 40, 41).
Should p4OXand cAMPutilize a similartarget sequence,
the role ofcAMP-dependentkinases in transcriptional acti-vation by p40X should not be overlooked. Since mutations (H19 and H21) within the highly conserved pentameric subset(62)of thecAMPoctamer(46)notonlyresultedinthe loss of biological activity but also in the inability ofthe HTLV-I 21-bp sequence to bind the 120- and 180-kDa
proteins (Fig. 3),wesuggestthat thesepolypeptidescouldbe
participating factors in a p40x-interactive cellular cascade.
Recently, Montmniny and Bilezikjian (45) purified a 43-kDa
proteinfrom PC12 cells that boundspecificallytothe
cAMP-responsiveelement of the somatostatingene.Linand Green
(41)havealsoindependentlyidentifieda46-kDaproteinfrom HeLa cells that bind to cAMP-inducible promoters by a
UV-cross-linking approach. We have also noted in our
analyseswith HeLacell nuclearproteins the variable
pres-enceofa46-kDaproteinthatcanbind to the HTLV-I21-bp
element (Fig. 5A, lanes 6 and 7, arrow; Fig. 6A, lane 7). Using an oligonucleotide affinity column containing the
HTLV-I21-bpsequence,wehavepreparatively purifiedthis
46-kDaspeciestohomogeneity,asassayed bysilverstaining
onSDS-PAGE(unpublished results).The exactrelationship betweenthe 43-kDa PC12protein,the 46-kDa HeLaprotein, and the higher-molecular-weight HeLa cell HTLV-I 21-bp-TGACGTCA
TGACG
A
CRE
HTLV-I
HTLV-II1
AAGGCTC TAGGCCC CAGGCGT
AAGGCCC TAGGCTC CAGGCGT
AAGGCTC ACGGCCC AAGGCTC
TGACGTCT TGACGTGT TGACGACA
TGACGTGT TGACGTCT TGACGACA
TGACGTCT TGACGTCC TGACGATT
cccccc
CCCCCT ACCCCT
CCCCCT COCCCC ACCCTC
CCCCCT CTCCCG ACCCCC VOL.62, 1988
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.612.86.539.73.467.2]sequence-binding proteins is unclear. However, in
gel
shiftassays with an HTLV-I
21-bp-sequence-containing probe,
wehaveascertained thatthepurified HeLa46-kDapolypep-tide is not responsible for the formation of the
presently
describedA, B,C, orDprotein-DNAcomplexes (Fig. 3and4). We suggestthat there maybe multipleproteins, perhaps
related, which have similar binding affinities but have
dif-ferent functionalpropertieswithregardtotheHTLV-I
21-bp
element. This isnot totally surprising since there are otherexamples in which morethanonedifferently sized
polypep-tide has been shown to interact with a single consensus
sequence (15, 56).
A functionalreason for p4Ox to actthrough ageneralized
cellular pathway may be the need to simultaneously affect many different genes. One report has suggested that an
overlap mayexistin theactionsofphorbol esterand cAMP
(7). The recent description of a phorbol-ester-responsive
coresequence 5'-TGACTCA-3' (2, 38)thatcloselyresemble
the cAMP consensus 5'-TGACGTCA-3' (46) is thus not
unexpected. We note with interest thattwo well-character-ized p40x-responsive promoters (interleukin-2a receptor [8]
and HTLV-ILTR [28]) are both alsophorbol ester
respon-sive. Hence, p4Ox may contemporaneously activate genes necessaryfor leukemogenesis(interleukin-2and itsreceptor)
and for viral replication (HTLV-I genomic sequences) throughcellular signal transduction mechanisms.
ACKNOWLEDGMENTS
Wededicate thisworktoGeorge Khourywhowas amentorand
a friend. His tireless leadership and incisive intellect were the
driving forcesbehind this andmanyotherprojects. Weareindebted
toPhilipSharp for providing his manuscript priortopublication. We
thank GaryHayward, Malcolm Martin, CarlWu,WarrenLeonard,
Gary Felsenfeld, PeterHowley, Arnold Rabson,and David Living-stonfor criticalreadings and discussions of the manuscript.Wealso
thankPamelaMellon forbringingtoourattentionthepresenceofa
cAMP consensus sequence in the HTLV-I 21-bp repeats. The
technicalassistanceof Janet Duvall and the secretarial assistance of
Carol Beckerarealso appreciated.
This study was supported in part by the National Institutes of
HealthintramuralAIDS-targetedantiviral programfrom the office
of the Director.
LITERATURECITED
1. Altman,R., D. Harrick,J. A. Garcia,and R. B. Gaynor. 1988.
Human T-cell leukemia virus types I andII exhibit different
DNase Iprotectionpatterns.J. Virol. 62:1339-1346.
2. Angel, P., M.Imagawa, R. Chiu, B. Stein, R. J. Imbra, H. J.
Rahmadorf,C. Jonat, P. Herlich, andM. Karin. 1987.Phorbol
ester-induciblegenescontaina commonciselement recognized
byaTPA-modulatedtrans-actingfactor. Cell49:729-739.
3. Berk, A.J., F. Lee,T. Harrison, J. Williams, and P. A. Sharp.
1979. Pre-early adenovirus Tgeneproduct regulates synthesis
of earlyviralmessengerRNAs.Cell 17:935-944.
4. Brady,J., K.-T.Jeang, J. Duvall, and G. Khoury. 1987.
Identi-fication of p40x-responsive regulatory sequences within the
HTLV-I LTR.J. Virol. 61:2175-2181.
5. Chatis, P., C. Holland,J. Hartley, W.Rowe, and N. Hopkins.
1983.Role ofthe 3' endof thegenomeindetermining the disease
specificityofFriend and Moloney murineleukemia virus.Proc.
Natl. Acad. Sci.USA 80:4203-4207.
6. Chodosh,L. A.,R. W.Carthew, and P. A. Sharp. 1986. A single
polypeptidepossessesthe binding andtranscriptional activities
of the adenovirus major late transcription factor. Mol. Cell.
Biol. 6:4723-4733.
7. Comb,M.,N. C. Birnberg,A.Seasholtz,E. Herbert, and H. M.
Goodman. 1986. A cyclic-AMP- and phorbol ester-inducible
DNAelement. Nature(London)323:353-356.
8. Cross, S. L., M. B. Feinberg, J. B. Wolf, N. J. Holbrook, F.
Wong-Staal, and W. Leonard. 1987. Regulation of the human interleukin-2 receptor alpha chain promoter: activation of a
nonfunctional promoterby thetransactivator gene of
HTLV-I.
Cell 49:47-56.9. Derse, D. 1987. Bovine leukemia virus transcription is
con-trolled by a virusencoded trans-acting factor andby cis-acting
response element. J. Virol. 61:2462-2471.
10. DesGrosellers, L., E. Rassart, and P. Jolicoeur. 1983. Thymo-tropism of murine leukemia virus is conferred by its long terminal repeat. Proc. Natl. Acad. Sci. USA 80:4203-4207. 11. Dignam, J. D., R. M. Lebowitz, and R. G. Roeder. 1983.
Accurate transcription initiation by RNA polymerase II in a soluble extract from isolated mammalian nuclei. Nucleic Acids Res. 11:1475-1489.
12. Dynan, W. S., and R. Tjian. 1983. Isolation oftranscription factors that discriminate between different promoters recog-nized by RNA polymerase
II.
Cell32:669-680.13. Feigenbaum, L., and G. Khoury. 1985. The role of enhancer elements in viral host range and
pathogenicity,
p. 181-194. In Banbury report 22: genetically altered viruses and the environ-ment. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.14. Felber, B. K., H. Paskalis, C.
Kleinman-Ewing,
F. Wong-Staai,and G. N. Paviakis. 1985. The pX protein of
HTLV-I
is a transcriptional activator of its long terminal repeats. Science 229:675-679.15. Franza, B. R., S. F. Josephs, M. Z. Gilman, W. Ryan, and B. Clarkson. 1987. Characterization of cellularproteins
recogniz-ing the HIV enhancer usrecogniz-ing a microscale DNA-affinity precipi-tation assay. Nature (London) 330:391-395.
16. Fried, M., and D. M. Crothers. 1981. Equilibria andkinetics of lac repressor-operator interactions by polyacrylamide gel elec-trophoresis. Nucleic Acids Res. 9:6505-6525.
17. Fujisawa, J., M. Seiki, M. Sato, and M. Yoshida. 1986. A transcriptional enhancer sequence of
HTLV-I
is responsible for trans-activation mediated byp40x
ofHTLV-I.
EMBO J. 4:713-718.18. Galas, D., and J. Schmitz. 1978. DNase footprinting: a simple method for the detection of protein-DNA binding specificity. Nucleic Acids Res. 5:3157-3170.
19. Gallo, R. C. 1986. The first human retrovirus. Sci. Am. 255:88-98.
20. Garner, M. M., and A. Revzin. 1981. A gel electrophoresis method for quantifying the binding of proteins to specific DNA region: application to components of the Escherichia coli lac-tose operon regulatory system. Nucleic Acids Res. 9:3047-3060. 21. Gaynor, R. B., D.
Hillman,
and A. J. Berk. 1984. Adenovirus early region1A protein activates transcription of a nonviral gene introduced into mammalian cells by infection or transfection. Proc. Natl. Acad. Sci. USA 81:1193-1197.22. Gendelman, H. E., W. Phelps, L. Feigenbaum, J. M. Ostrove, A. Adachi, P. M. Howley, G. Khoury, H. S. Ginsberg, and M. A. Martin. 1986. Trans-activation of the human immunodeficiency virus long terminal repeat sequence by DNA viruses. Proc. Natl. Acad. Sci. USA 83:9759-9763.
23. Giam, C.-Z., M. Nereberg, G. Khoury, and G. Jay. 1986. Expression of the complete HTLV-I pX coding sequence as a functional protein in Escherichia coli. Proc. Natl. Acad. Sci. USA 83:7192-7196.
24. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982. Recombinant genomes which express chloramphenicol acetyl-transferase in mammalian cells. Mol. Cell. Biol. 2:1044-1051.
25. Gottesman, M. M. 1980. Genetic approaches to cyclic AMP effects in cultured mammalian cells. Cell22:329-330.
26. Hardy, S., and T. Shenk. 1988. Adenovirus control regions activated by Ela and the cAMP response element bind to the same factor. Proc. Natl. Acad. Sci. USA 85:4171-4175. 27. Haseltine, W. A., J. Sodroski, R. Patarca, D. Briggs, D. Perkins,
and F. Wong-Staal. 1984. Structure of 3' terminal region of type II human lymphotropic virus: evidence for new coding region. Science 225:419-421.
28. Holbrook, N. J., J. D. Luethy, and J. Kin. 1987. Transient expression of foreign genes in lymphoid cells is enhanced by
on November 10, 2019 by guest
http://jvi.asm.org/
HTLV-I 21-bp BINDING PROTEIN 4509
phorbol ester. Mol. Cell. Biol. 7:2610-2613.
29. Inoue, J.-I., M. Seiki, T. Toniguchi, S. Tsumu, and M. Yoshida. 1986. Induction of interleukin-2 receptor gene expression by
p40"encoded by human T-cell leukemia virus typeI. EMBO J. 5:2883-2888.
30. Jeang, K.-T., C.-Z. Giam, M. Nerenberg, and G. Khoury. 1987. Abundant synthesis of functional human T-cell leukemia virus type I
p40x
protein in eucaryotic cells by using a baculovirus expression vector. J. Virol. 61:708-713.31. Jones, N., and T. Shenk. 1979. An adenovirus type 5 early gene function regulates expression of other early viral genes. Proc.
Natl. Acad. Sci. USA 76:3665-3669.
32. Josephs, S. F., F. Wong-Staal, V. Mazari, R. C. Gallo, J. Sodroski, M. D. Trus, D. Perkins, R. Patarca, and W. A. Haseltine. 1984. Long terminal repeat structure of an American isolate of type I human T-cell leukemia virus. Virology 139:340-345.
33. Kiyokawa, T., M. Seike, S. Iwashita, K. Imagawa, F. Shimuzu, and M. Yoshida. 1985. p27x"' and p21""', proteins encoded by the pX sequence of human T-cell leukemia virus typeI. Proc. Natl. Acad. Sci. USA 82:8359-8363.
34. Kingston, R. E., A. S. Baldwin, and P. A. Sharp. 1985. Tran-scription control by oncogenes. Cell 41:3-5.
35. Laimins, L. A., P. Gruss, R. Pozzatti, and G. Khoury. 1984. Characterization of enhancer elements in the long terminal repeat of Moloney murine sarcoma virus. J. Virol. 49:183-189. 36. Laimins, L. A., M. Holmgren-Konig, and G. Khoury. 1986. Transcriptional "silencer" element in rat repetitive sequences associated with the rat insulin 1 gene locus. Proc. Natl. Acad. Sci. USA 83:3151-3155.
37. Lee, T. H., J. E. Coligan, J. G. Sodoroski, W. A. Haseltine, S. Z. Slahuddin, F. Wong-Staal, R. C. Gallo, and M. Essex. 1984. Antigens encoded by the 3'-terminal regions of human T-cell leukemia virus: evidence for a functional gene. Science 226:57-61.
38. Lee, W., P.Mitchell, and R. Tjian. 1987. Purified transcription factor AP-1 interacts with TPA-inducible enhancer elements. Cell 49:741-752.
39. Lenz, J., D.Clelander,R. Crowther, R. Patarca, D. Perkins, and W. A. Haseltine. 1984. Determination of the leukemogenicity of a murine retrovirus by sequences within the long terminal repeat. Nature (London) 308:467-469.
40. Leza, M. A., and P. Hearing. 1988. Cellular transcription factor binds to adenovirus early region promoters and to a cyclic AMP response element. J. Virol. 62:3003-3013.
41. Lin, Y.-S., and M. R. Green. 1988. Interaction of a common cellular transcription factor, ATF, with regulatory elements in both Ela- and cyclic AMP-inducible promoters. Proc. Natl. Acad. Sci. USA85:3396-3400.
42. Mamouon, R. E., T. Astier-Gin, R. Kettmann, J. Deschamps, N.
Rebeyrotte, and B. J. Guillenaim. 1985. The pX region of the bovine leukemia virus is transcribed as a 2.1-kilobase mRNA. J. Virol. 54:625-629.
43. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
44. Manley, J. L., A. Fire, A. Cano, P. A. Sharp, and M. L. Gefter. 1980. DNA-dependent transcription of adenovirus genes in a soluble whole-cell extract. Proc.Nati. Acad. Sci. USA 77:3855-3859.
45. Montminy, M. R., and L. M. Bilezikjian. 1987. Binding of a nuclear protein to the cyclic-AMP response element of the somatostatin gene. Nature(London)328:175-178.
46. Montminy, M. R., K. A. Sevarino, J. A. Wagner, G. Mandel, and R. H. Goodman. 1986. Identification of a cyclic-AMP-responsive element within the rat somatostatin gene. Proc.Natl. Acad. Sci. USA83:6682-6686.
47. Nagashima, K., M. Yoshida, and M. Seiki. 1986. A singlespecies of pX mRNA of human T-cell leukemia virus type I encodes trans-activator p40x and two other phosphoproteins. J. Virol. 60:394-399.
48. Nerenberg, M., S. H. Hinrichs, R. K. Reynolds, G. Khoury, and
G. Jay. 1987. The tat gene of humanT-lymphotropicvirus type I induces mesenchymal tumorsintransgenic mice.Science 237: 1324-1329.
49. Nyborg, J. K., W. S. Dynan,I. S. Y.Chen,and W.Wachsman. 1988. Binding ofhost-cellfactortoDNAsequencesin thelong terminal repeat of T-cell leukemiavirustypeI:implicationsfor viral gene expression. Proc. Natl. Acad. Sci. USA 85:1457-1461.
50. Paskalis, H., B. K. Felber,andG.N.Pavlakis. 1986. cis-Acting sequences responsible for thetranscriptional activation of hu-man T-cell leukemiavirus type I constitute a conditional en-hancer. Proc. Natl. Acad. Sci. USA83:6558-6562.
51. Seiki, M., S. Hattori, Y. Hiryama, and M. Yoshida. 1983. Human adult T-cell leukemia virus: complete nucleotide se-quence of the provirusgenomeintegratedinleukemiacell DNA. Proc. Natl. Acad. Sci. USA80:3618-3622.
52. Seiki, M., S. Hattori,andM.Yoshida. 1982.Human adultT-cell leukemia virus: molecularcloningoftheprovirus DNA and the unique terminal structure. Proc. Natl. Acad. Sci. USA 79: 6899-6902.
53. Seiki, M., J. Inoue, T. Takeda, and M. Yoshida. 1986. Direct evidence thatp40" of human T-cell leukemia virus type I is a
trans-acting transcriptionalactivator. EMBO J. 5:561-565. 54. Shimotohno, K., M.Takano, T. Terunchi,and M. Miwa. 1986.
Requirement ofmultiple copies of a21-nucleotide sequence in the U3 region of humanT-cellleukemia virustype I and type II long terminal repeats fortrans-actingactivation oftranscription. Proc. Natl. Acad. Sci. USA83:8112-8116.
55. Shimotohno, K.,W. Wachsman, Y. Takhashi, D. W.Golde,M. Miwa, T. Sugimura, and I. S. Y. Chen. 1984. Nucleotide sequence of the 3'regionof ainfectious human T-cell leukemia virus typell genome. Proc. Natl.Acad. Sci. USA81:6657-6661. 56. Short, N. J.1988. Regulationoftranscription. Flexible
interpre-tation. Nature(London) 334:192-193.
57. Simons, A.,D. Tils, B. vonWilchen-Bergmann, and B. Muller-Hill. 1984. Possible ideal lac operator: Escherichia coli lac operator-like sequencesfromeukaryoticgenomes lack the
cen-tral G:C pair. Proc. Natl. Acad. Sci. USA 81:1624-1628. 58. Sodroski, J. G., C. A. Rosen, and W. A. Haseltine. 1984.
trans-Acting transcriptional activation of the long terminal
re-peat of humanT-lymphotropic viruses in infected cells. Science 225:381-385.
59. Spalholz, B. A., P.F. Lambert, C. L. Yee, and P. M. Howley.
1987. Bovinepapillomavirus transcriptionalregulation: localiza-tionoftheE2-responsive elements in thelong controlregion.J. Virol. 61:2128-2137.
60. Svensson, K.,andG.Akusjarvi.1984.Adenovirus 2earlyregion
1A stimulates expression of both viral and cellular genes. EMBO J. 4:789-794.
61. Treisman,R., M. R.Green, and T. Maniatis. 1983. cisandtrans
activation ofglobingenetranscription in transient assays. Proc. Natl. Acad. Sci. USA80:7428-7432.
62. Tsukada, T., J.S.Fink, G. Mandel, and R. H. Goodman. 1987. Identification of a region in the human vasoactive intestinal polypeptide gene responsible forregulation by cyclic AMP. J. Biol. Chem. 262:8743-8747.
63. Weber, J. T., K.Takio, K. Titani, and T. A. Steitz. 1982. The cAMP binding domains of the regulatory subunit of cAMP-dependent protein kinase and the catabolite gene activator proteinarehomologous. Proc. Natl. Acad. Sci. USA 79:7679-7683.
64. Wildeman,A. G., P. Sassone-Corsi, T. Grundstrum, M.Zenke,
and P.Chambon. 1984. Stimulation of in vitrotranscriptionfrom theSV40 early promoter by the enhancer involves a specific
trans-actingfactor. EMBO J. 3:3129-3133.
65. Wu,C., S.Wilson, B.Walker,I.Dawid,T.Paisley,V.Zimarino, and H. Ueda. 1987. Purification and properties ofDrosophila
heat shock activatorprotein. Science 238:1247-1253.
66. Yates, J. L., N. Warren, D. Reisman, and B. Sugden. 1984. A cis-acting element from the Epstein-Barr viral genome that permits stable replication of recombinant plasmids in latently infected cells. Proc. Natl. Acad. Sci. USA 81:3896-3900. VOL.62,1988