0022-538X/92/042226-06$02.00/0
Copyright © 1992, American SocietyforMicrobiology
Ar+
Plasma-Induced Damage
to
DNA
in
Bacteriophage A:
Implications for the Arrangement of DNA in the Phage Head
ELLIOT C.MENDELSON, WILLIAM W. NEWCOMB, ANDJAY C. BROWN*
DepartmentofMicrobiology and Cancer Center, University ofVirginia Health Sciences Center, Charlottesville, Virginia22908
Received 7 November1991/Accepted 10January1992
BacteriophageAwasbombarded withlow-energyAr+ ionswith the goal of determining whether particular
regions of the DNAgenome are found preferentiallyin the outer portion of the packaged DNA mass. The
strategywastofragment the DNAselectivelynearthesurface ofthevirusbyexposingintact phagetoAr+ions energetic enough to break covalent chemical bonds in DNAbut not energetic enough to penetrate deeply
beneath theviral capsid shell. Broken DNAwasthen isolated,and its genomic originwasidentified bySouthern
hybridization tomapped restriction fragments ofXDNA. Analysis ofsuch Southern blots revealed that all regionsoftheAgenome wererepresentedamongthe smallDNA fragmentsgeneratedduring all timesofAr+
bombardmentexamined. Dependingontheduration ofexposure,however, particular regions of thegenome werefoundtobe enrichedin thesmall-fragment population.After short periods ofexposure, sequencesfrom
the leftmost 10% and from the right half of the standard genetic map were enriched in the broken-DNA
fraction. Amongsequencesintheright half of thegenome,the enrichmentwasprogressivelymorepronounced
beginning inthe middle ofthegenetic map andproceeding toward the right end. In phage bombarded for longerperiods of time, rightwardsequences werepreferentiallydepletedinthesmall-fragment population. In contrast, when Ar+ bombardment wascarried outwith free ADNA rather than intactphage, small DNA
fragments arose uniformly from all regions ofthe genome at all times ofexposure examined. The results indicate that in the intact phage, DNAsequences from the right half and from theveryleftmostregionsofthe
genome havea tendencyto lie closer to the capsid than does the remainder of thegenome. Since DNA is packagedintotheprohead beginningattheleftend,ourresultssuggestthatpackagingoccursin suchaway
thatnewlyenteringDNA tends tobedisposed externallytothatpackaged atearliertimes.
DNA is found in a highly condensed state inside the capsidsofdouble-strandedDNAbacteriophagessuchasT4,
A, P22, and their relatives. The DNA concentration, for instance, has been calculated to beapproximately 0.5 g/ml
for most double-stranded DNA phages (6), and the DNA strandsarepackedverycloselytogether, thespacing being 2.6 to 2.7 nm, as determinedby small-angle X-ray diffrac-tion (7, 8). This is comparable to the value of 2.8 nm (12,
16) found for the interstrand spacing in crystalline B-form DNA.Further,DNAisvirtuallytheonlycomponent present inside the capsidof phage X; proteins make up the capsid shell,but theyare notfound in significant quantitiesinside the capsid cavity. During the viral replication cycle, free DNA isinserted intoanempty(i.e., DNA-free) proteinshell called the procapsid. A single DNA molecule enters the procapsidat aunique site called theportal vertex, and the
genomeis thenpackaged linearlyinadefined direction(1, 2, 20, 21).
Cryoelectron microscopic images of T4 and X (15) show
that in the intact phage, DNA is condensed into small domains where the DNA strandsarealignedinparallel.The domainsthemselves, however,areorienteddifferentlywith respect tooneanother in individualvirions,suggesting that one is not seeing different views ofan invariant structure, but rather individual virions differ in the location of DNA-aligned domains. The cryoelectron microscopic images, therefore, demonstrate some local strand alignment in the packaged DNA,but they do notaddress the issue of
long-range order.
*Correspondingauthor.
Conflictingresults have been obtained from studies that havedirectlytested forlong-rangeorderin packaged phage DNA. Ion etching studies of T4 and X, for example, have
indicated that thelast DNAtoentertheprocapsidliescloser to the surface of the virion than does the DNApackaged earlier (3, 5). When intact phage isphysicallyeroded inan
Ax+plasma,thelast DNApackagedisfoundtobe lostmore
readily than is the remainder of the DNA. In contrast, cross-linkingstudies have suggested a more random distri-bution of DNA in phage X. Widom and Baldwin (26) have
demonstrated thatanyregion of the Xgenomecan lie close enough to the capsid shell to be cross-linked to it by irradiation with UV light. Similarly, Haas et al. (13) have demonstrated that in the intactphage, anytworegionsofX
DNAcanlie closeenoughtooneanother tobecross-linked by bis-psoralen.
Theexperiments described herewereundertakento clar-ifythe issue oflong-rangeorderbyuseofanovelapplication
of ionetching methodology.IntactXphagewasexposedtoa low-energy (-1-keV) Ar+ plasmawhoseionsare energetic
enough to break covalent chemical bonds in DNA but not energetic enough to penetrate deeply beneath the capsid shellorto desorb (sputter) materialfromthevirus surface. Under suchconditions,DNA fromthe outermostregionsof theDNAmassisexpectedtobedamaged preferentially.We thereforeisolated thefirst-damagedDNA anddetermined its location in the phage genomebySouthern hybridization to mappedrestrictionfragmentsof wholeADNA.Specificityin the genomic location of the first-damaged DNA was
ex-pected to beobserved onlyif there exists some degree of long-rangeorderingin packagedX DNA.
2226
on November 9, 2019 by guest
http://jvi.asm.org/
MATERIALS AND METHODS
Phage growth and purification. Lambda phage was pre-pared from the temperature-inducible Eschenchia coli lambda lysogen c1857 GM119 (Sigma Chemical Co.), using a procedure similar to that described by Maniatis et al. (19). In a 2-liter flask, 300 ml of LB broth was inoculated with an overnight culture of lambda c1857 GM119 and incubated at 34°C until the culture reached an optical density at 630 nm of 0.9.Phagegrowth was induced by raising the temperature to 45°C for 30 min.Thereafter, the temperature was lowered to 37°C and growth was continued for 3 h. Cells were then pelleted, resuspended in 10 ml of SM buffer (10 mM MgSO4, 10 mM Tris [pH 7.5]), and lysed by addition of 0.5 ml of CHCl3 and 2 ,ug of DNase per ml. This crude lysate was
clarifiedbycentrifugation for 5 min at 12,000 xg in a Sorvall SS-34 rotor, and phage were purified from the lysate by two steps of CsCl density gradient ultracentrifugation (19). The band of purified phage was removed from the gradient, dialyzed extensively against SM buffer to remove CsCl, titered byplaquingon a permissive host (E. coli C600), and diluted to
1012
PFU/ml with SM buffer for use in the experiments described below. Immediately prior to use, phage suspensions were made 0.2 mg/ml in bovine serum albumintofacilitate spreading of virions onto plastic cover-slips. 32P-labeled X phage was prepared as described above except thatprior to phage induction, 5 mCi ofH332Po4 was added to 100 ml of culture.Ar+ bombardment. All phage and A DNA specimens were subjected to Ar+ bombardment after freeze-drying onto plastic coverslips (2.2 by 2.2 cm; Fisher Scientific catalog no. 12-547). For Ar+ exposure of intact phage, each
cover-slip was spread evenly with 10
RI
of SM buffer containing1010 phage (enough to cover -13% of the total coverslip surface) plus 0.2 mg of bovine serum albumin per ml and incubated for 10 min in a moist chamber to allow phage to adhere. Coverslips were then washed briefly (twice) in 10 mMammonium acetate-0.1 mM magnesium acetate, frozen rapidly in liquid nitrogen, and dried in vacuo. Promptly after drying, coverslips were placed virus side up in half of a 100-mm-diameter plastic petri dish and exposed to an Ar+ plasma as described below. DNA specimens (0.1 ,g per coverslip) were prepared for Ar+ bombardment in the same way except that coverslips were glow discharged prior to
beingspread with DNA and the washing steps were omitted. Ar+bombardment of freeze-dried specimens was carried out inamodified Polaron E5100 sputter coater whose properties have been described recently (5). Plasmas were produced in a cylindrical chamber 15 cm in diameter between two disk-shaped aluminum electrodes 14 cm in diameter and separated by4cm. Plasmas weredeveloped at a current of 1 mAin 100% Ar at a pressure of 100 mtorr (13.33 kPa). The expected depth of ion penetration (2.2 to 4.4 nm) was derived from the results of a study (22) in which ions with energies comparable to those employed here were used to bombard palmitic acid multilayers containing a radioactive layer buried at different depths beneath the surface of the overall multilayer. The ion penetration depth (range) was consideredtobe the thickness of thenonradioactive palmi-tate cover required to protect the reference (radioactive) layer temporarily from damage by energetic ions.
DNAisolation and Southern hybridization. DNA was iso-lated from coverslips containing Ar+-exposed phage or phage DNA by extraction overnight at 4°C in extraction buffer (6 M guanidine-HCl, 5 mM dithiothreitol, 10 mM
Tris-HCl [pH 7.5]; 0.3 ml per coverslip). Control studies
with radioactive phage or phage DNA demonstrated that this procedure resulted in solubilization of >92% phage DNA and >95% free DNA. Further analyses were performed with DNA pooled from threeidentically treated coverslips. DNA was recovered from extraction buffer with GeneClean (Bio 101, La Jolla, Calif.), subjected to electrophoresisovernight
on a 0.85% agarose gel (20 by 25 by 0.5 cm) in TPE buffer (19) (2 V/cm) with molecular weight markers derived from HindIII-digested X DNA, and visualized by staining the gel with 0.5,ug of ethidium bromide per ml. DNA fragments 1 kb or smaller were excised from each lane of the gel and prepared for use as probes in Southern hybridization exper-iments. DNA was isolated from gel slices with GeneClean and determined quantitatively with DNA Dipsticks (Invitro-gen, San Diego, Calif.). Aliquots of 10 ng were copied into radioactive form, using the random priming method(10) with
[a-32P]dCTP(3,000 Ci/mmol; Amersham,Arlington Heights, Ill.) as a source of radioactive label. Probe specific activities were in the range of 5 x 105 to 1 x 106 cpm/ng for experimental samples from Ar+-bombarded materials and 5
x 106 to 8 x 106 cpm/ng for unexposed whole A DNA. Southern hybridizations were carried out with DNA frag-ments produced by digestion of K DNA with each of eight restriction endonucleases,AccI, BclI,BglI, EcoRI,HindIll,
MluI, NdeI, and NruI. Two 0.3-,ug aliquots of A DNA were digested to completion with each of the eight enzymes, and the digests were subjected to electrophoresis on 0.85% agarose gels, as described above, in two identical blocks of eight lanes each. After electrophoresis, DNA was vacuum transferred from agarose gels onto supported nitrocellulose membranes (BAS-NC; Schleicher & Schuell, Keene, N.H.) for 4 to 6 h at a pressure of 30 cm of H20 in an LKB Vacugene apparatus. Vacuum rather than capillary transfer wasfound to be essential to achieve the degree of uniformity of DNA transfer required for the experiments described below. When transfer was complete, the DNA was fixed to the membranes by baking at 80°C in vacuo for 1 h. The membranes were then cut in half to produce sets of two identical blots. The two blots were hybridized with probes prepared, respectively, from Ar+-bombarded specimens or from whole A DNA as described above. All hybridizations and washings were performed in screw-cap glass tubes (38 by 200 mm; Bellco Glass Co., Vineland, N.J.) in a rotating incubator set at 3 rpm. Hybridizations were carried out at 68°C overnight in 10 ml of hybridization buffer (19) (5x
SSPE [5 xSSPE is 0.9 M NaCl, 50 mM NaH2PO4, and 5 mM EDTA, pH 7.4], 5 x Denhardt's solution, 0.5% sodium dodecyl sulfate [SDS], 150 ,ug of salmon sperm DNA per ml, 10%dextran sulfate).Thereafter, blots were washed twice at highstringency (30 min at 68°C in O.lx SSPE-0.1% SDS), wrapped in Saran Wrap, and subjected to autoradiography with Kodak XRP film, using Cronex Hi-Plus intensifying screens.
Dataanalysis.Autoradiograms were digitized by scanning in an LKB UltroScan XL laser densitometer, and the resulting curves from experimental and control lanes (for each restriction enzyme digest used) were scaled to one another by a best-fit approach. Individual bands were then integrated by using a first-order Gaussian curve fit (with the LKBGelscanprogram, version 1.2), and for each restriction fragment, the integrated area in the experimental band was expressed as a fraction of that in the control. At 1-kb intervals along thegenome, an average experimental/control ratio was then computed from the relevantrestriction frag-ments in asmany as possible of the eight restriction digests;
on November 9, 2019 by guest
http://jvi.asm.org/
9.3a..&...
6.6.&E_
4. 4-_l
2.3-_
[image:3.612.95.287.78.280.2]2.-c
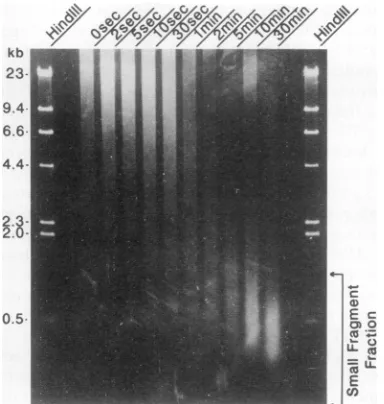
FIG. 1. Agarose gelelectrophoresis of DNA isolated from Ar+-bombarded phageX.Intact phagewasexposedtoanAr+ plasma for various timesasindicated above the lanes. DNAwasthen isolated from the phage andsubjectedtoelectrophoresison a0.85%agarose
gel beside molecular weight markers derived from HindIll-digested
XDNA.The region of the gelindicated bythearrows,
correspond-ingtothe small-fragment fraction (1kb and smaller), wasexcised
from each lane and used as a probe in Southern hybridization experiments.
thisaverageratiowasplottedasafunction ofposition along the Xgenome.
RESULTS
The experimental strategy used in this study depends criticallyon thepropertiesof theAr+ plasma.As indicated above, the appropriate plasma must contain ions able to break covalent chemical bonds in DNA but not able to penetratedeeplybeneath the virion surfaceor desorb DNA fragments.Inpractice,aplasma developedat acurrentof 1 mAwasfoundtohave the desired properties. Newcomb et al. (22)showedthat ions in suchaplasma penetrate biolog-ical materials to maximum depths of 2.2 to 4.4 nm or
approximatelyoneto twolayersof DNA(cf. Xphagehead
diameter of -65 nm
[14,
15]). When X phage containing32P-labeled DNAwasexposedtoa1 mAAr+ plasmaunder theconditionsused in thisstudy, no statistically significant lossof32plabelwasobserved after30minofbombardment (datanotshown). Similarly,noevidence of surface erosion was detected when X phage was bombarded for 30 min,
shadowed with Pt-C, and examined in the electron
micro-scope(datanotshown).
Figure 1 shows the results obtained when DNA was
isolated fromAr+-bombarded phage andsubjected to elec-trophoresis on 0.85% agarose gels. At all times of Ar+
exposuretested,DNAwasfoundtomigrateinabroadband
or smearoffragments that spannedawide rangeof
molec-ularlengths, e.g.,48to <3kb inphagebombarded for 10s.
The midpoint of thesmear, however, shifted to a
progres-sively lower molecular weight with longer durations of
exposure, suggesting that smaller DNA fragments were
beingproducedasphageabsorbedmorehits fromenergetic Ar+ ions. The smallest DNAfragments migrated inaband thatwas centered at -500bp and spannedthe range from
A B N
13- qw
6.9
35
3.5-
2.1-a-w___ -23
A,¢;
O
*_i i_ -9.4
-.
*Ail
_ -3.6
| -1.7
FIG. 2. Southern hybridization analysis of smallDNA fragments produced during exposure of phage X to an Ar plasma. Blots containing mapped restriction fragments of X DNA were probed with 32P-labeled whole X DNA (A) or 32P-labeled small DNA fragments (1 kb and smaller) produced by subjecting lyophilized phageX toanAr+plasma for30 s(B). Numbersatthe left and right indicate molecularsizesof selected bandsinkilobases.
-90 to -1,000 bp. Thirty seconds was the shortest Ar+ exposure time that produced DNA fragments migrating in thissmall-fragment fraction. After30 minofbombardment,
however, virtually all DNAwasfound in this region ofthe
gel.
For purposes of the studies described below, damaged DNAisdefinedoperationallyasthat migratingin the small-fragment fraction. This material was isolated from
Ar+-bombarded phage and radioactively labeled (10), and its
genomic origin was determined by using it to probe a
Southernblotcontaining mapped restriction fragments ofX DNA. As acontrol, free A DNAwasbombarded with Ar+ underthesameconditionsused forphage, and theresulting small-fragment DNA was subjected to the same analysis (i.e., as aprobein aSouthern hybridization experiment).
Figure2Bshows theautoradiograph obtainedwhenablot wasprobed with small-fragment DNAisolated from phage
that had been bombarded with Ar+ for 30s. Asister blot
probedwith wholeXDNAisshown inFig.2A.Thegenomic locations ofmajorrestrictionfragmentsareindicatedinFig.
3.Bycomparing Fig.2A andB,itcanbeseen at aqualitative
level that all restriction fragments labeled by the whole X
Acci
LL
13kb I II 11kb I 1 6.9kb I I 5.5kbIBcII I 8.8kb 114.5kbI
BgIll
L
EcoRi
L
Hindill
L
22kb
21kb
23kb
19kb 14.6kbI 6.3kb I I I
13kb I 11 9.7kb
I4.9kbI 5.6kb I 7.4kb I 5.8kb I
I I 1 9.4kb 11 6.6kb 14.3kbI
Miul
115.lkb 9.8kb I 11INdel L
28kbNrul 14.6kbI
26kb
1 13.Skti | l 1 8.4kb I 13.7ktlI 9.4kb I 6.7kb |
FIG. 3. A DNArestrictionmap fortheeightrestrictionenzymes shown inFig.2.Numbers indicate theapproximatemolecular sizes of the largerbands.
I
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.329.565.78.218.2] [image:3.612.326.567.546.696.2]13 k
0~~
Migration
-FIG. 4. Laser scans of theAccI lanes shown in Fig. 2. 23p_ labeledDNAprobeswerederivedfromAr+-bombarded phage (A) orwhole A DNA(B).Numbers indicatetheapproximate molecular sizesofthemajor restrictionfragments.
probe were also labeled by the probe prepared from ion-bombarded phage. Quantitatively, however, significant dif-ferences were found to exist in the intensity with which corresponding bandswere labeled. For example, the AccI 5.5-kb and the HindlIl4.3-kb fragmentswere more heavily
labeled in the experimental blot than in the control blot,
while the opposite patternwas observed in the case of the
MluI 2.2- and 2.0-kb fragments. Such quantitative differ-ences were seen more clearly when autoradiographs were digitized by scanning in a laser densitometer and
corre-sponding traces were scaled to each other. An example of suchananalysisis shown inFig. 4for the AccI lanes inFig.
2. Here itcan be seen that the 5.5- and 6.9-kb bandswere labeledmore heavily bytheexperimental probethanbythe controlprobe, while the extentsoflabeling of the 3.5- and 13-kbbandswere more nearly equal.
Tointegrate information from alleight restrictiondigests,
the datawere processed asfollows: (i) all autoradiographs
weredigitized,andcorrespondingtraces werescaled; (ii)for eachlabeledband, theintegrated intensity (peak area) inthe
experimentalblotwasdividedbythecorrespondingvalue in thecontrol; and(iii)the latterratiowasaveragedamongthe
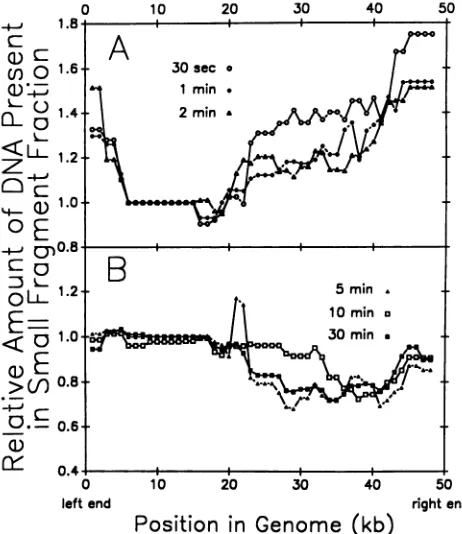
eightrestrictiondigestsat1-kbintervalsalongthe A genome, and the valuewasplottedas afunction of genomepositionas shown in Fig. 5. Such plots were produced for all Ar+ exposure times atwhich recovery of DNA from the small-fragment fractionwasabovebackground (i.e., .30 s).
Theplotswerefoundtobe oftwodifferent types, depend-ingonwhetherphage were exposed to Ar+ for short (30 sto 2min;Fig. 5A)orlong(5to30 min;Fig. SB)times. Inphage bombarded for shorttimes,thesmall-fragment fractionwas
systematically enriched in sequencesderived from theright
half of the standardgenetic map and in sequences from the leftmost -5 kb of the genome. Among the rightward se-quences,enrichmentwasprogressively greaterbeginningat
approximately themiddle of the genome andproceedingto theright end. The highest experimental/control ratios were -1.75 (Fig. 5A). In contrast, when phage were bombarded with Ar+ for 5 min orlonger, sequences from the left half of the genomewereenriched in thesmall-fragment population while rightward ones were depleted. Enrichment was
rea-sonably uniform over the leftmost half of the genome.
0 10 20 30 40 50
leftend rightend
[image:4.612.72.278.75.247.2]Position in Genome
(kb)
FIG. 5. Genomicorigin ofthe small DNAfragments produced duringAr+ bombardmentofphage X. The relativecompositionof the smallDNAfragment fractionwasdeterminedby laser densito-metryofautoradiograms from Southern blots suchasthose shown in Fig. 1. The integrated intensity (peak area) of each labeled restriction fragment was expressed as a ratio to the control and plotted with respect to genomic position. Points on the plots represent averages oftheexperimentally determined ratio forthe restriction fragments that overlapped a given 1-kb interval of the genome. (A) Relative composition of the small DNA fragments generated byexposing phageto anAr+plasmafor 30 s, 1min,and 2 min. (B) Relative composition of the small DNA fragments generated byexposing phageto anAr+plasmafor5, 10, and 30min.
Quantitatively, the degree of enrichment in leftward
frag-ments was slightly less than that observed for rightward speciesat shortAr+ exposure times.
A quite different pattern oflabeling was observed when free X DNA rather thanintact phagewas subjected to Ar+ bombardment. In this case, thesmall-fragmentfractionwas foundtocontainanearlyrandom subset of theXgenomeat allAr+ exposure times tested.Figure6showsrepresentative
results obtained when DNAwasbombarded for short(30 s)
and long (30 min) times. In studies with free DNA, the
experimental/control ratiowasfoundtobewithin the range of 0.91 to 1.06for all fragmentstested (i.e., in eight restric-tion endonuclease digests). Thecomparable rangefor stud-ies with intactphage was 0.68 to 1.75.
DISCUSSION
The purpose ofthisstudywas todetermine whether there existslong-range orderingin the arrangementof DNA in the head ofbacteriophage Xand,if so, toconsider theway(s)in which it may arise. Our experimental approachwas to use
energetic Ar+ ions, as found in ion plasmas, to introduce breaks selectively into the outermost strands of the pack-agedDNAmass.The first DNAdamagedwasthenisolated,
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.317.548.78.345.2]
cD-
4Q)->0
ci
0 10 20 30
1.8 l l l
30
seconds
1.6
Intact Phage
1.4
,r
1.2
1.0 *
Fr>,
eeA
U.0
1.2
1.0
0.8' 0.6'
nA
0
left end
1i0 20 30
Position
in
Genome
(
FIG. 6. Genomic origin of the small DNA frag
during Ar+ bombardmentof freegenomic DNA
radiograms werescanned bydensitometry and qu
scribed in Materials andMethods. (A)Relative co: small DNA fragmentpopulation produced byAr freeA DNA for 30 s compared with that prod bombardmentof intact phage for thesametime;(B)
freeDNA andphage exposedtotheAr' plasmaft
and itsoriginalong theA genome wasidentific
hybridization experiments. The results der
DNAsequencesfrom all regionsof thegenon
broken-DNA fraction. Thiswasthecasewith
damaged by the gentlest Ar+ bombardment breaks inDNA. Therefore,we conclude tha
the A genome can reside in the external j
packaged DNAmasswithin the range(one t( DNA) of the Ar+ ions used in this study analysis of the Southern hybridization expe
ever, revealed that allA DNA sequences we
represented inthefirst-damaged population. S therighthalf of thegenomeand from the lefti significantly enriched. This finding suggestec
morelikelythan the remainder of thegenom4
in the external portions of the packaged D comparable enrichment was observed wher
rather than intactphage was exposedto Ar4 that the enrichment observed is due to the DNAisarrangedin the phage headandnott greater sensitivity of rightward sequences t
energetic ions.
Whenphagewereexposedtotheion plasm 25min)rather than short periodsof time, the thedamaged-DNAfraction changed significa
sequences werefound to be depleted rather
We assume that depletion of rightward sequ
fromtheaccumulationofAr+-induceddamah most DNA in the phage head. Such accum
40 50 would alter the most external DNA too extensively for it to be copied accurately into a
32P-labeled
DNA probe. Thus, the less-damaged leftward sequences would be expected to predominate in Southern hybridization experiments as we have observed. This interpretation of our results is consis-tent with the view that rightward regions of the genome are enriched in externally disposed DNA while leftward ones are more prevalent toward the center of the phage head.Our results suggest that both ends of the genome tend to DNA be found at the
periphery
of the DNA mass. This resultagrees with
experiments involving
taillessmutantsof P2(18)
and P4 (17, 27). When DNA is isolated from these mutants, the genome is recovered as a topologically knotted structure with the cohesive ends annealed. This may indicate (2) that the two ends of the genome are positioned near one another DNA and atthe
portal
vertex. Our resultssuggest this may be the case with phage A as well. The latter conclusion was notanticipated.
Inphage
T4,
in which thefirst end of the DNA packaged is also the first end ejected (4), it is expected that both termini must be positioned at the surface of the DNA mass (1, 2, 4). However, inA,
the first end of the DNA 3ge packaged is the last ejected (9, 11, 25). It might have been anticipated that in X, the left terminus of the genome would be found at the center of the virus. Instead, both termini 40 50 appear tobe located at the periphery of the packaged DNArightend mass. Thisfinding providessupportfor theidea that there is
kb) aunitarypackaging structure utilized by all double-stranded
;ments produced DNA
bacteriophages (1,
2,
5).
Lanes of auto- The results
reported
here are inagreement
with thoseLantitated
asde- obtainedpreviously
inAr+plasma etching
studies ofphages
tmposition of the T4 and X. For example, in the case of T4, the last DNA bombardment of packaged into the prohead (which corresponds to rightward
luced by similar DNA inX [9, 11, 24]) was found to be lost more rapidly than
)similarplotsfor theremainder when the virus surface was physically eroded or 30 min. by energetic ions (3). Similarly, rightward DNA was selec-tively lost when the overall size of phage X was reduced by etching in an
Ar+
plasma (5). The present studies have added -d bySouthern the idea of a gradient of proximity to the capsid shell amongmonstrate that rightward sequences and evidence that the very leftmost ne occur in the portion of the X genome is enriched in more-external DNA.
ithe first DNA The present studies also complement previous analyses in
able to cause the way in which ion plasmas were used. In previous studies t all regions of (3, 5), the virus surface was eroded entirely (sputtered) by regions of the the action of energetic
Ar+
ions, and analyses wereper-)two layers of formed with the DNA remaining. The identity of external
z. Quantitative DNA was inferred as the difference between the whole and -riments, how- what remained after etching. In contrast, the experimental
Ire
not equally approach used in the present study does not involve physical sequences from loss of phage DNA. Rather, capsid-proximal DNA was ablemost5 kb were to be physically isolated and identified directly.
I that they are The tendency of last-packaged DNA to lie near the capsid e to be located shell as described here (and in previous studies [3,
5])
mostINA. Since no probably has its origin in the way DNA enters the phage
n free X DNA prohead. Once the head is filled, DNA strands are unlikely to we conclude rearrange significantly. Since XDNA is packaged beginning way in which at the left end (9, 11, 24), theobservedright end
external-left
o an inherently end internal polarity would be explained if DNA filled the o breakage by prohead in such a way that newly entering DNA tended to be disposed externally to that already present. The last DNA to
ka
for long (i.e., enter (i.e., the right end) would therefore be found on the composition of external surface of the overall condensate. In its pure form, ntly; rightward such an external-preferred packaging scheme must, how-than enriched. ever, constitute a simplification of the real situation. Asiences resulted shown by Widom and Baldwin (26) and by the present study,
ge
to the outer- all regions of the X genome can occur in the outermosttulated
damage portions of the mature DNA mass. As this latter observation30
minutes
4.
Free
.e
'. o .,.
IntactPhc
B
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.68.305.75.345.2]isclearly not compatible with astraightforward internal-to-external packaging scheme, weconclude that some qualifi-cation tothesimple form of the mechanism mustapply.Two possibilities suggest themselves. (i) One is to suppose that the DNA in general enters theprohead externally to DNA packagedearlier, but it is abletobecome rearranged tosome extent thereafter. The extent of rearrangement would pre-sumably decrease progressivelyastheproheadis filled with DNA. The result would be a population of mature phage withconsiderableindividualvariation in the waythe DNA is arranged. Inall,however, there would beatendency forthe
last-packaged DNA toreside nearer tothe capsidshell. (ii) Alternatively, DNA may be arranged in thephage head in such a way that it makes periodic excursions from the interior to the exterior of the overall packaged mass. This conceptis embodied in thespiral-foldandrelatedmodelsof DNA packing (3, 23). Since the X head is -65 nm in diameter, the required excursions could be accomplished with DNAlengths of <120 bp.
The existing electron micrographs of mature T4 and X preserved in the frozen-hydrated state (15) are probably
compatible with both DNA packaging schemes described above. Furtherinformation about DNA arrangementmight, however,be obtained ifone couldproduce electron micro-scopicimagesofproheads preservedin thefrozen-hydrated
state while in the process of being filled with DNA. Such
micrographsshouldatleastclarify the issue of whether DNA fills thecapsid cavityisotropicallyaspackaging proceeds or whether it is condensed in a specialized area such as the
portalvertexortheinner surface of thecapsidwall.
ACKNOWLEDGMENTS
Wegratefully acknowledge Susan Stilwell forcomputer programs and JohnBoring, CarlSchnaitman, andMitchSmith foradviceand helpful criticism.
Thiswork wassupported bygrant GM34036 from theNational Institutes of Health andby an award from the Jeffress Memorial Trust.
REFERENCES
1. Black, L. W.1988. DNApackagingin dsDNAbacteriophages, p. 321-373. In R. Calendar (ed.), The bacteriophages,vol. 2. PlenumPress,NewYork.
2. Black, L. W.1989. DNApackagingin dsDNAbacteriophages. Annu. Rev.Microbiol. 43:267-292.
3. Black, L. W., W. W. Newcomb, J. W. Boring, and J. C. Brown. 1985. IonetchingofbacteriophageT4:support for a spiral-fold model ofpackagedDNA. Proc.Natl.Acad. Sci.USA 82:7960-7964.
4. Black, L. W., and D. J. Silverman. 1978. Model for DNA packaging intobacteriophageT4heads. J. Virol. 28:643-655. 5. Brown, J. C., and W. W. Newcomb. 1986. Ion etching of
bacteriophage lambda: evidencethattheright end of the DNA is located at the outside of the phage DNA mass. J. Virol. 60:564-568.
6. Earnshaw,W.C.,andS. R.Casjens. 1980. DNA packagingby thedouble-stranded DNAbacteriophages. Cell21:319-331. 7. Earnshaw,W.C.,andS. C.Harrison. 1977. DNAarrangement
inisometricphageheads. Nature268:598-602.
8. Earnshaw,W.C.,S. C.Harrison,and F. A.Eiserling.1978. The structural organization ofDNA packagedwithin heads of T4 wild-typeisometric andgiant bacteriophages. Cell 14:559-568. 9. Emmons, S. 1974. Bacteriophage lambda derivatives carrying
twocopiesof thecohesive end site. J. Mol.Biol.83:511-525. 10. Feinberg, A. P., and B. Vogelstein. 1983. A technique for
radiolabelingDNA restriction endonucleasefragments tohigh specific activity.Anal.Biochem. 132:6-13.
11. Feiss, M., and A. Bublitz. 1975. Polarized packingof lambda chromosomes. J. Mol. Biol. 94:583-594.
12. Giannoni,G.,F.Padden,andH.Keith. 1969.Crystallizationof DNA from dilute solution. Proc. Natl. Acad. Sci. USA 62:964-971.
13. Haas,R.,R.Murphy, and C. Cantor. 1982.Testing modelsof the arrangement of DNA in phage lambda. J. Mol. Biol. 159:71-92.
14. Kemp, C. L., A. F. Howatson, and L. Siminovitch. 1968. Electron microscope studies of mutants of lambda bacterio-phage. Virology 36:490-502.
15. Lepault, J., J. Dubochet, W. Baschong, and E. Kellenberger. 1987.Organizationofdouble-stranded DNAinbacteriophages: a study by cryo-electron microscopy of vitrified samples. EMBO J. 6:1507-1512.
16. Lerman, L., L. Wilkerson, Jr., J.Venable, and B. Robinson. 1976. DNA packing in single crystals inferred from freeze-fracture-etchreplicas.J. Mol.Biol. 108:271-293.
17. Liu, L.F., J. L. Davis, and R. Calendar. 1981. Novel topolog-ically knottedDNAfromP4bacteriophage capsids: studies with DNAtopoisomerases. Nucleic Acids Res. 9:3979-3989. 18. Liu, L. F., L. Pevkocha, R. Calendar, andJ. C. Wang. 1981.
Knotted DNA from bacteriophage capsids. Proc. Natl. Acad. Sci. USA 78:5498-5502.
19. Maniatis, T., E. F. Fritsch,andJ. Sambroolk 1982. Molecular cloning: alaboratory manual. Cold Spring Harbor Laboratory, ColdSpring Harbor, N.Y.
20. Murialdo, H. 1991.Bacteriophagelambda DNAmaturation and packaging.Annu.Rev.Biochem. 60:125-153.
21. Murialdo, H., and A. Becker. 1978. Head morphogenesis of complex double-stranded deoxyribonucleic acid bacterio-phages. Microbiol. Rev. 42:529-576.
22. Newcomb, W. W., T. A. Johnston, and J. C. Brown.1987.Ar' plasma etching ofpalmitic acidmultilayers: differential erosion
ratesofexposedandprotectedlayers. Langmuir 3:1000-1004. 23. Serwer, P. 1986. Arrangement ofdouble-stranded DNA
pack-aged inbacteriophage capsids. J. Mol. Biol. 190:509-512. 24. Sternberg, N.,and R. Weisberg. 1975. Packaging of prophage
andhostDNAbycoliphage lambda. Nature (London) 256:97-103.
25. Syvanen, M. 1975. Processing of bacteriophage lambda DNA during its assembly into heads.J. Mol. Biol.91:165-174. 26. Widom, J., and R. Baldwin. 1983. Tests of spool models for
DNApackaginginphage lambda.J. Mol.Biol. 171:419-437. 27. Wolfson,J.S., G. L. McHugh, D. C. Hooper, and M. N. Swartz.
1985. Knotting ofDNAmolecules isolatedfrom deletion
mu-tantsofintactbacteriophage P4. Nucleic Acids Res. 13:6695-6702.