0022-538X/92/010150-10$02.00/0
Copyright © 1992, American Society forMicrobiology
Distinct
RNA
Sequences
in the
gag
Region
of
Human
Immunodeficiency
Virus
Type
1 Decrease RNA
Stability
and Inhibit
Expression in the Absence of Rev
Protein
STEFANSCHWARTZ,"12 BARBARAK. FELBER,3 ANDGEORGEN. PAVLAKIS1*
Human Retrovirus
Section'
and Human RetrovirusPathogenesis Group,3 National CancerInstitute-Frederick CancerResearch andDevelopment Center, ABL-BasicResearch
Program, Frederick,
Maryland
21702-1201, andDepartmentofVirology, Karolinska Institute,Stockholm,
Sweden2Received 26 June1991/Accepted 24September 1991
The expression ofGag, Pol, Vif, Vpr, Vpu,and Envproteinsfrom
unspliced
andpartially spliced
humanimmunodeficiencyvirustype 1(HIV-1)mRNAsdependsonthe viral
protein Rev,
while theproduction
ofTat, Rev, and Nef frommultiply splicedmRNAs doesnotrequireRev. Toinvestigate
thedifference betweengag and tatmRNAs, wegeneratedplasmidsexpressing tat-gag hybridmRNAs. Insertion of thegag gene downstream ofthe tatopenreadingframe in thetatcDNA resulted in theinhibition of Tatproduction.
Thisinhibition was caused, at least in part, bya decrease in the stability of theproduced
mRNA. Deletions in gag defined a 218-nucleotideinhibitorysequence namedINS-1andlocatedatthe5' end of thegag gene. Further experiments indicated the presence of more than one inhibitory sequence in the gag-protease generegion
of the viral genome. The inhibitory effect ofINS-1 was counteracted by thepositive effect mediated by theRev-Rev-responsive element interaction, indicatingthat this sequence isimportantforRev-regulatedgag expression. TheINS-1 sequence did not contain any known HIV-1splicesites and acted
independently
ofsplicing. Itwas found to have anunusually highAU content(61.5% AU),acommonfeatureamong cellular mRNAs with shorthalf-lives. These resultssuggest that HIV-1 and possibly other lentiviruses have evolvedtoexpress unstable mRNAs which require additional regulatory factors for their expression. This strategy inay offer the virus several advantages, includingthe
ability
toenter a stateof loworlatentexpressioninthe host.The genome of human immunodeficiency virus type 1
(HIV-1) contains thegag,pol, andenvgenes common toall retroviruses. Inaddition tothesegenes, the HIV-1 genome
contains several open reading frames (ORFs) encoding the
accessory andregulatoryproteins Tat, Rev, Nef, Vpu, Vif,
and Vpr. The expression ofall of these ORFs from one promoter is
accomplished
primarily by the generation of alternativelyspliced-mRNAs grouped into three size classes: full length (9kb),
intermediate (4 to 5 kb), and small,multiply spliced (2 kb). The full-length and intermediate mRNAsencoding Gag, Pol, Env,
Vpu,
Vif, Vpr, and Tat-1(21, 38, 51, 52) contain the cis-acting Rev-responsive ele-ment(RRE)(15,16,19, 25,26,36, 47).Theexpressionofthe
full-length
and intermediate mRNAs depends on the viralRev protein (15, 17, 21, 25, 51, 52, 57). In the absence of Rev, most ofthe full-length and intermediate mRNAs are
further spliced, generating the class of small, multiply
splicedmRNAs encoding Tat, Rev, and Nef (15, 17, 18, 25,
51,52). These
mRNAs
lack the RRE and are the only HIV-1mRNAsthat can beexpressed efficiently in the absence of Rev.
The Revprotein actsposttranscriptionallytoincrease the transportand utilizationof allviral mRNAs that contain the RRE (4, 16, 19, 25, 26, 36). Rev binds to RRE (5, 12-14, 27,
35, 41, 66). Several lines of evidence from in vivo experi-ments
indicate
thatthe Rev-RRE interaction, although nec-essary, is not sufficient for the activation ofRRE-containing mRNAs andthatadditional cellular factorsinteractingwith Rev arenecessary for Revfunction(1,5). ThemechanismofRev
function has been the subject of intense study. It has* Correspondingauthor.
beenshownthat Revincreasesthehalf-life of RRE-contain-ing HIV-1 mRNAs (19), promotes theirtransport from the nucleus to thecytoplasm(16, 19, 26, 36),and promotes their efficient translation(2, 4, 5, 25, 57).
Experiments with constructsexpressing mutant gag and envmRNAslacking splice sites showed that only lowlevels ofthesemRNAs accumulate in the absence of Rev andthat
theirexpression isRevdependent (19, 31, 36). Theseresults led to theconclusion that Revactsindependently of splicing (19, 36). The inability of gag and env mRNAs to be
effi-ciently expressedin theabsence of Rev ledtotheprediction
that they contain sequences other than splice sites which
inhibit their expression (16, 19, 25, 47). It was shown that env sequences lower the levels ofchloramphenicol acetyl-transferase (CAT) produced by CAT-HIV hybrid mRNAs
(47). It wasproposed that this inhibitory effectis mediated by sequences-named cis-actingrepressivesequences within the env region. The nature and mode of action of these
sequences have remained unclear. The presence of
inhibi-tory sequences thataffecttheaccumulationand utilization of gag and env mRNAs was also proposed on the basis of
measurements of viral mRNAsandproteins (17, 19, 25, 42, 57). Although it was suggested that these sequences affect thestability and utilization of viralmRNAs, themechanisms bywhich they mediate the decreased levels ofgag andenv expressionare notunderstood.
Examinationof the effects of Rev on thelocalization and
splicingofhybridglobin-RREmRNAsledto theconclusion that splice sitesarenecessaryforRev regulation (9). Itwas proposedthatregulationbyRev involves the dissociation of
splicingcomponents and pre-mRNA. It was alsosuggested thatcomplex formation between Ulsmall nuclear RNA and env mRNA is necessary for the formation of unspliced
150
on November 10, 2019 by guest
http://jvi.asm.org/
mRNA thatis subject to regulation by Rev (34). In view of
these results, the question arises as to whether the
postu-lated inhibitory sequences contain elements recognized by
the splicing machinery or whether additional,
splicing-inde-pendent mechanisms inhibit viral mRNA transport and
sta-bility.
Wedesignedexperiments to identify inhibitory sequences
within the HIV-1 genome and to study their function in
detail. Here we report theidentificationand characterization
of an inhibitory sequence in the HIV-1 gag gene. On the
basis ofthe characterization, we have named this sequence
INS-1. The relationship of INS-1 to cis-acting repressive sequences is not clear and will require additional
character-ization. The INS-1 element does not contain any functional
splice sites and acts in cis by lowering steady-state mRNA
levels. Thus, INS-1appears to function, at least in part, at
the level ofmRNA stability. This effect could beovercome
by the Rev-RRE interaction. Therefore, sequences other
than
splice
sites are important for regulation by theHIV-1Rev protein. Our experiments suggest that mechanisms
affecting mRNA stability areimportant for the regulation of HIV-1 and possiblyother complex retroviruses.
MATERIALS AND METHODS
Recombinant
plasmids.
Thefollowing
plasmidshavebeen describedpreviously:pNL1.4.7
(50) (seeFig.1B), pHCMVs-rev(5), andpHCMV-SEAP(6). To generate RRE- andgag-containing tat cDNAs, we clonedtheHIV-1 RRE upstream
of the HIV-1 gag gene in the polylinker of pBluescript KS(-) (Stratagene). The 330-bp RRE fragment from p330 (58) was cloned into theEcoRI andBamHI sites in pBlue-script KS(-). The resulting plasmid was opened at the
BamHI site and ligated to the BssHII-ApaI gag
fragment
from pMcgag (19), which was derived from the HIV-1
molecular clone HXB2. The gag
fragment
has a deletion between nucleotides (nt) 257 and 330 which removes the major 5'splice
site at nt 289 and theproposed
packaging
signal upstream of the 5'
splice
site (10,33).
Theresulting
plasmid contained the
330-bp
RREupstream ofgag inthepBluescript KS(-)
polylinker
and wasusedto generatethe various RRE-gagfragments
that were cloned into the tatcDNA
expression
plasmid pNL1.4.7. RRE-gag
fragments
weregeneratedby restriction enzyme
digestion
otby
poly-merase chain reaction(PCR) (48, 49)
withoneoligonucleo-tideprimeratthe5'endofthe RREand theotheratvarious locations in the gag gene. Thevarious
RRE-gag
fragments
werecloned intotheBamHI siteof
pNL1.4.7 (50),
resulting
in
pTRG(330-1552),
pTRG(330-1397),
pTRG(330-1174),
pTRG
(330-1053),
pTRG(330-961),
pTRG(330-691),
pTRG(330-631),
pTRG(330-480),
pTRG(330-413),
andpTRG(330-354).
Thenumbering ofthe revisedHXB2 sequence startsat thefirst nucleotide of the R
region
in the 5'long
terminal repeat(LTR) (39,45, 46, 58). Thenumbersin
parentheses
indicate the5' and 3'positions
ofthe gagfragment
containedineachplasmid.
To generate deletions in the 5' end of the gagfragment, wedigested
plasmid
pTRG(330-691)
withBamHIand
religated
it. Thisprocedure
removed thegagfragment
andresulted inpTR,
which
containedaunique
BamHIsiteimmediately
downstreamof
the RRE. The various 5' gagdeletions of the gag
fragment
from nt 330 to 691 weregenerated
by
PCR and subcloneddirectly
into the BamHIsite of pTR. This
procedure
resulted inpTRG(355-631),
pTRG(414-631), and
pTRG(481-631).
pTG(330-1552)
wasgenerated by
subcloning
oftheBssHII-ApaI
gagfragment
from pMcgag (19) into the BamHI site of
pNL1.4.7
(50).
pTG(961-1552)
was constructed by excising the 5' end ofgag in
pTG(330-1552)
with BssHIIand
PstI. To constructpNL17R,
we PCR amplified the pl7gag coding sequencefrompMcgag andsubsequently cloned itinto the EcoRV site
ofthe
pBluescript KS(-)
vector. A premature stop codonwas introduced immediately after the last codon of
p179ag
through
oneof the PCRprimers.The gagfragmentdoes notcontain the
major
HIV-1splice
donor. The 330-rt RREfragment
described above was subcloned into the Hindlllsite downstream of
p17
resulting
inpBS17R. p17 a9 and RREwereexcisedas onefragment
bydigestion
ofpBS17R
withBssHII andBamHI.TheBssHII-BamHI fragment from
pBS17R
was subcloned intoBssHII-BamHI-digested
plas-mid
pNL1.4.7,
resulting in the eucaryotic expressionplas-mid
pNL17R
(seeFig.
6A).Cells and transfections.HL3T1 cells contain silent
copies
of the HIV-1 LTR promoter linked to the CAT gene
(65).
HLtatcellsconstitutively
express thetruncated,
butfunc-tional,
Tat-1protein (50).
Thevariousamounts ofplasmids
used for transfection were
adjusted
to 17,ug/0.5
ml ofprecipitate
per 60-mmplate
(approximately
106cells).
withpUC19 carrier DNA and were transfected
by
the calciumphosphate
coprecipitation technique (23)
asdescribedprevi-ously (19, 20).
Onemicrogram
oftat-gagplasmid
was usedfor each transfection unless indicated otherwise. This amount
usually
resulted in 20 to40% transfectedcells,
asdetermined
by
immunofluorescence. Cellswereharvested20h
posttransfection.
Toquantitate
Tatproduction
from allTat-expressing plasmids,
wedetermined theamountofplas-midthatresulted inaCATresponsein the linearrangeofthe assay.All
Tat-expressing plasmids
wereserially
dilutedandtransfected into HL3T1 cells to determine the amount of
plasmid
thatresultedinaCATresponse in the linear rangeofthe assay. The CAT valueswerenormalized tothe amount
of
plasmid
used for transfection. Cotransfection with aplasmid
(pHCMV-SEAP) producing
secretedhumanplacen-tal alkaline
phosphatase
(SEAP)
(6)
wasusedas acontrol for transfectionefficiency.
Comparable
levels of SEAP wereproduced
in the different transfections.RNAextractions and Northern
(RNA)
blotting.
TotalRNA wasextracted 1day
posttransfection by
theheparin-DNase
procedure (32).
Nuclear andcytoplasmic
fractions wereseparated
as describedby
Greenberg
and Ziff(24),
and theRNA was
prepared by
theheparin-DNase
procedure
(32).
Thefilterswere
hybridized
to agel-purified, nick-translated,
PCR-amplified
DNAfragment
spanningnt 8304 to 9008 at the 3' end ofthe HXB2 genome. Measurements ofRNAstability
wereperformed
by
the addition ofactinomycin
D(final
concentration,
10,ug/ml)
to the culture medium of transfected HL3T1 cells 20 hposttransfection.
Total RNA was harvestedatdifferent times afteractinomycin
D treat-ment. The RNA blots werequantitated
with the AMBISradioanalytic
imaging
system.Immunoprecipitations,
Westernimmunoblotting,
andCATassays. At
day
1posttransfection,
HLtat cells(50)
weremetabolically
labeled with[35S]cysteine
for 3 h. The cellswere
lysed
in 0.5x RIPAbuffer,
immunoprecipitated
withanti-Tat
antiserum,
andanalyzed by
electrophoresis
on15%denaturing
polyacrylamide
gels
asdescribedpreviously
(18).
ForWestern
immunoblotting,
transfectedcellswerelysed
in0.5x
RIPA buffer andsubjected
toelectrophoresis
on 15%denaturing
polyacrylamide gels
asdescribedpreviously
(25).
Theblots werehybridized
toHIV-1
patient
serum, and theproteins
were visualized with the enhancedchemilumines-cence Western
blotting
system
(Amersham).
CAT assays wereperformed
asdescribedpreviously
(22).
on November 10, 2019 by guest
http://jvi.asm.org/
A
330 1552
l l
G
B
289
I
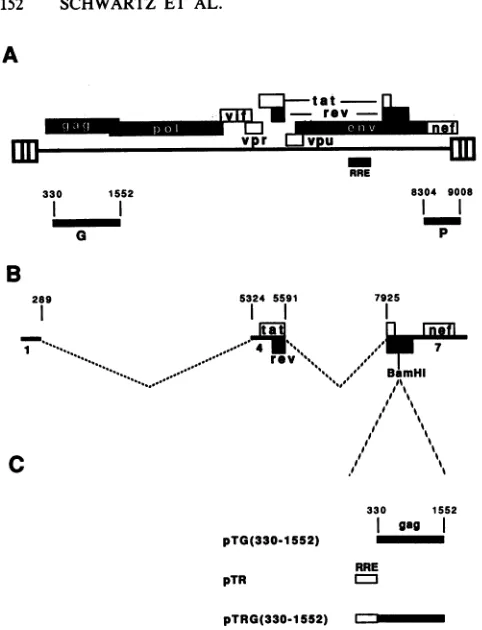
FIG. 1. (A) Structure oi black bars indicate ORFs. marked G indicates the ga
1552). The shaded barmar
Northern blotanalysis. (B) nucleotidepositionsof the numbering of the revised s
HXB2 used inourexperim4
region in the 5' LTR (39, describedby Muesingeta] coding regions fortat,rev,; BamHIrestriction site into
gagfragmentsweresubclor cloned into tat cDNA pN1 (330-1552),andpTRG(330-] fragment containing theRE fragment spanningnt330t4
Insertion ofgagsequer tion of Tat. Studies of shown thatgag cannot
(17, 19, 25, 57). Incontr
as tat mRNA 1.4.7 (Fig absence of Rev. To dete
adominant inhibitoryeff
wegenerated plasmids e hybrid mRNAs. Three
pNL1.4.7 were constru both RRE and gag seqi
downstream of the tat pTG(330-1552), pTR, a
and are shown in Fig indicate the gag sequen gag fragment was inser theseplasmids.Thegag
tatAUG hasa strong consensus sequencefor translational initiation which efficiently blocks the expression of down-tat stream ORFs (50). The RRE was inserted upstream of the ;[vitltrev - gag fragment. The exact position of these elements in
relationtoeach otherwasnotfoundtobe
important (see
also vprVPU
DI Fig. 6, pNL17R). Several investigators have shown that the RRE RRE retains activity when placed at different locations 8304 9008 within the transcription unit (19, 36). Tat production from l l theseplasmidswasanalyzedandquantitated bytransfection p into HL3T1 cells, which contain silent copies of the HIV-1 LTR promoter linked to the CAT gene. Upon transfectionwithaTat-producing plasmid, suchas tatcDNA
pNL1.4.7,
5324 5591 7925 CAT expression was activated in a dose-dependent manner l l l (Fig. 2A, inset). Therefore, under these conditions, then [nefl levels of CAT
activity
reflect theamountsof Tatprotein.
mvI*TTo assess Tat production from pTG(330-1552), pTR, and
rev ,. BBamHl
pTRG(330-1552),
wetransfected theseplasmids
into HL3T1 cells in the absenceorpresence ofRev-expressingplasmid pHCMVsrev (5)and determined the levels of CAT activation(Fig. 2A). Tatproductionwas comparedwith that obtained , ' fromwild-type tat cDNA pNL1.4.7. Thisplasmidexpressed
.0'
',high
levels ofTat and wasnot affectedby
the presence of Rev. In theexperimentshown inFig. 2A, pTR expressedTat 330 1552 at almost the same levels aspNL1.4.7, although
a smallpT32gag
inhibitory
effect of the RRE could be observed in severalpTG(330-1 552) other
experiments.
Coexpression
of Rev increased Tat pro-RRE duction from pTR slightly, furtherindicating
that the RRE pTR exerted a small inhibitory effect on Tat expression underthese conditions.
Interestingly,
Tatexpression
from pTRG(330-1552)pTG(330-1552)
was 10-fold lower thanfrompNL1.4.7,
indi-f the HIV-1 genome. Open, shaded,or cating that the presence of gag sequences substantially
The RRE is indicated. The black bar inhibited Tat expression. Rev did not affect Tat expression Ig fragment that is present in pTG(330- from the former plasmid, since it did not contain the RRE. A
rked
P indicates the probe used for the similar inhibitory effect on Tat production was observed exons in tat mRNA are indicated. The when pTRG(330-1552), containing both gag and RREse-sequence ofthe HIV-1 molecular clone quences, was transfected into HL3T1 cells. Tat expression
ntsstarts atthef
fIrst nuleotide
ofeR
from this plasmid was elevated in the presence of Rev,,45, 46, 58). The exons are named as
demonstrating
that theinhibitory
effect exertedby
the gag1. (38) andSchwartz et al. (50, 51).The sequencescould beatleast
partially
overcomeby
Rev.andnefare indicated. The location of the To verify that CAT protein levels reflected the levels of which the variousRRE, gag, or RRE- Tat protein, we also analyzed the production of Tat from ied is indicated.(C) Fragmentsthat were pNL1.4.7 and pTRG(330-1552) directly by immunoprecipi-1552j. Open bars indicatethe 330-nt
Styp
tation ofTat. pNL1.4.7
andpTRG(330-1552)
were trans-{E(58),and blackbars indicate the3 ag fectedinto
HLtat cells, a HeLa-derived cell line constitu-o1552. tively producing the functional truncated one-exon Tatprotein
Tat-1.This 14-kDaprotein
couldbedistinguished
insize fromthe
complete
16-kDaTatprotein
(Tat-2)produced
RESULTS by tat cDNA pNL1.4.7 and its derivatives.Radioimmuno-precipitation
with anti-Tat antiserum demonstrated thaticesintatmRNAinhibitstheproduc- pNL1.4.7 expressed
high
levelsofTatboth in the absencethe
expression
of HIV-1 gag have andin the presence ofRev (Fig. 2B). Tatproduction frombe
expressed
in the absence of RevpTRG(330-1552)
was undetectable in the absence of Rev,rast, multiply spliced mRNAs, such while cotransfection with Rev-producing pHCMVsrev
re-. 1), areexpressed
efficiently
in the sulted inhigh levels ofTat. The resultsofthese experimentsrminewhethergag sequences have were in agreement with the CAT measurements (Fig. 2A),
rectontheexpression oftatmRNA, verifying that the expression of Tat from pTRG(330-1552)
-xpressing
tat-gag andtat-RRE-gag could be substantially increased by the Rev-RREinterac-different derivatives of tat cDNA tion.
cted and contained gag, RRE, or Deletion mutant construction and analysis. To define the uences inserted at the BamHI site location of inhibitory sequences in gag more precisely, we
ORF. These
plasmids
were named generateda series of progressive deletions in the 3' end ofLnd
pTRG(330-1552), respectively, the gag fragment in pTRG(330-1552) (Fig. 3A). Tatproduc-. 1. The numbers in parentheses tion from each mutant was assessed following transfection
ices inserted in the construct. The into HL3T1 cells. Cotransfection with aplasmid (pHCMV-ted downstream ofthe tat ORF in SEAP) producing SEAP (6) was used as a control for
ORFcannotbe
translated,
sincethe transfectionefficiency.
The results of the CATmeasure-1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.54.295.57.372.2]50
N
cmJ
I
-0U 25
ng pNL1.4.7
0
*
Ui
)cm C CM
CLCL gnI- inV-m~~~~C
0. 0.
O O ~~~0.
B
Rev: l
-+
- +1l..u4
_ _
-
Tat-2- Tat-2
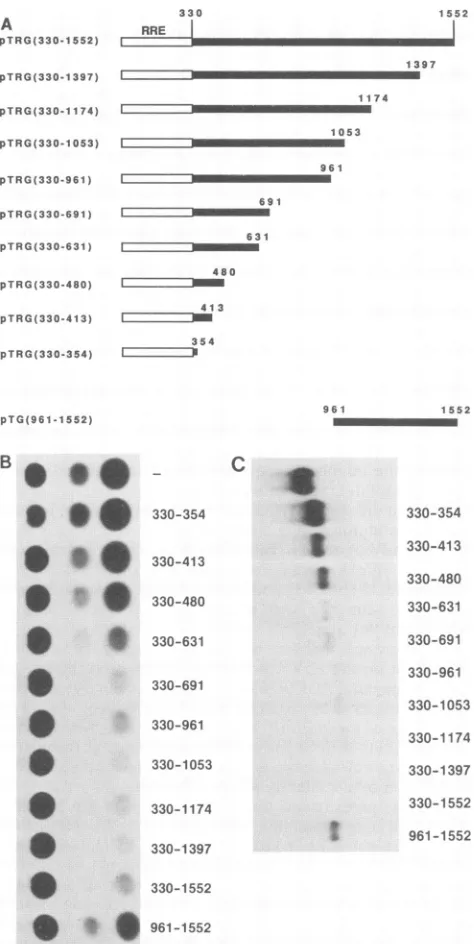
ments are shown in Fig. 3B. The plasmid containing the smallest gag fragment that resulted in a significant inhibitory
effect was pTRG(330-631). There was a gradual decrease in inhibition when smaller gag fragments were used (see below and Fig. 3C). These results defined the 3' border of the inhibitory sequence of gag as nt 631.
gag sequences inhibit Tat expression by decreasing the steady-state levels of tat mRNAs. To investigate the
mecha-nism of function of the gag inhibitory sequences, we also
studied the expression ofthe various plasmids at the RNA
level. Northern blotanalysis demonstrated that
gag-contain-ingtatcDNAs thatexpressed low levels of Tat protein also
expressed lowlevelsofmRNA (Fig.3C). Quantitation of the mRNA levels revealed that low Tat production could be 300 explained to a large extent by low mRNA levels. The RNA
results alsodefined the3' borderof the
inhibitory
sequence as nt 631, in agreement with the results of the protein measurementspresented in Fig. 3B.To investigate whether other inhibitory sequences were present in the 3' end of the gag gene, we deleted the sequencesfrom nt 330 to 961 in pTG(330-1552), resulting in
pTG(961-1552) (Fig. 3A). Analysis of the expression of
proteinand RNA from pTG(961-1552) revealed that higher
protein andRNA levels were detected with this plasmid than
with pTRG(330-1552) (Fig. 3B and C), demonstrating that
Itto
some of theinhibitory
sequences had been deleted.How-ever, pTG(961-1552) produced lower protein and mRNA
levels thandidtatcDNA pNL1.4.7 (Fig. 3B and C), indicat-ing that additional sequences in the 3' end of gag also affected mRNA levels. Taken together, these results dem-onstrated that a strong inhibitory sequence was located between nt 330and631and thatanindependent butweaker
inhibitory sequence was present in the 3' end of gag, between nt 961and 1552.
To determine whether the inhibitory effect of gag was
dependentonthe HIV-1 promoter, we inserted the tat-gag
DNA
fragment
TRG(330-691) 3' tothe humancytomegalo-viruspromoter. Theresulting plasmids produced onlyvery + low levels oftat-gag mRNA in the absence of Rev, while
cotransfection withRevresulted inanincrease inthe mRNA levels (datanot shown), demonstrating that the INS-1 ele-m mentacted independently of thepromoter.
o We next
investigated
the effect ofRev on the levels ofcn RRE- and
gag-containing
tat mRNAs. RNA productionfrom three plasmids, pTRG(330-354), pTRG(330-691), and
pTRG(330-1552), which contain gag fragments ofdifferent
lengths,wasquantitatedasdescribedinMaterialsand
Meth-ods in the absence orpresence of
Rev-expressing plasmid
pHCMVsrev
(Fig.
4).pTRG(330-354) produced
high
levels of RNA whilepTRG(330-691)andpTRG(330-1552) producedFIG. 2. (A) CATactivitymeasured aftertransfections ofHL3T1 cells with the various plasmids indicated at the bottom of the histogram.Eachplasmidwastransfected in the absence(-)orthe presence(+) ofRev-expressingplasmid pHCMVsrev.CATactivity
measured after transfection withpNL1.4.7was set at100%.(Inset)
Titrationexperimentsverified that the CAT responsewas propor-tionaltothelevelofTat-producingplasmid. Cellsweretransfected with the indicated concentrations ofpNL1.4.7, and the produced
CAT protein was quantitated. (B)
Immunoprecipitations
of Tat proteinfrom cells transfected withpNL1.4.7orpTRG(330-1552)
in theabsence(-)orthe presence(+)ofRev.Tat-2is thecomplete
16-kDaTatprotein producedfrompNL1.4.7and
pTRG(330-1552).
Tat-1 is thetruncated 14-kDaTatprotein
constitutively
produced by
HLtatcells.
A
1oO
-o o
50
+
t-
tI-it qi.
-1
z z
0. 0.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.67.307.84.721.2]A
pTRG(330o1 552) pTRG(330-1397) pTRG(33011174)
pTRG(330-1 053)
pTRG(330-961)
330
RRE
1552 I
13g97
1 174
10 3 1.
9g61
691
pTRG(330-69 1)
6 31
pTRG(330-631)
48 0 pTRG(330-480) I
413
pTRG(330-413) Iz
-3 54
pTRG(330-354) ._
pTG{961 -1552)
B is
*
it
C
330-354
330-413
330-480
a
...s
II
I
330-631
.
.
0
330-691
330-961
330-1053
330-1174
330-1397
330-1552
[image:5.612.360.501.84.210.2]'T 961-1552
FIG. 3. (A) Schematic structures of the containingdifferentdeletionsatthe 3' endofgd
plasmids are showntothe left.Open bars ind fragment containing the RRE, and black ba fragments. The nucleotide positions of the fragmentsareindicated.(B)CAT activity inext
transfected with the different plasmids indica numbersindicatethegag sequencescontainec plasmid. The minus sign indicates transfecti pNL1.4.7,which didnotcontainany gagsequ blotanalysis of total RNApreparedfromHL
withthe differentplasmids shown inpanel A.T thegagsequencescontained ineach transfecte signindicates transfectionwithtatcDNA pNL containany gag sequences.
Rev:
F[
+-
F
1
+
r~~~~~~~~~~~~
FIG. 4. Northern blot analysis of total RNA prepared from HL3T1 cellstransfected with theplasmidsindicatedatthetop. The
plasmidsweretransfected in the absence(-)orthepresence(+)of pHCMVsrev.
verylow mRNA levels in the absenceof Rev(Fig. 3C and 4). Coexpression of Rev had a small effect on the RNA levels 6 1 15 5 2 produced frompTRG(330-354). Incontrast, Rev resulted in a marked increase (5- to 10-fold) in the mRNA levels produced from pTRG(330-691) andpTRG(330-1552).
There-fore, Rev overcomes the inhibitory effect of the gag se-quences by increasing the steady-state levels of mRNAs 330-354 containing both RRE and gag sequences.
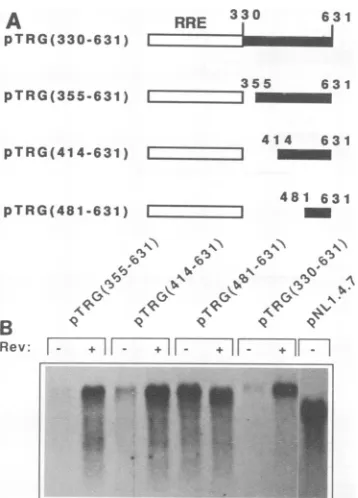
Aninhibitory element is located between nt 414 and 631 in 330-413 the p17gag gene. To define the shortest inhibitory sequence
330-480 within the proximal part of the gag gene, we constructed
|
330-480
progressive deletions at the 5' end of the gag fragment in330-631 pTRG(330-631) (Fig.
5A).
Thisplasmid contained the short-est gag sequence thatdisplayed a full inhibitory effect, as330-691
determined by the 3' deletions (Fig.3C).
Three different330-961 5'-end deletions were generated, resulting in pTRG(355-631),
pTRG(414-631), and pTRG(481-631) (Fig. 5A). These plas-330-1053 mids were transfected into HL3T1 cells in the absence or presenceofRev-expressingplasmidpHCMVsrev, andtheir 330 1174 phenotypes were determined by Northern blot analysis.
330-1397
Only pTRG(481-631)
expressed high
levels ofmRNA both in the absence and in the presence ofRev, demonstratingthat 330-1552 a larger fragment of gag was required for inhibition (Fig.a
SB).
pTRG(355-631)
andpTRG(414-631)
displayed
thesame961-1552 phenotype as parent plasmid pTRG(330-631). Efficient
expression ofthese twoplasmids required the presence of
Rev. Therefore, pTRG(414-631) contained the shortest gag
fragment that elicitedastronginhibitory effect.
The Northern blots were probed with a fragment that
hybridized to the 3' endof the mRNAs (Fig. 1A) and that RRE-gag fragments should detect all mRNAs. Since only the expected full-ag. The names of the
length
mRNAwasproduced
from eachexpression
plasmid,licate the 330-ntStyI efficientsplicing of thetat-RRE-gagmRNAsdid not occur. rs indicate the gag In conclusion, these experiments identified a 218-nt cis-3' ends of the gag acting element in gag (nt 414 to 631) named
INS-1. INS-1
tracts of HL3T1cells actedindependently of splicing and lowered the steady-state ted in panel A. The levels ofgag-containing tat mRNAs.ion with tat cDNA The expression of
p17gag
protein is Rev dependent. Theences. (C) Northern experiments described above identified an inhibitory RNA
3T1 cells transfected element contained within the coding regionof
p7
gag. The'henumbersindicate data
predicted
thatp7
gag protein couldnotbeexpressed
in dplasmid. The minus the absence of theRev-RREinteraction.To testthishypoth-1.4.7,which didnot esis and to verify the results obtained with tat-gag hybrid constructs, weanalyzedRNA andprotein production from a
pl7gag
expression plasmid. We cloned thep179ag
codingsequence immediately followed by a terminator codon
to-9
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.58.295.100.572.2]A
RRE 330 631pTRG(330-631) El
35 5 6 31
pTRG(355-631) -61 II
414 631
pTRG(414-631) IE I _
pTRG(481-631) I I
N>"N lb'
C.
B
Rev:
NI
4 8 1 63 1
1N>
S g, -A t.
Ve
I +
I
+1 ''' - +r--FIG. 5. (A) Structures of the RRE-gag fragments containing different deletions at the 5' end of the gag fragment present in plasmid pTRG(330-631). Thenamesof the plasmidsareshowntothe left. Open bars indicate the 330-nt RREfragment, and black bars indicategagsequences.Thegagregion nucleotidesincludedin each
plasmid are indicated. (B) Northern blot analysis of total RNA
prepared from HL3T1 cells transfected withtheplasmids indicated atthetop.Thetat-gagplasmids containing the variousgagdeletions
shown inpanelAweretransfected into HL3T1 cells in the absence
(-)orthepresence(+)of Rev.
gether witha330-nt RRE fragmentdownstream of the HIV-1 LTRpromoter.This procedure resulted in plasmid pNL17R (Fig. 6A). This plasmid wastransfected into HLtat cells in the absence or presence of plasmid pHCMVsrev. RNA analysis of the transfected cells revealed thatverylow levels ofp179a9mRNAwereproduced in the absence of Rev, while high mRNA levels were detected in the presence of Rev (Fig. 6B). The levels of p179a9 mRNA produced in the presence of Revwerecomparableto those produced bytat cDNApNL1.4.7. To assess the production ofp179a9from
pNL17R,wesubjectedextractsoftransfected cellsto West-ernblotanalysis. Very low levels ofp179a9weredetectedin the absenceof Rev, while cotransfection withpHCMVsrev resulted indramatically increased levels ofp179a9(Fig. 6C). Deletionof RRE frompNL17Rdid notaltertheexpression in theabsence ofRev, demonstrating that the RRE did not negatively affectp179a9expression (datanot shown).
Theseresults verifiedthataninhibitoryelement is located inthecoding region ofp179a9and thatthis elementactsby decreasing the steady-state levels ofmRNA. The negative effect ongag expression exerted by INS-1 could be
over-come by the Rev-RRE interaction, demonstrating that the expression ofpl7gag isRev dependent.
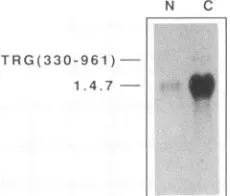
Inhibitory element INS-1 affects the RNA levels in the nucleus.Theexperimentsdescribed above indicated that the decreasein Tatprotein expressioncausedby INS-1couldbe accounted forby the low mRNA levels. We next investi-gated whetherthe differencesbetween total tat-gagand tat mRNA levels reflected differences in the nuclear pools of
transfected cells. Nuclear and cytoplasmic RNA fractions were prepared from HL3T1 cells cotransfected with
pNL1.4.7 and pTRG(330-961) and analyzed by Northern
blotting. tat-gag mRNAswere found in lower levels than tat mRNA, both in the nucleus and in the cytoplasm of
trans-fected cells (Fig.7). The low levels of mRNATRG(330-961) inthe nucleus could only be detected by longer exposures of the blots. These results indicated that the INS-1 element
decreasedthe levelsofRNA alreadyin the nucleus. The INS-1element decreases the half-lives of gag-containing RNAs. Tofurtherstudy the effect ofthe INS-1 element on RNAlevels, we measuredthestability ofgag-containingtat mRNAs. HL3T1 cells cotransfected with pNL1.4.7 and
pTRG(330-961) were treated with actinomycin D to block
transcription, and totalRNA wasanalyzed at various times.
This allowed simultaneous comparison of the stabilities of the two different tat mRNAs in the same cells under the sameexperimental conditions. gag-containingmRNAs were less stable than tat mRNA 1.4.7(Fig. 8A). We next
deter-minedthe half-lives of tat mRNA 1.4.7 and gag-containing mRNA TRG(330-961). To accurately quantitate the low mRNAlevels producedby gag-containingmRNA
TRG(330-961), we cotransfected 3 ,ugofpTRG(330-961) and1 ,ug of pNL1.4.7. Quantitation ofone of three independent
experi-mentsisshownin Fig. 8B. tat mRNA had ahalf-life of4 h,
and the half-life oftat-gag mRNA was approximately 1 h
(Fig. 8B). These results indicated that the INS-1 element
contained sequences that decreased mRNA stability. The
differences inRNAstabilitymeasured here may not account
forthewholeinhibitory effect exerted bytheINS-1element.
Since the stability measurements were performed on total RNA, of whichthe majorcomponent is cytoplasmic RNA,
the stability of the nuclear RNA pool was not measured directly. The stabilities oftat mRNA and tat-gag mRNAs maybedifferent inthenucleusand inthe cytoplasm. Further
experiments are necessary to determine whether the
de-creaseinmRNA
stability
istheonly mechanism of action of INS-1.Inspection of the INS-1 sequence revealed that it is AU
rich
(61.5%)
compared with the complete HIV-1 genome.Figure9showsaplot of theAU content withinINS-1.Two
regions ofvery high AU content are apparent. Deletion of
one of these AU-rich regions in mutant pTRG(481-631) abolished the inhibitory effect of INS-1, suggestingthatthe instability of mRNA caused by INS-1 may be associated withtheAU-rich subregions. Interestingly, eitherof the two AU subregions does not act alone, as demonstrated by
mutants pTRG(330-480) and pTRG(481-631). This result suggeststhat redundantsequenceelementsor,
alternatively,
a complex structure of thecomplete
218-nt sequence is required for inhibition.DISCUSSION
We have identified and characterizedan
inhibitory
RNAelement named INS-1 and located in the HIV-1
p17gag
coding sequence. Evidence also exists for the presence of
morethan oneINS element in thegag-protease gene
region
of the virus. Theinhibitory
effect ofINS-1 could be over-come, at leastpartially, by
the Rev-RREinteraction,
dem-onstratingthatthis sequence is
important
forRev-regulated
viral
expression.
Since the gag element decreased the
stability
of themRNAs, we askedwhether any
homology
existed between gagand cellularmRNAswithshorthalf-lives.Regulation
ofmRNA
stability
has beenknowntobe animportant
regula-N
lb bt
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.100.279.79.328.2]A
gagp4 repv
-p 17 p24 p7 vpr vpU
gag
RRE
{-IE
pNL17Rtat
- - pNL1.4.7
pl7gag -28S
-18S
FIG. 6. (A) Structureof the HIV-1 genomeand ofexpression plasmids pNL17RandpNL1.4.7. Open, shaded, orblack bars indicate ORFs. TheRRE isindicated. (B) Northern blotanalysisoftotal RNA from HLtat cells transfected withpNL17Rin thepresence(+)or
absence (-) ofpHCMVsrevorwithtatcDNApNL1.4.7. (C)Westernblotanalysisofextractsfrom HLtat cells transfected withpNL17R in thepresence (+)orabsence (-) ofpHCMVsrev.
tory step in gene expression (for reviews see references 8 and 11 and references therein). In the mRNAs studied to date,itappearsthat there existregions controlling instability rather thanstabilityand thatmorethanoneinstability region maybe present in thesamemRNA. Amongthe mRNAs for which instabilityisimportantinregulating expressionarethe
mRNAsencoding productsinvolved ingrowth control,such
as c-fos, c-myc, lymphokines, and cytokines. Histone and transferrin receptor mRNAs are also regulated by mRNA stability.
TRG(330-961 ) -1.4.7
-N C
FIG. 7. Northern blot analysis of nuclear (N) and cytoplasmic (C)RNAs prepared from HL3T1 cellscotransfected with both tat
cDNApNL1.4.7 and pTRG(330-961). Transfectionswereperformed
asdescribed in Materials and Methods. Nuclear and cytoplasmic RNAswereseparatedasdescribedby Greenberg and Ziff (24). The positionsofthe RNAsareindicatedonthe left.
Sequences specifyingRNAinstability have been mapped toAU-rich regions in the 3' untranslated regionsin cellular mRNAsencoding granulocyte-macrophage colony-stimulat-ingfactor(54),c-myc(28, 44), andc-fos (56, 61, 63).Recent reports have also described elements which are located in
the coding regions ofc-myc and c-fos mRNAs and which affect thestabilityof these mRNAs(55, 64). ThegagINS-1 sequence has an AU content of61.5%, a content which is high compared with that of tat mRNA and most cellular mRNAs, which have AU contents of about 50%. The pentanucleotide AUUUA, which has been suggested to be
an important determinant of RNA instability in cellular
mRNAs (54), wasnot present in INS-1, indicating that the
gag sequence contains a novel regulatory RNA element.
Further mutagenesis experiments and comparisons with
other elementsarerequiredtodefine theinstability determi-nantsinmoredetail. Inthiscontext,it isinteresting that nine copies of the sequence AUUUA exist within the gag-pol region ofHIV-1.Theseelementsmaybeassociated with the
additional instability regions suggested by our data.
Se-quence analysis has shown that all the studied lentiviruses
contain AU-rich regions (for a review, see reference 61a),
suggesting that these viruses may have similar regulatory
circuits.
Rev proteinactson RRE-containing mRNAs by promot-ingnuclearexportand mRNAutilization (4,16, 19, 26, 36). It has beenreported that splice sites are required for Rev
regulation and that they act by retaining the unspliced Pl 7!
[II}I--B
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.155.453.85.388.2] [image:7.612.115.230.576.674.2]A hours: 0 1 2 4
TRG(330-961) -1.4.7
FIG. 8. (A) Analysis of mRNA stabilityw microgram of pNL1.4.7 and3 ,ug of pTRG( fected into HL3T1 cells. The cells weretreal 20 hposttransfection. RNA was harvested at addition of actinomycin D (indicated at the positions of the two different mRNAs are in Quantitation of the Northernblots with the imaging system. Symbols: l,1.4.7 mRNAle mRNA levels. One representativeexperime:
mRNAs in the nucleus (9, 34). The pa sites in the Rev regulatory pathway
experiments performedwithmutated gl It was postulated thatthe Rev-RRE in
the unsplicedorpartially spliced mRN.
some complex. Additional
experiment:
expressing molecularconstructs also suj
of5'splice sitesupstreamof theenv AU
(34). Therequirement for splice sitesd
general, since gag-containing plasmid regulation byRev inthe absence of and
sites (19). Furthermore, tat mRNA cont
3'
splice
sites thatcanbeactivated bytlupstream5'splice site (9a). However,t
3'
splice
siteson tatmRNAdoesnotin(3, 18, 38, 50), indicating that not all N
inhibit theexpression of viralmRNAsir All retroviruses express Gag and Gz
unspliced
mRNAs that contain unuseretroviral splice sitesareinefficientbyd
and a mix of unspliced and spliced r
generated to produce the correct
pros
viral proteins. Thus, if unused splice retention or RNA
degradation,
we wretroviralgag-polmRNAswould be de evidence suggests that they are not. sarcoma virusGag and Pol proteins arn
[image:8.612.325.558.93.315.2]400 500 S6
FIG. 9. Graphic representation of the i INS-1 region. Two subregions with very h within INS-1. The deletions affecting thes phenotypesareshownatthe bottom.
6 8 B
Iog(cpm)
,ith actinomycin D. One
'330-961)
werecotrans-ted with actinomycin D various times after the topof the figure). The idicated on the left. (B) AMBIS radioanalytic
wvels;0,TRG(330-961) ntof three is shown.
trticipation of splice
Iwas suggested by
lobin splice sites(9).
teraction dissociates
Asfrom the
spliceo-s with mutated
env-ggested
thenecessityIG for Revregulation oes notappear to be s are dependent on
2.2
-2.0
-1.84
1.6
1.4
1.2
0 2 4 6 8 10
hours
yfunctional 5' splice malian cells in the absence of other viral
proteins (37,
62).
Intains three functional contrast,
study
ofthe expression of Gagand Envproteins
heintroductionofan of
complex retroviruses,
even in the absence of any func--hepresenceof these tionalsplice sites, strongly
suggests that thecorresponding
ihibitTatexpression mRNAsaredefective inexpressionunlessviral Rev(orRex) viral splice sites can protein ispresent.Thisevidencesuggests thattheseviruses
itheabsenceofRev. may contain
specific
inhibitory RNA sequences which areag-Pol proteins from notpresentintheless
complex
retroviruses.-d
splice
sites. The The INS-1element affects the abundance ofgag-contain-esign (29,30,59,60),
ing
mRNAs. This effect may be different from that of the nRNAs is thereforesplice
sites in HIV-1 env, which have been showntoinhibitportion
of structural thetransport of RNAs from the nucleustothecytoplasm(9,
sites led to nuclear
34).
An alternativeexplanation
for the low levels ofgag-ould expect that all
containing
RNAs maybe that these mRNAs areprocessed
,fective.Theexisting
differently
from most cellular mRNAs orby an alternative For example, Rouspathway
thatrequires
Rev.Theseproposals
arenotmutuallyeexpressed in mam-
exclusive,
and the results of different laboratories suggestthat the viral transcripts are defective for more than one reason. To explain the different observations, we have proposed that Rev acts as a chaperone to guide
RRE-containing
mRNAsthrough aspecific
transport andutiliza-tion pathway which is initiated immediately after
transcrip-tionandleadstoefficient translation(4). Thispathwaymust
provide partial protection from splicing and
degradation
50% AU within the nucleus. This hypothesis is consistent with all theexperimental results and
explains
thestabilization,
trans-port, andtranslational effects.
According
to thisview,
theRev-RRE interaction is a
positive
interaction needed to330x1
lowcorrect
apreexisting
defect on the viral mRNAs caused byseveral INS elements. Thesplice
sites inenv(9, 34)
and330-480 high the
INS-1
sequence in the gag gene may act at different- 355-631 low stages of posttranscriptional processing of the HIV-1
-
414-831
low mRNAs. Sequences similar in function toINS-1may also be- 481-631 high presentintheenv
region. Regions
in theenvgene have been AU content within the shown to down-regulate CAT expression when linked to the igh AU contents exist CAT gene (47) and to inhibit the expression of Tat from a e subregions and theirhybrid
tat-envcDNA(40). Similarly,wehave evidence thatinstability sequences also exist in the gag gene of other
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.68.317.549.694.2]complex retroviruses (58a). It has also been shown that
human T-cell lymphotropic virus types I andII contain
inhibitorysequencesin the U5regionof theLTR(7, 53).The
function of these sequences has not been characterized in
detail.
The presence ofmRNA-destabilizing sequences such as INS-1 in thegaggene mayreflect arequirement for low or
no expression of structural proteins at certain stages of the
viral life cycle. This requirement may be necessary for a
low-levelorlatentinfectiontooccur. Ithasbeenshown that
some monocytoid cell clones silently infected with HIV-1 produce only multiply spliced and singly spliced mRNAs, while full-length gag-pol mRNAs arenot detected inthese cells (43). These results further suggest a requirement for
down-regulation of the
gag-pol
mRNA levels for a silentinfection to occur, indicating that the expression of these
mRNAs isselectively inhibited.
Inconclusion,wepresentevidencearguingthatunspliced
HIV-1 mRNAs contain distinctsequencesotherthan splice
sites which decrease the stability of the viral mRNAs and inhibit the expression of these mRNAs in the absence
of Rev. Given these results, it is possible that multiple
elements, acting by more than one mechanism, affect the
transport, stability, and utilization of HIV-1 mRNAs. It is reasonable to hypothesize that these inhibitory sequences,
notpreviously found in other retroviruses, have an
impor-tantfunctioninthe life cycle ofHIV-1 andotherlentiviruses.
ACKNOWLEDGMENTS
S.S. isagraduatestudentattheKarolinskaInstitute,Stockholm, Sweden.WearegratefultoM. Powersforoligonucleotide synthe-sis, M. Campbell and J. Harrison for technical assistance, V. Ciminale and G. Nasioulas for discussion, N. Tavernarakis for participatinginsomeexperiments,and A. Arthurand J.Hopkinsfor editorialassistance.
This research was sponsored by the National Cancer Institute
undercontractN01-CO-74101with ABL.
REFERENCES
1. Ahmed, Y. F., S. M. Hanly, M. H. Malim, B. R. Cullen, and W. C. Greene. 1990. Structure-function analysis ofthe HTLV-I
rex and HIV-1 rev RNA response elements: insights into
the mechanism ofrex and rev function. Genes Dev.
4:1014-1022.
2. Arrigo, S. J., and I. S. Y. Chen. 1991. Rev is necessary for
translationbutnotcytoplasmic accumulationof HIV-1 vif,vpr,
andenv/vpu2 mRNAs. Genes Dev. 5:808-819.
3. Arya,S. K., andR. C. Gallo. 1986.Three novelgenesof human
T-lymphotropicvirustypeIII: immunereactivity of their
prod-ucts with sera from acquired imnmune deficiency syndrome
patients. Proc.Natl. Acad. Sci. USA 83:2209-2213.
4. Benko,D. M.,B. K. Felber, J. E. Harrison, and G. N. Pavlakis. Submittedfor publication.
5. Benko, D. M., R. Robinson, L. Solomin, M. Mellini, B. K.
Felber, and G. N. Pavlakis. 1990. Binding of trans-dominant
mutantrevprotein ofhuman immunodeficiency virustype1to
the cis-actingrevresponsive elementdoesnotaffect the fate of
viralmRNA. NewBiol.2:1111-1122.
6. Berger,J.,J. Hauber,R. Hauber, R. Geiger,and B. R. Cullen.
1988. Secretedplacental alkalinephosphatase: apowerful new
quantitative indicator of gene expression in eukaryotic cells.
Gene 66:1-10.
7. Black, A. C., I. S. Y. Chen, S. Arrigo, C. T. Ruland, T.
Aflogiamento,E. Chin,and J. D. Rosenblatt. 1991. Regulationof
HTLV-IIgeneexpressionbyrexinvolves positive andnegative
cis-acting elements in the 5' long terminal repeat. Virology
181:433-444.
8. Brawerman, G. 1987. Determinants of messenger RNA
stabil-ity. Cell 48:5-6.
9. Chang, D. D., andP. A. Sharp. 1989. Regulation by HIV Rev depends upon recognitionof splice
sites.
Cell59:789-795. 9a.Ciminale, V.,S. Schwartz, and G. N. Paviakis. Unpublishedresults.
10. Clavel, F., and J. M. Orenstein. 1990. A mutant of human immunodeficiency virus with reduced RNA packaging and ab-normal particle morphology. J. Virol. 64:5230-5234.
11. Cleveland, D. W., and T. J.Yen.1989. Multipledeterminants of eukaryotic mRNAstability. NewBiol. 1:121-126.
12. Cochrane, A., C. H. Chen, and C. A. Rosen. 1990. Specific interaction of the human immunodeficiency virus Rev protein with a structured region in the env mRNA. Proc.
Natl.
Acad. Sci. USA 87:1198-1202.13. Daefler, S., M. E. Klotman, and F. Wong-Staal. 1990. Trans-activating rev protein of the human immunodeficiency virus 1 interacts directly and specifically with its target RNA. Proc. Natl. Acad. Sci. USA 87:4571-4575.
14. Daly, T., K. Cook, G. Gray, T. Maione,and J. Rusche. 1989. Specific binding of HIV-1 recombinant Rev protein to the Rev-responsive element in vitro. Nature (London) 342:816-819.
15. Dayton, A. I., E. F. Terwilliger, J. Potz, M. Kowalski, J. G. Sodroski, and W. A. Haseltine. 1988. Cis-acting sequences responsive to the rev gene product of the human immunodefi-ciency virus. J. Acquired Immune Defic. Syndr. 1:441-452. 16. Emerman, M., R. Vazeux, and K. Peden. 1989. The rev gene
product of the human immunodeficiency virus affects envelope-specific RNA localization. Cell 57:1155-1165.
17. Feinberg, M. B., R. F. Jarrett, A. Aldovini, R. C.
Gallo,
and F. Wong-Staal. 1986.HTLV-IIIexpression and production involve complex regulation at the levels of splicing and translation of viral RNA. Cell 46:807-817.18. Felber, B. K., C. M. Drysdale, and G. N. Pavlakis. 1990. Feedback regulation of human immunodeficiency virus type 1 expression by the Rev protein. J. Virol. 64:3734-3741. 19. Felber, B. K., M. Hadzopoulou-Cladaras, C.
Cladaras,
T.Cope-land, and G. N. Pavlakis.1989. rev protein of human immuno-deficiency virus type 1 affects the stability and transport of the viral mRNA. Proc. Natl. Acad. Sci. USA86:1495-1499. 20. Felber, B. K., H. Paskalis, C. Kleinman-Ewing,F. Wong-Staal,
and G. N. Pavlakis. 1985. The pX protein of
HTLV-I
is a transcriptional activator of its long terminal repeats. Science 229:675-679.21. Garrett, E. D., L. S. Tiley, and B. R. Cullen. 1991. Rev activates expression of the human immunodeficiency virus type 1 vif and vpr gene products. J. Virol. 65:1653-1657.
22. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982. Recombinant genomes which express
chloramphenicol
acetyl-transferase in mammalian cells. Mol. Cell. Biol. 2:1044-1051. 23. Graham, F. J., and A. J. Van der Eb. 1973. A new technique forthe assay of infectivity of human adenovirus 5 DNA. Virology 52:456-460.
24. Greenberg, M. E., and E. B. Ziff. 1984. Stimulation of 3T3 cells induces transcription of the c-fos proto-oncogene. Nature (Lon-don) 311:433-437.
25. Hadzopoulou-Cladaras, M.,B. K. Felber, C. Cladaras, A. Atha-nassopoulos, A. Tse, and G. N.Pavlakis. 1989. The rev (trslart) protein of human immunodeficiency virus type 1 affects viral mRNA and protein expression via a cis-acting sequence in the env region. J. Virol. 63:1265-1274.
26. Hammarskjold, M. L., J. Heimer, B.
Hammarskjold,
I. Sang-wan, L. Albert, and D. Rekosh. 1989. Regulation of human immunodeficiency virus env expression by the rev gene product. J. Virol. 63:1959-1966.27. Heaphy, S., C. Dingwall, I. Ernberg, M. Gait, S. Green, J. Karn, A. Lowe, M. Singh, and M. Skinner. 1990. HIV-1 regulator of virion expression (Rev) protein binds to an RNA stem-loop structure located within the Rev response element region. Cell 60:685-693.
28. Jones, T., and M. Cole. 1987. Rapid cytoplasmic
turnover
of c-myc mRNA: requirement of the 3' untranslated sequences.on November 10, 2019 by guest
http://jvi.asm.org/
Mol. Cell. Biol. 7:4513-4521.
29. Katz, R. A., M. Kotler, and A. M. Skalka. 1988. cis-Acting intron mutations that affect the efficiency of avian retroviral RNA splicing: implication for mechanisms of control. J. Virol. 62:2686-2695.
30. Katz, R. A., and A. M. Skalka. 1990. Control of retroviral RNA splicing through maintenance of suboptimal processing signals. Mol. Cell. Biol. 10:696-704.
31. Knight, D. M., F. A. Flomerfelt, and J. Ghrayeb. 1987. Expres-sion of the art/trs protein of HIV and study of its role in viral envelope synthesis. Science236:837-840.
32. Krawczyk, Z., and C. Wu. 1987. Isolation of RNA for dot hybridization by heparin-DNase I treatment of whole cell ly-sate.Anal. Biochem. 165:20-27.
33. Lever, A., H. Gottlinger, W. Haseltine, and J. Sodroski. 1989. Identification ofa sequencerequired for efficient packaging of human immunodeficiency virus type 1 RNA into virions. J. Virol. 63:4085-4087.
34. Lu, X., J. Heimer,D.Rekosh, andM.-L.Hammarskjold. 1990. Ul small nuclear RNA plays a direct role in the formation ofa rev-regulated human immunodeficiency virus env mRNA that remains unspliced. Proc. Natl. Acad. Sci. USA 87:7598-7602.
35. Malim, M., L.Tiley, D.McCarn, J. Rusche, J. Hauber, and B. Cullen.1990. HIV-1structural geneexpression requiresbinding of the Rev trans-activator to its RNA target sequence. Cell 60:675-683.
36. Malim,M.H., J. Hauber, S. Le, J.V. Maizel,andB. R.Cuilen.
1989. The HIV-1 rev transactivatoracts through a structured target sequence to activate nuclear export ofunspliced viral mRNA. Nature(London)338:254-257.
37. Morris-Vasios,C., J.P.Kochan,and A. M. Skalka.1988.Avian sarcoma-leukosis virus pol-endo proteins expressed indepen-dently in mammalian cells accumulate in the nucleus butcanbe directedtoother cellular compartments. J. Virol. 62:349-353. 38. Muesing, M. A., D. H.Smith, C. D. Cabradilla, C. V. Benton,
L.A.Lasky,and D. J.Capon. 1985. Nucleic acidstructureand expression of the human AIDS/lymphadenopathy retrovirus. Nature (London) 313:450-458.
39. Myers,G. 1990. Human retroviruses and AIDS. Acompilation
and analysis of nucleic acid and amino acid sequences. Los AlamosNational Laboratory, LosAlamos, N.M.
40. Nasioulas, G., S. Schwartz, G. N. Paviakis, and B. K. Felber. Unpublishedresults.
41. Olsen, H. S., P. Nelbock, A. W. Cochrane, and C. A. Rosen. 1990.Secondary structureis themajordeterminant for interac-tion of HIVrevproteinwith RNA. Science 247:845-848. 42. Pavlakis, G.N., and B. K. Felber. 1990. Regulationof
expres-sionof human immunodeficiency virus. New Biol.2:20-31. 43. Pomerantz,R.J., D.Trono,M. B.Feinberg,and D. Baltimore.
1990. Cells nonproductively infected with HIV-1 exhibit an
aberrantpatternof viral RNA expression:amolecular model for latency. Cell 61:1271-1276.
44. Rabbitts,P.H.,A.Forster,M. A.Stinson,and T. H.J.Rabbitts. 1985. Truncation of exon 1 from the c-myc gene results in prolongedc-mycmRNA stability. EMBO J. 4:3727-3733. 45. Ratner,L.,A.Fisher,L.L.Jagodzinski,R.S.Liou,H.Mitsuya,
R. C. Gallo, and F. Wong-Staal. 1987. Complete nucleotide sequencesof functional clonesof the virus associated with the acquired immunodeficiency syndrome, HTLV-III/LAV. Hae-matol. Bluttransfus. 31:404-406.
46. Ratner,L.,A.Fisher,L. L.Jagodzinski,H.Mitsuya,R.S.Liou,
R. C. Gallo, and F. Wong-Staal. 1987. Complete nucleotide sequences of functional clones oftheAIDS virus. AIDS Res. Hum. Retroviruses3:57-69.
47. Rosen, C. A., E. Terwilliger, A. Dayton, J. G. Sodroski, and W. A.Haseltine.1988.
Intragenic
cis-acting
artgene-responsive
sequences of the human immunodeficiency virus. Proc. Natl. Acad. Sci. USA 85:2071-2075.48. Saiki,R.K.,D. H.Gelfand,S.Stoffel,S.J.Scharf,R.Higuchi, G. T. Horn, K. B. Mullis, and H. A. Ehrlich. 1988. Primer-directedenzymatic amplificationof DNA withathermostable DNApolymerase.Science 239:487-491.
49. Saiki, R. K.,S.Scharf, F. Faloona,K. B.Mullis, G. T.Horn, H. A.Erlich,and N.Arnheim. 1985.Enzymatic amplificationof
,B-globin genomic sequences and restriction site analysis for
diagnosisof sickle cell anemia. Science230:1350-1354. 50. Schwartz, S.,B.K.Felber,D.M.Benko,E. M.Fenyo,andG. N.
Paviakis. 1990. Cloning and functional analysis of multiply spliced mRNAspeciesof humanimmunodeficiencyvirus type 1. J.Virol. 64:2519-2529.
51. Schwartz, S., B. K. Felber,E. M. Fenyo, and G. N. Pavlakis. 1990. Envand Vpuproteinsof humanimmunodeficiencyvirus type 1areproducedfrommultiplebicistronic mRNAs. J. Virol. 64:5448-5456.
52. Schwartz, S.,B. K.Felber,and G. N.Pavlakis. 1991.Expression
of humanimmunodeficiencyvirustype-1 vif and vprmRNAsis Rev-dependentandregulated bysplicing. Virology183:677-686. 53. Seiki, M.,A.Hikikoshi,and M. Yoshida.1990.TheU5sequence isacis-acting repressiveelementforgenomic RNA
expression
of humanTcellleukemia virus type 1.Virology 176:81-86. 54. Shaw, G.,andR. Kamen.1986. A conserved AU sequence from
the 3' untranslatedregion ofGM-CSF mRNA mediates selec-tive mRNAdegradation. Genes Dev.46:659-667.
55. Shyu, A.-B., J. Belasco,andM.Greenberg. 1991. Twodistinct
destabilizing elements in thec-fos message trigger
deadenyla-tion as a first step in rapidmRNA decay. Genes Dev. 5:221-231.
56. Shyu, A.-B., M. Greenberg, and J. Belasco. 1989. The c-fos
transcript is targeted for rapid decay by two distinct mRNA
degradation pathways.Genes Dev. 3:60-72.
57. Sodroski, J., W.C.Goh, C.Rosen,A.Dayton,E.Terwilliger,
and W. A.Haseltine.1986. Asecondpost-transcriptional
trans-activator gene required for the HTLV-III
replication.
Nature(London)321:412-417.
58. Solomin, L.,B. K. Felber, andG. N. Pavlakis. 1990. Different sites of interaction forRev, Tev, and Rex
proteins
within therev-responsiveelementof human
immunodeficiency
virustype 1.J. Virol. 64:6010-6017.58a.Solomin, L., S. Schwartz, G. N. Pavlakis, and B. K. Felber.
Unpublishedresults.
59. Stoltzfus,C.M.,andS.J.Fogarty.1989.
Multiple
regions
in the Rous sarcoma virus src gene intron act in cis to affect the accumulation ofunsplicedRNA.J. Virol.63:1669-1676. 60. Stoltzfus, C. M., S. K. Lorenzen, and S. L. Berberich. 1987.Noncoding region between the env and src genes of Rous sarcoma virus influences
splicing efficiency
atthe src gene 3'splicesite. J. Virol. 61:177-184.
61. Treisman, R. 1985. Transient accumulation ofc-fos RNA
fol-lowing serum stimulation
requires
a conserved 5' elementof c-fos sequences. Cell 42:889-902.61a.Wagner,R.R.,and H.Fraenkel-Conrat(ed.).1991. The Retro-viridae, vol. 1.PlenumPublishing Corp., NewYork.
62. Wills,J.W.,R. C.Craven,andJ.A. Achacoso.1989.Creation and expression of
myristylated
forms of Rous sarcoma virusGag protein
inmammalian cells. J. Virol. 63:4331-4343.63. Wilson, T., and R. Treisman. 1988. Removal of
poly(A)
and consequentdegradation
ofc-fos mRNAfacilitatedby
3' AU-richsequences. Nature(London)
336:396-399.64. Wisdom, R.,and W. Lee. 1991. The
protein-coding
region
of c-myc mRNA containsasequencethatspecifies
rapid
mRNA turnoverand inductionby
protein
synthesis
inhibitors. Genes Dev.5:232-243.65. Wright, C. M., B. K. Felber,H. Paskalis,and G. N.Pavlakis. 1986.Expressionandcharacterization of the trans-activator of HTLV-III/LAV virus. Science 234:988-992.
66. Zapp,M.,and M.Green.1989.
Sequence-specific
RNAbinding
bytheHIV-1rev