Copyright © 2003, American Society for Microbiology. All Rights Reserved.
Positive Regulation of CXCR4 Expression and Signaling by
Interleukin-7 in CD4
⫹
Mature Thymocytes Correlates with
Their Capacity To Favor Human Immunodeficiency
X4 Virus Replication
Nathalie Schmitt, Laurent Cheˆne, David Boutolleau, Marie-The´re`se Nugeyre,
Eric Guillemard, Pierre Versmisse, Catherine Jacquemot,
Franc¸oise Barre´-Sinoussi, and Nicole Israe¨l*
Unite´ de Biologie des Re´trovirus, Institut Pasteur, 75724 Paris Cedex 15, France
Received 16 December 2002/Accepted 27 February 2003
The emergence of X4 human immunodeficiency virus type 1 (HIV-1) variants in infected individuals is associated with poor prognosis. One of the possible causes of this emergence might be the selection of X4 variants in some specific tissue compartment. We demonstrate that the thymic microenvironment favors the replication of X4 variants by positively modulating the expression and signaling of CXCR4 in mature CD4ⴙ
CD8ⴚ CD3ⴙ thymocytes. Here, we show that the interaction of thymic epithelial cells (TEC) with these
thymocytes in culture induces an upregulation of CXCR4 expression. The cytokine secreted by TEC, interleu-kin-7 (IL-7), increases cell surface expression of CXCR4 and efficiently overcomes the downregulation induced by SDF-1␣, also produced by TEC. IL-7 also potentiates CXCR4 signaling, leading to actin polymerization, a process necessary for virus entry. In contrast, in intermediate CD4ⴙ CD8ⴚ CD3ⴚ thymocytes, the other
subpopulation known to allow virus replication, TEC or IL-7 has little or no effect on CXCR4 expression and signaling. CCR5 is expressed at similarly low levels in the two thymocyte subpopulations, and neither its expression nor its signaling was modified by the cytokines tested. This positive regulation of CXCR4 by IL-7 in mature CD4ⴙthymocytes correlates with their high capacity to favor X4 virus replication compared with
intermediate thymocytes or peripheral blood mononuclear cells. Indeed, we observed an enrichment of X4 viruses after replication in thymocytes initially infected with a mixture of X4 (NL4-3) and R5 (NLAD8) HIV strains and after the emergence of X4 variants from an R5 primary isolate during culture in mature thymocytes.
The chemokine receptors CXCR4 and CCR5 have been identified as the main coreceptors for human immunodefi-ciency virus type 1 (HIV-1) (4). A predominance of R5 HIV-1 isolates is observed during the establishment of HIV-1 infec-tion as well as throughout the asymptomatic period. Subse-quently, in about 40% of AIDS patients, an evolution towards an enrichment of X4 viruses occurs (44), concomitant with a drastic CD4 T-cell depletion (13, 28, 43, 51, 54, 58). The mech-anisms involved in the emergence of X4 variants early in chil-dren (50) and at later stages of infection in adults are poorly understood. Possibly, replication of X4 variants is enhanced in specific tissue compartments because of a more efficient usage of the CXCR4 coreceptor.
The thymus is known to be a critical site for pathogenic HIV infection in humans (20, 41, 45) and in animal models of AIDS (2, 55, 63). Thymuses from pediatric patients undergoing an accelerated disease process (8, 26, 35, 38) or from adults with AIDS (14, 48) show severe thymocyte depletion associated with a profound disorganization of the thymic epithelial net-work. Thymic failure may also be responsible for the fact that, among patients undergoing antiretroviral therapies for
ad-vanced disease, the CD4 cell counts often remain below nor-mal levels despite long-term suppression of viral load (32, 62). Conversely, the levels of CD4 counts progressively increase in patients producing drug-resistant HIV strains which display low replication efficiencies in thymocytes (31, 56).
We and others have shown that thymocytes are cellular targets of HIV infection (42, 47, 61). Furthermore, we have shown that HIV-1 takes advantage of activation signals re-quired for thymocyte differentiation for its replication in thy-mocytes. We demonstrated that interaction of human thymo-cytes with thymic epithelial cells (TEC) is required for HIV replication (47). This interaction leads to the cosecretion of two cytokines crucial for virus replication, tumor necrosis fac-tor (TNF) and interleukin-7 (IL-7), that further synergize with IL-6, IL-1, and granulocyte-macrophage colony-stimulating factor (GM-CSF) (11, 12). However the interaction with TEC appears to be efficient only in the case of mature singly positive CD4⫹CD8⫺CD3⫹(SP CD4⫹) thymocytes (11). These results
correlate with previous studies showing that, in simian immu-nodeficiency virus-infected macaques, most of the infected thy-mocytes are located in the medulla (30, 63), where mature thymocytes reside. However, we showed that the intermediate CD4⫹CD8⫺CD3⫺thymocytes were able to replicate the virus
in the presence of TNF and IL-7 (11). Specifically, restriction of viral replication in these intermediate cells was due to the lack of TNF secretion. However, in vivo, a lack of TNF might
* Corresponding author. Mailing address: Unite´ de Biologie des Re´trovirus, Institut Pasteur, 25 rue du Dr Roux, 75724 Paris Cedex 15, France. Phone: 33 1 4568 8944. Fax: 33 1 4568 8957. E-mail: nisrael @pasteur.fr.
5784
on November 8, 2019 by guest
http://jvi.asm.org/
be compensated by the TNF secreted by activated macro-phages that infiltrate the thymus during infection (20, 41).
In this study, we investigated the possibility that the thymus may be a site of enrichment for X4 variants. We studied the influence of the thymic microenvironment on the level of ex-pression and signaling of the two main coreceptors CXCR4 and CCR5 in mature SP CD4⫹and intermediate thymocytes.
We focused on the signals induced during TEC-thymocyte interaction and particularly on the cytokines involved in the induction of virus replication. We then correlated the efficiency of regulation of these coreceptors to the ability of each of these two thymocyte populations to favor replication of X4 versus R5 variants compared to peripheral blood mononuclear cells (PBMC).
MATERIALS AND METHODS
Reagents. (i) Cytokines and chemokines.Human recombinant cytokines IL-1 and IL-6 were purchased from PeproTech, Inc. (Rocky Hill, N.J.); IL-7, TNF-␣, GM-CSF, stromal cell-derived factor 1␣(SDF-1␣), and macrophage inflamma-tory protein 1␣(MIP-1␣) were from R&D Systems (Minneapolis, Minn.).
(ii) Viruses.HIV-1 NL4-3 and NLAD8 were produced by transfecting COS-7 cells with molecular clone pNL4-3 (1) or pNLAD8 (60). NLAD8 and NL4-3 provirus differ only in theenvgene sequence conferring a CXCR4 tropism on NL4-3 and a CCR5 tropism on NLAD8. Transfections of COS-7 cells were performed with SuperFect (Qiagen S.A, Courtaboeuf, France). Cell superna-tants were harvested 2 and 3 days after transfection.
Primary isolates were from HIV-1-infected individuals of Vietnamese (34) (V.CT8), Italian (49) (J2758), or Central African Republic (37) (11111D and 9614C) cohorts. They were propagated on phytohemagglutinin (PHA)–IL-2-activated PBMC. The 50% tissue culture infective dose (TCID50) was deter-mined by using serial dilutions of virus and calculated by the method of Ka¨rber (27). The HIV-1 p24gagantigen concentration was determined in culture super-natants with a Coulter HIV-1 p24 antigen assay (Beckman Coulter France S.A, Villepinte, France).
Cell culture conditions. (i) Preparation and culture of thymic cell populations.
Thymus fragments were obtained during elective cardiac surgery on HIV-sero-negative children (ages, 6 days to 24 months).
(a) Preparation of TEC.TEC were obtained as previously described (47) and seeded in selective medium: McCoy 5A containing 20% fetal calf serum (FCS), 1 mML-glutamine, 10 mM HEPES, antibiotics (Gibco BRL Life Technologies,
Paisley, Scotland), 500 ng of hydrocortisone (Sigma-Aldrich Chemical Co., St. Louis, Mo.)/ml, and 5⫻10⫺9M cholera toxin (Interchim, Montluc¸on, France) prior to coculture.
(b) Preparation and culture of thymocyte subpopulations.SP CD4⫹ and immature CD4⫺/⫹CD8⫺CD3⫺(consisting of a pool of triple negative CD4⫺ CD8⫺CD3⫺[TN] and intermediate CD4⫹CD8⫺CD3⫺thymocytes) thymocyte populations were obtained by negative selection as previously described (11). The thymocytes were then cultured in McCoy 5A, 10% FCS, 1 mML-glutamine,
10 mM HEPES, and antibiotics. Where indicated, specific cytokines were added at the initiation of the cultures and then every 3 or 4 days.
(c) TEC-thymocyte coculture.Autologous cocultures of TEC and mature SP CD4⫹or immature CD4⫺/⫹CD8⫺CD3⫺subpopulations were performed 3 days after thymic excision. A ratio of 2⫻105thymocytes per 104TEC per 1 ml per well (24 wells/plate) was used in each experiment.
(ii) Preparation and culture of PBMC.PBMC were obtained from the blood of HIV-seronegative donors by Ficoll-Hypaque gradient separation (Amersham Biosciences, Orsay, France). PBMC were then cultured in RPMI 1640 (Gibco BRL), 10% FCS, 1 mML-glutamine, 10 mM HEPES, and antibiotics and
acti-vated for 3 days with 0.5g of PHA (Murex Biotech Ltd., Dartford, England)/ml and then for 2 days with 540 IU of IL-2 (Proleukin; Chiron, Suresnes, France)/ml prior to infection.
(iii) Culture of human glioma cell line U87.U87 cell lines stably expressing CD4 or coexpressing CD4 and the chemokine receptor CXCR4 or CCR5 (15, 16) were cultured in Dulbecco’s modified Eagle medium–glucose (4.5 g/liter)– Glutamax–sodium pyruvate (Gibco BRL) containing 10% FCS, 10 mM HEPES, and antibiotics.
Infection of PBMC or thymocytes.Freshly isolated mature SP CD4⫹or im-mature CD4⫺/⫹CD8⫺CD3⫺thymocytes (107) or 5⫻106activated PBMC were infected with HIV-1 molecular clones or primary isolates for 1 h at 37°C. Cells
were then washed three times and cultured in RPMI 1640 containing 10% FCS, 1 mML-glutamine, 10 mM HEPES, and antibiotics, with 10 ng of TNF-␣/ml and
10 ng of IL-7/ml for thymocytes or 540 IU of IL-2/ml for PBMC. Culture supernatants were collected at different times postinfection and assayed for the presence of HIV-1 p24 antigen.
Characterization of viral coreceptor usage.To analyze viral coreceptor usage, U87 cells expressing CD4 CXCR4 or CD4 CCR5 were seeded into 96-well plates at a concentration of 104cells/well in Dulbecco’s modified Eagle medium– glucose (4.5 g/liter)–Glutamax–sodium pyruvate containing 10% FCS, 10 mM HEPES, and antibiotics. Twenty-four hours later, the cells were infected with serial dilutions of virus for 2 h at 37°C, washed four times, and cultured in fresh medium. HIV p24 antigen was determined at days 7 and 10 postinfection. CD4 U87 cells were included in each experiment as a control. The TCID50of each supernatant tested for coreceptor usage on CD4-CXCR4 and CD4-CCR5 U87 cells was calculated by the method of Ka¨rber (27).
Immunostaining of CXCR4 and CCR5.After incubation with mouse immu-noglobulin G1 control (679.1Mc7; Beckman Coulter), cells were immunostained by monoclonal antibodies 12G5-phycoerythrin (PE) for CXCR4 (BD-Pharmin-gen, San Diego, Calif.), 2D7-PE for CCR5 (BD-Pharmingen), or immunoglob-ulin G1-PE (679.1Mc7; Beckman Coulter). Immunostaining was analyzed with an XL-4C cytofluorometer (Beckman Coulter).
Determination of CXCR4 mRNA levels using real-time PCR.Total RNA was isolated from 107thymocytes with an RNeasy DNase extraction minikit (Qiagen S.A.). RNAs were reverse transcribed in a 20-l reaction volume by using a TaqMan reverse transcription kit and random hexamers (Perkin-Elmer Corp., PE Applied Biosystems, Foster City, Calif.). Reverse transcription was per-formed on a GeneAmp PCR system 9700 (PE Applied Biosystems) for 30 min at 48°C. Quantitative PCR was performed in special optical 96-well microtiter plates (PE Applied Biosystems) on an ABI Prism 7700 sequence detector system (PE Applied Biosystems). CXCR4 cDNA amplifications were performed with previously described oligonucleotides (25) and the SYBR Green PCR master mix (PE Applied Biosystems). To normalize for differences in the amount of total RNA added to the reaction mixture, quantification of 18S RNA with primers and probes purchased from PE Applied Biosystems was performed. Reverse transcriptions were performed in duplicate, and each reverse transcrip-tion product was submitted to real-time PCR at two dilutranscrip-tions.
Actin polymerization.The function of CXCR4 was evaluated by monitoring actin polymerization following the addition of SDF-1␣as previously described (9, 23). Briefly, 107thymocytes per ml were incubated at 37°C in RPMI medium containing 10 mM HEPES and SDF-1␣(1g/ml). Every 15 s, 100l of cell suspension was added to 400l of 100 mM fluorescein isothiocyanate-labeled phalloidin (Sigma), 0.125 mg ofL-␣-lysophosphatidylcholine (Sigma)/ml, and 4.5% formaldehyde in phosphate-buffered saline. Fixed cells were analyzed with an XL-4C cytofluorometer (Beckman Coulter). Similar experiments were per-formed to evaluate the actin polymerization induced through CCR5. For this purpose, 100 ng of MIP-1␣/ml was used as a ligand.
Detection of SDF-1␣levels.SDF-1␣levels were determined by a commercial enzyme-linked immunosorbent assay (Quantikine human SDF-1␣immunoassay; R&D Systems) according to the manufacturer’s instructions.
Statistical analysis.Because of the variability in the studied parameters dis-played by the different thymuses from different donors, we performed statistical analyses using the nonparametric Mann-Whitney U test. The level of significance was set at aPvalue of⬍0.05.
RESULTS
CXCR4 is highly expressed on thymocytes in contrast to CCR5, and CXCR4 levels are higher on immature (CD4ⴚ/ⴙ
CD8ⴚ CD3ⴚ) than on mature (CD4ⴙCD8ⴚ CD3ⴚ)
thymo-cytes.We first analyzed the specific expression of CXCR4 and CCR5, the major HIV coreceptors, on the two subsets of thymocytes able to produce the virus in the thymic microenvi-ronment, namely, mature SP CD4⫹and intermediate (CD4⫹
CD8⫺CD3⫺) thymocytes. The levels of expression of CXCR4
and CCR5 were determined by flow cytometry at the surfaces of freshly isolated mature SP CD4⫹and immature (isolated for
technical convenience as a pool of intermediate and TN) thy-mocytes. In the experiment whose results are shown in Fig. 1, CXCR4 was expressed on 88% of immature thymocytes, with
on November 8, 2019 by guest
http://jvi.asm.org/
a mean fluorescence intensity (MFI) of 11.6, and on 56% of mature thymocytes, with an MFI of 1.5. The proportion of labeled cells obtained from 13 independent experiments per-formed on thymuses from individual donors ranged from 86 to 97% (median, 94%) with an MFI of 4.1 to 12.9 (median, 7.3) for immature CD4 thymocytes and from 17 to 70% (median, 44%) with an MFI of 0.36 to 4.2 (median, 1.3) for mature thymocytes. These data indicate that CXCR4 is highly ex-pressed on the two cell populations, but this level was signifi-cantly higher on immature than on mature thymocytes (P⬍
0.001). In contrast, expression of CCR5 was similarly low (P⫽
0.7) in both populations. The rate of CCR5-labeled cells ranged between 1 and 19% (median, 6.9%), with an MFI between 0.02 and 0.4 (median, 0.3). These levels of coreceptor expression on thymocytes were compared to that on CD4⫹
cells within activated PBMC, which are well known as efficient target cells for both X4 and R5 viruses. At the time when we usually start HIV infection and under the proper conditions for activation (see Materials and Methods), 91% of these periph-eral CD4⫹cells expressed CXCR4 and 3.3% expressed CCR5.
However, these levels of expression varied in culture towards a decrease of CXCR4-labeled (78%) and an increase of CCR5-labeled (51%) cells when IL-2 culture was prolonged (for in-stance, after 14 days), as already reported (6, 10).
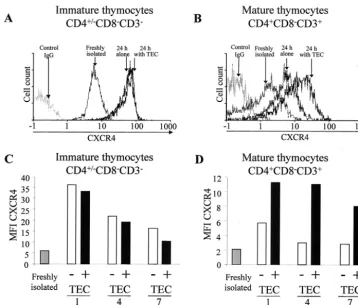
TEC upregulate CXCR4 expression in mature thymocytes.
We next examined whether TEC-thymocyte interaction, which favors HIV replication, may regulate coreceptor expression in thymocytes. Flow cytometry analysis of CXCR4 cell surface expression was performed on mature and immature thymo-cytes that had been either freshly isolated or cultured for 24 h alone or with TEC as previously described (47). A significant increase of CXCR4 expression was detected on the mature (P
⬍0.01) as well as on the immature (P⬍0.01) thymocytes after 24 h in culture, as shown by the shift of the peak of CXCR4-labeled cells (Fig. 2A and B). This increase, observed when thymocytes were removed from the thymic microenvironment, suggested that some factor(s) present in the thymic microen-vironment negatively regulates CXCR4. It should be pointed
out that the effect of the negative regulation is lower for the mature thymocytes (P⬍0.001). This may reflect the fact that interaction with TEC significantly increased CXCR4 expres-sion in mature (P⬍0.01) but not immature thymocytes (Fig. 2A and B), suggesting that some positive factor(s) secreted by TEC plays a role in CXCR4 expression in mature thymocytes but has no significant effect on immature thymocytes. This is indeed the case, since TEC increased CXCR4 expression in mature (Fig. 2B) but not in immature (Fig. 2A) thymocytes. The absence of an effect of TEC in immature thymocytes is not due to maximum levels of CXCR4 being already present, since TEC could not prevent the decrease of CXCR4 observed dur-ing prolonged culture (Fig. 2C). In contrast, TEC partially counteracted this decrease when it occurred on mature thymo-cytes (Fig. 2D).
SDF-1␣and IL-7 secreted by TEC have opposite effects on the modulation of CXCR4 expression on thymocytes.We next investigated the factors secreted by TEC that are involved in the regulation of CXCR4 expression in thymocytes. Since an increase of CXCR4 expression on thymocyte surfaces was de-tectable by 3 h in culture and culminated at 12 h (data not shown), we postulated that a putative negative regulatory fac-tor might act by masking or causing internalization of CXCR4, as has been described for SDF-1␣ (3, 36, 52). Indeed, we detected this chemokine in the supernatants of TEC cultures (mean of 190 pg/ml/105 cells after 4 days in culture). The
production of this chemokine by TEC is in agreement with a recent report (22) showing SDF-1␣staining in epithelial cells in sections of human thymus. As shown in Fig. 3A and B, in both mature and immature thymocytes, SDF-1␣inhibited in a dose-dependent manner the increase of CXCR4 expression induced during the culture. Even though TEC are able to secrete this negative regulatory factor, their interaction with mature thymocytes results in an overall positive regulatory effect. Therefore, we considered the possibility that additional factors present in the thymic microenvironment might act to increase CXCR4 expression. We particularly focused on the cytokines that we have previously shown to induce HIV-1 rep-lication in thymocytes, namely IL-7, IL-6, IL-1, TNF, and GM-CSF (11). As shown in Fig. 3B, among these cytokines, only IL-7 was able to enhance CXCR4 expression on mature and immature thymocytes (P ⬍ 0.01). This effect was weak but significant in immature thymocytes (P ⬍ 0.01). These data correlate with IL-7 receptor (IL-7R) expression levels, which are higher on mature than on immature thymocytes, as previ-ously shown (19). A dose-dependent effect of this cytokine was demonstrated on the mature subpopulation, with an optimal effect at 1.25 ng/ml (Fig. 3B). We then determined the effects on CXCR4 expression following cultivation of thymocytes in the presence of both SDF-1␣and IL-7. As shown in Fig. 3C, similar to the results at 24 h of cultivation mentioned above, a dose of 0.1g of SDF-1␣/ml led to about 50% inhibition of CXCR4 expression on the immature and mature populations at 12, 24, and 84 h. This decrease was overcome in mature (but not immature) thymocytes by IL-7 at a concentration of 1.25 ng/ml. In conclusion, SDF-1␣exerts a negative regulatory ef-fect on CXCR4 expression on the two subpopulations, whereas IL-7 overcomes this effect mainly in mature thymocytes.
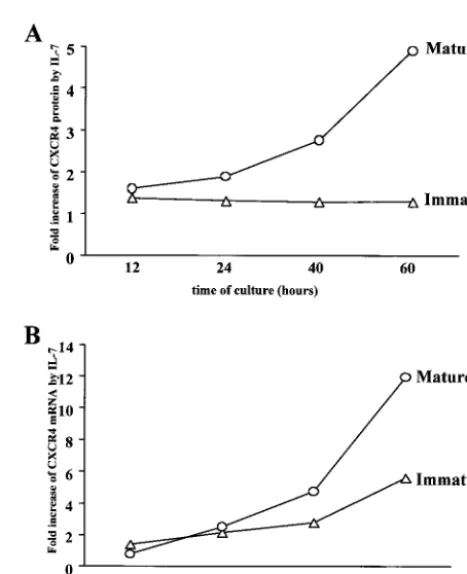
IL-7 positively regulates CXCR4 expression at the RNA level in mature thymocytes.To further investigate the possible
FIG. 1. Comparison of the levels of expression of CXCR4 and CCR5 in mature SP CD4⫹and immature CD4⫺/⫹CD8⫺CD3⫺
thy-mocytes. Immunostainings of CXCR4 and CCR5 were performed on freshly isolated thymocytes and analyzed by flow cytometry. White area indicates isotypic control. Percentages of labeled cells are given. The results are representative of independent experiments carried out with thymuses from 13 donors.
on November 8, 2019 by guest
http://jvi.asm.org/
regulatory role of IL-7 on CXCR4 expression at the transcrip-tional level, we determined the kinetics of CXCR4 mRNA expression in mature and immature thymocytes in the presence or absence of IL-7 (Fig. 4B). We analyzed in parallel the levels of cell surface CXCR4 protein (Fig. 4A). The values for mRNA or CXCR4 protein in the presence of IL-7 were ex-pressed as increase (-fold) compared with levels in untreated cells. As shown in Fig. 4B, in the mature thymocytes, a signif-icant increase in CXCR4 mRNA levels (P ⬍ 0.03) was ob-served at 40 and 60 h of culture. This was correlated with an increase at the CXCR4 protein levels (P⬍0.05). In immature thymocytes, as shown before, the increase of CXCR4 protein was lower (increase, 1.3-fold) and correlated here with a lower increase at the mRNA level. The difference in mRNA increase after 60 h in culture between mature and immature thymocytes was significant (P⬍0.03).
IL-7 increases CXCR4 signaling, leading to actin polymer-ization in mature thymocytes.CXCR4 can regulate HIV entry by internal signaling, in particular by inducing actin polymer-ization, because the entry of the virus needs disintegration and reconstitution of the membrane structure, which are associated with actin turnover (24). Therefore, we investigated whether IL-7, which increases CXCR4 expression, might also partici-pate to increase or prolong the induction of actin polymeriza-tion through CXCR4 binding. Since the level of IL-7R was
much higher on mature than on immature thymocytes (19), we hypothesized that IL-7 might sustain signaling through CXCR4 preferentially in mature thymocytes. SDF-1␣was used as a ligand to reveal the capacity of signaling of CXCR4. The kinetics of actin polymerization were determined in immature and mature thymocytes following culture for 40 h in the pres-ence or abspres-ence of IL-7. This period in culture is sufficient to free the IL-7R, which was masked or internalized following interaction with IL-7 within the microenvironment (data not shown). As shown in Fig. 5, actin polymerization, as deter-mined by immunostaining of filamentous actin (F actin), was rapidly induced following binding of SDF-1␣ to its receptor. This induction occurred at a higher level in mature than in immature thymocytes (P⬍0.03) in the absence of IL-7, and IL-7 increased and prolonged this induction only in mature thymocytes (P⬍0.01).
We also tried to investigate the kinetics of actin polymeriza-tion following the ligapolymeriza-tion of MIP-1␣(the most specific ligand of CCR5) with CCR5. No consistent signal was observed in any experimental condition (data not shown).
These data suggest that entry of X4 viruses might be favored over that of R5 viruses in thymocytes and particularly in ma-ture thymocytes following exposure to IL-7.
The selection of X4 versus R5 viruses was particularly fa-vored in mature thymocytes compared to that observed in
FIG. 2. Influence of TEC on CXCR4 expression in mature SP CD4⫹and immature CD4⫺/⫹CD8⫺CD3⫺thymocytes. Cell surface expression
of CXCR4 was analyzed on immature (A) and mature (B) thymocytes by flow cytometry at the time of isolation or after 24 h in culture alone or with TEC. Immunostaining of CXCR4 was determined in immature and mature thymocytes just after isolation or after 1, 4, or 7 days of culture with or without TEC. (C and D) Results are shown as MFI of CXCR4-labeled cells. The experiment presented here is representative of independent experiments carried out with thymuses from five donors.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:4.603.112.471.72.377.2]immature thymocytes or PBMC. Since IL-7 upregulates CXCR4 expression and signaling in mature thymocytes, these cells might have a higher propensity to favor replication of X4 viruses. Nevertheless, the higher CXCR4 constitutive level ex-pressed on immature thymocytes might compensate for the lower signaling capacity and might also permit high-level se-lection of X4 variants. We thus investigated the capacity of each subpopulation to favor X4 virus replication compared to that of PBMC.
The two populations of thymocytes or PBMC were each infected with the molecular clones pNL4-3 or pNLAD8. These two viruses differ from each other only in theenvgene, which confers CXCR4 or CCR5 tropism, respectively. The multiplic-ities of infection (MOI) used for NL4-3 and NLAD8 (respec-tively, 1.6 ⫻ 10⫺4 and 2.8 ⫻ 10⫺3) were chosen to obtain
consistent levels of replication, even with NLAD8, despite the low level of CCR5 coreceptors on thymocytes. The same MOI were also used to infect the PBMC. In order to evaluate the
minimal ratio necessary to observe any significant selection, we also performed infections with a mixture of NL4-3 and NLAD8 in known proportions (X4/R5 ratio, 1/103, 1/104, or 1/105).
Each distinct infected cell population was then cultured in the optimal conditions required for virus replication. Therefore, the infected thymocytes were cultured in the presence of IL-7 and TNF and the PBMC were cultured with IL-2 (as described in Materials and Methods).
We also verified that IL-7 has no direct effect on the level of HIV replication or on selection of X4 variants on activated T lymphocytes. Indeed we treated IL-2–PHA-activated PBMC from two healthy donors with IL-7 prior to infection with NL4-3 or NLAD8. IL-7 did not modify the level of replication of NL4-3 or of NLAD8 (data not shown).
Since the duration of the culture was 25 days, fresh thymo-cytes or PBMC were added to the culture at day 12, in order to provide new targets for the virus. Kinetics of virus replication was established for each cell type and for each viral inoculum by determination of p24 concentration in the culture
superna-FIG. 3. Effects of SDF-1␣and IL-7 on CXCR4 expression in ma-ture SP CD4⫹and immature CD4⫺/⫹CD8⫺CD3⫺thymocytes. Cell
surface expression CXCR4 was analyzed by flow cytometry on imma-ture or maimma-ture thymocytes, freshly isolated and after 24 h in culimma-ture in the presence of various concentrations of SDF-1␣(A), after 24 h in culture in the presence of various cytokines (10 ng/ml each or the indicated concentrations of IL-7) (B), or freshly isolated and after 12, 36, and 84 h in the presence of SDF-1␣(0.1g/ml) or a combination of SDF-1␣ (0.1g/ml) and IL-7 (1.25 ng/ml) (C). The results are shown as MFI of CXCR4-labeled cells and are expressed as means plus standard deviations of triplicate values for each thymus. The results are representative of independent experiments performed with thymuses from three donors.
FIG. 4. Upregulation of CXCR4 protein and mRNA levels in ma-ture SP CD4⫹ and immature CD4⫺/⫹ CD8⫺ CD3⫺ thymocytes by
IL-7. CXCR4 expression was monitored at the protein (A) and mRNA (B) levels in mature and immature thymocytes after 12, 24, 40, and 60 h in culture in the presence or absence of IL-7 (10 ng/ml). (A) CXCR4 protein expression level was determined by CXCR4 cell surface im-munostaining and analysis of labeled cells by flow cytometry. The results are shown as increase in MFI induced by IL-7 compared to untreated cells. (B) CXCR4 mRNA levels were determined by using a real-time kinetic quantitative reverse transcription-PCR. To normalize for differences in the amounts of total RNA, 18S rRNA was amplified as a control. The results are shown as increase of the normalized amount of CXCR4 mRNA induced by IL-7 compared to the amount in untreated cells. The data are representative of independent exper-iments carried out with thymuses from four donors.
on November 8, 2019 by guest
http://jvi.asm.org/
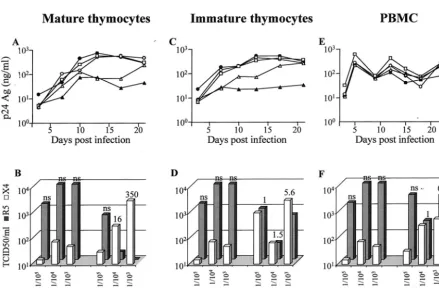
[image:5.603.303.537.300.588.2]tant (Fig. 6A, C, and E). To characterize the evolution of viral coreceptor usage during cell culture, we determined the ability of viral supernatants, before infection (initial inoculum) and at the end of the culture, to infect U87 cells coexpressing CD4 and CXCR4 or CD4 and CCR5. Viral tropism evolution was evaluated by calculating the ratios of viral titer (X4/R5) ob-tained in the two cell lines.
Despite the higher MOI, NLAD8 replicated less efficiently than NL4-3 in both mature and immature thymocytes (Fig. 6A and C), whereas the replication levels of the two viruses were similar in PBMC (Fig. 6E).
Infection of mature thymocytes with the mixtures (X4/R5, 1/103, 1/104, or 1/105) rapidly led to replication levels higher
than those observed with NLAD8 used alone, in both cell populations. The replication level attained with the X4/R5 mixtures at ratios of 1/103and 1/104was of the same order of
magnitude as that observed with NL4-3 alone, suggesting an enrichment of X4 versus R5 virus. Moreover, TCID50
deter-mined in CD4 CXCR4 and CCR5 U87 cells clearly demon-strated that mature thymocytes initially inoculated with
mix-FIG. 5. Influence of IL-7 on CXCR4 signaling leading to actin polymerization in mature SP CD4⫹ and immature CD4⫺/⫹ CD8⫺
CD3⫺thymocytes. Thymocytes were cultured for 40 h in the presence
[image:6.603.74.513.316.604.2](solid lines) or absence (dashed lines) of 10 ng of IL-7/ml. Immuno-staining of intracellular F actin was performed with fluorescein iso-thiocyanate-labeled phalloidin and was analyzed by flow cytometry after the addition of 1g of SDF-1␣/ml at time zero, in immature (triangles) and mature (circles) thymocytes. Results are shown as per-cent intracellular F-actin (MFI) relative to the value before addition of SDF-1␣. The data are representative of independent experiments car-ried out with thymuses from five donors.
FIG. 6. Comparison of mature SP CD4⫹thymocytes, immature CD4⫺/⫹CD8⫺CD3⫺thymocytes, and PBMC in their capacity to select X4
(NL4-3) versus R5 (NLAD8) viruses. (A, C, and E) Kinetics of HIV replication in mature (A) and immature (C) thymocytes and in PBMC (E). Cells were infected at MOIs of 1.6⫻10⫺4for NL4-3 (filled circle), 2.8⫻10⫺3for NLAD8 (filled triangle), or 2.8⫻10⫺3for a mixture of both at TCID50ratios (NL4-3/NLAD8) of 1/105(open triangles), 1/104(open squares), and 1/103(open circles). Infected thymocytes were cultured with IL-7 (10 ng/ml) and TNF-␣ (10 ng/ml). PHA–IL-2-activated PBMC were infected and maintained with IL-2 (540 IU/ml). Freshly isolated thymocytes or activated PBMC were added to the cultures of infected thymocytes or PBMC every 12 days in order to provide new targets for the virus. HIV replication was determined by measuring the p24gagconcentration in the culture supernatants every 3 or 4 days. (B, D, and F) Tropism
of viruses recovered from the supernatants of infected mature (B) and immature (D) thymocytes and in PBMC (F). Supernatants of thymocytes or PBMC infected with virus mixtures at ratios of 1/105, 1/104, and 1/103were harvested at day 21 for thymocytes and day 24 for PBMC. These supernatants and the initial mixtures used for infection (initial inoculum) were serially diluted and used to infect CD4-CXCR4 and CD4-CCR5 U87 cells. Replication of X4 and R5 viruses was determined by measuring the p24gagconcentrations in culture supernatants at day 7. X4 and R5
TCID50were then calculated by the method of Ka¨rber (27). The ratios of TCID50per milliliter for X4 versus R5 are given above the bars. ns, no significant ratio (⬍0.02) due to a too-low value for CD4-CXCR4 U87 cells.
on November 8, 2019 by guest
http://jvi.asm.org/
tures containing 103 and 104times more R5 than X4 viruses
produced 16 and 350 times more X4 than R5 after 21 days of culture (Fig. 6B). In the immature thymocytes, an enrichment of X4 viruses was also observed at day 21 postinfection but to a lower extent than in mature cells (Fig. 6D and B). In PBMC, the X4/R5 TCID50ratio balanced towards a similar level of
replication of X4 and R5 variants (Fig. 6F). Thus, it appears that, compared to PBMC, mature and to a lesser extent im-mature thymocytes are more efficient in selecting replication of X4 versus R5 virus.
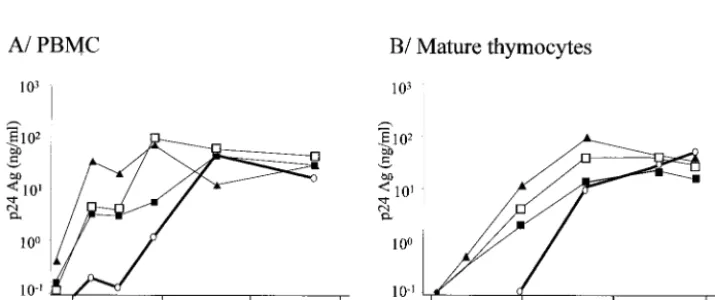
Mature thymocytes favor the emergence of X4 variants from an R5 primary isolate.To determine whether emergence of X4 variants could occur after replication of primary isolates in mature thymocytes, four R5 clinical isolates with no detectable X4 variants, according to their inability to grow in CD4-CXCR4 U87 cells, were inoculated onto mature thymocytes at the highest available titer (Table 1). The origin of each viral isolate and its coreceptor usage evolution are detailed in Ma-terials and Methods and in Table 1. All four R5 isolates effi-ciently replicated in PBMC and in mature thymocytes (Fig. 7). Viruses obtained at day 32 postinfection were tested for their coreceptor usage.
For one out of the four R5 isolates tested (V.CT8), X4
variants could be detected only in the supernatants of mature thymocytes (Table 1).
DISCUSSION
In the present report we studied the influence of the thymic microenvironment on the control of expression and signaling
of CXCR4 and CCR5 in mature SP CD4⫹ and immature
intermediate thymocytes, which were shown to allow virus rep-lication in the presence of the cytokines of the thymic micro-environment. We also studied the consequences of this control on the capacity of these cells to replicate R5 or X4 HIV-1. Just after isolation, CCR5 is expressed at similarly low levels in mature SP CD4⫹ and immature thymocytes, while CXCR4
[image:7.603.44.541.82.170.2]expression is much higher in both subsets, particularly in im-mature cells. The latter result is consistent with previous stud-ies indicating that CXCR4 expression decreases with thymo-cyte maturation in cells freshly isolated from fetal thymuses (5) or from thymuses obtained after cardiac surgery in children (64). Careful examination of coreceptor expression in mature and immature thymocytes, cultured as isolated populations or in the presence of TEC, demonstrate that CXCR4 but not CCR5 (data not shown) expression is regulated by the thymic
[image:7.603.116.474.517.667.2]FIG. 7. Kinetics of replication of the primary isolates described in Table 1 in PBMC or mature thymocytes. PBMC (A) and mature thymocytes (B) were infected with various primary HIV-1 isolates described in Table 1. Freshly isolated thymocytes or PHA–IL-2-activated PBMC were infected with the primary isolates at MOI of 1.6⫻10⫺3for V.CT8 (open circles), 9.5⫻10⫺4for 11111D (filled triangles), 3.5⫻10⫺4for J2758 (open squares), and 2.25⫻10⫺3for 9614C (filled squares). Infected thymocytes were cultured with IL-7 (10 ng/ml) and TNF-␣(10 ng/ml). Infected PBMC were maintained with IL-2 (540 IU/ml). Freshly isolated thymocytes or activated PBMC were added to the cultures every 10 days in order to provide new targets for the virus. HIV replication was determined by measuring the p24gagconcentration in the culture supernatants.
TABLE 1. Comparison of tropism evolution of primary isolates in PBMC or mature thymocytesa
Isolate Origin
Initial tropism Final tropism
R5 X4 PBMC Thymocytes
R5 X4 R5 X4
V.CT8 Vietnam 1.8⫻102 ⬍101 3.2⫻102 ⬍101 1⫻103 2⫻102
11111D Central African Republic 3.2⫻103 ⬍101 3.2⫻103 ⬍101 3.2⫻104 ⬍101
J2758 Italian cohort 3.2⫻102 ⬍101 5.6⫻103 ⬍101 5.6⫻102 ⬍101
9614C Central African Republic 5.6⫻104 ⬍101 1⫻103 ⬍101 5.6⫻102 ⬍101
aThe tropisms of viruses recovered from the supernatants of infected mature thymocytes or PBMC were determined at day 32 postinfection. Initial isolates and supernatants from day 32 were serially diluted and used to infect CD4-CXCR4- and CD4-CCR5-expressing U87 cells.⫻4 and R5 virus replication was determined by measuring the p24gagconcentration in the culture supernatants at day 7. The X4 and R5 values are TCID50/ml calculated by the method of Ka¨rber (27). The threshold of detection on U87 cells is 101TCID50/ml.
on November 8, 2019 by guest
http://jvi.asm.org/
microenvironment. Indeed, the microenvironment taken as a whole exerts a negative regulatory effect on CXCR4 expression in the two subpopulations, whereas TEC alone increase CXCR4 expression essentially in the SP CD4⫹ thymocytes.
The negative effect was very likely induced by SDF-1␣, which we and others have found to be secreted by TEC (data not shown and reference 22). The negative effect of SDF-1␣ on CXCR4 expression likely results from the masking or the in-ternalization of CXCR4 (3). Nevertheless, coculture between TEC and thymocytes resulted in a net increase of CXCR4 expression in mature cells, suggesting that factors involved in TEC-thymocyte interaction can overcome the negative effect of SDF-1␣ on CXCR4 expression. This increase was due to soluble factors secreted by TEC, since supernatant of TEC cultures had the same effect (data not shown). The factor(s) involved in the positive regulation of CXCR4 was searched for among those involved in HIV replication, such as TNF, IL-7, IL-6, IL-1, and GM-CSF (11). Among these cytokines, we identified one, IL-7, capable of enhancing CXCR4 expression and of overcoming the down-regulation of CXCR4 induced by SDF-1␣. This enhancement correlates with the expression level of the IL-7R, which is much higher in mature than in immature thymocytes, as previously shown (19). Thus, the pos-itive effect of TEC mainly on mature thymocytes underlines the possibility that IL-7 might be the main positive factor of the microenvironment. However, since IL-7-neutralizing antibod-ies are not available, we were unable to exclude the possibility that other factors may play a synergistic role. Furthermore, independently of their microenvironment, the thymocytes themselves might maintain a high level of CXCR4 through the secretion of IL-2 and IL-4, which have been shown to increase CXCR4 expression in mature thymocytes (42, 61; also data not shown).
We then attempted to elucidate the mechanisms by which IL-7 modulates expression and function of CXCR4. First, we demonstrated that the effect of IL-7 on CXCR4 expression takes place at the mRNA level and that this positive regulatory effect was more efficient in mature than in immature thymo-cytes in accordance with the IL-7R content. Second, we deter-mined that SDF-1␣, used as a ligand, induced CXCR4 signal-ing, leading to actin polymerization (24), more efficiently in mature than in immature thymocytes. IL-7 increased and pro-longed this induction specifically in mature thymocytes. Since induction of actin polymerization was obtained especially in mature thymocytes even in the absence of the addition of exogenous IL-7, we suggest that IL-7, within the microenviron-ment, might increase CXCR4 function by inducing signaling molecules which might remain partially activated at the time chosen here for the actin polymerization assay (40 h after thymocyte isolation). Such an activated molecule might be protein kinase B, which was shown to be activated by IL-7 (59) and to prolong CXCR4 signaling (40). Addition of IL-7 to the culture medium might potentiate this mechanism.
The level of expression of CCR5 was not modified by any culture conditions (data not shown). However, the thymocytes secreted MIP-1␣, MIP-1, and RANTES (data not shown), and it is thus possible that these CC chemokines might main-tain CCR5 expression at a low level. This possible autocrine regulation of CCR5 by the CC chemokines might explain why the addition of cytokines was unable to modify the level of
CCR5. The binding of MIP-1␣to CCR5 did not induce actin polymerization in the thymocytes, irrespective of the culture conditions. These data are reminiscent of the fact that MIP-1
is unable to induce significant chemotaxis of thymocytes (57). In addition to the difference in the level of expression of CXCR4 and CCR5, the main difference in their activity sug-gested a possible selection in favor of X4 viruses, particularly in the mature thymocytes.
To evaluate the minimal proportion of X4 virus necessary for a possible selection, we first used mixtures of NL4-3 and NLAD8 in various proportions. The replication kinetics assay showed that an X4/R5 ratio of 1/104was sufficient to
prefer-entially produce X4 viruses after infection of thymocytes. This selective process was more efficient in mature than in imma-ture thymocytes. The capacity of maimma-ture and immaimma-ture thymo-cytes to enrich for X4 variants was higher than that of PBMC, since in these cells a lower proportion of X4 viruses was pro-duced after 24 days. This was expected, since PBMC display a higher level of CCR5 than thymocytes (data not shown) and prolonged activation of CD4⫹T cells from PBMC leads to a
decrease in CXCR4 expression (6) and an increase in CCR5 expression (10). This difference could also be explained by the fact that CD4⫹cells from PBMC are highly sensitive to
X4-induced cytopathogenic effects (29) while X4 replication in thymocytes and especially in mature thymocytes is not associ-ated with a high level of apoptosis of infected cells (19).
This selection of X4 variants in mature thymocytes was also observed when these cells were infected by one primary isolate out of four tested. In contrast, PBMC were unable to select X4 variants from this primary isolate. The fact that the other three primary isolates did not give rise to any X4 variants might be due to a real lack of X4 variants in the initial isolates or to a very high dilution of X4 viruses (X4/R5 ratio⬍1/104). Indeed,
we think that the selection of X4 viruses is mainly due to an enrichment of preexisting X4 variants rather than to muta-tional events in theenv gene, since X4 variants could not be obtained from an R5 molecular clone, at least in vitro, over the culture period (even after 74 days of culture) (data not shown). However, the evolution by mutations, which is dependent upon a high number of replication cycles, is not excluded in vivo in the mature thymocytes because of their permanent activation state within their microenvironment (12) (in contrast to T lymphocytes, which are mostly resting).
Recent data in the literature indicate the potential use of the coreceptor CCR8 by HIV-1. However, none of the molec-ular clones or isolates used was able to productively infect CD4-CCR8 U87 cells (data not shown). This negative result strengthens the importance of the two main coreceptors, CXCR4 and CCR5, in the thymus.
In this paper, we emphasize the importance of the thymic microenvironment and particularly of IL-7 in the positive con-trol of expression and signaling of CXCR4 and the fact that such a control favors the replication of X4 viruses in the thy-mus, mainly in mature thymocytes. This more effective repli-cation of X4 variants might be particularly efficient in children, because they display a high thymic function.
This interpretation is supported by a series of published data. Children whose thymuses are infected progress rapidly towards AIDS (8, 26, 35, 38). This might be sometimes related to the fact that in infants infected by their seropositive mothers
on November 8, 2019 by guest
http://jvi.asm.org/
who progressed rapidly to AIDS (three of nine), X4 isolates appeared very early in the course of the disease (mean, 7.7 months after birth) (50). It is indeed a general observation that emergence of X4 viruses is often correlated with accelerated progress of the disease in children (13, 54) and adults (28, 43, 51, 58). However, the particularly fast clinical evolution in children with X4 variants has been explained (13) by increased T-cell depletion due to cell destruction (as in adults) but ag-gravated by an impairment of production of new T cells (again suggesting an insult to the thymus).
In middle-aged adults, the thymus, albeit less active, is still functional (17). Furthermore, 50% of adult HIV patients ex-hibit a rebound of thymic activity (56) capable of generating new naive T cells (53). Therefore, emergence of X4 variants in adult thymus is plausible, especially since high levels of IL-7 in plasma observed in HIV patients (39), which might reflect this rebound, have been associated with the emergence of X4 vari-ants (33). Furthermore, it has been shown that X4 varivari-ants develop more in circulating naive T cells (7). As these cells are normally quiescent in the blood and may not support active replication, a proliferation step must take place. This happens to compensate for the destruction of CD4⫹T cells (21) by the
virus, and the number of infected naive T cells is therefore a function of the decline of the CD4⫹T cells. In this hypothesis
it is not clear when naive T cells become infected, although in the case of thymus infection, these naive cells may derive from infected thymocytes. In any case, X4 variants in the blood might require an amplification period before being detectable. The length of time necessary for this amplification might there-fore be related to the number of infected recent thymic emi-grants (which is higher in children) and the rate of CD4⫹T cell
loss (again higher in children). In conclusion, we think that infection of the thymus is a factor of acceleration of the ap-pearance of X4 variants and thus of disease progression. This process might occur early in the course of the disease, for instance, in children whose thymuses were infected in utero, or later, since the thymus may also be infected at later stages (as shown in simian immunodeficiency virus-infected macaques [46]).
The participation of the thymus is probably not the sole possibility to explain the emergence of X4 variants, especially in advanced disease. At this stage, the consequences of gener-alized immune activation (18) might influence the amplifica-tion of X4 viruses in the blood compartment.
ACKNOWLEDGMENTS
N. Schmitt and L. Cheˆne contributed equally to this work. We thank Sonia Berrih-Aknin (Hoˆpital Marie Lannelongue, Le Plessis-Robinson, France) and F. Leco (Hoˆpital Necker, Paris, France) for providing us with thymuses and Gabriella Scarlatti (San Raffaele Scientific Institute, Milan, Italy) and Elisabeth Menu (Institut Pasteur, Paris, France) for providing us with primary HIV-1 isolates. We thank R. H. Bassin and M. Derrien for careful reading of the manuscript and G. Pancino and M. C. Mu¨ller-Trutwin for helpful discussions.
This work was supported by the Agence Nationale pour la Recher-che sur le SIDA (ANRS). L. Cheˆne was the recipient successively of a fellowship from Ensemble contre le SIDA (Fondation pour la Recher-che Me´dicale, Paris, France) and of a fellowship from Pasteur-Weiz-mann, N. Schmitt was the recipient of a fellowship from the French Ministry of Education and Research (MENESR), and E. Guillemard was the recipient of a fellowship from the Agence Nationale pour la Recherche sur le SIDA (France).
REFERENCES
1. Adachi, A., H. E. Gendelman, S. Koenig, T. Folks, R. Willey, A. Rabson, and M. A. Martin.1986. Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhuman cells transfected with an infectious molecular clone. J. Virol.59:284–291.
2. Aldrovandi, G. M., G. Feuer, L. Gao, B. Jamieson, M. Kristeva, I. S. Chen, and J. A. Zack.1993. The SCID-hu mouse as a model for HIV-1 infection. Nature363:732–736.
3. Amara, A., S. L. Gall, O. Schwartz, J. Salamero, M. Montes, P. Loetscher, M. Baggiolini, J. L. Virelizier, and F. Arenzana-Seisdedos.1997. HIV core-ceptor downregulation as antiviral principle: SDF-1␣-dependent internaliza-tion of the chemokine receptor CXCR4 contributes to inhibiinternaliza-tion of HIV replication. J. Exp. Med.186:139–146.
4. Berger, E. A., P. M. Murphy, and J. M. Farber.1999. Chemokine receptors as HIV-1 coreceptors: roles in viral entry, tropism, and disease. Annu. Rev. Immunol.17:657–700.
5. Berkowitz, R. D., K. P. Beckerman, T. J. Schall, and J. M. McCune.1998. CXCR4 and CCR5 expression delineates targets for HIV-1 disruption of T cell differentiation. J. Immunol.161:3702–3710.
6. Bermejo, M., J. Martin-Serrano, E. Oberlin, M. A. Pedraza, A. Serrano, B. Santiago, A. Caruz, P. Loetscher, M. Baggiolini, F. Arenzana-Seisdedos, and J. Alcami.1998. Activation of blood T lymphocytes down-regulates CXCR4 expression and interferes with propagation of X4 HIV strains. Eur. J. Im-munol.28:3192–3204.
7. Blaak, H., A. B. van’t Wout, M. Brouwer, B. Hooibrink, E. Hovenkamp, and H. Schuitemaker.2000. In vivo HIV-1 infection of CD45RA⫹CD4⫹T cells is established primarily by syncytium-inducing variants and correlates with the rate of CD4⫹T cell decline. Proc. Natl. Acad. Sci. USA97:1269–1274. 8. Blanche, S., M. Tardieu, A. M. Duliege, C. Rouzioux, F. Le Deist, K. Fuku-naga, M. Caniglia, C. Jacomet, A. Messiah, and C. Griscelli.1990. Longi-tudinal study of 94 symptomatic infants with perinatally acquired human immunodeficiency virus infection. Evidence for a bimodal expression of clinical and biological symptoms. Am. J. Dis. Child.144:1210–1215. 9. Bleul, C. C., R. C. Fuhlbrigge, J. M. Casasnovas, A. Aiuti, and T. A.
Springer.1996. A highly efficacious lymphocyte chemoattractant, stromal cell-derived factor 1 (SDF-1). J. Exp. Med.184:1101–1109.
10. Bleul, C. C., L. Wu, J. A. Hoxie, T. A. Springer, and C. R. Mackay.1997. The HIV coreceptors CXCR4 and CCR5 are differentially expressed and regu-lated on human T lymphocytes. Proc. Natl. Acad. Sci. USA94:1925–1930. 11. Cheˆne, L., M. Nugeyre, E. Guillemard, N. Moulian, F. Barre´-Sinoussi, and
N. Israe¨l.1999. Thymocyte-thymic epithelial cell interaction leads to high-level replication of human immunodeficiency virus exclusively in mature CD4⫹CD8⫺CD3⫹thymocytes: a critical role for tumor necrosis factor and interleukin-7. J. Virol.73:7533–7542.
12. Cheˆne, L., M. T. Nugeyre, F. Barre´-Sinoussi, and N. Israe¨l.1999. High-level replication of human immunodeficiency virus in thymocytes requires NF-B activation through interaction with thymic epithelial cells. J. Virol.73:2064– 2073.
13. Correa, R., and M. A. Munoz-Fernandez.2001. Viral phenotype affects the thymic production of new T cells in HIV-1-infected children. AIDS15:1959– 1963.
14. Davis, A. E., Jr.1984. The histopathological changes in the thymus gland in the acquired immune deficiency syndrome. Ann. N. Y. Acad. Sci.437:493– 502.
15. Deng, H., R. Liu, W. Ellmeier, S. Choe, D. Unutmaz, M. Burkhart, P. Di Marzio, S. Marmon, R. E. Sutton, C. M. Hill, C. B. Davis, S. C. Peiper, T. J. Schall, D. R. Littman, and N. R. Landau.1996. Identification of a major co-receptor for primary isolates of HIV-1. Nature381:661–666.
16. Deng, H. K., D. Unutmaz, V. N. KewalRamani, and D. R. Littman.1997. Expression cloning of new receptors used by simian and human immunode-ficiency viruses. Nature388:296–300.
17. Douek, D. C., R. D. McFarland, P. H. Keiser, E. A. Gage, J. M. Massey, B. F. Haynes, M. A. Polis, A. T. Haase, M. B. Feinberg, J. L. Sullivan, B. D. Jamieson, J. A. Zack, L. J. Picker, and R. A. Koup.1998. Changes in thymic function with age and during the treatment of HIV infection. Nature396:
690–695.
18. Feinberg, M. B., J. M. McCune, F. Miedema, J. P. Moore, and H. Schuite-maker.2002. HIV tropism and CD4⫹T-cell depletion. Nat. Med.8:537. 19. Guillemard, E., M. T. Nugeyre, L. Cheˆne, N. Schmitt, C. Jacquemot, F.
Barre´-Sinoussi, and N. Israe¨l.2001. Interleukin-7 and infection itself by human immunodeficiency virus 1 favor virus persistence in mature CD4⫹CD8⫺CD3⫹thymocytes through sustained induction of Bcl-2. Blood
98:2166–2174.
20. Haynes, B. F., L. P. Hale, K. J. Weinhold, D. D. Patel, H. X. Liao, P. B. Bressler, D. M. Jones, J. F. Demarest, K. Gebhard-Mitchell, A. T. Haase, and J. A. Bartlett.1999. Analysis of the adult thymus in reconstitution of T lymphocytes in HIV-1 infection. J. Clin. Investig.103:453–460.
21. Hazenberg, M. D., J. W. Stuart, S. A. Otto, J. C. Borleffs, C. A. Boucher, R. J. de Boer, F. Miedema, and D. Hamann.2000. T-cell division in human immunodeficiency virus (HIV)-1 infection is mainly due to immune
on November 8, 2019 by guest
http://jvi.asm.org/
tion: a longitudinal analysis in patients before and during highly active antiretroviral therapy (HAART). Blood95:249–255.
22. Hernandez-Lopez, C., A. Varas, R. Sacedon, E. Jimenez, J. J. Munoz, A. G. Zapata, and A. Vicente.2002. Stromal cell-derived factor 1/CXCR4 signaling is critical for early human T-cell development. Blood99:546–554. 23. Howard, T. H., and W. H. Meyer.1984. Chemotactic peptide modulation of
actin assembly and locomotion in neutrophils. J. Cell Biol.98:1265–1271. 24. Iyengar, S., J. E. Hildreth, and D. H. Schwartz.1998. Actin-dependent
receptor colocalization required for human immunodeficiency virus entry into host cells. J. Virol.72:5251–5255.
25. Jinquan, T., H. H. Jacobi, C. Jing, C. M. Reimert, S. Quan, S. Dissing, L. K. Poulsen, and P. S. Skov.2000. Chemokine stromal cell-derived factor 1␣
activates basophils by means of CXCR4. J. Allergy Clin. Immunol.106:313–320. 26. Joshi, V. V., J. M. Oleske, S. Saad, C. Gadol, E. Connor, R. Bobila, and A. B. Minnefor.1986. Thymus biopsy in children with acquired immunodeficiency syndrome. Arch. Pathol. Lab. Med.110:837–842.
27. Ka¨rber, G.1931. Beitrag zur kollektiven behandlung pharmakogisher reih-envesuche. Arch. Exp. Pathol. Pharmakol.162:956–959.
28. Koot, M., R. van Leeuwen, R. E. de Goede, I. P. Keet, S. Danner, J. K. Eeftinck Schattenkerk, P. Reiss, M. Tersmette, J. M. Lange, and H. Schuite-maker.1999. Conversion rate towards a syncytium-inducing (SI) phenotype during different stages of human immunodeficiency virus type 1 infection and prognostic value of SI phenotype for survival after AIDS diagnosis. J. Infect. Dis.179:254–258.
29. Kwa, D., J. Vingerhoed, B. Boeser-Nunnink, S. Broersen, and H. Schuite-maker.2001. Cytopathic effects of non-inducing and syncytium-inducing human immunodeficiency virus type 1 variants on different CD4⫹ -T-cell subsets are determined only by coreceptor expression. J. Virol.75:
10455–10459.
30. Lackner, A. A., P. Vogel, R. A. Ramos, J. D. Kluge, and M. Marthas.1994. Early events in tissues during infection with pathogenic (SIVmac239) and nonpathogenic (SIVmac1A11) molecular clones of simian immunodefi-ciency virus. Am. J. Pathol.145:428–439.
31. Lecossier, D., F. Bouchonnet, P. Schneider, F. Clavel, and A. J. Hance.2001. Discordant increases in CD4⫹T cells in human immunodeficiency virus-infected patients experiencing virologic treatment failure: role of changes in thymic output and T cell death. J. Infect. Dis.183:1009–1016.
32. Lederman, M. M., and H. Valdez.2000. Immune restoration with antiretro-viral therapies: implications for clinical management. JAMA284:223–228. 33. Llano, A., J. Barretina, A. Gutierrez, J. Blanco, C. Cabrera, B. Clotet, and
J. A. Este.2001. Interleukin-7 in plasma correlates with CD4 T-cell depletion and may be associated with emergence of syncytium-inducing variants in human immunodeficiency virus type 1-positive individuals. J. Virol.
75:10319–10325.
34. Menu, E., T. X. Truong, M. E. Lafon, T. H. Nguyen, M. C. Mu¨ller-Trutwin, T. T. Nguyen, A. Deslandres, G. Chaouat, Q. T. Duong, B. K. Ha, H. J. Fleury, and F. Barre´-Sinoussi.1996. HIV type 1 Thai subtype E is predom-inant in South Vietnam. AIDS Res. Hum. Retrovir.12:629–933. 35. Meyers, A., A. Shah, R. H. Cleveland, W. R. Cranley, B. Wood, S. Sunkle, S.
Husak, and E. R. Cooper.2001. Thymic size on chest radiograph and rapid disease progression in human immunodeficiency virus 1-infected children. Pediatr. Infect. Dis. J.20:1112–1118.
36. Misse, D., M. Cerutti, N. Noraz, P. Jourdan, J. Favero, G. Devauchelle, H. Yssel, N. Taylor, and F. Veas.1999. A CD4-independent interaction of human immunodeficiency virus-1 gp120 with CXCR4 induces their cointer-nalization, cell signaling, and T-cell chemotaxis. Blood93:2454–2462. 37. Muller-Trutwin, M. C., M. L. Chaix, F. Letourneur, E. Begaud, D.
Beau-mont, A. Deslandres, B. You, J. Morvan, C. Mathiot, F. Barre-Sinoussi, and S. Saragosti.1999. Increase of HIV-1 subtype A in Central African Repub-lic. J. Acquir. Immune Defic. Syndr.21:164–171.
38. Nahmias, A. J., W. S. Clark, A. P. Kourtis, F. K. Lee, G. Cotsonis, C. Ibegbu, D. Thea, P. Palumbo, P. Vink, R. J. Simonds, S. R. Nesheim, et al.1998. Thymic dysfunction and time of infection predict mortality in human immu-nodeficiency virus-infected infants. J. Infect. Dis.178:680–685.
39. Napolitano, L. A., R. M. Grant, S. G. Deeks, D. Schmidt, S. C. De Rosa, L. A. Herzenberg, B. G. Herndier, J. Andersson, and J. M. McCune.2001. In-creased production of IL-7 accompanies HIV-1-mediated T-cell depletion: implications for T-cell homeostasis. Nat. Med.7:73–79.
40. Pallard, C., A. P. Stegmann, T. van Kleffens, F. Smart, A. Venkitaraman, and H. Spits.1999. Distinct roles of the phosphatidylinositol 3-kinase and STAT5 pathways in IL-7-mediated development of human thymocyte pre-cursors. Immunity10:525–535.
41. Papiernik, M., Y. Brossard, N. Mulliez, J. Roume, C. Brechot, F. Barin, A. Goudeau, J. F. Bach, C. Griscelli, R. Henrion, and R. Vazeux.1992. Thymic abnormalities in fetuses aborted from human immunodeficiency virus type 1 seropositive women. Pediatrics89:297–301.
42. Pedroza-Martins, L., K. B. Gurney, B. E. Torbett, and C. H. Uittenbogaart.
1998. Differential tropism and replication kinetics of human immunodefi-ciency virus type 1 isolates in thymocytes: coreceptor expression allows viral entry, but productive infection of distinct subsets is determined at the postentry level. J. Virol.72:9441–9452.
43. Penn, M. L., J. C. Grivel, B. Schramm, M. A. Goldsmith, and L. Margolis.
1999. CXCR4 utilization is sufficient to trigger CD4⫹T cell depletion in HIV-1-infected human lymphoid tissue. Proc. Natl. Acad. Sci. USA96:663–668. 44. Richman, D. D., and S. A. Bozzette.1994. The impact of the syncytium-inducing phenotype of human immunodeficiency virus on disease progres-sion. J. Infect. Dis.169:968–974.
45. Rosenzweig, M., D. P. Clark, and G. N. Gaulton.1993. Selective thymocyte depletion in neonatal HIV-1 thymic infection. AIDS7:1601–1605. 46. Rosenzweig, M., M. Connole, A. Forand-Barabasz, M. P. Tremblay, R. P.
Johnson, and A. A. Lackner.2000. Mechanisms associated with thymocyte apoptosis induced by simian immunodeficiency virus. J. Immunol.165:3461– 3468.
47. Rothe, M., L. Cheˆne, M. Nugeyre, F. Barre´-Sinoussi, and N. Israe¨l.1998. Contact with thymic epithelial cells as a prerequisite for cytokines enhanced HIV-1 replication in thymocytes. J. Virol.72:5852–5861.
48. Savino, W., M. Dardenne, C. Marche, D. Trophilme, J.-M. Dupuy, D. Pek-ovic, N. Lapointe, and J.-F. Bach.1986. Thymic epithelium in AIDS. An immunologic study. Am. J. Pathol.122:302–307.
49. Scarlatti, G., V. Hodara, P. Rossi, L. Muggiasca, A. Bucceri, J. Albert, and E. M. Fenyo¨.1993. Transmission of human immunodeficiency virus type 1 (HIV-1) from mother to child correlates with viral phenotype. Virology
197:624–629.
50. Scarlatti, G., E. Tresoldi, A. Bjorndal, R. Fredriksson, C. Colognesi, H. K. Deng, M. S. Malnati, A. Plebani, A. G. Siccardi, D. R. Littman, E. M. Fenyo, and P. Lusso.1997. In vivo evolution of HIV-1 co-receptor usage and sensitivity to chemokine-mediated suppression. Nat. Med.3:1259–1265. 51. Schuitemaker, H., M. Koot, N. A. Kootstra, M. W. Dercksen, R. E. de Goede,
R. P. van Steenwijk, J. M. Lange, J. K. Schattenkerk, F. Miedema, and M. Tersmette.1992. Biological phenotype of human immunodeficiency virus type 1 clones at different stages of infection: progression of disease is asso-ciated with a shift from monocytotropic to T-cell-tropic virus population. J. Virol.66:1354–1360.
52. Signoret, N., M. M. Rosenkilde, P. J. Klasse, T. W. Schwartz, M. H. Malim, J. A. Hoxie, and M. Marsh.1998. Differential regulation of CXCR4 and CCR5 endocytosis. J. Cell Sci.111:2819–2830.
53. Smith, K. Y., H. Valdez, A. Landay, J. Spritzler, H. A. Kessler, E. Connick, D. Kuritzkes, B. Gross, I. Francis, J. M. McCune, and M. M. Lederman.
2000. Thymic size and lymphocyte restoration in patients with human im-munodeficiency virus infection after 48 weeks of zidovudine, lamivudine, and ritonavir therapy. J. Infect. Dis.181:141–147.
54. Spencer, L. T., M. T. Ogino, W. M. Dankner, and S. A. Spector.1994. Clinical significance of human immunodeficiency virus type 1 phenotypes in infected children. J. Infect. Dis.169:491–495.
55. Stanley, S. K., J. M. McCune, H. Kaneshima, J. S. Justement, M. Sullivan, E. Boone, M. Baseler, J. Adelsberger, M. Bonyhadi, J. Orenstein, C. H. Fox, and A. S. Fauci.1993. Human immunodeficiency virus infection of the human thymus and disruption of the thymic microenvironment in the SCID-hu mouse. J. Exp. Med.178:1151–1163.
56. Stoddart, C. A., T. J. Liegler, F. Mammano, V. D. Linquist-Stepps, M. S. Hayden, S. G. Deeks, R. M. Grant, F. Clavel, and J. M. McCune.2001. Impaired replication of protease inhibitor-resistant HIV-1 in human thymus. Nat. Med.7:712–718.
57. Taylor, J. R., Jr., K. C. Kimbrell, R. Scoggins, M. Delaney, L. Wu, and D. Camerini.2001. Expression and function of chemokine receptors on human thymocytes: implications for infection by human immunodeficiency virus type 1. J. Virol.75:8752–8760.
58. Tersmette, M., J. M. Lange, R. E. Y. de Goede, F. de Wolf, J. K. M. Eeftink-Schattenkerk, P. T. Schellekens, R. A. Coutinho, J. G. Huisman, J. Goudsmit, and F. Miedema.1989. Association between biological properties of human immunodeficiency virus variants and risk for AIDS and AIDS mortality. Lancet1:983–985.
59. Thelen, M.2001. Dancing to the tune of chemokines. Nat. Immunol.2:129– 134.
60. Theodore, T. S., G. Englund, A. Buckler-White, C. E. Buckler, M. A. Martin, and K. W. Peden.1996. Construction and characterization of a stable full-length macrophage-tropic HIV type 1 molecular clone that directs the pro-duction of high titers of progeny virions. AIDS Res. Hum. Retrovir.12:191– 194.
61. Uittenbogaart, C. H., D. J. Anisman, B. D. Jamieson, S. Kitchen, I. Schmid, J. A. Zack, and E. F. Hays.1996. Differential tropism of HIV-1 isolates for distinct thymocyte subsets in vitro. AIDS10:9–16.
62. Weiss, L., P. Ancuta, P. M. Girard, H. Bouhlal, A. Roux, N. H. Cavaillon, and M. D. Kazatchkine.1999. Restoration of normal interleukin-2 production by CD4⫹T cells of human immunodeficiency virus-infected patients after 9 months of highly active antiretroviral therapy. J. Infect. Dis.180:1057–1063. 63. Wykrzykowska, J. J., M. Rosenzweig, R. S. Veazey, M. A. Simon, K. Hal-vorsen, R. C. Desrosiers, R. P. Johnson, and A. Lackner.1998. Early regen-eration of thymic progenitors in rhesus macaques infected with simian im-munodeficiency virus. J. Exp. Med.187:1767–1778.
64. Zaitseva, M. B., S. Lee, R. L. Rabin, H. L. Tiffany, J. M. Farber, K. W. Peden, P. M. Murphy, and H. Golding.1998. CXCR4 and CCR5 on human thymo-cytes: biological function and role in HIV-1 infection. J. Immunol.161:3103– 3113.