Copyright © 1998, American Society for Microbiology. All Rights Reserved.
Differential Tropism and Replication Kinetics of Human
Immunodeficiency Virus Type 1 Isolates in Thymocytes:
Coreceptor Expression Allows Viral Entry, but
Productive Infection of Distinct Subsets
Is Determined at the Postentry Level
LIVIA PEDROZA-MARTINS,
1KEVIN B. GURNEY,
1BRUCE E. TORBETT,
2 ANDCHRISTEL H. UITTENBOGAART
1,3,4,5*
Department of Microbiology & Immunology,
1Department of Pediatrics,
3UCLA AIDS Institute,
4and Jonsson Comprehensive Cancer Center,
5UCLA School of Medicine, Los Angeles,
and The Scripps Research Institute, La Jolla,
2California
Received 1 June 1998/Accepted 24 August 1998
Human thymocytes are readily infected with human immunodeficiency virus type 1 (HIV-1) in vivo and in
vitro. In this study, we found that the kinetics of replication and cytopathic effects of two molecular isolates,
NL4-3 and JR-CSF, in postnatal thymocytes are best explained by the distribution of chemokine receptors used
for viral entry. CXCR4 was expressed at high levels on most thymocytes, whereas CCR5 expression was
restricted to only 0.1 to 2% of thymocytes. The difference in the amount of proviral DNA detected after infection
of fresh thymocytes with NL4-3 or JR-CSF correlated with the levels of CXCR4 and CCR5 surface expression.
Anti-CCR5 blocking studies showed that low levels of CCR5 were necessary and sufficient for JR-CSF entry in
thymocytes. Interleukin-2 (IL-2), IL-4, and IL-7, cytokines normally present in the thymus, influenced the
ex-pression of CXCR4 and CCR5 on thymocytes and thus increased the infectivity and spread of both NL4-3 and
JR-CSF in culture. NL4-3 was produced by both immature and mature thymocytes, whereas JR-CSF
produc-tion was restricted to the mature CD1
2/CD69
1population. Although CXCR4 and CCR5 distribution readily
explained viral entry in mature CD69
1and immature CD69
2cells, and correlated with proviral DNA
distri-bution, we found that viral production was favored in CD69
1cells. Therefore, while expression of CD4 and
ap-propriate coreceptors are essential determinants of viral entry, factors related to activation and stage-specific
maturation contribute to HIV-1 replication in thymocyte subsets. These results have direct implications for
HIV-1 pathogenesis in pediatric patients.
Human immunodeficiency virus (HIV) infection of the
thy-mus leads to loss of thymocytes and eventual thymic atrophy
(8, 29, 50, 53). While the role of the thymus in regeneration of
the immune system of HIV-infected adults has not been
es-tablished, the thymus is required for T-cell generation in
chil-dren (18, 39). Therefore, HIV infection of thymocytes and
thymic emigrants may have an impact on disease progression
in children. We and others have previously shown that NL4-3,
a molecularly cloned highly cytopathic CXCR4-tropic virus, as
well as certain pediatric HIV type 1 (HIV-1) isolates, are able
to replicate in immature and mature thymocyte subsets, while
JR-CSF, a relatively noncytopathic CCR5-tropic isolate, and
selected pediatric isolates have a more restricted tropism for
mature thymocyte subsets (27, 33, 64, 71, 73a). In addition,
interleukin-2 (IL-2), IL-4, and IL-7, cytokines implicated in
thymic subset expansion and maturation, have distinct effects
on HIV-1 replication (69, 70, 72, 73, 78). NL4-3 and some
pediatric isolates from rapid disease progressors replicated
faster in the presence of IL-4 plus IL-7 than in the presence of
IL-2 plus IL-4. In contrast, JR-CSF and isolates obtained from
pediatric patients with a slow disease progression replicated
faster in the presence of IL-2 plus IL-4 (20, 71, 72, 73a).
Surface expression of CD4 and of specific chemokine
core-ceptors allows HIV-1 entry into cells (13, 15, 17, 21). HIV-1
primary isolates can use CXCR4, CCR5, both receptors
(du-altropic isolates), or a number of other reported
seven-trans-membrane, G-protein-coupled chemokine receptors (3–5, 12,
14, 21, 34, 36, 60, 80). In adults, the critical role of CCR5 in
transmission and disease progression has been suggested by
genetic studies correlating resistance or delay of HIV-1
infec-tion with the presence of CCR5 mutainfec-tions that result in no or
low expression of CCR5 (25, 37, 55, 61). In children, the role
of CCR5 in transmission and disease progression has been
assessed in a cross-sectional study of children born to mothers
seropositive for HIV-1. Heterozygosity for CCR5
D
32 was not
associated with transmission but was associated with a slower
development of HIV-related disease in children (42).
Consis-tent with reports of studies of HIV-1-infected adults (12), viral
isolates obtained from children at early disease stages were
CCR5 tropic, while those from later stages of disease used
CXCR4 as a coreceptor (56). Early acquisition of CXCR4
usage by these viral isolates was associated with rapid disease
progression (12, 56).
In the thymus, where CD4 is expressed on more than 95% of
the cells, the distribution of HIV coreceptors would be
ex-pected to be an important determinant of tropism. Wide
dis-tribution of CXCR4 surface expression on fetal thymocytes has
been recently reported (31), while expression of the
corecep-tors CCR5, CCR8, and STLR-33/GPR15 on total thymocytes
has been reported at the mRNA level (36, 46, 51, 54, 68).
* Corresponding author. Mailing address: Department of
Microbi-ology and ImmunMicrobi-ology, UCLA School of Medicine, Los Angeles, CA
90095-1747. Phone: (310) 825-1982. Fax: (310) 206-1318. E-mail:
9441
on November 9, 2019 by guest
http://jvi.asm.org/
Other chemokine receptors, such as CCR4 (49), not yet
iden-tified as HIV coreceptors, are also present in the thymus.
Finally, three unique thymic orphan chemokines,
macrophage-derived cytokine (MDC), thymus- and activation-regulated
cytokine (TARC), and thymus-expressed cytokine (TECK),
whose as yet unidentified receptors could potentially support
HIV-1 entry have been detected (19, 24, 77).
After viral entry, the activation state of the target cell
de-termine its ability to reverse transcribe, integrate, and support
HIV replication (63, 65, 82). In peripheral blood mononuclear
cells (PBMC), for example, full reverse transcription requires
at least progression to the G
1bphase of the cell cycle and
therefore is dependent on the activation state of the cell (32,
82). Thymocytes are a heterogeneous population of cells in
terms of differentiation and activation. In this study, we
exam-ined HIV replication in thymocyte subsets defexam-ined by the
ex-pression of surface molecules that are commonly used as
mark-ers of T-cell development: CD1, CD69, and CD45RA. The
CD1 molecule is expressed at high levels in CD3
2/lowthymo-cytes and therefore identifies immature thymothymo-cytes (7).
Down-regulation of CD1 correlates with acquisition of functional
maturation of thymocytes (52). During the process of positive
selection, the activation marker CD69 is expressed on 10% of
CD4
1/CD8
1double-positive thymocytes and at high levels on
mature single-positive CD4
1and CD8
1cells (67). However,
CD69 expression is absent on thymocytes that emigrate from
the thymus to the periphery (52, 76). By contrast, the CD45RA
antigen, a marker of naive cells in the periphery, is expressed
only on mature CD3
1/high/CD1
2thymocytes that are ready to
leave the thymus (66).
We took advantage of the differential tropism of JR-CSF
and NL4-3 for thymocyte subsets to study the distribution and
the usage of CCR5 and CXCR4 as HIV coreceptors on freshly
isolated postnatal thymocytes. The chemokine receptors CCR5
and CXCR4 have been reported as coreceptors for JR-CSF
and NL4-3, respectively, in PBMC and transfected cell lines
(17, 60, 80). We found that postnatal thymocytes expressed
high levels of CXCR4 and low levels of CCR5. In postnatal
thymocytes, CXCR4 was broadly distributed on immature and
mature subsets, as previously reported for fetal thymocytes
(31). Nevertheless, CCR5 expression on a low percentage of
thymocytes is necessary and sufficient to support replication of
a CCR5-tropic isolate. We also demonstrate that both CXCR4
and CCR5 support viral entry into CD69
1and CD69
2cells,
whereas only the CD69
1thymocyte subset sustained a highly
productive infection. These results help explain the reported
HIV-1-induced pathogenesis of the thymus by distinct HIV-1
tropic isolates.
MATERIALS AND METHODS
Reagents and monoclonal antibodies.Recombinant human IL-2 (1.53106
U/ml) and IL-4 (0.7 mg/ml) were provided by Amgen, Inc. (Thousand Oaks, Calif.). Recombinant human IL-7 (100mg/ml) was a gift from Immunex Corp. (Seattle, Wash.). 7-Amino-actinomycin D (7-AAD) was obtained from Sigma (St. Louis, Mo.). Actinomycin D (AD) was obtained from Boehringer Mannheim (Indianapolis, Ind.). Normal mouse immunoglobulin G (IgG; 3 mg/ml) was obtained from Caltag (Burlingame, Calif.). Monoclonal antibodies to CD8, CD4, CD3, CD45RA, and CD69 conjugated with fluorescein (FITC), phycoerythrin (PE), or peridinin chlorophyll protein (PerCP) and goat anti-mouse IgG-FITC were obtained from Becton Dickinson Immunocytometry Systems (BDIS; San Jose, Calif.). The antibodies KC57-FITC and KC57-PE, which identify intracel-lular HIV p24gagantigen expression (10, 40), CD1-PE, CD45RA-PE, and the
unconjugated antibodies CD45RA and CD69, used for thymocyte subset sepa-rations, were obtained from Coulter/Immunotech (Hialeah, Fla.). Unconjugated CXCR4 and CXCR4-PE (12G5) were obtained from Pharmingen (San Diego, Calif.). Unconjugated monoclonal antibodies to the chemokine receptors CCR-3 (7B11) (21, 23) and CCR-5 (2D7) (79) were obtained through the AIDS Re-search and Reference Reagent Program, Division of AIDS, National Institute of Allergy and Infectious Diseases, National Institutes of Health. CXCR-4 (12G5)
was a gift from James Hoxie (16). The monoclonal antibody to CCR-5 (3A9) was a gift from LeukoSite, Inc. (80). CD4-IgG was a gift from Genentech (San Francisco, Calif.).
Freshly isolated, nonstimulated PBMC were used as the positive control for CCR5 detection. In the same PBMC adult donors, CCR5 was present in 14 to 20% of the lymphocytes stained with 2D7 but only 2 to 3% of cells stained with 3A9. However, staining with antibodies 3A9 and 2D7 gave similar results on monocytes from these donors and in freshly isolated postnatal thymocytes. To rule out the possibility that the epitopes recognized by these antibodies were not exposed on the surface of thymocytes, cells were permeabilized and stained intracellularly with antibody 2D7-PE (58). The intracellular level of CCR5 was below the detection level on thymocytes but was detectable in PBMC, although at lower levels than on the cell surface.
HIV infection and thymocyte cultures.Normal pediatric thymuses were ob-tained in the course of corrective cardiac surgery. Single-cell suspensions and nylon wool purification were done as previously described, and thymocytes were cultured at 13107to 23107cells/ml in serum-free medium
(albumin-trans-ferrin-IMDM [Iscove’s modified Dulbecco’s medium]; Irvine Scientific, Santa Ana, Calif.) supplemented with delipidated bovine serum albumin (BSA; Sigma) at 1,100mg/ml, transferrin (Sigma) at 85mg/ml, glutamine at 2 mM (0.3 mg/ml), and penicillin-streptomycin at 25 U/ml–25mg/ml (73, 78). Thymocytes were cultured in the presence or absence of the cytokines IL-2 (20 U/ml), IL-4 (20 ng/ ml), and IL-7 (200 U/ml).
Two molecular clones of HIV-1, the non-syncytium-inducing, CCR5-tropic clone JR-CSF (33) and the syncytium-inducing, CXCR-4-tropic hybrid clone NL-4-3, were used for these studies (1). Virus stocks of JR-CSF were prepared from 24-h harvests of supernatants from PBMC infected with the supernatant of COS cells electroporated with plasmid pYKJR-CSF. Virus stocks of NL4-3 were prepared from 24-h harvests of supernatants from CEM cells (CCRF-CEM) infected with the supernatant of COS cells electroporated with plasmid pNL4-3. Virus stocks were stored at270°C and treated with DNase (2mg/ml; Worthing-ton, Lakewood, N.J.) for 30 min at room temperature in the presence of 0.01 M MgCl2before infections. Heat-inactivated controls were obtained by incubating
DNase-treated viruses at 65°C for 45 min. All infections were standardized by determining infectious units (IU) in limiting dilution studies using phytohemag-glutinin (PHA)-stimulated PBMC (81, 82). For thymocyte infections, JR-CSF was used at 10- to 20-fold higher multiplicity of infection (MOI) than NL4-3 unless otherwise indicated.
Thymocytes were infected and cultured as previously described (72). Briefly, virus infection was accomplished by incubating thymocytes with 30 to 200 ng of viral p24/107cells in the presence of Polybrene (10mg/ml; Sigma) for 1 to 2 h at
37°C. Control thymocytes were sham infected in the presence of Polybrene with supernatant from uninfected cells that were used for preparing the virus stocks. After infection, the cells were washed extensively in A-IMDM and resuspended in serum-free medium in the presence of cytokines. On day 1 postinfection and weekly thereafter, the supernatant was removed and the cells were fed with fresh medium and cytokines. Virus expression was assessed by measuring p24 antigen in the supernatant by enzyme-linked immunosorbent assay (Coulter, Hialeah, Fla.).
Blocking studies using antibodies to chemokine receptors.Thymocytes were preincubated with antibodies to CCR5 (2D7; 1 to 5mg/107cells) and/or CXCR4
(12G5; 5 to 10mg/107cells) or CD4-IgG (100mg/107cells) at 4°C for 1 to 2 h
before infection. The antibodies and CD4-IgG were present during infection and throughout the duration of the experiment. On day 1 and weekly thereafter, the medium was removed and fresh medium containing the antibody was added, while CD4-IgG was added on days 1 and 7 only.
Isolation of thymocyte subsets.Magnetic beads were used to isolate thymocyte subsets. In initial experiments, magnetic beads coated with goat anti-mouse IgG-plus-IgM antibody (Kirkegaard & Perry, Gaithersburg, Md.) were used (73, 78). Since the Kirkegaard & Perry beads are no longer available, Dynal (Lake Success, N.Y.) M280 magnetic beads coated with sheep anti-mouse IgG were used in later experiments. Comparisons showed that subset purities were similar in assays using the beads from the two manufacturers. CD45RA- and CD69-positive and -negative subsets were obtained as follows. Magnetic beads were preincubated at 108beads/ml in A-IMDM containing 1% BSA to prevent
non-specific binding to thymocytes and then coated with the CD45RA or CD69 monoclonal antibody (1.25 tests of the antibody as determined by the manufac-turer/108beads/ml) for at least 18 h at 4°C. Beads were washed once to remove
excess unbound antibody immediately before use. For depletion of CD45RA1 cells, thymocytes were combined with CD45RA-coated beads at a bead-to-cell ratio of 1:2 and rotated at 4°C for 1 h. Cells bound to beads were removed with a magnet (Collaborative Research, Inc., Bedford, Mass.) and subjected to a second round of depletion. The CD45RA-depleted cells were then combined with the CD69-coated beads at a bead-to-cell ratio of 2:1 and rotated at 4°C for 1 h. The CD45RA-depleted cells bound to CD69-coated beads (CD691 popu-lation) were magnetically removed, and unbound cells (CD692population) were subjected to a second round of depletion. Following separation, the depleted subsets were immunophenotyped and analyzed by flow cytometry. Both posi-tively and negaposi-tively immunoselected subsets were used for infection and culture experiments. Their viability, as determined by trypan blue dye exclusion, was .96%.
on November 9, 2019 by guest
http://jvi.asm.org/
Immunofluorescent staining and flow cytometry.Surface and cytoplasmic im-munophenotyping of thymocytes with directly conjugated antibodies were done as previously described (57, 58). When unconjugated antibodies were used, cells were washed in phosphate-buffered saline (PBS) containing 1% BSA (PBS-BSA). After blocking with 50ml of human AB serum to prevent nonspecific protein binding, thymocytes (13105to 53105) were incubated with optimal
amounts of unconjugated monoclonal antibody for 20 min at 4°C in a total volume of 100ml and then washed with 3 ml of PBS-BSA. Goat anti-mouse IgG-FITC antibody was added for 20 min at 4°C in the presence of 50ml of human AB serum. Cells were washed with 3 ml of PBS-BSA and incubated for 10 min at 4°C with 50ml of mouse IgG (3 mg/ml) diluted 1:15 in PBS-BSA to prevent nonspecific protein binding before incubation with directly conjugated PE- or PerCP-labeled antibodies for 20 min at 4°C. To exclude dead cells, the thymocytes were incubated in a solution of 2mg of 7-AAD per ml in PBS for 20 min at 4°C protected from the light. The cells were washed in PBS and incubated in 1% paraformaldehyde solution in PBS containing 4mg of AD per ml (57, 59). The samples were subjected to flow cytometric analysis in the paraformalde-hyde-AD solution.
A FACScan flow cytometer equipped with a standard filter setup (BDIS) was used in these experiments. A minimum of 10,000 events was acquired on each sample. Multiparameter data acquisition and analysis were performed with Cell Quest software (BDIS).
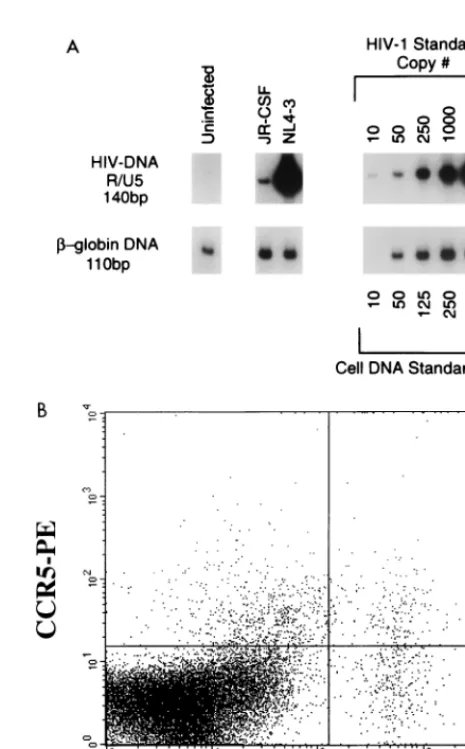
Quantitative DNA PCR.At 16 to 20 h postinfection, 106thymocytes were
removed from the cultures, washed once in PBS, lysed in urea lysis buffer (4.7 M urea, 1.3% [wt/vol] sodium dodecyl sulfate, 0.23 M NaCl, 0.67 mM EDTA [pH 8.0], 6.7 mM Tris-HCl), and then subjected to multiple phenol-chloroform ex-tractions and ethanol precipitation. Total nucleic acids obtained from thymocytes were subjected to quantitative DNA PCR as described previously (2, 81, 82). HIV DNA was detected by using the32P-end-labeled M667-AA55 primer pair
specific for the R/U5 region of the viral long terminal repeat (LTR) (81, 82). For detection of full-length reverse transcripts, the M667-M661 primer pair specific for the LTR/gag region was used (32, 82). Products obtained after 25 cycles of amplification were resolved on a 6% polyacrylamide gel. Standard curves for HIV-1 DNA were generated by using various dilutions of plasmid pYKJR-CSF linearized with EcoRI, which does not digest viral sequences. The dilutions were made into DNA from normal human PBMC (10mg/ml). To normalize for cellular DNA, replicate samples were analyzed for humanb-globin gene se-quences (35, 81) by 25 cycles of amplification. Standard curves for human DNA were generated from two- and fivefold dilutions of PBMC DNA. Values were obtained by interpolation from the standard curves, using a radioanalytic imaging system (Ambis, San Diego, Calif.).
RESULTS
Cell surface expression of CCR5, CCR3, and CXCR4 on
thymocytes from children.
Postnatal thymus specimens
ob-tained from 18 children (both sexes, 15 days to 4 years old)
were used for these studies. Freshly isolated thymocytes were
immunophenotyped with antibodies to CCR5 (2D7 and/or
3A9) and CXCR4 (12G5) to determine the thymic distribution
of chemokine receptors that are reportedly the coreceptors for
JR-CSF and NL4-3, respectively, in transfected CD4
1cells
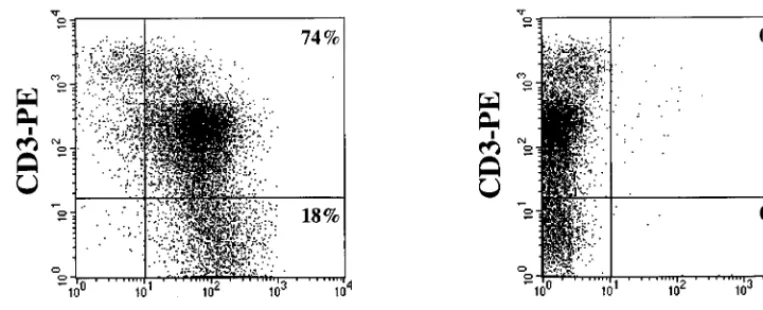
and PBMC (60, 80). In all specimens analyzed, more than 95%
of postnatal thymocytes expressed CXCR4, while the
percent-ages of CCR5
1cells ranged from 0.2 to 1% (mean
6
standard
deviation
5
0.45%
6
0.22%). A representative experiment is
shown in Fig. 1. The same coreceptor expression profile was
found in thymocyte single-cell suspensions before and after
nylon wool purification to enrich for T cells (data not shown).
Figure 1 shows that high levels of CXCR4 expression were
found in the immature CD3
2and CD3
1/lowsubsets, while the
mature CD3
1/highsubset contained CXCR4
1and CXCR4
2cells, as previously reported for fetal thymocytes (31). The
determination of CCR5 expression on distinct thymocyte
sub-sets by immunofluorescence methods was hampered by the low
numbers of CCR5
1thymocytes. CCR3 surface expression was
not detectable with antibody 7B11 in any of the eight
thymo-cyte samples tested (not shown).
Cytokines that favor HIV production by thymocytes
upregu-late CCR5 and CXCR4 surface expression.
Expression of
CCR5 and CXCR4 is tightly regulated on PBMC by
stimula-tory signals, mitogens, and cytokines such as IL-2 and IL-10 (6,
9, 38, 44, 45, 62, 80). We have previously shown that cytokines
involved in thymocyte maturation distinctly regulate the
ex-pression of JR-CSF and NL4-3 in thymocyte subsets in vitro
(72). To investigate the effect of these cytokines on chemokine
receptor expression, cells were immunophenotyped at day 0
and cultured in serum-free medium in the presence of IL-2,
IL-4, IL-7, IL-2 plus IL-4, or IL-4 plus IL-7 for 2 weeks. These
cytokines affect proliferation and differentiation of different
subpopulations, which results in different proportions of cells
expressing high levels of CD3 (i.e., a CD3
1/highpopulation)
(72, 73, 78). Cell surface phenotype was determined weekly,
and the cursors were set in order to analyze coreceptor
expres-sion in the CD3
1/highpopulation (Fig. 2; Table 1).
Thymocytes cultured in the presence of IL-2 plus IL-4 or
IL-4 plus IL-7 showed increased levels of CXCR4 expression
as measured by fluorescence intensity and by the absence of
CXCR4
2cells (Fig. 2A). IL-4 alone was sufficient to increase
[image:3.612.109.491.73.228.2]the levels of CXCR4 expression (Table 1). Interestingly, IL-4
increased the expression of CXCR4 in the mature CD3
1/highpopulation that expresses low levels of CXCR4 in freshly
iso-lated thymocytes. In thymocytes cultured with IL-2 alone,
there was a threefold increase in the fluorescence intensity and
in the percentage of CXCR4
1/CD3
1/highcells (Table 1), but
the mature CD3
1/highpopulation that did not express CXCR4
FIG. 1. Distribution of chemokine receptor expression on freshly isolated human thymocytes. Thymocytes were isolated by nylon wool separation and phenotyped with CD3-PE and nonlabeled antibodies to the chemokine receptors CXCR-4 (antibody 12G5) and CCR5 (antibody 2D7), followed by goat anti-mouse IgG-FITC (GAM-FITC). Appropriate isotype control antibodies (IgG2a and IgG1) followed by goat anti-mouse IgG-FITC were used to set the cursors. The percentage of cells staining with the isotype control antibodies followed by goat anti-mouse IgG-FITC was 0%. Dead cells were excluded from the analysis by using 7-AAD.on November 9, 2019 by guest
http://jvi.asm.org/
was also expanded (data not shown). Thymocytes cultured with
IL-7 showed this same profile (Table 1).
In contrast, the percentage of cells expressing CCR5
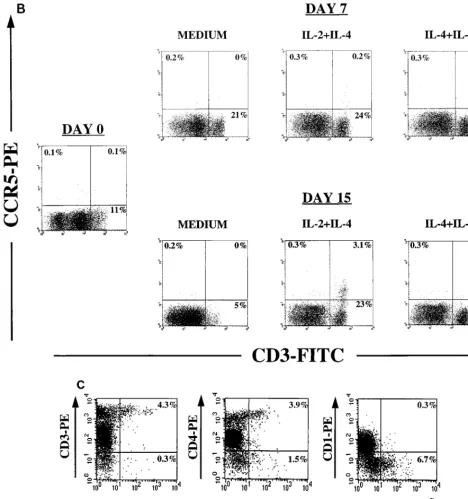
in-creased after 2 weeks of culture with IL-2 plus IL-4 but not in
the presence of IL-4 plus IL-7 (Fig. 2B). In six of seven
thy-mocyte culture experiments, IL-2 and IL-4 synergistically
in-creased the percentages of CCR5-expressing cells from 0.2 to
1% on day 0 to 1 to 6% after 2 weeks of culture. No effect of
IL-4 or IL-7 alone on CCR5 expression was seen, whereas
upregulation of CCR5 expression in the CD3
1/highby IL-2
alone was observed in only one of seven experiments. Further
analysis of CCR5 distribution in thymocytes cultured in IL-2
plus IL-4 showed that CCR5 was expressed on the CD3
1/high/
CD4
1/high/CD1
2thymocyte subset, in which we have
previ-ously observed JR-CSF expression (Fig. 2C) (71).
NL4-3 and JR-CSF replication kinetics in thymocytes
cor-relate with the expression levels of CXCR4 and CCR5.
The
role of chemokine receptors in the different kinetics of
repli-cation of JR-CSF and NL4-3 in thymocytes was studied in
vitro. Levels of CCR5 and CXCR4 expression were assessed
before and after infection on freshly isolated thymocytes.
Twenty-four hours after infection with JR-CSF (200 IU/10
4cells) or NL4-3 (10 IU/10
4cells), 10
6cells were taken and
analyzed by PCR for proviral DNA content as described
pre-viously (81). The level of proviral DNA in thymocytes infected
with JR-CSF was significantly lower than the level of proviral
DNA detected in thymocytes infected with NL4-3 (Fig. 3A),
despite the 20-fold-higher MOI of JR-CSF, as determined in
PHA-stimulated PBMC. This finding suggests that the
differ-ence observed between the replication kinetics of the two
vi-ruses is determined at the entry level. The copy number of
NL4-3 proviral DNA in thymocytes (more than 50 copies/ng)
correlated with the high numbers of cells expressing CD4 and
CXCR4 in the thymus. The low level of JR-CSF proviral DNA
in thymocytes (approximately 1 copy/ng) correlated with the
low level of CCR5 surface expression (0.4%) on the specimen
analyzed on day 0 (Fig. 1 and 3). Nevertheless, at 2 weeks
postinfection p24 levels in the supernatant of JR-CSF infected
cells reached 110 ng/ml. Notably, in all of eight infection
ex-periments, thymus specimens containing at most 1% CCR5
1 [image:4.612.59.547.67.428.2]cells at the time of infection (day 0) were able to sustain
FIG. 2. Effects of cytokines on chemokine receptor expression. Thymocytes were cultured for 2 weeks in serum-free medium alone or supplemented with IL-2 plus IL-4 or IL-4 plus IL-7. Before culture (day 0) and on days 7 and 15, cells were removed for immunophenotyping to examine expression of chemokine receptors by flow cytometry. Appropriate isotype control antibodies and single-color staining with CD3-FITC were used to set the cursors defining the CD31/highpopulation (A and B).
Dead cells were excluded from the analysis by using 7-AAD. (A and B) CXCR4 and CCR5 expression on thymocyte subsets was determined by using the antibodies CD3-FITC, CXCR4-PE (12G5), and CCR5-PE (2D7). (C) In a different experiment, immunophenotyping was performed on day 12 of culture to identify the distribution of CCR5 on thymocyte subsets that respond to IL-2 plus IL-4. Thymocytes were stained with CD3-PE, CD4-PE, or CD1-PE in combination with nonlabeled antibody to CCR5 (2D7) and then with goat anti-mouse IgG-FITC (GAM-FITC). The percentage of cells staining with the IgG1 isotype control antibody for CCR5 followed by goat anti-mouse IgG-FITC was 0%.
on November 9, 2019 by guest
http://jvi.asm.org/
JR-CSF production, as measured by p24 levels in the
super-natant.
To determine if JR-CSF was produced by the small subset of
CCR5
1thymocytes, cell surface staining was combined with
intracellular staining for HIV Gag proteins with the KC57
antibody (10, 40, 71). As observed for uninfected thymocytes,
expression levels of both CXCR4 and CCR5 increased on
mature thymocytes during culture of infected cells with IL-2
plus IL-4, although the percentage of CCR5
1positive cells
detected remained below 10% (Fig. 3B). At 2 weeks after
infection with JR-CSF, KC57
1cells were detected in both the
CCR5
1and CCR5
2populations (Fig. 3B). In subsequent
ex-periments, at later time points, JR-CSF expression was
de-tected only in the CCR5
2population, in a manner reminiscent
of the presence of HIV expression in the CD4
2thymocyte
subpopulation at late stages of infection (30).
Therefore, the slower replication of JR-CSF compared to
that of NL4-3 in thymocytes correlated with lower levels of
proviral DNA after infection. This observation could be
attrib-uted at least in part to the differences in the availability of cells
expressing CD4 and the appropriate coreceptor, presumably
CCR5 and CXCR4, at the time of infection.
Low levels of CCR5 support replication of JR-CSF in
thy-mocytes.
The presence of JR-CSF in the CCR5
2population
[image:5.612.66.535.73.572.2]could indicate downregulation of CCR5 on JR-CSF-infected
cells. However, JR-CSF adaptation to CXCR4 in culture and/
or usage of an alternative coreceptor could not be excluded in
the experiments described above. The antibody 2D7 was used
FIG. 2—Continued.
on November 9, 2019 by guest
http://jvi.asm.org/
to determine if JR-CSF replication in thymocytes could be
prevented by blocking the coreceptor CCR5 (79). In all of four
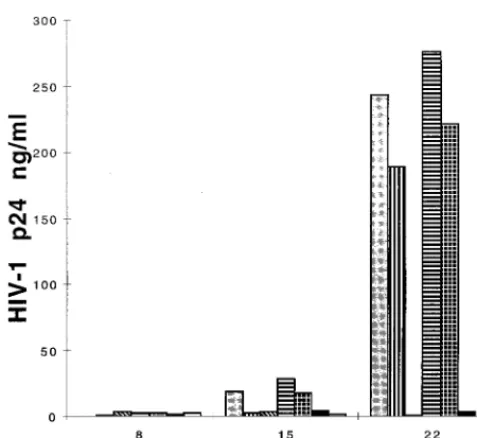
experiments, the p24 levels were reduced up to 50-fold in
JR-CSF-infected cells cultured in the presence of 2D7, while in the
presence of the 12G5 antibody to CXCR4 (16, 41) there was
no reduction in p24 levels. As can be seen in Fig. 4, thymocytes
infected with JR-CSF (30 IU/10
4cells) produced high levels of
p24 at 3 weeks postinfection. However, there was a delay in
HIV expression in the presence of 1
m
g of 2D7 per ml, while
with 5
m
g/ml the p24 levels were barely detectable up to 3 weeks
postinfection. Pretreatment and culture of thymocytes in the
presence of antibodies to both CXCR4 and CCR5 gave the
same results as treatment with antibody to CCR5 alone,
indi-cating poor, if any, usage of CXCR4 by JR-CSF in this system.
Addition of 5
m
g of 2D7 per ml 1 h after infection of
thymo-cytes with JR-CSF decreased p24 peak levels to 16 ng/ml,
compared to 354 ng/ml in thymocytes cultured in the absence
of antibody.
To determine if the effect of 2D7 on HIV replication on
thymocytes was due to blocking of CCR5 coreceptor function
and not to another effect of 2D7 on the cells which prevented
them from producing virus, thymocytes were pretreated with 5
m
g of 2D7 per ml and infected with either JR-CSF or NL4-3.
Production of NL4-3 was not blocked by antibody to CCR5 but
could be partially blocked by antibody to CXCR4 at 10
m
g/ml
(data not shown). The p24 data were confirmed by intracellular
staining with the KC57 antibody 2 and 3 weeks after infection
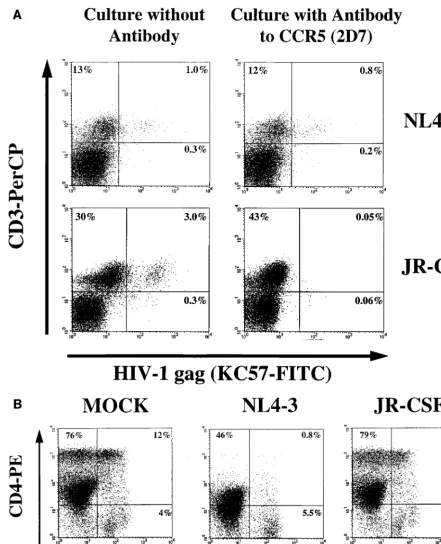
(Fig. 5). After infection with JR-CSF, KC57 expression was
detected in 3% of the untreated thymocytes, as opposed to
0.05% of the cells in the presence of 5
m
g of CCR5 antibody
per ml. In thymocytes infected with NL4-3, percentages of
KC57
1cells were similar in the nontreated thymocytes and in
the thymocytes treated with antibody to 2D7 at 2 weeks
postin-fection (Fig. 5A). In addition, when 2D7 was present, a
pro-found depletion of CD4
1thymocytes, a hallmark of NL4-3
infection, had already taken place (Fig. 5B). Cells infected with
JR-CSF in the presence of 2D7 did not have a CD4/CD8
profile significantly different from that of mock infected cells
cultured with 2D7 (Fig. 5B).
Taken together, these results suggest that JR-CSF uses
CCR5 as a coreceptor in thymocytes. Furthermore, they
indi-cate that very low levels of CCR5 surface expression can
sup-port replication of CCR5-tropic viruses in the thymus.
JR-CSF and NL4-3 production by different thymocyte
sub-sets is determined at the postentry level.
We used the CD69
and CD45RA molecules as markers of thymocyte development
to further characterize the thymocyte subsets susceptible to
JR-CSF and NL4-3 productive infection. Thymocytes subsets
at different stages of maturation were obtained by using
anti-body-coated magnetic beads to negatively and positively select
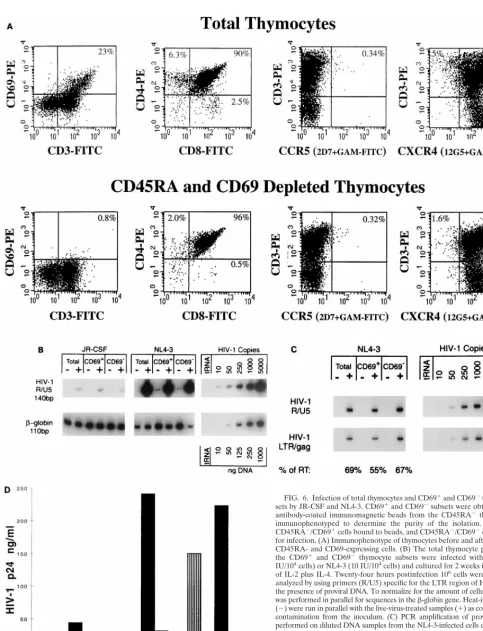
specific subsets. As shown in Fig. 6A, thymocytes expressing
CD69 are found in the mature CD3
1/highsubset, which
in-cludes CD4 and CD8 single-positive cells and 5 to 10% of the
CD4
1/CD8
1thymocytes as previously described (67, 76).
Ma-ture CD3
1/high/CD45RA
1cells were removed before CD69
depletion to eliminate the most mature CD4 and CD8
single-positive thymocytes that are CD69
2but express CD45RA (7,
52, 66, 76). In the experiment shown in Fig. 6, this procedure
removed all but 0.8% of the CD69
1cells, which included
[image:6.612.50.289.90.185.2]single-positive CD4
1and CD8
1cells, but did not remove the
FIG. 3. CXCR4 and CCR5 expression levels correlate with the amount of NL4-3 and JR-CSF proviral DNA after infection. Thymocytes were infected with JR-CSF (200 IU/104cells) or NL-4-3 (10 IU/104cells) and cultured with IL-2
plus IL-4. CCR5 expression on day 0 is shown in Fig. 1. (A) Twenty-four hours postinfection, 106cells were removed and analyzed by using primers (R/U5)
[image:6.612.310.543.262.637.2]specific for the LTR region of HIV-1 to detect the presence of proviral DNA. To normalize for the amount of cellular DNA, PCR was performed in parallel for sequences in theb-globin gene. (B) JR-CSF-infected thymocytes were cultured with IL-2 plus IL-4. At 13 days postinfection, cells were subjected to surface staining with CCR5-PE/CD3-PerCP followed by intracellular staining with KC57-FITC.
TABLE 1. Effects of cytokines on CXCR4 expression
in thymocytes
aCytokine
Mean fluorescence intensity of CXCR4 on CD31/high
cells (% CD31/high/CXCR41cells)b
Expt 1 Expt 2 Expt 3 Expt 4
None
47 (3)
38 (2)
37 (6)
37 (5)
IL-2
60 (9)
118 (21)
139 (28)
ND
IL-4
444 (22)
354 (24)
359 (29)
256 (15)
IL-7
ND
70 (25)
ND
160 (24)
IL-2
1
IL-4
431 (29)
489 (33)
361 (35)
ND
IL-4
1
IL-7
ND
398 (33)
ND
326 (17)
aFreshly isolated thymocytes were cultured for 2 weeks in serum-free medium
in the presence or absence of cytokines. Surface expression of CXCR4 was determined by flow cytometry using directly conjugated antibodies specific for CD3 (FITC) and CXCR4 (PE), with 7-AAD used to exclude dead cells.
bThe geometric mean of the fluorescence intensity of CXCR4 in the CD31/
highpopulation and the percentage of these cells in the gated population were
calculated with the Cell Quest software. Cursors were set by using isotype controls for all cytokine conditions within an individual experiment. Single-color staining with CD3-FITC was used to identify the CD31/highpopulation. Note
that the percentage of total CD31/highcells depended on the cytokine used (Fig.
2A). ND, not determined.
on November 9, 2019 by guest
http://jvi.asm.org/
cells expressing CCR5 or CXCR4 (Fig. 6A). In other
experi-ments, CCR5 expression was very low or below the detection
level in CD69
2cells while present in low levels in the CD69
1population; therefore CCR5 expression could not be
as-cribed to specific thymocyte subsets defined by CD69
ex-pression (data not shown). Given the low levels of CCR5
surface expression on thymocytes and the results obtained in
the blocking experiments, we tried to determine which
thymo-cyte subsets expressed CCR5 on the basis of their susceptibility
to JR-CSF infection (see below).
The total population and the immunoselected thymocyte
subsets were infected with either JR-CSF (100 IU/10
4cells) or
NL4-3 (10 IU/10
4cells), and virus production was monitored
by measuring p24 levels in the culture supernatants for up to 3
weeks. Proviral DNA levels in the distinct thymocyte subsets
were assessed by using PCR primers detecting partial and
full-length reverse transcription. Figure 6B shows that the amount
of proviral DNA in the total population infected by NL4-3 or
JR-CSF correlated with the expression levels of the respective
coreceptors as shown in Fig. 3A. JR-CSF and NL4-3 proviral
DNA could be detected in both CD69
2and CD69
1popula-tions, suggesting that viral entry occurred in both subsets.
However, NL4-3 copy number in each subset was at least
1,000-fold higher than JR-CSF copy number in the same
sub-set. The copy number of NL4-3 DNA was slightly higher in the
CD45RA
2/CD69
2cells than in the CD45RA
2/CD69
1cells,
[image:7.612.51.290.68.287.2]while the low copy number of JR-CSF DNA did not permit a
quantification of proviral levels in the different subsets. These
results indirectly suggest that CCR5 was expressed in both
populations, albeit at very low levels, confirming the phenotype
determined by flow cytometry (Fig. 6A). The ability of the
different subsets to complete reverse transcription after NL4-3
infection was assessed by amplifying the DNA samples with
primers detecting the LTR/gag region (32, 81, 82) as shown in
Fig. 6C. Full-length reverse transcripts were present in the
total population and in both CD69
1and CD69
2thymocyte
subsets at relative levels (
.
50%) that indicate completion of
reverse transcription in all subsets (Fig. 6C). Yet, in five of five
experiments, the levels of p24 were higher in the supernatant
of CD69
1cells than in the supernatant of CD69
2cells after
infection with JR-CSF or NL4-3 (Fig. 6D and data not shown).
This difference in viral expression was observed in the presence
of the appropriate coreceptors and of similar amounts of
pro-viral DNA in both thymocyte subsets (Fig. 6A, B, and D).
These results suggest that postentry events determine the
ability of HIV to preferentially replicate in the more mature
CD69
1thymocyte subset. Furthermore, full reverse
transcrip-tion and low levels of p24 expression were detected in CD69
2cells infected with NL4-3, indicating that late events in the
virus cycle are possibly involved in the differential tropism of
HIV for different thymocyte subsets. The expression level of a
given virus isolate (JR-CSF or NL4-3) in these different
thy-mocyte subsets was not determined at the entry level, although
the differences between expression of different virus isolates in
a given subset (i.e., CD69-depleted cells) could be explained by
availability of the respective coreceptors.
DISCUSSION
In this study, we have demonstrated that the distribution of
CXCR4 and CCR5 on thymocytes is a major determinant for
NL4-3 and JR-CSF tropism and determines the replication
kinetics of these two isolates (71). The majority of freshly
isolated postnatal thymocytes from uninfected children
ex-pressed moderate to high levels of CXCR4, in comparison to
CCR5 expression, which was present at low levels on 0.1 to 2%
of the thymocyte population. Although we have shown that
expression of CXCR4 and CCR5 on thymocytes was necessary
for viral entry, additional host factors were required for a
highly productive infection in the CD69
1thymocyte subset.
This was evident in studies demonstrating that both the CD69
1and CD69
2cell populations allowed NL4-3 and JR-CSF entry,
whereas only the CD69
1population was identified as highly
susceptible to NL4-3 and JR-CSF productive infection.
CCR5 expression in fresh thymocytes, determined by both
surface and intracellular staining, was detected on few cells.
Underestimation of CCR5 expression could be occurring in
our system due to downregulation of CCR5 in thymocytes by
ligand occupation or virus binding. This is unlikely because low
levels of CCR5 mRNA were also detected by reverse
transcrip-tion-PCR (data not shown). In addition, Wu et al. reported
that 2D7 recognizes the chemokine binding site and does not
downregulate CCR5 expression (79). Furthermore, while low
levels of CCR5 could be detected on thymocytes with 2D7, this
antibody could block JR-CSF infection of thymocytes as
pre-viously reported for other cell types (60, 79, 80). JR-CSF usage
of alternative coreceptors on thymocytes cannot be excluded
by our studies (4, 54). However, an indirect effect of CCR5
blocking by 2D7 on such putative receptors affecting JR-CSF
and not NL4-3 replication would be necessary to explain our
data. For example, a link between mutations in CCR2 and the
level of expression of CCR5 has been proposed (56). However,
we favor the explanation that CCR5
2cells expressing HIV
originated as CCR5
1cells that have either internalized CCR5
due to virus binding or matured into CCR5
2cells.
In the postnatal thymus, CXCR4 was present at high levels
in immature CD1
1/CD3
1/lowthymocytes and at lower levels in
most but not all of the CD3
1/high/CD69
2/CD45RA
1thymo-cytes, cells that have the potential to leave the thymus (52, 76).
Our results further suggest that there are fewer
CCR5-express-FIG. 4. The antibody (2D7) to CCR5 is able to block productive infection of thymocytes by JR-CSF. Thymocytes were preincubated in the presence or ab-sence of antibody to CCR5 (2D7) or CXCR4 (12G5) for 2 h before infection with JR-CSF (30 IU/104cells). The antibodies were present during the infection and
throughout the culture with IL-2 plus IL-4. HIV replication was detected by measuring p24 antigen in the culture supernatants on days 8, 15, and 22 postin-fection. Preincubation conditions: no antibody (gray bars), 1mg of 2D7 (verti-cally striped bars), 5mg of 2D7 (diagonally striped bars), 10mg of 12G5 (hori-zontally striped bars), 1mg of 2D7 plus 10mg of 12G5 (checkered bars), 5mg of 2D7 plus 10mg of 12G5 (black bars), and 100mg of CD4-IgG (white bars).
on November 9, 2019 by guest
http://jvi.asm.org/
ing thymic emigrants than CXCR4-expressing thymic
emi-grants, which is consistent with reported studies demonstrating
low numbers of CCR5-expressing cells in the cord blood (48).
This finding is also in agreement with the fact that in adults,
CXCR4 expression in circulating T cells is detected mainly
in the naive CD26
low/CD45RA
1/CD45RO
2population, while
CCR5 is expressed mostly in the effector/memory CD26
high/
CD45RA
low/CD45R0
1population that has previously
under-gone activation (6, 80).
In PBMC, CXCR4 is upregulated within 72 h upon
stimu-FIG. 5. The antibody to CCR5 (2D7) specifically blocks expression of JR-CSF in thymocytes. Thymocytes were preincubated in the presence or absence of 5mg of CCR5 antibody per ml for 2 h before infection with JR-CSF (30 IU/104cells) or NL4-3 (1.5 IU/104cells). The antibody was present during infection and throughout
the culture with IL-2 plus IL-4. (A) At 2 weeks postinfection, cells were subjected to surface staining with CD3-PerCP followed by intracellular staining with KC57-FITC. (B) To determine the effect of CCR5 antibody on thymocytes, uninfected and infected cells were immunophenotyped with CD4-PE and CD8-PerCP and intracellularly stained with KC57-FITC 2 weeks postinfection. CD4-PE/CD8-PerCP expression is shown.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.76.518.68.613.2]FIG. 6. Infection of total thymocytes and CD691and CD692thymocyte sub-sets by JR-CSF and NL4-3. CD691and CD692subsets were obtained by using antibody-coated immunomagnetic beads from the CD45RA2thymocytes and immunophenotyped to determine the purity of the isolation. The resulting CD45RA2/CD691cells bound to beads, and CD45RA2/CD692cells were used for infection. (A) Immunophenotype of thymocytes before and after depletion of CD45RA- and CD69-expressing cells. (B) The total thymocyte population and the CD691and CD692thymocyte subsets were infected with JR-CSF (100 IU/104cells) or NL4-3 (10 IU/104cells) and cultured for 2 weeks in the presence
of IL-2 plus IL-4. Twenty-four hours postinfection 106cells were removed and
analyzed by using primers (R/U5) specific for the LTR region of HIV-1 to detect the presence of proviral DNA. To normalize for the amount of cellular DNA, PCR was performed in parallel for sequences in theb-globin gene. Heat-inactivated virus (2) were run in parallel with the live-virus-treated samples (1) as controls for DNA contamination from the inoculum. (C) PCR amplification of proviral DNA was performed on diluted DNA samples from the NL4-3-infected cells described above to detect the presence of fully reverse transcribed (RT) proviral DNA (LTR/gag) in parallel with the partially reverse transcribed (R/U5) proviral DNA. (D) HIV replication was detected by measuring p24 antigen in the culture supernatants of JR-CSF-infected CD691cells (vertically striped bars), JR-CSF-infected CD692 cells (white bars), NL4-3-infected CD691cells (black bars), and NL4-3-infected CD692cells (gray bars) on days 1, 5, and 12 postinfection.
on November 9, 2019 by guest
http://jvi.asm.org/
lation with PHA or anti-CD3, while increased CCR5
expres-sion on stimulated T cells requires addition of IL-2 for 2 to 3
weeks (6, 80). These culture conditions form the basis of the
slow/low versus rapid/high biological phenotype of CCR5 and
CXCR4 tropic primary isolates in PBMC (5). In both PBMC
and the SCID-hu mouse, the distribution of thymocyte
core-ceptors described in this study is a major determinant of the
biological phenotype of NL4-3 and JR-CSF (27, 28, 64, 71, 74).
The expression of CXCR4 on the immature CD3
2/CD4
1/low/
CD8
1thymocytes may lead to a rapid productive infection and
destruction of this actively proliferating cell population. We
have found that cultures containing IL-4 increased the level of
CXCR4 expression in the mature CD3
1/highthymocyte subset,
thereby increasing the number of NL4-3 targets. The high
lev-els of CXCR4 expression in freshly isolated immature
thymo-cytes, detected in all specimens analyzed, may be related to the
presence of IL-4 in the subcortical area where immature
thy-mocytes responding to IL-4 are found (22, 75). Consistent with
this notion, Papiernik et al. reported that pathological
abnor-malities in fetuses aborted from HIV-1-seropositive women
were present mainly in the cortex (47). Our observations
fur-ther suggest that immature thymocyte subsets from children
may be infected in vivo with CXCR4-tropic HIV isolates, as
observed in the SCID-hu model (27). Confirmation of a similar
effect of IL-4 on upregulation of CXCR4 expression in the
periphery might signify that the proposed shift from a Th1 to
Th2 pattern of cytokine synthesis could favor the propagation
of CXCR4-tropic viruses in late stages of diseases (11).
Fur-thermore, a Th2-like cytokine pattern has been observed in
perinatally infected children progressing to AIDS (26).
Increased CCR5 expression in thymocytes was observed only
in cultures containing IL-2 in combination with IL-4. As seen
in stimulated PBMC, upregulation of CCR5 expression in
thy-mocytes required the presence of IL-2 for at least 2 weeks (6,
80). The slower replication of JR-CSF in thymocytes was
ini-tially due to low availability of CCR5 and was reflected in the
low levels of viral entry detected by PCR. The increase in
JR-CSF production seen in IL-2 plus IL-4-supplemented cultures
was presumably from upregulation of CCR5 on mature
thy-mocytes and proliferation of these cells, thereby allowing viral
spread. It is noteworthy that high levels of virus could be
produced by very few infected cells, suggesting that a mature
thymocyte population expressing CCR5 is highly permissive to
JR-CSF replication.
We have found that both NL4-3 and JR-CSF replicate
pref-erentially in the CD69
1thymocyte population. This
popula-tion includes cells at various stages of maturapopula-tion from the less
mature CD1
1/CD4
1/CD8
1cells through the single-positive
CD4
1or CD8
1populations (52, 76). Since JR-CSF is not
produced in immature CD1
1cells (71), we conclude that the
thymocyte subset producing high levels of JR-CSF is a mature
subset that has downregulated CD1, but not yet CD69, and
therefore is not ready to leave the thymus. In this CD1
2/
CD69
1subset, NL4-3 production is also highly favored, but
the broad distribution of CXCR4 expression allows NL4-3
entry into the immature CD1
1/CD69
2populations, thereby
accounting for the low level of NL4-3 production seen in the
immature thymocyte subset. Detection of full-length proviral
DNA in all populations confirms that while coreceptor
expres-sion is a major determinant of tropism, cellular factors
ex-pressed at specific stages of T-cell development affect
post-entry events and can determine HIV replication in the thymus.
In this regard, it should be noted that in vivo, the CD69
1pop-ulation consists of thymocytes that are activated during the
process of positive selection (43, 76) and thus should be
per-missive for viral entry and replication.
We have previously proposed that pediatric isolates able to
infect immature thymocytes might have a greater impact on
disease progression (71). Here we show that a CXCR4-tropic
isolate could produce this effect. We are now in the process of
determining whether coreceptor use of isolates obtained from
children with rapid and slow disease progression correlates
with specific receptor use and subsequent loss of thymocytes.
In this regard, the early acquisition of CXCR4 tropism in rapid
progressors observed by Scarlatti et al. could be associated with
CXCR4 targeting in the thymus (56).
It has been proposed that differences in the expression levels
of CCR5 due to genetic factors can affect the rate of disease
progression in adults and children, where heterozygosity for
the CCR5
D
32 deletion substantially reduces disease
progres-sion (42, 61). It is clear that in our in vitro conditions, at a low
MOI, the threshold of CCR5 expression required for
replica-tion in thymocytes is very low. Although it takes longer, CD4
depletion occurs in SCID-hu mice infected with JR-CSF (27).
Since in our system the contribution of stromal elements
(po-tentially CCR5 positive) could not be evaluated, we cannot
de-termine the full contribution of CCR5 for HIV pathogenesis
on the thymus. Stanley et al. have shown that JR-CSF causes a
more pronounced disruption of stromal elements than a
T-tropic virus (64). The usage of coreceptors other than CCR5
and CXCR4 by pediatric isolates in the thymus needs to be
investigated.
In conclusion, our studies indicate that the ability of
thymo-cyte subsets to support HIV productive infection is determined
by the presence of the appropriate coreceptor and by cellular
factors related to the state of maturation of the cells that affect
postentry events in the virus replication cycle.
ACKNOWLEDGMENTS
The first two authors contributed equally to this work.
This work was supported by grants from the National Institutes of
Health (HD 29341, HD 29341-S1, AI 28697, and DK49886), by UARP
SRF01, and by student awards to K.B.G. from the Elizabeth Glaser
Pediatric AIDS Foundation and the UCLA AIDS Institute (Esther
Hays Graduate Student Award).
We thank Hillel Laks and his colleagues and staff for providing the
thymus specimens; Jerome Zack and Irvin Chen for use of
biocontain-ment facilities; Esther Hays, Beth Jamieson, John Ferbas, and
Debo-rah Anisman-Posner for helpful discussions and critical reviews of the
manuscript; and Deborah Anisman-Posner, Silvia Neagos, Kris
Con-ners, and Prista Charuworn for excellent technical assistance.
REFERENCES
1. Adachi, A., H. E. Gendelman, S. Koenig, T. Folks, R. Willey, A. Rabson, and
M. A. Martin.1986. Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhuman cells transfected with an infectious molecular clone. J. Virol. 59:284–291.
2. Aldrovandi, G. M., G. Feuer, L. Gao, B. Jamieson, M. Kristeva, I. S. Y. Chen,
and J. A. Zack.1993. The SCID-hu mouse as a model for HIV-1 infection. Nature 363:732–736.
3. Alkhatib, G., C. Combardiere, C. C. Broder, Y. Feng, P. E. Kennedy, P. M.
Murphy, and E. A. Berger.1996. CC CKR5: a RANTES, MIP-1a, MIP-1b receptor as a fusion cofactor for macrophage-tropic HIV-1. Science 272: 1955–1958.
4. Bazan, H. A., G. Alkhatib, C. C. Broder, and E. A. Berger. 1998. Patterns of CCR5, CXCR4, and CCR3 usage by envelope glycoproteins from human immunodeficiency virus type 1 primary isolates. J. Virol. 72:4485–4491. 5. Bjorndal, A., H. Deng, M. Jansson, J. R. Fiore, C. Colognesi, A. Karlsson, J.
Albert, G. Scarlatti, D. R. Littman, and E. M. Fenyo.1997. Coreceptor usage of primary human immunodeficiency virus type 1 isolates varies according to biological phenotype. J. Virol. 71:7478–7487.
6. Bleul, C. C., L. Wu, J. A. Hoxie, T. A. Springer, and C. R. Mackay. 1997. The HIV coreceptors CXCR4 and CCR5 are differentially expressed and regu-lated on human T lymphocytes. Proc. Natl. Acad. Sci. USA 94:1925–1930. 7. Blue, M., H. Levine, J. F. Daley, K. R. Branton, and S. F. Schlossman. 1989.
Expression of CD1 and class I MHC antigens by human thymocytes. J. Im-munol. 142:2714–2720.
on November 9, 2019 by guest
http://jvi.asm.org/
8. Calabro, M. L., C. Zanotto, F. Calderazzo, C. Crivellaro, A. Del Mistro, A.
De Rossi, and L. Chieco-Bianchi. 1995. HIV-1 infection of the thymus: evidence for a cytopathic and thymotropic viral variant in vivo. AIDS Res. Hum. Retroviruses 11:11–19.
9. Carroll, R. G., J. L. Riley, B. L. Levine, Y. Feng, S. Kaushal, D. W. Ritchey,
W. Bernstein, O. S. Weislow, C. R. Brown, E. A. Berger, C. H. June, and D. C. St. Louis.1997. Differential regulation of HIV-1 fusion cofactor expression by CD28 costimulation of CD41T cells. Science 276:273–276.
10. Chassagne, J., P. Verrelle, C. Dionet, F. Clavel, F. Barre-Sinoussi, J. C.
Chermann, L. Montagnier, J. C. Gluckmann, and D. Klatzmann.1986. A monoclonal antibody against LAV gag precursor: use for viral protein anal-ysis and antigenic expression in infected cells. J. Immunol. 136:1442–1445. 11. Clerici, M., and G. M. Shearer. 1994. The Th1-Th2 hypothesis of HIV
infection: new insights. Immunol. Today 15:575–581.
12. Connor, R. I., K. E. Sheridan, D. Ceradini, S. Choe, and N. R. Landau. 1997. Change in coreceptor use correlates with disease progression in HIV-1-infected individuals. J. Exp. Med. 185:621–628.
13. Deng, H., R. Liu, W. Ellmeier, S. Choe, D. Unutmaz, M. Burkhart, P.
DiMarzio, S. Marmon, R. E. Sutton, C. M. Hill, C. B. Davis, S. C. Peiper, T. J. Schall, D. Littman, and N. R. Landau.1996. Identification of a major co-receptor for primary isolates of HIV-1. Nature 381:661–666.
14. Doranz, B. J., J. Rucker, Y. Yi, R. J. Smyth, M. Samson, S. C. Peiper, M.
Parmentier, R. G. Collman, and R. W. Doms.1996. A dual-tropic primary HIV-1 isolate that uses fusin and theb-chemokine receptors CKR-5, CKR-3, and CKR-2b as fusion cofactors. Cell 85:1149–1158.
15. Dragic, T., V. Litwin, G. P. Allaway, S. R. Martin, Y. Huang, K. A.
Naga-shima, C. Cayanan, P. J. Maddon, R. A. Koup, J. P. Moore, and W. A. Paxton.1996. HIV-1 entry into CD41cells is mediated by the chemokine receptor CC-CKR5. Nature 381:667–672.
16. Endres, M. J., P. R. Clapham, M. Marsh, M. Ahuja, J. Davis Turner, A.
McKnight, J. F. Thomas, B. Stoebenau-Haggarty, S. Choe, P. J. Vance, T. N. C. Wells, C. A. Power, S. S. Sutterwala, R. W. Doms, N. R. Landau, and J. A. Hoxie.1996. CD4-independent infection by HIV-2 is mediated by fusin/CXCR4. Cell 87:745–756.
17. Feng, Y., C. C. Broder, P. E. Kennedy, and E. A. Berger. 1996. HIV-1 entry cofactor: functional cDNA cloning of a seven-transmembrane, G protein-coupled receptor. Science 272:872–877.
18. Gaulton, G. N., J. V. Scobie, and M. Rosenzweig. 1997. HIV-1 and the thymus. AIDS 11:403–414.
19. Godiska, R., D. Chantry, C. J. Raport, S. Sozzani, P. Allavena, D. Leviten, A.
Mantovani, and P. W. Gray.1997. Human macrophage-derived chemokine (MDC), a novel chemoattractant for monocytes, monocyte-derived dendritic cells, and natural killer cells. J. Exp. Med. 185:1595–1604.
20. Hays, E. F., C. H. Uittenbogaart, L. W. Vollger, J. Brewer, and J. A. Zack. 1992. In vitro studies of HIV-1 expression in thymocytes from infants and children. AIDS 6:265–272.
21. He, J., Y. Chen, M. Farzan, H. Choe, A. Ohagen, S. Gartner, J. Busciglio, X.
Yang, W. Hofmann, W. Newman, C. R. Mackay, J. Sodrovski, and D. Gabuzda.1997. CCR3 and CCR5 are co-receptors for HIV-1 infection of microglia. Nature 385:645–649.
22. He, W., Y. Zhang, Y. Deng, and D. Kabelitz. 1995. Induction of TCR-gd expression on triple-negative (CD3-CD4-CD8-) human thymocytes. J. Im-munol. 154:3726–3731.
23. Heath, H., S. Qin, P. Rao, L. Wu, G. LaRosa, N. Kassam, P. D. Ponath, and
C. R. Mackay.1997. Chemokine receptor usage by eosinophils. The impor-tance of CCR3 demonstrated using an antagonistic monoclonal antibody. J. Clin. Investig. 99:178–184.
24. Hieshima, K., T. Imai, M. Baba, K. Shoudai, K. Ishizuka, T. Nakagawa,
J. Tsuruta, M. Takeya, Y. Sakaki, K. Takatsuki, R. Miura, G. Opdenakker, J. Van Damme, O. Yoshie, and H. Nomiyama.1997. A novel human CC chemokine PARC that is most homologous to macrophage-inflammatory protein-1a/LD78a and chemotactic for T lymphocytes, but not for mono-cytes. J. Immunol. 159:1140–1149.
25. Huang, Y., W. A. Paxton, S. M. Wolinsky, A. U. Neumann, L. Zhang, T. He,
S. Kang, D. Ceradini, Z. Jin, K. Yazdanbakhsh, K. Kunstman, D. Erickson, E. Dragon, N. R. Landau, J. Phair, D. D. Ho, and R. A. Koup.1996. The role of a mutant CCR5 allele in HIV-1 transmission and disease progression. Nat. Med. 2:1240–1243.
26. Hyjek, E., H. W. Lischner, T. Hyslop, J. Bartkowiak, M. Kubin, G.
Trinch-ieri, and D. Kozbor.1995. Cytokine patterns during progression to AIDS in children with perinatal HIV infection. J. Immunol. 155:4060–4071. 27. Jamieson, B. D., S. Pang, G. M. Aldrovandi, J. Zha, and J. A. Zack. 1995. In
vivo pathogenic properties of two clonal human immunodeficiency virus type 1 isolates. J. Virol. 69:6259–6264.
28. Jamieson, B. D., C. H. Uittenbogaart, I. Schmid, and J. A. Zack. 1997. High viral burden and rapid CD41cell depletion in human immunodeficiency virus type 1-infected SCID-hu mice suggests direct viral killing of thymocytes in vivo. J. Virol. 71:8245–8253.
29. Joshi, V. V., J. M. Oleske, S. Saad, C. Gadol, E. Connor, R. Bobila, and A. B.
Minnefor.1986. Thymus biopsy in children with acquired immunodeficiency syndrome. Arch. Pathol. Lab. Med. 110:837–842.
30. Kitchen, S. G., C. H. Uittenbogaart, and J. A. Zack. 1997. Mechanism of
human immunodeficiency virus type 1 localization in CD4-negative thymo-cytes: differentiation from a CD4-positive precursor allows productive infec-tion. J. Virol. 71:5713–5722.
31. Kitchen, S. G., and J. A. Zack. 1997. CXCR4 expression during lymphopoi-esis: implications for human immunodeficiency virus type 1 infection of the thymus. J. Virol. 71:6928–6934.
32. Korin, Y. D., and J. A. Zack. 1998. Progression to the G1b phase of the cell
cycle is required for completion of human immunodeficiency virus type 1 reverse transcription in T cells. J. Virol. 72:3161–3168.
33. Koyanagi, Y., S. Miles, R. T. Mitsuyasu, J. E. Merill, H. V. Vinters, and
I. S. Y. Chen.1987. Dual infection of the central nervous system by AIDS viruses with distinct cellular tropisms. Science 236:819–822.
34. Kozak, S. L., E. J. Platt, N. Madani, Ferro, Jr., K. Peden, and D. Kabat. 1997. CD4, CXCR-4 and CCR-5 dependencies for infections by primary patient and laboratory-adapted isolates of human immunodeficiency virus type 1. J. Virol. 71:873–882.
35. Lawn, R. M., A. Efstratiadus, C. O’Connell, and T. Maniatis. 1980. The nucleotide sequence of the humanb-globin gene. Cell 21:647–651. 36. Liao, F., G. Alkhatib, K. W. C. Peden, G. Sharma, E. A. Berger, and J. M.
Farber.1997. STRL33, a novel chemokine receptor-like protein, functions as a fusion cofactor for both macrophage-tropic and T cell line-tropic HIV-1. J. Exp. Med. 185:2015–2023.
37. Liu, R., W. A. Paxton, S. Choe, D. Ceradini, S. R. Martin, R. Horuk, M. E.
MacDonald, H. Stuhlmann, R. A. Koup, and N. R. Landau.1996. Homozy-gous defect in HIV-1 coreceptor accounts for resistance of some multiply-exposed individuals to HIV-1 infection. Cell 86:367–377.
38. Loetscher, P., M. Seitz, M. Baggliolini, and B. Moser. 1996. Interleukin-2 regulates CC chemokine receptor expression and chemotactic responsive-ness in T lymphocytes. J. Exp. Med. 184:569–577.
39. Mackall, C. L., T. A. Fleisher, M. R. Brown, M. P. Andrich, C. C. Chen, I. M.
Feuerstein, M. E. Horowitz, I. T. Magrath, A. T. Shad, S. M. Steinberg, L. H. Wexler, and R. E. Gress.1995. Age, thymopoiesis, and CD41T-lymphocyte regeneration after intensive chemotherapy. N. Engl. J. Med. 332:143–149. 40. Martin, S. J., P. M. Matear, and A. Vyakarnam. 1994. HIV-1 infection of
human CD41T cells in vitro. J. Immunol. 152:330–342.
41. McKnight, A., D. Wilkinson, G. Simmons, S. Talbot, L. Picard, M. Ahuja, M.
Marsh, J. A. Hoxie, and P. R. Clapham.1997. Inhibition of human immu-nodeficiency virus fusion by monoclonal antibody to a coreceptor (CXCR4) is both cell type and virus strain dependent. J. Virol. 71:1692–1696. 42. Misrahi, M., J. Teglas, N. N’Go, M. Burgard, M. Mayaux, C. Rouzioux, J.
Delfraissy, and S. Blanche. 1998. CCR5 chemokine receptor variant in HIV-1 mother-to-child transmission and disease progression in children. JAMA 279:277–280.
43. Moore, N. C., J. Girdlestone, G. Anderson, J. J. T. Owen, and E. J.
Jenkin-son. 1995. Stimulation of thymocytes before and after positive selection results in the induction of different NF-kB/Rel protein complexes. J. Immu-nol. 155:4653–4660.
44. Moriuchi, H., M. Moriuchi, and A. S. Fauci. 1997. Cloning and analysis of the promoter region of CCR5, a coreceptor for HIV-1 entry. J. Immunol.
159:5441–5449.
45. Moriuchi, M., H. Moriuchi, W. Turner, and A. S. Fauci. 1997. Cloning and analysis of the promoter region of CXCR4, a coreceptor for HIV-1 entry. J. Immunology 159:4322–4329.
46. Napolitano, M., A. Zingoni, G. Bernardini, G. Spinetti, A. Nista, C. T.
Starlazzi, M. Rocchi, and A. Santoni.1996. Molecular cloning of TER-1, a chemokine receptor-like gene expressed by lymphoid tissues. J. Immunol.
157:2759–2763.
47. Papiernik, M., Y. Brossard, N. Milliez, J. Roume, C. Brechot, F. Barin, A.
Goudeau, J.-F. Bach, C. Griscelli, R. Henrion, and R. Vazeux.1992. Thymic abnormalities in fetuses aborted from human immunodeficiency virus type 1 seropositive women. Pediatrics 89:297–301.
48. Peng-Yang, L., J. L. Riley, R. G. Carroll, C. H. June, J. Hoxie, B. K.
Patterson, Y. Ohshima, R. J. Hodes, and G. Delespesse.1998. Productive infection of neonatal CD81T lymphocytes by HIV-1. J. Exp. Med. 187:1139– 1144.
49. Power, C. A., A. Meyer, K. Nemeth, K. B. Bacon, A. J. Hoogewerf, A. E.
Proudfoot, and T. N. C. Wells. 1995. Molecular cloning and functional expression of a novel CC chemokine receptor cDNA from a human baso-philic cell line. J. Biol. Chem. 270:19495–19500.
50. Prevot, S., J. Audouin, J. Andre-Bougaran, R. Griffais, A. Le Tourneau, J. G.
Fournier, and J. Diebold.1992. Thymic pseudotumorous enlargement due to follicular hyperplasia in a human immunodeficiency virus sero-positive pa-tient. Am. J. Clin. Pathol. 97:420–425.
51. Raport, C. J., J. Goslings, V. L. Schweickart, P. W. Gray, and I. F. Charo. 1996. Molecular cloning and functional characterization of a novel human CC chemokine receptor (CCR5) for RANTES, MIP-1band MIP-1a. J. Biol. Chem. 271:17161–17166.
52. Res, P., B. Blom, T. Hori, K. Weijer, and H. Spits. 1997. Downregulation of CD1 marks acquisition of functional maturation of human thymocytes and defines a control point in late stages of human T cell development. J. Exp. Med. 185:141–151.
53. Rosenzweig, M., D. P. Clark, and G. N. Gaulton. 1993. Selective thymocyte
on November 9, 2019 by guest
http://jvi.asm.org/
depletion in neonatal HIV-1 thymic infection. AIDS 7:1601–1605. 54. Rucker, J., A. L. Edinger, M. Sharron, M. Samson, B. Lee, J. F. Berson, Y.
Yi, B. Margulies, R. G. Collman, B. J. Doranz, M. Parmentier, and R. W. Doms.1997. Utilization of chemokine receptors, orphan receptors, and her-pesvirus-encoded receptors by diverse human and simian immunodeficiency viruses. J. Virol. 71:8999–9007.
55. Samson, M., F. Libert, B. J. Doranz, J. Rucker, C. Liesnard, C. Farber, S.
Saragosti, C. Lapoumeroulie, J. Cognaux, C. Forceille, G. Muyldermans, C. Verhofstede, G. Burtonboy, M. Georges, T. Imai, S. Rana, Y. Yi, R. J. Smyth, R. G. Collman, R. W. Doms, G. Vassart, and M. Parmentier.1996. Resis-tance to HIV-1 infection in caucasian individuals bearing mutant alleles of the CCR5 chemokine receptor gene. Nature 382:722–725.
56. Scarlatti, G., E. Tresoldi, A. Bjorndal, R. Frederiksson, C. Colognesi, H. K.
Deng, M. S. Malnati, A. Plebani, A. G. Siccardi, D. R. Littman, E. M. Fenyo, and P. Lusso. 1997. In vivo evolution of HIV-1 co-receptor usage and sensitivity to chemokine-mediated suppression. Nat. Med. 3:1259–1265. 57. Schmid, I., W. J. Krall, C. H. Uittenbogaart, J. Braun, and J. V. Giorgi. 1992.
Dead cell discrimination with 7-amino-actinomycin D in combination with dual color immunofluorescence in single laser flow cytometry. Cytometry 13: 204–208.
58. Schmid, I., C. H. Uittenbogaart, and J. V. Giorgi. 1991. A gentle fixation and permeabilization method for combined cell surface and intracellular staining with improved precision in DNA quantification. Cytometry 12:279–285. 59. Schmid, I., C. H. Uittenbogaart, B. Keld, and J. V. Giorgi. 1994. A rapid
method for measuring apoptosis and dual-color immunofluorescence by single laser flow cytometry. J. Immunol. Methods 170:145–157.
60. Simmons, G., D. Wilkinson, J. D. Reeves, M. T. Dittmar, S. Beddows, J.
Weber, G. Carnegie, U. Desselberger, P. W. Gray, R. A. Weiss, and P. R. Clapham.1996. Primary, syncytium-inducing human immunodeficiency virus type 1 isolates are dual-tropic and most can use either Lestr or CCR5 as coreceptors for virus entry. J. Virol. 70:8355–8360.
61. Smith, M. W., M. Dean, M. Carrington, C. Winkler, G. A. Huttley, D. A.
Lomb, J. J. Goedert, T. R. O’Brien, L. P. Jacobson, R. Kaslow, S. Buch-binder, E. Vittinghoff, D. Vlahov, K. Hoots, and M. W. Hilgartner.1997. Contrasting genetic influence of CCR2 and CCR5 variants on HIV-1 infec-tion and disease progression. Science 277:959–965.
62. Sozzani, S., S. Ghezzi, G. Iannolo, W. Luini, A. Borsatti, N. Polentarutti, A.
Sica, M. Locati, C. Mackay, T. N. C. Wells, P. Biswas, E. Vicennzi, G. Poli, and A. Mantovani.1998. Interleukin 10 increases CCR5 expression and HIV infection in human monocytes. J. Exp. Med. 187:439–444.
63. Spina, C. A., J. C. Guatelli, and D. D. Richman. 1995. Establishment of a stable, inducible form of human immunodeficiency virus type 1 DNA in quiescent CD4 lymphocytes in vitro. J. Virol. 69:2977–2988.
64. Stanley, S. K., J. M. McCune, H. Kaneshima, J. S. Justement, M. Sullivan,
E. Boone, M. Baseler, J. Adelsberger, M. Bonyhadi, J. Orenstein, C. H. Fox, and A. S. Fauci.1993. Human immunodeficiency virus infection of the hu-man thymus and disruption of the thymic microenvironment in the SCID-hu mouse. J. Exp. Med. 178:1151–1163.
65. Stevenson, M., T. L. Stanwick, M. P. Dempsey, and C. A. Lamonica. 1990. HIV-1 replication is controlled at the level of T cell activation and proviral integration. EMBO J. 9:1551–1560.
66. Tedder, T. F., L. C. Clement, and M. D. Cooper. 1985. Human lymphocyte differentiation antigens HB-10 and HB-11. I. Ontogeny of antigen expres-sion. J. Immunol. 134:2983–2988.
67. Testi, R., J. H. Phillips, and L. L. Lanier. 1988. Constitutive expression of a phosphorylated activation antigen (Leu 23) by CD3brighthuman thymocytes.
J. Immunol. 141:2557–2563.
68. Tiffany, H. L., L. L. Lautens, J. Gao, J. Pease, M. Locati, C. Combadiere, W.
Modi, T. I. Bonner, and P. M. Murphy.1997. Identification of CCR8: a human monocyte and thymus receptor for the CC chemokine I-309. J. Exp. Med. 186:165–170.
69. Ueno, Y., T. Boone, and C. H. Uittenbogaart. 1989. Selective stimulation of human thymocyte subpopulations by recombinant IL-4 and IL-3. Cell. Im-munol. 118:382–393.
70. Ueno, Y., E. Hays, L. Hultin, and C. H. Uittenbogaart. 1989. Human thy-mocytes do not respond to interleukin-2 after removal of mature “bright” CD5 positive cells. Cell. Immunol. 124:239–251.
71. Uittenbogaart, C. H., D. J. Anisman, B. D. Jamieson, S. Kitchen, I. Schmid,
J. A. Zack, and E. F. Hays.1996. Differential tropism of HIV-1 isolates for distinct thymocyte subsets in vitro. AIDS 10:F9–F16.
72. Uittenbogaart, C. H., D. J. Anisman, J. A. Zack, A. Economides, I. Schmid,
and E. F. Hays.1995. Effects of cytokines on HIV-1 production by thymo-cytes. Thymus 23:155–175.
73. Uittenbogaart, C. H., S. Higashitani, I. Schmid, L. W. Vollger, T. Boone, and
L. T. Clement.1990. Interleukin-4 induces expression of the CD45RA anti-gen on human thymocyte subpopulations. Int. Immunol. 2:1179–1187. 73a.Uittenbogaart, C. H., et al. Unpublished data.
74. Valentin, H., M.-T. Nugeyre, F. Vuillier, L. Boumsell, M. Schmid, F.
Barre-Sinoussi, and R. A. Pereira.1994. Two subpopulations of human triple-negative thymic cells are susceptible to infection by human immunodefi-ciency virus type 1 in vitro. J. Virol. 68:3041–3050.
75. Vandekerckhove, B. A. E., A. Barcena, D. Schols, S. Mohan-Peterson, H.
Spits, and M. Roncarolo.1994. In vivo cytokine expression in the thymus. J. Immunol. 152:1738–1743.
76. Vanhecke, D., G. Leclercq, J. Plum, and B. Vandekerckhove. 1995. Charac-terization of distinct stages during the differentiation of human CD691CD31 thymocytes and identification of thymic emigrants. J. Immunol. 155:1862– 1872.
77. Vicari, A. P., D. J. Figueroa, J. A. Hedrick, J. S. Foster, K. P. Singh, S.
Menon, N. G. Copeland, D. J. Gilbert, N. A. Jenkins, K. B. Bacon, and A. Zlotnik.1997. TECK: a novel CC chemokine specifically expressed by thymic dendritic cells and potentially involved in T cell development. Immunity 7: 291–301.
78. Vollger, L. W., and C. H. Uittenbogaart. 1993. Interleukin-7 promotes the generation of phenotypically mature CD45RA positive human thymocytes in-vitro. Cytokine 5:157–168.
79. Wu, L., G. LaRosa, N. Kassam, C. J. Gordon, H. Heath, N. Ruffing, N. Chen,
J. Humblias, M. Samson, M. Parmentier, J. P. Moore, and C. R. Mackay.
1997. Interaction of chemokine receptor CCR5 with its ligands: multiple domains for HIV-1 gp120 binding and a single domain for chemokine bind-ing. J. Exp. Med. 186:1373–1381.
80. Wu, L., W. A. Paxton, N. Kassam, N. Ruffing, J. B. Rottman, N. Sullivan, H.
Choe, J. Sodrovski, W. Newman, R. A. Koup, and C. R. Mackay.1997. CCR5 levels and expression pattern correlate with infectability by macrophage-tropic HIV-1, in vitro.