0022-538X/90/010061-08$02.00/0
Copyright©D 1990, American Societyfor Microbiology
Regulation of pp60Ocsrc and Its Interaction with Polyomavirus
Middle T Antigen in Insect Cells
HELEN PIWNICA
WORMS,lt*
NIDHI G. WILLIAMS,' SENG H. CHENG,2ANDTHOMAS M. ROBERTS'Dana-Farber CancerInstituteandDepartmentofPathology, HarvardMedical School, Boston, Massachusetts02115J, andLaboratory ofCellular Regulation, IntegratedGeneticsInc., Framingham,Massachusetts 017012
Received21June1989/Accepted15September 1989
High yields of soluble, biologically active pp60csrc and middle t antigen (MTAg) of polyomavirus were
produced in insect cells, using a baculovirus expression system. In mammalian cells, pp60csrc undergoes a
regulatory phosphorylationonTyr-527 in vivo and is autophosphorylatedonTyr-416in vitro. In insect cells, pp60C-srC was phosphorylated primarily on Tyr-416, although Tyr-527 was detectable at a low level. A
kinase-negativemutantofpp60C`c? was notphosphorylatedoneither Tyr-527or Tyr-416in insect cells and
thus isanexcellentbiochemicalreagent tosearchfor the regulatory kinase thatusuallyphosphorylates Tyr-527 inmammalian cells.MTAg synthesized in insectcells wasnotphosphorylatedontyrosine residues in vivoor
invitro, suggesting that it didnotassociate withany endogenoustyrosinekinases. However, MTAg isolated
fromcellscoinfected with viruses encoding both MTAg andpp60's'Cwasphosphorylatedontyrosine residues
both in vivo and in vitro.
pp60-csrc is the cellular homolog of the retroviral trans-forming protein
pp60v-src.
pp60c-src andpp60v srcare struc-turally andfunctionally related. Bothareassociatedwith theplasmamembrane because ofan amino-terminal
myristyla-tion (3, 44). Bothpossessintrinsic tyrosine kinase activities
(12, 23, 32),although the kinase activity of pp60v-src has been estimated to be 10- to50-fold higher than that ofpp60c-src when measured in vitro (21, 24). Both proteins are
phos-phoproteins, although their sitesof tyrosine phosphorylation differ invivo. pp60`csrc is phosphorylated onSer-17 (11, 22)
and Tyr-527 (13) in vivo. Tyr-527 is located in the unique carboxyl terminus of pp60j-src and, therefore, is absent in pp6O -src. Data from a number of laboratories support the hypothesis that phosphorylation of Tyr-527 suppresses the
kinaseactivity and transforming potential of pp60-src in vivo (5, 6, 14, 16, 17, 24, 28, 29, 40). However, the kinase responsible for this negative regulation hasnotbeen identi-fied. Tyr-416 is the site of phosphorylation in vivo in (i) pp60v-src(37,46);(ii) transformingmutantsofpp60c`src (5, 9, 24, 28, 40), and (iii) that fraction of pp6Oc-src associated with the middle t antigen (MTAg) of polyomavirus (6). In each
case, phosphorylation of Tyr-416 correlates withenhanced kinaseactivity. Thus, full activation of pp6Oc-srcmayrequire
concomitantchanges inphosphorylationatbothTyr-527 and Tyr-416.
MTAg is the transforming protein of polyomavirus(47). It has no known intrinsic catalytic activities; instead, it is
thought to function by interacting with and altering the activitiesof cellularproteins. Three tyrosine kinases known tointeract withMTAgarepp60c-src (19), pp62c-Yes (30),and
pp59c-fyf
(7, 31). MTAg has been shown to activate the kinase activity of pp6Oc-src and pp62c-Yes (2, 30). Genetic studies have demonstratedthataninteraction withpp60csrcis necessary for MTAg-mediated transformation (8, 20). How MTAg altersthe specifickinaseactivityofpp60f-src is unclear. The shift inphosphorylation fromTyr-527 to Tyr-416doesnot accountforthe entire activation(5),Complexed
*Correspondingauthor.
tPresent address: Department of Physiology, Tufts Medical
School, 136HarrisonAve., Boston,MA 02111.
pp60-src has also been showntobephosphorylatedon one or moreamino-terminal tyrosine residues (50). These
phos-phorylations have been detected in vivo only in thepresence
of sodium orthovanadate or in vitro in immune complex
kinase assays. Thus, their biological significance remains
unresolved. Inaddition, onlyasmallproportion of the total
amount ofMTAg and pp60csrc presentwithin acell atany
given time is found stably associated (1, 20). Levels of the complexcanbeincreasedonly marginally by overexpressing either pp6Ocsrc or MTAg (39, 41, 43) in mammalian cells.
Factors controlling the association of these two molecules
are presently not known. Unfortunately, the two proteins fail to interact when they are coproduced in bacteria (H.
Piwnica-Worms, D. Pallas, and T. M. Roberts, unpublished data) or in yeast cells (S. Kombluth and H. Hanafusa, personal communication) orwhen they are cotranslated in
vitro inarabbitreticulocytelysate (H. Piwnica-Worms, and
T. M. Roberts, unpublished data). A complete biochemical analysis awaits the development of a system in which
purified components can be added and theireffectscan be
assessedindividually in vitro.
Asa first step towards the development ofan assay, we
have overproduced MTAg and avian
pp60,csrc,
using abaculovirusexpression system.We haveanalyzedthe post-translationalmodifications undergone by pp60csrc in insect cells, including myristylationandphosphorylation. In addi-tion, by coinfectingcells with recombinant virusesencoding
bothpp60"srcandMTAg,wehaveobtainedpp60c-src-MTAg
complexes.
MATERIALS AND METHODS
Celllines.Spodopterafrugiperda (Sf9)cellswereobtained from the AmericanTypeCulture Collection.Allprocedures relating to viral propagation, isolation, and plaquing were
performed assuggested by Summersand Smith (49). Generationofc-srcbaculoviruses. pSP68-4(RBS) (39) was
restrictedwithHindIII, andaBamHI linkerwasinsertedto generate pSP68-4RBS(Bam). pSP68-4RBS(Bam) was
re-stricted with NcoI, and a BclI linker (39) was inserted to generate pSP68-4RBS(Bcl-Bam). pSP68-4RBS(Bcl-Bam)
was restricted with BclI and BamHI, and the fragment 61
on November 10, 2019 by guest
http://jvi.asm.org/
encoding pp60csrc was isolated. This c-src fragment was cloned into BamHI-restricted and phosphatased pAC373 (49) to generate pAC373(c-src). A mixture of 2 jig of
pAC373(c-src)and 1jig of purifiedwild-type viral DNA was
transfectedintoSf9 cells.Four days later, medium
superna-tants wereremovedandcentrifugedat 1,000rpm for 10 min.
Clarifiedsupernatantscontainingwild-type and recombinant
viruseswere plaqued on amonolayer of Sf9 cells. Occlusion
negative plaques were picked and seeded onto 2.5 x 106
cells. Three days later, cells and mediumsupernatants were removed and centrifuged at 1,000 rpm for 10 min. The supernatant containing the virus was stored at 4°C. The
pelletedcells were lysedandimmunoprecipitated with EC10
serum (a monoclonal antibody specific for avian pp60csrc
[36]),andkinase assays wereperformedinvitro as described
previously (39). All lysates tested positive for the presence
of kinase-active pp6O-src. One supernatantwasselected and
taken through three rounds of plaque purification. A c-src
mutant encoding methionine rather than lysine at position 295 was constructed by site-directed mutagenesis as de-scribedpreviously (40). The Lys-295 to Met mutant was then
cloned intopAC373 togenerate pAC373(295) and
recombi-nantviruses were generated asdescribed above.
Generation of MTAg baculovirus. p890 (26)was restricted
withBclI andBamHI,and the fragmentencodingMTAg was
cloned intopAC373 which had been linearizedwith BamHI
and phosphatased. Therecombinantplasmidwasdesignated
pAC373(MTAg). A mixture of pAC373(MTAg) and
wild-type viral DNAwas transfected into Sf9 cells asdescribed
above for pp60-src.Twentyocclusion-negativeplaques were
picked and seeded onto cells as described above. After 6
days, cells and medium supernatants were removed and
clarified. Viral supematants were stored at4°C. Cellswere
lysedand normalized for protein content with the Bio-Rad
Laboratories protein assay kit, and proteinswere resolved
by 10% sodium dodecyl sulfate-polyacrylamide gel
electro-1
2
3
4
5
6
7
__
~~-pp60
phoresis (SDS-PAGE). Proteins were blotted onto nitrocel-lulose and probed with PY19 serum (polyclonal antibody specific for polyoma t antigens [34]) as described in the
protoblot immunoblottingsystem from Promega Biotec. One
viral stock was selected and taken through three rounds of plaquepurification.
Time course of protein production and accumulation.Cells, 3 x
106,
were seededinto each of 13 60-mm tissue culturedishes. Oneplatewasmockinfected, sixwereinfectedwith
recombinant baculovirusencoding
pp6O-src,
and theremain-ingsixwereinfected with recombinantbaculovirusencoding
MTAg. Cells were lysed at 15, 20, 24, 29, 40, and 52 h
postinfection (p.i.), and their protein concentration was
measured with the Bio-Rad protein assay kit. Total cell protein, 30 jug, was then resolved by SDS-PAGE,
trans-ferred electrophoretically to a 0.45-jim nitrocellulosefilter.
Thefilterwas thenblockedin 1% bovine serumalbumin in
TBST (10 mM Trishydrochloride [pH 8.0], 150 mM NaCl,
0.05% Tween 20) for 1 h at 37°C. The pp60c-src blot was
incubatedovernight with EC10serum (1: 2,500dilution),and
the MTAg blot was incubated for 4 h with PY19 serum (1:2,500 dilution). Both blots were then washed, incubated
with anti-mouse (pp60c-src blot) oranti-rabbit (MTAg blot)
alkaline phosphatase-conjugated immunoglobulin G for 45
min, and then developed by using the enzymatic color
reaction kit from PromegaBiotec.
Myristylation
ofpp6c-src.Cells,
3 x 106,wereseededinto eachoftwo60-mmtissue culture dishes. Afterattachment,the cellswereinfected with recombinantbaculovirus
encod-ing pp60c-srcatamultiplicity of infection of10. At 40hp.i.,
one plate was rinsed in methionine-free medium and then
labeledinmethionine-free mediumsupplementedwith 2 mM
glutamine and 0.67 mCi of [35S]methionine per ml. The
second plate was labeled with 2 mCi of[3H]myristic acid
(Dupont, NEN ResearchProducts) in afinal volume of1.5
ml. The[3H]myristic acidwaslyophilized and redissolvedin
12 34
5
6
7
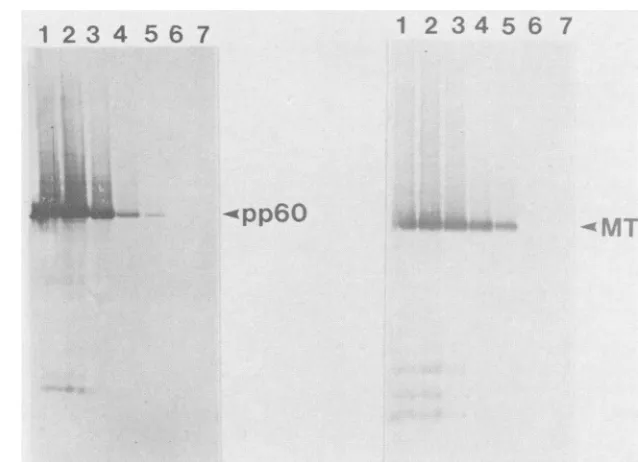
FIG. 1. MTAg andpp6Oc-srcproduction and accumulation. Sf9 cells were either mock infected(lane 7, bothpanels) orinfected with recombinantbaculovirusesencodingpp6O-src(leftpanel)orMTAg(right panel)at amultiplicityof infection of10. Cellswereharvestedat various times after infection; cell lysates were prepared, and proteins were resolved by SDS-PAGE. The proteins were blotted onto nitrocellulose andprobed with EC10 (left panel)orPY19(right panel)serum. Lane 1,52 hp.i.;lane2,40hp.i.;lane3,29hp.i.;lane4,24 hp.i.; lane 5, 20hp.i.;lane6, 15hp.i.; lane 7, mock infected.
on November 10, 2019 by guest
http://jvi.asm.org/
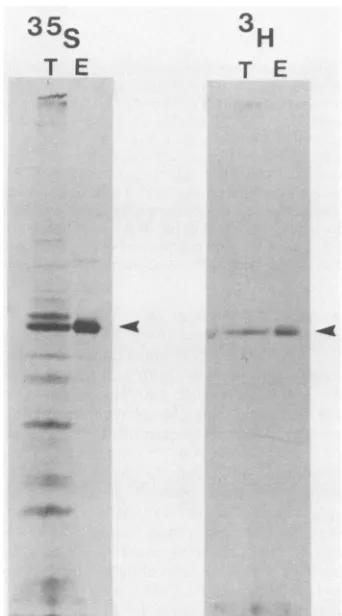
[image:2.612.146.465.446.677.2]15,ul of dimethyl sulfoxide prior to labeling. After 2 h, cells were lysed and proteins were analyzed directly by SDS-PAGE or immunoprecipitated with EC10 serum prior to SDS-PAGE.
Intrinsic and associated kinase activities. Cells, 9 x 106, were seeded intoeach of three 100-mm tissue culture dishes. After attachment, cells were mock infected or infected with recombinant baculovirus encoding MTAg or bothpp60-src and MTAg. Cell lysates were prepared at 40 h p.i. and normalized forprotein content. A 200-,ug amount of protein from each lysate was immunoprecipitated with eitherEC10 orPY19 serum, and kinase assays wereperformed in vitro. In vivo labeling and phosphopeptide mapping. Sf9 cells were infected with recombinant virus encoding
pp60c-src,
MTAg, or bothpp6Oc-src and MTAg each at a multiplicity of infection of 10. At 36 h after infection, cells were rinsed oncein phosphate-free medium and then incubated for 2 h in
phosphate-freemedium supplemented with 2 mM glutamine,
1.5% dialyzed calfserum, and 2mCiof32P-labeled P,per ml.
Cell lysates were prepared in a phosphate-RIPA buffer (10
mMsodiumphosphate [pH 7.2], 150 mM NaCl, 1% Nonidet P-40,0.1%sodium deoxycholate, 0.1% SDS, 2 mM EDTA)
supplemented with 2 mM phenyl-methylsulfonyl fluoride,
0.1% Aprotinin, and 2 mM sodium orthovanadate, and
pp60c-srcwasimmunoprecipitated with eitherEC10 orPY19
serum. Staphylococcus V8 and tryptic protease mapping was performed as described previously (14, 30), and
cyano-genbromidemappingwasperformedasdescribed by Jove et
al. (25). Phosphoamino acid analysis was performed as
described previously (35).
RESULTS
cDNAsencoding normal pp60c-src,a c-src mutant
contain-ing methionine rather than lysine at position 295, and the
MTAgofpolyomavirus werecloned intothe BamHI site of
pAC373 (49) to generate pAC373(c-src), pAC373(295), and
pAC373(MTAg), respectively. Sf9 cells were then
trans-fected with a mixture of wild-type baculoviral DNA and
pAC373(c-src), pAC373(295),orpAC373(MTAg).Fivedays
after transfection, medium supernatants containing both
wild-type and recombinant viruses were removed and
plaqued on a monolayer of Sf9 cells. Recombinant virus
plaqueswereidentified visually by theirocclusion-negative
phenotype (49). Recombinant virus was
purified
fromcon-taminating
wild-type virusby
three rounds ofplaque purifi-cation.Protein production and accumulation. The time course of
protein production
and accumulation in insect cells wasexamined asfollows. Sf9 cells were infected with
recombi-nant viruses encodingeither
pp6Oc-src (Fig.
1,leftpanel)
orMTAg(Fig. 1,right panel)at amultiplicity of infection of10.
Atvarious times afterinfection, cell lysates were
prepared
and protein production was examined
by
Westernblotting
(immunoblotting).
pp6Jc
srcandMTAgwerefirst detected20hp.i. (lane 5), increased
linearly
untilabout 40 h(lane
2),andcontinued to accumulate
gradually
from 52 to72 h(lane
1and data not shown). Breakdownproductswere detectable
by about 30 h p.i. (lane 3),
though
the bulk oftheprotein
remained stable for at least 3
days (data
notshown). By
Coomassie blue
staining,
we estimated thatpp6c-src
andMTAgcomprise
approximately
1% oftotal cellprotein.
pp60CSrCismyristylatedininsectcells.
pp({-csrc
ismyristy-latedin avianandmammalian cells
(3,
44)
andinyeast cells (29). To determine whether it wasmyristylated
in insectcells, we infected cells with recombinant baculoviruses
encoding pp6Oc-src and thenlabeled the infected cells with
either [3H]myristic acid(Fig. 2, right panel) or
[35S]methio-nine (Fig. 2,left panel).Total cell lysates or
immunoprecip-itates prepared by using antiserum specific for pp6Oc-src were then resolved by SDS-PAGE. Although pp6f4-src was the major protein labeled with
[35S]methionine,
severaladdi-tional proteins were also labeled during this time period.
Since the labeling period was started at 40 h p.i. when host protein synthesis is shut off, these proteins represent late virus-specific proteins.pp6Oc-srcwas the onlyprotein labeled with [3H]myristic acid. We do not know the fraction of
pp60-)srcthatismyristylated.
Intrinsic and associated kinase activities of pp60cs1c and MTAg in insect cells. To determine whether (i) pp6Oc-src
producedin insect cells was enzymatically active, (ii) MTAg
could associate with any endogenous insect tyrosine ki-nases, and (iii) MTAg and pp60-csrc would associate when
coproduced in insect cells, we performed the following
experiments: insectcells were mock infected (Fig. 3, lane 1)
orinfected with recombinant baculovirusesencoding MTAg
(lane 2) or MTAg and
pp6fc-src
(Fig. 3, lanes 3 and 5). Celllysates were prepared and immunoprecipitated with either
EC10serum (34)specific for avianpp6Oc-src(lane 5) or PY19 serum (36) which recognizes MTAg (lanes 1, 2, 3, and 4). Kinase assays were performed in vitro. MTAg was
phos-phorylatedin immunecomplex kinase assays in the presence
35s
T E
3T
T
E
FIG. 2. Myristylation ofpp6csrc. Sf9 cells were infectedwith recombinant virus encoding pp6Ocsrcandat 40hp.i. labeledwith either[35S]methionine(leftpanel)or[3H]myristicacid(rightpanel). Cell lysateswere preparedafter2hoflabeling, andproteins were
resolved directly by SDS-PAGE (T) or immunoprecipitated with EC10serumpriortoSDS-PAGE(E). Exposuretimeswere16 h for 35Gand 96 hfor'H.Arrowheads denoteppcswre.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.350.521.355.663.2]1
2
3
4
5
[image:4.612.97.262.79.377.2]SRCP--4MTe
30FIG. 4. Association betweenpp60csrc and MTAg in vivo. (Top) Cellswereinfected with virusesencoding MTAg (lane 1), pp6c-src (lane 4), or bothMTAg and pp60c-src (lanes 2 and 3). At 36 hp.i., cellswerelabeled with
32pi.
Celllysateswereprepared and immu-noprecipitated with either PY19 (lanes1and2)orEC10(lanes 3 and 4) serum.Immunoprecipitateswereresolvedby SDS-PAGE. (Bot-tom) Lanes1, 2, and 3 are the results ofphosphoamino acid analysis performedonMTAg isolated from lanes 1, 2, and 3, respectively, of thegel shown in the top panel. Lane 4 is the result of phosphoamino acidanalysis performedonpp6OcsrC isolated from lane 4 of the gel shown in thetop panel.FIG. 3. Intrinsic and associated kinase activities ofp60c-srcand MTAg. Sf9 cells were mock infected (lane 1) or infected with recombinantviruses encoding either MTAg(lane 2) orMTAg and pp6Oc-src (lanes3 and5). Celllysateswerepreparedat40hp.i. and immunoprecipitatedwithEC10 (lane 5)or PY19(lanes 1,2, 3,and4) serum, and kinase assays were performed in vitro. Lysates were also prepared from NIH 3T3 cells transformed by MTAg(lane 4 [10])andimmunoprecipitated withPY19serum, andkinase assays wereperformed invitro. The protein with the greatest electropho-reticmobility in lane4is a MTAg breakdownproduct (39).Forlanes 1, 2, 3,and 5, 200 ,ug of total cellproteinwasimmunoprecipitated, whereas 1.5 mg of totalcell proteinwasimmunoprecipitated in lane4.
(lanes3and5)butnotin the absence(lane 2) of
pp6Oc-src.
Inimmunoprecipitates prepared with EC10 serum (lane 5),
bothpp60csrc and MTAgphosphorylation isevident. EC10
serum immunoprecipitates both complexed and free pp60
c-src,
both ofwhichcanbephosphorylatedinvitro.This accounts for the significant phosphorylation ofpp6Oc-src inthisexperiment. Inimmunoprecipitates preparedwith PY19
serum(lane 3), onlythepp6Ocsrc complexed toMTAg was
immunoprecipitated and, since MTAg is thepreferred
sub-strate, phosphorylation ofpp6Ocsrc is less evident. MTAg was also phosphorylated in vivo by pp6Ocsrc when both
proteinswerecoproduced in insect cells (Fig. 4). Cells were
infected with viruses encoding
pp6fc-src
(Fig. 4, lane 4) orMTAg (lane 1) or with both recombinant viruses (lanes 2 and 3)andwerelabeledwith
32Pi.
Cell lysates were prepared andimmunoprecipitated with either PY19 (lanes 1 and 2) or
EC10 (lanes3and 4) serum. pp6Ocsrccoimmunoprecipitated
with MTAg, using antisera specific for MTAg (lane 2), and
MTAg coimmunoprecipitated with
pp6Ocsrc,
using antiseraspecific forpp6Oc-src (lane 3). pp6Oc-src wasphosphorylated
primarily on tyrosine and serine residues (Fig. 4, bottom
panel). MTAg was phosphorylated on serine when
synthe-sized aloneininsect cells (lane 1)andwasphosphorylatedon
tyrosine as well as serine when coproduced with pp6oc-src (lanes 2 and 3). We also found that MTAg reacted with an
anti-phosphotyrosine antibodyinimmunoblots fromlysates
prepared from cells coinfected with viruses encoding both MTAg andpp6Oc-src butnotfromlysatesprepared from cells infected with virus encoding MTAg only (datanot shown). Thus, MTAgwas phosphorylatedon tyrosine both in vitro and in vivo only when coproduced with pp6oc-src in insect cells.
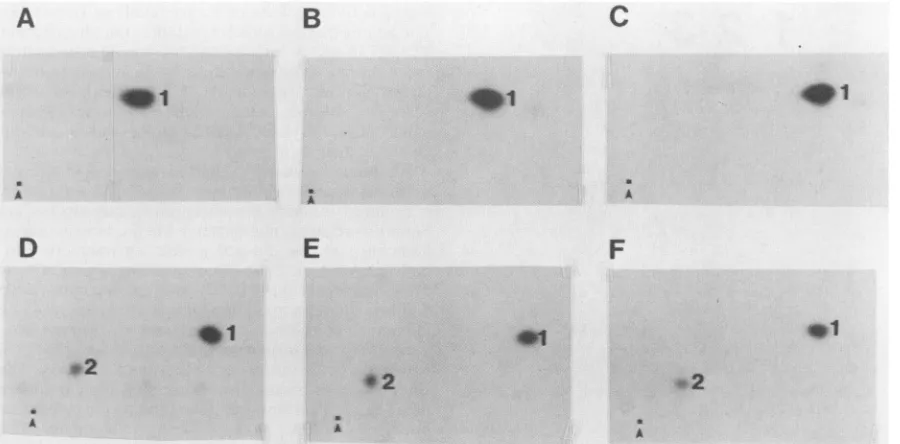
pp60j-srcisphosphorylatedprimarilyonTyr-416 and Ser-17
ininsectcells.To determine where thepp6Oc-src produced in insect cells was phosphorylated in vitro and in vivo, two-dimensional tryptic phosphopeptide mapping was per-formed.Tyr-416wasthe majorsite ofautophosphorylation
(Fig. 5A, B, andC).Tryptic digests of pp6O-src synthesized
and labeled in insect cells gave rise to two major tryptic
phosphopeptides (Fig. SD). Species2migratedat aposition
similartothetryptic peptide containingSer-17,and species
1 migrated at a position similar to the tryptic peptide
containing Tyr-416.Toverify this, wemixedtryptic digests
ofpp6Oc-srcsynthesizedininsect cells withtryptic digestsof
a c-src mutant thatwehave previously shown to be
phos-phorylatedonSer-17and Tyr-416 (40) (Fig. SE).Wedonot
know what fraction of pp6ocsrc produced in insect cells is
phosphorylatedonSer-17,but phosphotyrosine was present in greaterquantities thanphosphoserine by phosphoamino
acid analysis (Fig. 4, bottom panel) and more label was
incorporated into the peptide containing Tyr-416 than into
that containing Ser-17 (assuming equal recovery of both
peptides; Fig. 5D). Thus, it is unlikely that there is a
stoichiometricphosphorylation of Ser-17.
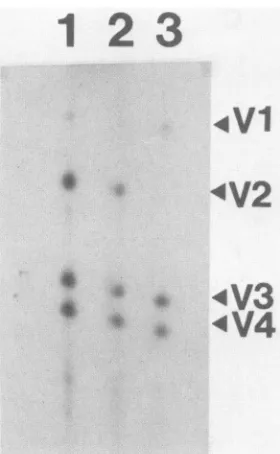
Since the phosphopeptide containing Tyr-527 is often
difficultto recoverduring trypticphosphopeptidemapping,
wealsoperformedone-dimensional cyanogen bromide
map-ping (data not shown). Using this method, we found that
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.336.533.81.268.2]A
A
D
B
E
C
A
F
@1
.2
*1
.2
;
AA
FIG. 5. Tryptic phosphopeptide mapping ofpp6Oc-srclabeled in vitroand in vivo. pp6Oc-srcwasimmunoprecipitated fromlysates of (A) infectedSf9cells or (B) NIH 3T3 cells overproducingpp6O-src,andautokinase assays were performed in vitro.Alternatively,pp6Oc-scwas immunoprecipitatedfrom 32P-labeledlysates of (D)infected Sf9 cells or (E) NIH 3T3 cells expressing a c-src mutant in which Tyr-527 is replaced by Phe. Immunoprecipitated proteins were digested with trypsin, separated by electrophoresis and chromatography in two dimensions, and autoradiographed. (C) Mix of phosphopeptides from panels A andB;(F) mix ofphosphopeptides from panels D and E. Spots 1and 2 designate the phosphopeptides containing Tyr-416 and Ser-17, respectively. The origins are marked witharrowheads. Electrophoresis atpH 8.9 was performed in thehorizontal dimension with the anode at the right.
pp6&c-src
was also phosphorylated to a small extent on Tyr-527(-10%of the level seen at Tyr-416). To distinguishbetweenthe possibility thatTyr-527 was being
phosphory-lated by a kinase present in Sf9 cells other than pp6c-src
itself, weexpressed akinase-negativemutantofpp60csrcin
insect cells and compared its phosphorylation pattern with
thatofnormal
pp60>src,
usingone-dimensional staphylococ-calV8protease mapping(aprotocol in whichphosphate onTyr-527 is stable). The kinase-negative mutant was
con-structed by replacing lysine residue 295 in pp60csrc with
methionine. Lys-295 is in the ATP-binding domain of
pp60c-src
and is essential for kinase activity (25, 48). TheLys-295toMet mutant hasbeenshown to be phosphorylated
onTyr-527 in aviancells (25). Ifanendogenous insect kinase
wasresponsible for phosphorylatingTyr-527, then this
res-idue would beexpected tobephosphorylated in theLys-295
mutant;if,onthe otherhand,pp60csrcwasitselfresponsible
for thisphosphorylation,then theLys-295mutantwouldnot
bepredictedtobephosphorylatedonTyr-527.The presence
ofV8
fragments
1, 3, and 4(Fig. 6) indicated that the Lys-295 mutant wasphosphorylated within its amino terminus,pre-sumably on Ser-17. However, the carboxy-terminal V8
fragment (V2) containing Tyr-416 and Tyr-527 was not
present in the mapofthe Lys-295 mutant (Fig. 6, lane 3),
indicating
thatthismutant wasnotphosphorylated
oneitherTyr-527orTyr-416.
DISCUSSION
Inthisreport, wedescribetheapplication ofabaculovirus
expressionsystem to(i) produce large
quantities
ofsoluble,
biologically
activepp60csrc
and MTAg;(ii) produce large
quantities ofakinase-negativemutantof
pp60-src
tobe usedas substrate to identify and purify kinases that
regulate
pp60c-src
inmammaliancells;
and(iii) develop
asystemforthe study ofMTAg-pp60c-src interactions invitro. The
bac-ulovirus expression system utilizes a helper-independent
virus (Autographa californica nuclear polyhedrosis virus)
whichcanbe grown tohigh titers incells adapted to Spinner
culture(49).Thus,highlevelsofrecombinantproteincanbe
obtained with relative ease.We haveobtained large
quanti-ties of soluble, biologically active pp6Oc-src and MTAg,
putting intherealm ofpossibilityathree-dimensional
struc-turalanalysis of both proteins.Inaddition, baculovirusesare
noninfectious to vertebrates andtheirpromoters have been
showntobe inactiveinmammalian cells(4). This isaclear
advantage overother eucaryotic viralvector systems when
expressing oncogenes or other potentially dangerous
pro-teins. Further, unlike bacterial expression systems, the
baculovirus systemis a
eucaryotic expression
system and,therefore, utilizesmanyoftheprotein
modification,
process-ing, and transport systemswhichoccurinhighereucaryotic cells.
Two of the three posttranslational modifications of
pp60c-src
whichusually
occurinhigher
eucaryotic
cells alsotake place in insect cells. These are
myristylation
andphosphorylation on Ser-17.
Myristylation
has also beenshown to occur in yeast cells (29). In mammalian
cells,
Ser-17 is
phosphorylated by
acyclic AMP-dependent protein
kinase(11, 38). Ahomologous enzymewiththe
capacity
torecognize avian
pp6Oc-src
must therefore exist in Sf9cells.Theonlyinvertebratehomolog ofc-srcthat hasbeencloned
andsequencedis from
Drosophila
melanogaster.
The aminoterminus of the
Drosophila
genediverges
greatly
from its vertebrate homolog,but it does encode aserineatposition
26which appearstobehomologous
toserine17 in vertebratepp60c-src
(45).Phosphorylation
ofTyr-527,
the third modifi-cation ofpp60c-src
thatusually
occurs in mammaliancells,
was also detectable at low levels in Sf9 cells. That theon November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.86.536.78.300.2]4V1
:V2
4V3
* 4s
*V4
FIG. 6. Staphylococcol V8protease mapping. pp60csrc(lanes 1 and2)and theLys-295to Metc-srcmutant(lane 3)were
immuno-precipitated from 32P-labeled lysates of infected Sf9 cells and resolvedbySDS-PAGE. Gel bandscontainingtherespective
pro-teins wereexcised and digestedwith staphylococcalV8 protease, and the proteolytic digestion products were resolved on a 15% polyacrylamide-SDSgel.V8fragments1, 3,and 4 are derivedfrom the amino terminusofpp6Oc-src,and V8fragment2 isderived from thecarboxylterminus ofpp6Oc-src.
Lys-295mutantwasnotphosphorylatedonTyr-527suggests that this low level ofphosphorylation was due to auto- or
transphosphorylation by pp6Ocsrcitself.
Drosophila c-src has been shown to encode a tyrosine residue ata position homologous to Tyr-527 in vertebrate c-src (45). Thus, the invertebrate src homolog may be
regulatedby amechanism similar to that used in vertebrate cells. The findingthat the Lys-295 mutantofpp6Oc-src was not phosphorylated on Tyr-527 in insect cells can be
ex-plainedinseveralways. (i)A527 kinasehomologmaynot be
presentininsect cells. (ii)The 527kinasemaybepresentin insect cells but its activity may be saturated by the large quantitiesof the c-srcmutantbeing produced. However,the
Ser-17 kinasewas not saturated under theseconditions.(iii)
The Tyr-527 kinase may be more labile than the Ser-17
kinase.
pp60csrc
is synthesizedlate in the viral replication cycle, long after host protein synthesis has been shut off. Thus, the Tyr-527 kinase may have been present prior toviralinfection butmayhaveturnedoverbythetimepp6Oc-src synthesis began. (iv) The determinants that make up the recognition site for the 527 kinase may not be shared betweenavianpp6Oc-srcanditsDrosophila homolog. Thus,it
is possible that an insect 527 kinase exists but does not recognize avian pp60csrc. (v) Finally, it is still formally possible that
pp60c-src
is itself the 527 kinase, but that itfunctions poorlyin yeastand insect cells(15, 16, 29).
Recent studies strongly suggest that Tyr-527 is a major
regulatory site inpp6Ocsrc. Phosphorylation ofthis residue has been shown to suppress the kinase activity and trans-forming potential of pp6Ocsrc Other members of the src
familyoftyrosinekinasespossessaTyr-527 homolog. Thus,
suppression by phosphorylation at this residue may be a
general mechanism for regulating thebiochemical and
bio-logicalactivities of the entiresrcfamilyoftyrosinekinases.
Thefinding that theLys-295mutantisnotphosphorylatedon Tyr-527 in insect cells makes the baculovirus system ideal
for preparinglarge quantities of this mutant to be used for
identifying and isolating the 527 kinase from mammalian
cells. In addition, other regulatory kinases, besides that which phosphorylates Tyr-527, may also be identified
by
using the approach.We have previously reported a region in the
carboxyl
terminus of
pp6Ocsrc,
borderedby Asp-516andPro-525,thatis required for MTAg association in mammalian cells
(9).
Since thisregion is not conserved in theDrosophilapp6Oc-src homolog (45), we did not predict an interaction to occur betweenMTAg and the insecthomolog of pp6Ocsrc. Indeed, MTAg synthesized in Sf9 cells was not detectably associated with any endogenous insect kinases when assayed in vivoor in immunecomplex kinase reactionsassayed in vitro. How-ever,MTAg did form a complex with avianpp6Oc-src in cells coinfected with viruses encoding both proteins. These re-sults make the baculovirus expression systempreferable to other expression systems for studying complex formation because: (i) preparations ofMTAg from higher
eucaryotic
cells would be expected to be contaminated with endoge-nous pp6Oc-src and (ii) pp6Oc-src and MTAg do not interact when coproduced in yeast cells (Kornbluth and Hanafusa, personal communication) or bacteria (Piwnica-Worms etal.,unpublished data) or when cotranslated in vitro, using a
rabbit reticulocytelysate (Piwnica-Worms and Roberts, un-published data). In addition, overproduction of either com-ponent in mammalian cells does not significantly increase levels of complex formation (39, 41, 43). These results suggest that there may be aninterplaybetween both positive
andnegativefactors which act inconjunctiontoregulate the
interactionof pp6Ocsrc and MTAg in mammalian cells. For
example, the posttranslational modifications undergone by
MTAg (i.e., phosphorylations) may be a positive factor
regulating complexformation in mammalian cells (42). We
caneasily test this hypothesis by comparing the
posttrans-lational modications undergone byMTAg ininsect cells and
thosewhichoccurinmammalian andyeast systems. Factors that inhibit complex formation in mammalian cells may include regulatory proteins, such as the 527 kinase, that compete with MTAgfor pp6Ocsrc binding. The baculoviral
expressionsystem may be the systemof choice for
identify-ing and purifying these various factors from mammalian cells.
Complex formation between MTAg and pp6Oc-src was
assayed both in vivo and in vitro. We found that MTAg
coimmunoprecipitatedwithpp6Ocsrc using antisera specific
for pp6Oc-src, and thatpp6Oc-srccoimmunoprecipitated with
MTAg, using MTAg-specific sera. In addition, MTAg was
phosphorylated on tyrosine in vivo only when coproduced
withpp6Oc-src and in vitro only when pp6Ocsrc was present in
the immunoprecipitates. Because the majority ofpp6O-src
producedin insect cells is not phosphorylated on Tyr-527, its
kinaseactivityisconstitutively high. It has, therefore, been difficult to determine whether the pp6Ocsrc associated with
MTAgisactivatedcompared with its unassociated
counter-part. Todate, we have been unable to measure any substan-tial enhancement in kinase activity when pp6csrc is
com-plexed to MTAg in insect cells. We are currently mapping
the complexed pp6Oc-src and MTAg to identify any novel
posttranslational modifications that occur as a result of
complex formation.
There remains uncertainty as to whether the complex formedbetween MTAgandpp6Ocsrc in insectcells is
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.108.248.77.304.2]tional. Because of the large quantities of protein
being
produced, the complexformationweobserved could be due
to a nonspecific aggregation of the two proteins. Using
antisera specific for MTAg, we have observed
coimmuno-precipitation ofpp6fvsrc and the v-fmsgene product when
eitheris coproduced withMTAg ininsectcells (althoughnot
to the same extent as with pp60-src). Neither of these
proteins has beendemonstrated to associate with MTAg in
mammalian cells. However, we do not observe
MTAg-pp6&c-src
complexes sedimenting to the bottom ofsucrosegradients, aswouldbeexpected forlarge nonspecific
aggre-gates. We also donot observea major shift inthe
sedimen-tation of eitherMTAgofpp60csrc from coinfectedcells when
compared with their sedimentation from singly infected
cells. When isolated from mammalian cells, the complex
sediments in the region of the gradient expected for a
220-kilodaltoncomplex (19, 20). These resultsmight suggest
that we have no complex formation inside insect cells, but
ratherthetwoproteinsassociateafterthecellsarelysedand
incubatedfor extended periods, as isthe case during
immu-noprecipitations. Alternatively, the complex isolated from
mammalian cells may contain proteins in addition to
pp60c-src and MTAgwhich contribute toitsgreater
sedimen-tation velocity, and thecomplex formedin insectcells may
be deficient in these other proteins. Several proteins have
been shown to associate with MTAg in mammalian cells
(33), and a
potential
phosphatidylinositol
kinaseofapproxi-mately 85 kilodaltonshas been shown tobe associatedwith
MTAg-pp60csrc
complexes
isolated from mammalian cells(18, 27). Further studies will
be required
to determinewhether MTAg and pp60csrc form afunctional
complex
ininsect cells.
Regardless of whether a functional
complex
is formedbetweenpp60c-srcandMTAgwithininsect cellsatthe timeof
theirsynthesis, thissystem
provides
apowerful
tool forthestudy ofpp60csrc and MTAg interactions in vitro. We are
currently purifying large
quantities
of MTAg, free ofcon-taminating tyrosine kinases, andlarge
quantities
ofpp6&-src
as afirststeptowardsthe
development
ofasystem in whichpurifiedcomponents canbe added and their
ability
todrivecomplex formation canbe assessed
individually.
ACKNOWLEDGMENTS
Wethank R. Jovefor adviceonthecyanogenbromide mapping, Debbie Morrison for the baculovirus encoding the v-fms gene product, and A. Desai for secretarial assistance.
This work was supported by Public Health Service grants CA30002 and CA43803 to T.M.R. and R29 CA50767 to H.P.-W. from the National Cancer Institute.
LITERATURE CITED
1. Bolen, J. B., V. DeSeau, J. O'Shaughnessy, and A. Shokreh. 1987. Analysis of middle tumor antigen and pp60csrc interac-tions in polyomavirus-transformed ratcells. J. Virol. 61:3299-3305.
2. Bolen, J. B., C.J. Thiele, M. A. Israel, W. Yonemoto, L. A. Lipsich, and J. S. Brugge. 1984. Enhancement of cellular src
geneproduct associatedtyrosylkinaseactivity following
poly-omavirusinfection and transformation. Cell38:767-777. 3. Buss, J. E.,and B. M.Sefton. 1985.Myristic acid, a rarefatty
acid, is thelipid attachedtothe transformingprotein ofRous sarcomavirus and itscellularhomolog.J.Virol.53:7-12. 4. Carbonell, L. F., M. J. Klowden, and J. K. Miller. 1985.
Baculovirus-mediatedexpressionofbacterial genesin
dipteran
andmammalian cells. J. Virol. 56:153-160.5. Cartwright, C. A., W. Eckhart, S. Simon, and P. L. Kaplan.
1987. Cell transformationby
pp60csrc
mutatedin the carboxy-terminalregulatory domain. Cell49:83-91.6. Cartwright,C. A.,P. L. Kaplan, J.A.Cooper,T.Hunter,and W. Eckhart. 1986. Altered sites of
tyrosine
phosphorylation
inpp60c-src
associated withpolyomavirus
middle tumorantigen.
Mol. Cell. Biol.6:1562-1570.
7. Cheng,S.H.,R.Harvey,P.C.Espino,K.Semba,T.Yamamoto,
K.Toyoshima,and A. E. Smith. 1988.Peptideantibodiestothe human c-fyngene product demonstratepp59fyn is
capable
ofcomplexformation with the middlet
antigen
ofpolyoma
virus.EMBOJ. 7:3845-3856.
8. Cheng,S.H., W.Markland,A. F.Markham,and A. E.Smith. 1986. Mutations around the NG59 lesion indicate an active association ofpolyomavirus middle-T
antigen
withpp60c-src
isrequiredfor cell transformation. EMBOJ. 5:325-334.
9. Cheng,S.H.,H.Piwnica-Worms,R.W.Harvey,T.M.Roberts,
and A. E. Smith. 1988. The carboxy terminus of
pp6('-src
is involved in complex formation with the middle-Tantigen
ofpolyomavirus. Mol. Cell. Biol. 8:1736-1747.
10. Cherington, V., B. Morgan, B. M.
Spiegehnan,
and T. M. Roberts.1986.Recombinant retroviruses that transduceindivid-ualpolyoma tumor
antigens:
effectsongrowth
anddifferentia-tion. Proc. Natl. Acad. Sci. USA 83:4307-4311.
11. Collett,M.S.,E.Erickson,and R. L. Erikson.1979.Structural
analysis oftheavian sarcomavirustransforming
protein:
sitesof
phosphorylation.
J. Virol.29:770-781.12. Collett, M. S., A. F. Purchio, and R. L. Erikson. 1980. Avian
sarcomavirustransforming protein p60srcshows
protein
kinaseactivity specificfortyrosine. Nature(London)285:167-169.
13. Cooper,J. A.,K. L. Gould, C. A. Cartwright, andT. Hunter. 1986.
Tyr5"
isphosphorylated
inpp60csrc:
implications
forregulation.
Science 231:1431-1434.14. Cooper, J. A., and C. S. King. 1986.
Dephosphorylation
orantibodybindingto thecarboxyterminus stimulates
pp6Oc-src.
Mol. Cell. Biol.6:4467-4477.
15. Cooper, J. A.,and A. MacAuley. 1988. Potential
positive
andnegative
autoregulation
ofpp60C-src
by
intermolecularautophos-phorylation.Proc. Natl.Acad. Sci. USA 85:4232-4236.
16. Cooper, J. A., and K. Runge. 1987. Avian pp60-src is more
active when
expressed
in yeastthan in vertebrate fibroblasts.Oncogene 1:297-310.
17. Courtneidge, S. A. 1985. Activation of the
pp60csrc
kinaseby
middle Tantigen binding
orby
dephosphorylation.
EMBOJ. 4:1471-1477.18. Courtneidge, S. A., and A. Heber. 1987. An 81kd
protein
complexed
with middle tantigen
andpp60-src:
apossible
phosphatidylinositol
kinase. Cell 50:1031-1037.19. Courtneidge, S. A., and A. E. Smith. 1983.
Polyoma
virustransforming protein
associates with theproduct
ofthe c-srccellular gene.Nature
(London)
303:435-439.20. Courtneidge, S. A., and A. E. Smith. 1984. The
complex
ofpolyoma
virus middle-Tantigen
andpp60c-src.
EMBO J. 3: 585-591.21. Coussens, P. M., J. A.
Cooper,
T. Hunter, and D.Shalloway.
1985. Restriction ofthe in vitro and in vivo
tyrosine protein
kinaseactivities ofpp60c-sc
relativetopp6o-src.
Mol. Cell. Biol. 5:2753-2763.22. Cross,F.R.,and H.Hanafusa.1983.Local
mutagenesis
ofRoussarcomavirus: the
major
sites oftyrosine
phosphorylation
ofpp6Oc-src
aredispensible
fortransformation. Cell34:597-607.23. Hunter, T.,and B. W.Sefton. 1980.
Transforming
geneproduct
of Rous sarcoma virus
phosphorylated tyrosine.
Proc. Natl. Acad. Sci. USA 77:1311-1315.24. Iba,H.,F. R.Cross,E.A.Garber,and H.Hanafusa.1985.Low level of cellular
protein
phosphorylation by nontransforming
overproduced p60c-sc.
Mol.Cell. Biol. 5:1058-1066.25. Jove,R., S. Kornbluth,and H. Hanafusa. 1987.
Enzymatically
inactivepp60csrc
with alteredATP-binding
site isfully
phos-phorylated
on itscarboxy-terminal regulatory region.
Cell 50: 937-943.26. Kaplan,D. R., B.Bockus, T.M. Roberts, J. Bolen,M. Israel,
and B. S. Schaflhausen. 1985.
Large-scale
production
ofpoly-omamiddle T
antigen by using
genetically
engineered
tumors.Mol.Cell. Biol.5:1795-1799.
27. Kaplan,D.R.,M.Whitman,B.
Schaffhausen,
D.C.Pallas,M.on November 10, 2019 by guest
http://jvi.asm.org/
White,L. Cantley, and T. M. Roberts.1987. Common elements in growth factor stimulationandoncogenic transformation: 85kd phosphoprotein and phosphatidylinositol kinase activity. Cell 50:1021-1029.
28. Kmiecik, T. E.,andD.Shafloway. 1987.Activation and
suppres-sion of pp60c-src transformingability bymutation ofitsprimary sites of tyrosinephosphorylation. Cell49:65-73.
29. Kornbluth, S.,R. Jove, andH. Hanafusa.1987.Characterization of avian and viral pp60src proteins expressed in yeast. Proc. Natl. Acad. Sci. USA 84:4455-4459.
30. Kornbluth, S., M. Sudol, and H. Hanafusa.1987.Association of polyomavirus middle-T antigen with c-yes protein. Nature
(London)325:171-173.
31. Kypta, R. M., A. Hemming, and S. A. Courtneidge. 1988. Identificationandcharacterization of pp59fyn(asrclike protein tyrosine kinase) innormaland polyomavirustransformedcells. EMBOJ.7:3837-3844.
32. Levinson, A. D., H. Oppermann, H. E. Varmus, and J. M. Bishop. 1980. Thepurified product of the transforminggeneof
avian sarcoma virus phosphorylates tyrosine. J. Biol. Chem. 255:11973-11980.
33. Pallas, D. C., V. Cherington, W. Morgan, J. DeAnda, D. Kaplan, B. Schaffhausen, and T. M. Roberts.1988.Cellularproteinsthat associate with themiddle and smalltantigensofpolyomavirus. J. Virol.62:3934-3940.
34. Pallas, D. C., C. Scley, M. Mahoney, E. Harlow, B. S. Schaff-hausen, and T. M. Roberts. 1986. Polyomavirus smalltantigen: overproduction inbacteria andutilization formonoclonal and polyclonal antibodyproduction.J. Virol.60:1075-1084. 35. Pallas,D. C., and F.Soloman. 1982.Cytoplasmic microtubule
associated proteins: phosphorylationatnovel sites is correlated with their incorporationinto assembled microtubules. Cell 30: 407-414.
36. Parsons, S. J., D. J. McCarley, C. M. Ely, D. C.Benjamin,and T.Parsons. 1984. MonoclonalantibodiestoRoussarcomavirus
pp6O-srcreactwithenzymatically activecellular pp6Osrcof avian andmammalian origin.J.Virol. 51:272-282.
37. Patschinsky, T., T. Hunter, F. S. Esch, J. A. Cooper, andB. M.
Sefton.1982.Analysis of thesequenceof amino acids surround-ingsites oftyrosine phosphorylation. Proc. Natl. Acad. Sci. USA 79:973-977.
38. Patschinsky, T., T. Hunter, and B. M. Sefton. 1986. Phosphor-ylationof thetransforming protein ofRoussarcomavirus:direct
demonstration of the phosphorylation ofserine17and identifi-cation of an additional site of tyrosine phosphorylation in
pp60v-srcofPragueRous sarcomavirus. J. Virol. 59:73-81. 39. Piwnica-Worms, H., D. R. Kaplan, M. Whitman, and T. M.
Roberts. 1986. Retrovirus shuttle vector for study of kinase activities ofpp6Oc-src synthesized in vitro and overproduced in vivo. Mol.Cell. Biol. 6:2033-2040.
40. Piwnica-Worms, H., K. B. Saunders, T. M. Roberts, A. E. Smith,andS. H.Cheng. 1987.Tyrosinephosphorylation regu-lates thebiochemical and biologicalproperties of pp6Ocsrc. Cell 49:75-82.
41. Raptis, L.,H.Lamfrom,and T. L.Benjamin.1985. Regulation of cellularphenotype andexpression of polyomavirus middlet antigenin ratfibroblasts. Mol. Cell. Biol. 5:2476-2485. 42. Schaffhausen, B. S., and J. Benjamin. 1981. Comparison of
phosphorylation oftwopolyomavirus middleTantigens in vivo and in vitro. J. Virol. 40:184-196.
43. Schaffhausen,B.S.,B.J. Bockus,K. L.Berkner,D.Kaplan,and T. M. Roberts. 1987. Characterization of middle T antigen expressed by usinganadenovirusexpression system.J. Virol. 61:1221-1225.
44. Schultz,A.M.,L. E.Henderson,S.Oroszlan,E. A.Garber,and H. Hanafusa. 1985. Aminoterminalmyristylation of the protein kinase p60srC, aretroviral transforming protein. Science 227: 427-429.
45. Simon, A.M.,B.Dress,T.Kornberg, andJ.M. Bishop. 1985. The nucleotide sequence and the tissuespecific expression of Drosophilac-src.Cell 42:831-840.
46. Smart, J. E.,H.Oppermann,A. P.Czervlofsky,A. F.Purchio, R.L.Erikson, and G. M.Bishop. 1981.Characterization of sites fortyrosinephosphorylation in the transformed protein of Rous sarcoma virus (pp6ov-src) and its normal cellular homologue (pp6Oc-src). Proc.Natl.Acad. Sci. USA 78:6013-6017. 47. Smith, A. E., and B. K. Ely. 1983. The biochemical basis of
transformationby polyoma virus. Adv. Viral Oncol. 3:3-30. 48. Snyder,M.A., J.M.Bishop, J.P.McGrath, and A. D.Levinson.
1985.AmutationattheATPbinding site of pp6Ov-src abolishes kinaseactivity, transformation, and tumorigenicity. Mol. Cell. Biol. 5:1772-1779.
49. Summers,M.D.,and G. E. Smith. 1987.Amanual of methods for baculovirus vectors and cloned insect cell culture proce-dures. TexasAgricultural Experiment Station, College Station. 50. Yonemoto, W., M.Javis-Morar, J. S.Brugge, J. B. Bolen, and M. A. Israel.1985.Tyrosine phosphorylation within the amino-terminal domain ofpp60c-src molecules associated with polyoma virus middle-sizedtumorantigen. Proc. Natl. Acad. Sci. USA 82:4568-4572.