0022-538X/90/020621-09$02.00/0
Copyright © 1990, American Society for Microbiology
The Human
Immunodeficiency Virus
Type
1-Specific
Protein
vpuIs
Required for Efficient Virus Maturation and Release
THOMAS KLIMKAIT,l* KLAUS STREBEL,1 M. DAVIDHOGGAN,1MALCOLM A. MARTIN,' AND JAN M. ORENSTEIN2
Laboratory of Molecular Microbiology, NationalInstitute ofAllergy and Infectious Diseases, Bethesda, Maryland
20892,1
andDepartment of
Pathology,
George
Washington University
MedicalSchool,
Washington,
D.C.200362Received25July 1989/Accepted2October1989
A deletion mutation affectingvpu wasintroduced intoaninfectious molecular clone ofhuman
immunode-ficiency virustype1, and theresultantphenotypewasexamined afterinfection of humanT lymphocytes. The
absence ofvpuresulted inanaccumulation of cell-associated viral proteins and impairedthe release ofprogeny
virions. Both electron microscopic and biochemical analysesindicated thata large proportion of themutant particleswasattachedtothesurface of infectedcells.Significant variation in the size and shape of theseprogeny
virions was observed. In addition, intracytoplasmic particles, some of which formed aberrant budding structures, were visualized inT cellsinfected with thevpumutant. Indirectimmunofluorescenceanalysesof culturesinoculated with wild-typevirus withuseofa vpu-specificantiserum demonstrated thatvpuismainly
localizedtoaperinuclearregion in the cytoplasm of virus-producing cells.
Human immunodeficiency virus type 1 (HIV-1) contains numerous openreadingframes (ORFs), many of whichare
found in other lentiviruses. The vpu ORF, however, is
unique to HIV-1, not being present in the genomes of the
closely related primatelentiviruses HIV-2 and simian immu-nodeficiency virus (SIV) (10, 12). The vpugeneproductisa
small,81-amino-acid protein that is expressed in vivo (3, 16, 23). Because of its structural similarity to the membrane protein M2 encoded by influenza virus and the previously reported 4-to10-foldreduction inprogenyvirion production
associated withtheinterruptionof the HIV-1 vpuORF (23), wesuggested thatvpumight playarolein virion maturation,
efficient particle release from HIV-1-infected T lympho-cytes, orboth.
In this report, we have extended our analysis of vpu
functionduring productive virus infection of human T cells. We show that vpu is expressed only in the cytoplasm of
infected cells. In the absence ofvpu, an accumulation of cell-associated virion proteins associated with increased cytopathicitywasobserved. The results of cell fractionation studies and electron microscopic analyses indicate that the
vpumutantphenotype in infected T cells is characterizedby
adefectinvirion release.
MATERIALS AND METHODS
Cells and viruses. The human T-lymphocytic cell line A3.01(4)waspropagated in RPMI 1640 supplemented with 10% fetal bovine serum (FBS). Human peripheral blood leukocytes collected from healthy donors werepurified by centrifugation inlymphocyte separation medium (Organon Teknika, Durham, N.C.) and stimulated with 0.25 ,ug of phytohemagglutinin per ml in RPMI 1640 containing 10% FBS 3 days before infection. After HIV infection, the medium was supplemented with 10% interleukin-2. The
tissue culture mediumwaschangedeverythirddayorwhen the cell concentration exceeded 2 x 106/ml. HeLa cells (ATCCCCL2)orHeLa CD4 cells(a gift fromR.Axel)were
propagated in Dulbecco modified Eagle medium (DMEM) containing 10% FBS. Filtered supernatants, harvested
ap-*Correspondingauthor.
proximately 48 hposttransfection,wereusedasvirus stocks
for wild-type or mutant virus. Confluent cell monolayers
wereroutinely split 1 to5.
Constructionof the vpumutant. Adeletionmutant of the infectious molecular HIV-1 clone pNL4-3 (1) was
con-structedby site-specific oligonucleotide-directed mutagene-sis of the2.7-kilobase EcoRI-BamHI fragment cloned into M13. A deletion of 48nucleotidesimmediately following the initiator methioninecodon of the vpuORF plusa
7-nucleo-tideinsertionresulting inaminus-1 frameshift andpremature termination after 14 missense codons wasintroduced (Fig.
1). The mutated EcoRI-BamHI fragmentwasreinserted into
the infectious pNL4-3 proviral clone; the mutation was
confirmedby sequencing, using the dideoxy-chain termina-tion method (19).
Transfection of HeLa cells. HeLa cells were grown to
about 70%confluency ina25-cm2 flask. Beforetransfection, the medium was replaced with 5 ml ofDMEM containing
10% FBS. Calciumphosphate-precipitated plasmidDNA(20
,ug)
was addedto the cellsfor 4h,
after which the mediumwas removed and the cells were subjected to a glycerol
shock for 2min(9). The cultureswerethen washed twicein DMEM and maintained in 5 ml ofDMEM supplemented with 10% FBS.
RTassay. Reversetranscriptase (RT) assays oncell-free mediumsamples (8)wereperformedaspreviouslydescribed
(24). Samplesfor intracellular RTactivitywerepreparedas follows. Infected-cell suspensions (1 ml) were pelleted and suspendedin 1 mlofphosphate-bufferedsaline (PBS). Cells were disrupted by three cycles of freezing on dry ice and thawing ina37°Cwaterbath. Aftervortexingand sedimen-tation of cell debris, 10-,ul samples of supernatant were subjected toRT assay.
Western immunoblot analysis. Cells were pelleted and
lysed in CHAPSbuffer(50mM Trishydrochloride [pH 8],5 mMEDTA, 100 mMNaCl,0.5% CHAPS [Behring Diagnos-tics, LaJolla, Calif.]) containing0.5% sodiumdeoxycholate. Insoluble material was removed by centrifugation, the
ly-sates were boiled for 5 min in anequal volume ofsample buffer (14), and the proteins were separated on 10 to 20% sodiumdodecyl sulfate-polyacrylamide gradient gels. After 621
on November 10, 2019 by guest
http://jvi.asm.org/
622 KLIMKAIT ET AL.
A
1 2 3 4 5 6 7 8 9 kb
.~~~~~~~~~~~~~~~~~~~~
vpu
| _ rev -_
_~~~~~~~~~~~~~~~4
gagInetY
%,pr
B
SA5961 +
...CATCTCCTATGGCAGGAAGAAGCGG sis
6-revlit
AGACAGCGACGAAGAGCTCATCAGAACAGTCAGACTCATCAAGCTTCTCTATCAAAGCAGTAAGTAGTACATGTA 66so
rev 'Pt tat
Pt-,.--VpU
wts M 9 P r Z v a Z v a L V v a Z Z Z a Z v v gr s Z v
ATGCAACCTATAATAGTAGCAATAGTAGCATTAGTAGTAGCAATAATAATAGCAATAGTTGTGTGGTCCATAGTA 13s
AULATGCTTTCTTT- CAATAGTTGTGTGGTCCATAGTA
wtFiz z Y Rx x L R Q R X Z D R L Z D A ZZ H X A X ATCATAGAATATAGGAAAATATTAAGACAAAGAAAAATAGACAGGTTAATTGATAGACTAATAGAAAGAGCAGAA 62t
AUF N H R I *
Wt > D S G N H S H GC V S A L V N M G V H M G H1 H A P
GACAGTGGCAATGAGAGTGAAGGAGAAGTATCAGCACTTGTGGAGATGGGGGTGGAAATGGGGCACCATGCTCCT6285
L-.,env
wt VN D Z D D L
TGGGATATTGATGATCTGTAGTGCT...
6305
FIG. 1. (A)Genetic map ofHIV-1 indicatingtheposition ofthevpugene in the viral genome.(B)Nucleotidesequenceof the vpu gene present inthe parentalwild-type (wt)plasmid pNL4-3 and the deletionmutantvpuDEL-1(AU).Thebeginningof the firstcodingexonofrev
and thesplicedonor site(SD)atthe end of the firstexonsofrevandtatareindicated. SA marks theputativespliceacceptor for vpu. The deduced amino acid sequences forwild-typeanddeleted vpuarealso shown. Thebeginningof theenvcodingregionis marked. Numbers indicate the nucleotide distance from the beginning of the5' long terminal repeat.
transferto
0.22-gim-pore-size
nitrocellulosemembranes, thefilterswereincubated with specific antiseraand
251I-labeled
proteinAandthen subjected toautoradiography (21).
Immunohistochemistry. In a modification ofa previously describedmethod(18), cells(3 x 105)were washed in PBS andcentrifugedontoglass slides(5 min at 250 x g),usinga
Cytospin centrifuge (Shandon Inc., Pittsburgh, Pa.). The slides were air dried, and the cells were fixed either in
ice-coldacetone for 5 min or in
3.7%
paraformaldehyde for 10 min. The dried samples were blocked for 1 h in PBScontaining
5% bovine serumalbumin. The seraused fortheprimary antibody reaction were preadsorbed with
acetone-fixed uninfected A3.01
cells
and cleared by centrifugation (9,000 x g for1 min). The anti-vpu serum raised in rabbitsagainstavpufusion protein synthesizedinEscherichia coli (F.Maldarelli, manuscriptinpreparation)and serum from an HIV-1seropositive individualwere each diluted 1:50. After
reaction with the primary antibodies, thecells were
exten-sivelywashed for 1 h with PBS containing 1% bovine serum
albumin. Fluorescein isothiocyanate-conjugated goat
anti-rabbitimmunoglobulin G(IgG) (Organon Teknika,
Malvern,
Pa.) for the anti-vpureaction ortetramethylrhodamine
isothio-cyanate-coupled anti-human IgG antibodies, developed in goat
(Qrganon
Teknika), were used in concentrations rec-ommended by the manufacturer as secondary antibodies. Aftera40-min incubation, the slides were washed twice inPBS, rinsed in distilled water, and analyzed by using a fluorescence-activatedlightmicroscope (Zeiss, Oberkochen,
FederalRepublic ofGermany).
Electron microscopy. At different times after infection,
samples of the infected lymphocyte cultures containing
ctlution B
A
101
103
10 3x10 6x10 104
4 3x10
FIG. 2. Endpoint dilution titration of wild-type (A) and vpuDEL-1(B) virus. Filteredculture mediumfromwild-type-and vpuDEL-1-infected lymphocyte cultures was adjusted to equivalent RT activity(approximately6x 106cpm/ml). The indicated dilutions of supernatantwereusedtoinfect A3.01 cellsinsixfoldreplicas.An autoradiogram of a 32P-RTassay performed on cell-free superna-tants16days postinfectionis shown.
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.157.468.74.414.2] [image:2.612.326.563.546.661.2]A.
E - 15
0
v-.0 CD
iF.1
A
?*
RTC.)
0
S
_ _
...
_ :1.
EuI
B
0
to
I
1
a: c.
10
S
10
AU
(x10Qcpm)
15
10
5.
5 7 9
-m
n
m
wt
F
1-1111 0
1-
_ I I I I I I time 11 14 17 5 7 9 11 14 17 (daysp1.)..
49
-gP1600 -gp120
__v . -p55
-t -p41
t(daysp.i.)
B. wt 'U
;
v . ...J;X4W~ ~ ~ ~ \.
FIG. 3. Cytopathicity of wild-type (wt) and vpuDEL-1
(A&U)
virus. (A) Cell viability of A3.01 cells after infection with wild-type pNL4-3 or vpuDEL-1 was monitored at the indicated times by trypan blue exclusion. RT activity in corresponding supernatant sampleswasalsodetermined. (B) Cell fusionwasassayed in HeLa CD4 cellstransiently transfected withequivalentamounts of wild-type or vpuDEL-1 plasmid DNA as described in Materials and Methods. The formation of syncytia was scored under a phase-contrast light microscope 20 h after transfection (magnification, x62). Syncytia are marked witharrowheads. Supernatant RT activ-ity for vpuDEL-1 was only 1/10 the amount produced in the transfection with wild-type virus.
approximately 106cells werepelleted by low-speed centrif-ugation (320 x g for 10min) and fixed for 15 min at room temperature in5mlofa2.5%glutaraldehyde solutionin 100 mM cacodylate buffer as previously described (17, 20). Samples were stored at4°C until they were processed for
ultrastructural studies.
RESULTS
Molecularcharacteristicsof the vpu deletion mutant. A vpu deletion mutation(vpuDEL-1),identical to thatpresentin a naturally occurring HIV-1isolate designatedNY-5 (2),was introducedinto theinfectious molecularclonepNL4-3 (1)as described in Materials and Methods. Unlikeour
previously
describedlinker insertion mutant, which had the
potential
to encode the amino-terminal 32 residues of vpu (23), the deletionpresent in vpuDEL-1 allowedonly
thesynthesis
of a15-residue missensepeptide.
The deduced missense amino acids ofthe mutated vpu lack anyhomologywith the authentic
protein
(Fig.
1). TheFIG. 4. Accumulation ofHIVproteins inlymphocytes infected with the vpu mutant. At the indicated times, cells and culture mediumwereseparated bycentrifugation (9,000xgfor 3 min). The supernatantfractionwasassayed forRTactivity, andthelysedcell pellets were electrophoretically separated and analyzed by immu-noblotting, using human anti-HIV-1 serum. vpu is notidentified in the blotsince thepredominantp17gagprotein comigrates with vpu and obscures it(23). wt,Wild type;
AU,
vpuDEL-1mutant.splice donor sites for the first coding exons oftat and rev
immediately5'ofthe vpu gene areintact, and the envgene
located 3' from the vpu mutation is preserved (Fig. 1).
Western
analysis of transfection lysatesor radioimmunopre-cipitated lysatesofvpuDEL-1-infectedTcellsindicatedthat noneoftheotherHIVORFs is affectedby thevpudeletion.The banding
patterns
and relative band intensities ofwild-type and mutant
proteins
were indistinguishable (data notshown).
Theinfectivity of mutant virionsisunaffectedby the dele-tion. The infectivity of vpuDEL-1 virus
preparations
wascompared
with that oftheisogenic
wild-type virus inend-point
titrationexperiments.
Wild-type
andvpuDEL-1
virus stocks wereprepared
byfiltering
supernatants from HeLacells 48 h after transfection. After normalization for RT
levels, A3.01 cellsin 96-well microdilution plates
(103
cellsper well) were infected with serial dilutions of each virus stock; each dilutionwasplatedinsixreplicas.Theinfectious
titerof each stockwasdetermined
by
RTassayoftheculturesupernatant.
Wild-type
and vpuDEL-1 HIV-1 had similar infectivities, although vpuDEL-1 consistentlygenerated
lower virus
yields
at each dilution(Fig. 2).
It should beemphasized
that in thisinfectivity
assay,comparable
amounts(RT activity) ofcell-freefiltered virus were exam-ined;aggregatedorcell-associatedformsofvpuDEL-1 (see
below) were
probably
eliminated from the test inoculum.Furthermore, immunoblot
analyses
ofwild-type
or vpuDEL-1particles, obtainedby
pelleting
viruspresent ineachfiltrateby
ultracentrifugation,
wereindistinguishable,
indicating
similarproteincompositions
(data
notshown).
In contrast, infection ofA3.01 cells with
vpuDEL-1
re-sulted in amore
rapid
onsetofcytopathicity
incomparison
with wild-typevirus. Greater cellkilling
was evident in ther%
Lin .,ft,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.53.559.69.390.2] [image:3.612.281.555.69.322.2]624 KLIMKAIT ET AL.
As ofratioRT
-
0-, 1.6
ID 1.2
0
°.S0.8
= 0.4
0
.B
Bs
:0*5
50F
A
0- AU
--*wt
5 0 5
5 10 15 t
(daysp.i.)
wt
AU
[/,1
2L 0
1c
FIG. 5. Cell-associated RTactivity. Samples of A3.01 cell cul-tures infected with equivalent amounts (3 x 106 cpm of
32P-RT
activity per ml) ofwild-type (wt) or vpuDEL-1 (AU) virus were
sedimented at the indicated times after infection. (A) Ratio of cell-associated RT activity, liberated byfreezing and thawingthe cellpelletororiginally present in themedium,plottedasafunction of timepostinfection.Thecell-associated viruswasfurther fraction-ated (B) byseparating virions loosely attached tothe cell surface from intracellular virusbyvortexingandcentrifugationtogenerate thesurface fraction. Thepelletedcellswerethenresuspendedin the original volume and disrupted by freezing and thawing (cytosol fraction).
vpuDEL-1-infected cultures after day10 eventhough
signif-icantly less progeny virus (as measured by RT
assay)
was produced (Fig. 3A). Similarly, vpuDEL-1proviral
DNA induced larger and more numeroussyncytia
in transient transfections of HeLa CD4+ cells than did thewild-type
provirus(Fig. 3B)yet
generated
only10%
of theRTactivity
observed in the lattertransfection.
Lack of vpu leads to an accumulation of viralproteinsand cell-associated HIV particles.The effect ofthe vpu mutation
onthe
synthesis
ofHIVproteins
and the releaseofprogenyvirions into the
supernatant
medium is depicted in Fig. 4.Cells andmediawere collected at the indicated timesafter
infectionwithwild-typeorvpuDEL-1 virus and analyzed by immunoblotting (Fig. 4B) or RT assay (Fig. 4A),
respec-tively. Duringtheearly phase (day 9) oftheinfectioncycle,
lysatesfrom
vpuDEL-1-infected
cells contained nearly five times more viral protein than comparable samples from awild-type virusinfection. Even atlatertimes,theamountof
on
B
~4. .
4~~~~~4
'w kj.' ,- :r
10~~~~
.. . ^ e j; or ., R~~~~~~~~r'v
.. ^< SX :E ;0S
*8'
iaS
t',01
^X^s~~~~~
FIG. 6. Low-magnification transmission electron micrographs otA3.01cellsintectedwith wild-type (A) or vpuDEL-1 (B) virus showing virions associated with the cell surface. Magnification, x9,000.
ii
J. VIROL.
MLI
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.68.561.87.712.2]HIV-1-SPECIFIC PROTEIN vpu 625
A
4-f;
k!; X4; .
y,¢<....+Jr1';|
B~~~''X
w |@h. *
.iFiw
2 ,.b. vo
b
* #N
6
,^fs.
W;'lrav&,
'I--,2
*~~~~ *
OF,~ ~ ~~~ F
*
.~~~~~~~~~~~~~~Ilk
: w
s. '.
r9
'if
C
W'W ;S4 ;".
i
VF
..
;4,
-4.
p:e
-4U
p.
cell-associated HIV proteins in cells infectedwith the vpu
mutantwasconsiderably higher. Acorrection for the lower
number of viable cells in the vpuDEL-1 infection at the indicated time points would result inan evengreater
accu-mulation of proteins. In contrast, and as reported earlier
(23), significantly more(fivefoldintheexperiment shown in Fig. 4) wild-type virus than vpuDEL-1wasreleased into the
medium.
N
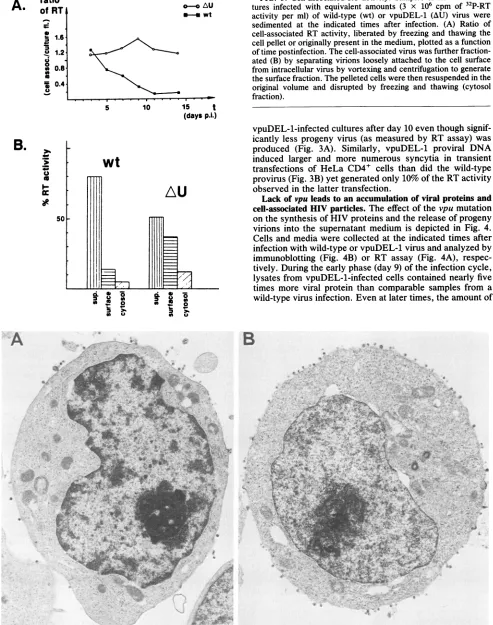
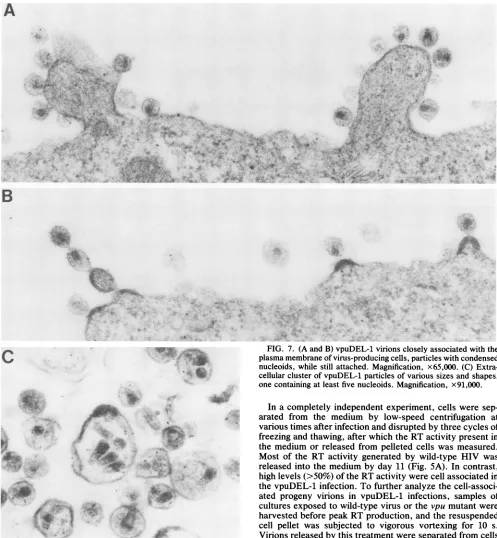
FIG. 7. (A and B) vpuDEL-1 virions closely associated with the plasma membrane of virus-producing cells, particles with condensed nucleoids, while still attached. Magnification, x65,000. (C) Extra-cellular cluster ofvpuDEL-1 particles of various sizes and shapes, onecontainingatleastfive nucleoids. Magnification, x91,000.
Ina completely independent experiment, cellswere
sep-arated from the medium by low-speed centrifugation at
various times after infection and disrupted bythree cyclesof freezing and thawing, after which theRTactivitypresent in the medium orreleasedfrom pelleted cells wasmeasured.
Most of the RT activity generated by wild-type HIV was
released into the medium by day 11 (Fig. 5A). In contrast,
highlevels(>50%) of theRTactivitywerecell associated in
thevpuDEL-1 infection. Tofurtheranalyze the cell-associ-ated progeny virions in vpuDEL-1 infections, samples of culturesexposed towild-type virusorthevpumutant were
harvestedbefore peakRTproduction, and the resuspended cell pellet was subjected to vigorous vortexing for 10 s.
Virions releasedbythistreatmentwereseparatedfrom cells by centrifugation (surface fraction), and the remaining cell pelletwasresuspendedinmedium,frozen and thawedtwice, andcentrifugedafinal time(cytosol fraction). Nearly75% of
thecell-associated RT invpuDEL-1 infectionswasreleased by vortexing (Fig. SB). A simultaneous trypan blue exclu-sion assay indicated that most of the vortexed cells were viable (data notshown). These results suggest that a large
proportion of the cell-associated vpuDEL-1 particles may
have beenlooselyattached tothe cell surface.
Electron microscopic examination ofvpuDEL-l-infected T cells.Approximately 10i A3.01 cellswereinfected withequal
.t
VOL.64, 1990
44f,--.
,I
I
.6
Ik. "-,
X. :,. .,
A
*
AL:
JFT
*to.7
At
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.57.554.79.617.2]626 KLIMKAIT ET AL.
A
a s -f W }
1*
4v*
M..
,;42..r.-X"';,i'.F.,lki' ;;.A'A.< + tf '
It
'71k
ZU
i
ci;
i
~~t
iS
4
t ''
i + j
f
v~~~~~~S
i;
,
v 7g i F r; ; rr ;s rt~-4 .t'{:,^, ,, ..
Afr
s
5'.' X~~~~~~~~~~~~~~~~~~~~L
D
i'e
B
J. VIROL.
- P.
.~~~~~~~~~~~~~~~~~~~~~~7
*e"W
.,.P-W%
t.'-h
-P'
on November 10, 2019 by guest
http://jvi.asm.org/
amounts (based on RT activity) of wild-type or vpuDEL-1 at amultiplicity similar to that used for the experiment shown
in Fig. 3; samples were collected on day 7, 9, or 10 and examined by transmission electron microscopy. Although the percentages of virus-producing cells were comparable (by transmission electron microscopy and indirect
immuno-fluorescence) in the two cultures, up to fivefold more parti-cles were associated with the surface of cells infected with vpuDEL-1 (Fig. 6). A characteristic electron microscopic
feature of the vpuDEL-1 infection was the presence of progeny particles, physically attached to the plasma
mem-brane of infected cells, that contained prematurely con-densed nucleoids (Fig. 7A). In some instances, chains of vpuDEL-1 virions were observed extending from the plasma membrane (Fig. 7B). In addition, a greater proportion of the vpuDEL-1 particles appeared aberrant compared with wild-type HIV,varying in size and shape, and in some instances appeared to harbor multiple nucleoids (Fig.7C).
Another common feature of vpuDEL-1-infected A3.01 cells was the presence of intracytoplasmic vacuoles
contain-ing large numbers of virions in all stages of assembly,
including highly aberrant forms (Fig. 8). Atypical detached
single, double, and multimeric budding structures that
formed rosettelike structures were also found (Fig. 8C). Besides virions, the cytoplasmic vacuoles frequently
con-tained small vesicles similar to multivesicular bodies
typi-cally associated with Golgi elements (Fig. 8D).
vpuDEL-1-infected peripheral blood leukocytes exhibited a similar
phenotype (data not shown).
vpu is located in the cytoplasm of virus-producing cells. Acetone-fixed and permeabilized A3.01 cells, 9 days after
infection with wild-type HIV-1, were simultaneously
incu-bated with a monospecific vpu antiserum raised in rabbits and human serum with HIV-1 seroreactivity and then
exam-inedfor the presence of viral proteins by indirect
immuno-fluorescence as described in Materials and Methods. The human anti-HIV-1 serum,whichreactedwell with HIV gag and env but not vpu in immunoblots (data not shown), gave strongcytoplasmic fluorescence (Fig. 9A). The signal with theanti-vpuserum wasalso strong and could be localized to theperinuclear region ofthecytoplasm;no nuclear reactiv-ity wasapparent (Fig. 9B). In control infections with vpu-DEL-1, thereaction withthe serumfromtheHIV-1-infected individualwassimilarto thewild-typereaction (Fig. 9C),but no reactivity was observed with the anti-vpu serum (Fig.
9D). When cells fixed withparaformaldehyde to study
sur-face reactivity were examined with the same antibodies, intense staining was observed with the human anti-HIV-1
serum(Fig. 8E), presumablybecauseofthepresence of gag or env protein on the plasma membrane, but no reaction above background levelswasobservedwith the vpu antise-rum(Fig. 8F).
DISCUSSION
We have recently reported that the HIV-1 vpu gene product is anintegralmembrane
protein
that isexpressed
atanti-HIV-1 anti-vpu
FIG. 9. Localization of vpu by indirect immunofluorescence microscopy. Wild-type (A, B, E, and F)- or vpuDEL-1 (C and D)-infected lymphocyteswerefixed withacetone (A toD)orin3.7% paraformaldehyde-PBS (E and F). Samples were simultaneously incubated with human anti-HIV-1 serum and an anti-vpu serum raised in rabbits. Tetramethylrhodamine isothiocyanate-conjugated goat anti-human IgG antibody and a fluorescein isothiocyanate-coupled mouse anti-rabbitIgG antibody were used as secondary antibodies, and fluorescence of the same fields was evaluated. Magnifications: AandB, x504;CtoF, x332.
highlevels invirus-producing Tlymphocytes but is largely, if not completely, absent from progeny virions (23; this report). Mutations affecting the synthesis of vpu lead to a marked reduction in virus particle production and an asso-ciated intracellular accumulation of HIV structural proteins
(23). Inthis study, vpu was found only in the cytoplasmof productively infected cells by indirect immunofluorescence andappearedtobeconcentrated ina subcellularorganelle,
most likelytheGolgicomplex.
Oneofthe moststriking findings of this work camefrom the electron microscopic analyses, which suggest that vpu
playsacriticalroleduringthe release of virusparticlesfrom infectedTcells.Inthe absenceofvpu, increased numbers of
virus aggregates and budding structures, including many mature particles characterized by condensed cores, were found associated with or evenattached totheplasma mem-brane ofvirus-producing cells. Concomitantly, budding of progeny vpu mutant particles from intracytoplasmic mem-branes into vacuoles was also seen, explaining in part the intracellularaccumulationof HIVproteinsobservedin met-abolic labeling (22) and cell fractionation (Fig. 5) experi-ments. Thus, in additiontoresultinginimpaired release of
FIG. 8. Intracytoplasmic vacuoles withincells infectedwithvpuDEL-1-containing particles invarious stages ofassembly. Shown are budding vacuolar particles (A to D) as well as highly aberrant structures containing multiple nucleoids of aberrant shape (A and C; arrowheads)orinapparentcores(A).Matureand aberrant virionscontainingeccentricdensematerialor anucleoid withahollowappearance (AandC)canbeseen. Incompletelyseparatedadjacentbuddingunits formastalklikestructure(B)ordetachedrosettelike structures(C; stars)consistingofsingle andmultiplebuds locatedwithinvacuoles ofamultinucleated giantcell. (D)Two vacuoles that contain mature virionsas well assmall smooth vesicles (arrows). Three typical budding structures are presenton the plasma membrane on the right. Magnifications: AandB, x110,000;C, x64,000;D, x80,000.
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.320.558.64.312.2]628 KLIMKAIT ET AL.
hdex
4- vpu
A
2
I
,
p3A
FIG. 10. Hydropathicity profiles of the deduced amino acid
sequences for HIV-1 vpu, foot-and-mouth disease virus p3A, and
influe'nza virus M2 proteins. The relative hydropathicity index of each residue is shown as afu'nction of amino acid position in the
proteinandwascalcu'latedbythemethod ofKyteandDoolittle(13).
The border between hydrophobicdomain andhydrophilicC
termi-nusof eachprotein isindicatedbythe verticalbroken line.
virions from thecell surface oflymphocytes, theabsenceof
vpu results inthe budding and maturation of virus particles
at membrane sites ordinarily restricted from such activity.
Perhaps the high concentrations ofvpu associated with the
Golgicomplexincells infected withwild-typeviruspreclude
particleproduction atthese intracytoplasmic membranes.
The electron microscopic studies also demonstrated that
vpuDEL-1 infections ofT lymphocytes were characterized
bytheproductionofaberrant virusparticles, including some
with multiple nucleoids and others that were several times
larger than wild-type virions and contained uncondensed
nucleoids. Thiswasaprominentfeatureofintracytoplasmic
vpuDEL-1 virus production, in which multimeric budding
structures werefound. One explanation linking all of these
phenomenais that vpumediatesacritical and terminal step
of thebuddingprocess; in its absence, individual immature
progenyparticlesfailtoseparatefrom oneanotherandfrom
the plasma membrane. This perturbation of virus release
could lead to aretardation ofHIV maturation and
second-arilyresult inthe accumulation ofhighintracellular
concen-trations of viral
piroteins
as well as the diversion of virusproduction to intracellular sites. A paradox of vpuDEL-1 infectionswasthe earlierappearance ofvirus-induced
cyto-pathicity thatwas accompanied by significantredu'ctions in
progenyvirionproduction. The intracellular accumulation of
potentiallytoxic viralproteins invpuDEL-1-infectedT
lym-phocytesmight explain the observed reduced cellviability.
Similarly,
high
concentrations ofvirionstructuresassociated with essential cellular membranes could alter theircompo-sition and impair function by altering permeability and
transport properties. In addition, the potentially increased concentrations ofgp120 and
gp41
on the surface of vpuDEL-1-infected T lymphocytes could contribute to the morerapid killing of otherCD4+
cells in the culture by cell fusion.The evolution of small viral proteins like vpu may not be unique to HIV-1, since a number of other RNA viruses encode proteins that appear to functionduring final steps of virion production and show similarity to vpu. For example,
the size, hydropathicity, and domain structure of the mem-brane-associated influenza virus M2protein (15, 26) andthe
foot-and-mouth disease virus protein p3A (5) are strikingly similar to those of vpu (Fig. 10). Ahydrophobic N-terminal
anchor is followed by a very hydrophobic C terminus, with
at least 18 charged residues in each case. Other possible
homologs of vpu include the small NB protein of influenza B virus (25) and the SH protein of simian virus 5 (11). These proteins have been suggested to beinvolvedin the processof virus assembly (11, 25, 26). In contrast, neither HIV-2 nor SIV encodes a structural homolog of vpu (10, 12). It remains to be shown whether these two primate lentiviruses utilize another viral protein to functionally replace vpu.
Alterna-tively, a putative host cellprotein
may serve as a virus release factor in HIV-2 or SIV infections and obviate the requirement for a virus-encoded vpu-like protein.The intracytoplasmic production of virus particles charac-teristic of a wild type HIV-1 infection of monocytes-macro-phages (6, 7, 17) is reminiscent
of
the vpuDEL-1 infection of T cells described above. What is puzzling, however, is that HIV-1 andHIV-2 both produce intracytoplasmic virus par-ticles in infected monocytes even though only HIV-1 has the capacity to encode vpu. Possible explanations include (i) the presence of an inhibitor of vpu in monocytes-macrophages or (ii) the fact that vpu function in human T cells requires the synthesis of or the interaction with a cellular cofactor that is not expressed in monocytes. Experiments are in progress to more fullydissectthe role of vpu in both cell systems and to develop antagonists that specifically abrogate vpu function during the acute infection of T lymphocytes.ACKNOWLEDGMENTS
We thank Charles Buckler for computer analyses and Ronald Willey for valuable discussions.
T.K. was supported by afellowship from the Deutsche Forschungs-gemeinschaft, Bonn, Federal Republic of Germany.
LITERATURE CITED
1. Adachi, A., H. E. Gendelman, S. Koenig, T. Folks, R.Willey,A. Rabson, and M. A. Martin. 1986.Productionof acquired immu-nodeficiency syndrome-associated retrovirus in human and non-human cells transfected with an infectious molecular clone. J. Virol. 59:284-291.
2. Benn, S., R. Rutledge, T. Folks, J. Gold, L. Baker, J. McCor-mick, P. Feorino, P. Piot, T. Quinn, and M. Martin. 1985. Genomic heterogeneity of AIDS retroviral isolatesfromNorth America and Zaire. Science 230:949-951.
3. Cohen, E. A., E. F. Terwilliger, J. G. Sodroski, and W. A. Haseltine. 1988. Identification of a protein encoded by the vpu gene of HIV-1. Nature (London) 334:532-534.
4. Folks,T., S. Benn, A. Rabson, T. Theodore, M. D. Hoggan, M. Martin, M. Lightfoote, and K. Sell. 1985. Characterization of a continuous T-cell line susceptible to the cytopathic effects of the acquired immunodeficiency syndrome (AIDS)-associated retro-virus. Proc.Natl. Acad. Sci. USA 82:4539-4543.
5. Forss, S., K. Strebel, E. Beck, and H.Schaller.1984. Nucleotide sequence and genome organization of foot-and-mouth disease virus. Nucleic Acids Res. 12:6587-6601.
6. Gartner, S., P. Markovits, D. M. Markovitz, M. H. Kaplan, R. C. Gallo, and M. Popovic. 1986. The role of mononuclear J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.68.306.56.355.2]phagocytes in HTLV-III/LAV infection. Science 233:215-219. 7. Gendelman, H. E., J. M.Orenstein, M. A. Mirtin, C. Ferrua, R.
Mitra, T. Phipps, L. A. Wahl, H. C. Lane, A. S. Fauci, D. E. Burke, D.Skillman, and M. S. Meltzer. 1988. Efficient isolation and propagation ofhuman immunodeficiency virus on recombi-nant colony-stimulating factor 1-treated monocytes. J. Exp. Med.167:1428-1441.
8. Goff, S., P. Traktman, and D. Baltimore. 1981. Isolation and properties of Moloney murine leukemia virus mutants: use of a rapid assay for release of virion reverse transcriptase. J. Virol. 38:239-248.
9. Graham, F. L., andA.J. van der Eb. 1973.Anewtechnique for the assayofinfectivity of human adenovirus 5 DNA. Virology 52:456-457.
10. Guyader, M., M.Emerman, P. Sonigo, F. Clavel, L. Montagnier, and M.Alizon. 1987. Genome organization and transactivation of the human immunodeficiency virus type 2. Nature(London) 326:662-669.
11. Hiebert, S. W., C. D. Richardson, and R. A. Lamb. 1988. Cell surface expression and orientation in membranes of the 44-amino-acid SHprotein of simian virus5.J. Virol.62:2347-2357. 12. Hirsch, V., N. Riedel, and J. I. Mullins. 1987. The genome organization of STLV-3 is similar to that of the AIDS virus exceptforatruncated transmembraneprotein. Cell 49:307-319. 13. Kyte, J., and R. F. Doolittle. 1982. A simple method for displaying the hydropathic character ofaprotein. J. Mol. Biol. 157:105-132.
14. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (London) 227:680-685.
15. Lamb, R. A., S. L. Zebedee, and C. D. Richardson. 1985. Influenza virus M2 protein is an integral membrane protein expressedontheinfected cell surface. Cell 40:627-633. 16. Matsuda, Z., M. J. Chou, M. Matsuda, J. H. Huang, Y. M.
Chen, R. Redfield,K. Mayer,M. Essex,and T. H. Lee. 1988. Humanimmunodeficiency virus type1 has anadditionalcoding sequence inthecentral region of the genome.Proc.Natl.Acad. Sci. USA 85:6968-6972.
17. Orenstein,J. M., M. S. Melzer, T. Phipps, and H. E. Gendel-man. 1988. Cytoplasmic assembly and accumulation of human immunodeficiency virus types 1 and 2 in recombinant human colony-stimulatingfactor-i-treatedhumanmonocytes:an ultra-structural study. J. Virol.62:2578-2586.
18. Raff, M. C., K. L. Fields, S.-I. Hakomori, R. Mirsky, R. M. Pruss, andJ. Winter. 1979. Cell-type-specific markers for dis-tinguishing and studying neurons and the major classes of glial cells inculture. Brain Res. 174:283-308.
19. Sanger, F., S. Nicklen, andA.R.Coulson. 1977. DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463-5467.
20. Simionescu, N., and M. Simionescu. 1976. Galloylglucoses of lowmolecular weight as mordant in electronmicroscopy.J.Cell Biol. 70:608-621.
21. Symington,J.,U.Green,and K.Brackmann.1981. Immunoau-toradiographic detection of proteins afterelectrophoretic trans-fer from gels to diazo-paper: analysis of adenovirus encoded proteins. Proc. Natl. Acad. Sci. USA 78:177-181.
22. Strebel,K.,T.Klimkait,F.Maldarelli,andM.A.Martin.1989. Molecular and biochemical analysis of human immunodefi-ciency virus type1vpuprotein. J. Virol. 63:3784-3791. 23. Strebel,K.,T.Klimkait,and M. A.Martin. 1988.Anovel gene
of HIV-1, vpu, and its 16 kilodalton product. Science 241: 1221-1223.
24. Wiley,R.L.,D.H.Smith,L.A.Lasky,T.S.Theodore, P.L.
Eitrl,B. Moss,D.J. Capon, and M. A.Martin. 1988. In vitro mutagenesis identifiesaregion within the envelope gene of the humanimmunodeficiency virus that is critical for infectivity. J. Virol.62:139-147.
25. Williams, M. A.,and R. A. Lamb. 1986. Determination of the orientation ofanintegral membraneprotein and sites of glyco-sylation by oligonucleotide-directed mutagenesis: influenzaB virus NBglycoprotein lacksacleavablesignalsequenceand has anextracellularNH2-terminalregion. Mol. Cell. Biol. 6:4317-4328.
26. Zebedee, S. L., and R. A. Lamb. 1988. InfluenzaAvirus M2 protein: monoclonal antibody restriction of virus growth and detection of M2 in virions. J.Virol. 88:2762-2772.