JOURNALOFVIROLOGY,Aug. 1993,p. 4665-4675 0022-538X/93/084665-11$02.00/0
Copyright © 1993,American Society for Microbiology
Two
Distinct
Proteinase Activities Required for the
Processing of
a
Putative
Nonstructural Precursor Protein of
Hepatitis C Virus
MAKOTOHIJIKATA,1 HIROTOMIZUSHIMA,2 TSUYOSHI AKAGI,1 SHIGEHISA MORI,'
NOBUKO KAKIUCHI,1 NOBUYUKI KATO,' TORAHIKO TANAKA,1 KOICHI KIMURA,2
ANDKUNITADASHIMOTOHNO1*
Virology Division, National Cancer Center ResearchInstitute, 5-1-1,
Tsukiji,
Chuo-ku, Tokyo 104,1andDepartmentofAppliedBiological Science, Faculty of Science and Technology,
Science University of Tokyo, 2641, Yamazaki, Noda 278,2 Japan
Received 31 December 1992/Accepted 10 May 1993
Geneproducts of hepatitis C virus (HCV),apossiblemajorcausativeagentofposttransfusion non-A, non-B hepatitis,areconsidered tobe produced fromaprecursorpolyprotein viaproteolyticprocessing mediated by
either host cellorviralproteinases.Thepresence of HCV serineproteinase has beenproposedfromanalyses
of amino acidsequence homology. To examine theprocessing mechanism of the HCVprecursorpolyprotein, the amino-terminal region of the putative nonstructural protein region of the HCVgenome, containing the serineproteinasemotif,wasexpressed and analyzed by usinganin vitrotranscription/translation systemand a transient expression system in cultured cells. Two distinct proteinase activities which function in the production ofa70-kDaprotein (p70) from theprecursorpolyproteinweredetected. One of theseproteinase
activities, which cleaved the carboxyl(C)-terminalside ofp70,required thepresenceofthe serineproteinase motif, which is located in the amino(N)-terminal region of p70. That suggested that the predicted HCV serine proteinasewasfunctional. The otheractivity,whichwasresponsible for the cleavage of the N-terminal side of
p70,required the expression of the regionupstreamand downstream of thatcleavage site,includingthep70 serineproteinase domain. From the results ofpulse-chase analysis, using proteinase inhibitors coupled witha
point mutation analysis, the latter activitywasproposedtobe anovelzinc-dependent metalloproteinase. Infection with thehepatitis C virus (HCV) is reported to
beclosely related toposttransfusion non-A, non-B hepatitis (10, 27). The HCV genome has recently been cloned and sequenced (11, 23, 32, 33, 38, 39). Thepositive-strand RNA genomeofthis virus iscomposed of about 9,400 nucleotides andincludesasingle largeopenreading frame(ORF) encod-ing a polypeptide of 3010, 3011, or 3033 amino acid (aa) residues (11, 23, 32, 33, 38, 39). Analyses of amino acid sequencehomology have revealed that HCV is relatedtothe flaviviruses and the pestiviruses (10, 22, 23, 29, 38), suggest-ing that the genomicorganization of HCV is homologousto those of these two related viruses. Therefore, functional HCVgeneproductsseem tobeproduced by both cotrans-lational and posttranslational processing of the large poly-proteinprecursor,asin thecasesoftheserelated viruses (for
a review, see reference 35). Using an invitro processing system,wehave shown that threeprocessedproducts of the
putative viral structural proteins, p22, gp35, and gp7O, are
initially cotranslationally produced from the N-terminal
re-gion of the HCV precursor polyprotein, possibly byahost
cellsignal peptidase (18). However, the processing mecha-nism of the C-terminal region of the precursor polyprotein hasnotbeenclarified. This latter region, which constitutes about 75% of the HCV ORF 3' terminus, is suggested to encode thenonstructural(NS)proteins thatareessential for
replication of the HCVgenome;itcontains conserved amino acidsequencemotifs ofachymotrypsin-likeserine
protein-ase, RNAhelicase, and RNA-dependent RNApolymerase. Thesequence motifsarealsofound insequences of
flavivi-*Correspondingauthor.
rus andpestivirus NS proteins (22, 23, 29). Recently,
puta-tiveviral serineproteinaseswereshowntobefunctional in theproteolytic processing of viralprecursorpolyproteinsby invitro and in vivoexperiments. This is thecasein theNS proteins of yellow fever virus and denguevirus type 2 and type 4 (DEN2 and DEN4), all members of the Flavivirus genus, and it is also true forbovine viral diarrheavirus, a
pestivirus (5, 14, 34, 43); however, the predicted RNA helicase and RNApolymerase activitieswere notdetected. Thesedatastronglysuggestthatthestructurehomologousto serine proteinase found in the HCV ORF has enzymatic activity and isresponsible for processing the primary HCV geneproducts.
To determine the molecular mechanism of HCV viral protein production and gene organization, cloned cDNA fragments corresponding to the middle of the genome, in which theputative serine proteinase motif is located, were
expressedby using anin vitrotranscription/translation
sys-tem and in cultured cells by using a transient expression
system. Two distinctproteinase activities functioning in the production ofa70-kDapolypeptide (p7O)weredetected.p70 waspresumablyanHCVhomolog of the flaviviral NS3 and
the 80-kDa protein (p80) encoded by the bovine viral diar-rhea virusgenome,which hasaserineproteinase domain (2).
One oftheproteinase activities,whichcleavedthe C-termi-nal end of p70, required the alignment of predicted serine proteinase structure in the N-terminal portion of p70. The other, which cleavedthep70Nterminus,isproposedtobe zinc dependent and is required the polypeptide region
up-streamand downstream of thecleavagesite,whichincludes thecomplete serineproteinasemotif.
4665
Vol. 67,No. 8
on November 9, 2019 by guest
http://jvi.asm.org/
4666 HIJIKATA ET AL.
5' 3'
U"4Vnnma
.nu, wvviuei < ORF (9030nts, 3010 aa) o
5 1,0 1,5 2,0 25
3,0
XIC a..a. ,precursor.%Oj#vnl2Cmn7h 'polyprotein
p=zz p3o gpfU
/
Be RSE5Bsp A BhS.No
1 I 1i -. f w
I10 1 5
I
gp70
m IHDS
x102a.a.
N340
C1
C2 C3 C4
C5
- C6
[image:2.612.72.284.69.341.2]N340-1325 N722-1908 N722-1647 N722-1466 N722-1325 N722-1233 N722-1174 N722-1019 N898-1908 N992-1908 N1117-1908 N898-1325 N992-1325 FIG. 1. Constructions ofcDNAfragments .usedininvitro
tran-scription/translation experiments and regions encoded by these
constructs in the HCV ORF. HCV polyprotein precursors are
boxed. The p22, gp35, and gp70 are putative initial processing productsofthe structural protein region ofHCVORF(18). H, D, and Sindicate Hisataa1083, Aspataa1107,andSerataa1165of
theHCVprecursorprotein.These residuesarepredictedtoform the catalytic triad of the chymotrypsin-like serine proteinase, and a
highly homologous sequence motif has been found in the HCV
precursor polyprotein indicated by the shaded box. The cDNA fragmentsofN340, Cl, C2, C3, C4, C5, and C6werefrompN340
(18), clones 6,7, 8, and 9 (23),andclones 6and 18(39), respectively. The locations of BssHII(Bs),RsrII(R), SaclI (S),AvrII (A), Eco52I (E5), SphI (Sp),BamHI(Bh), SacI (Sa),and NsiI(Ns)restriction sitesareshownonthe scale for the amino acidposition.
MATERIALSANDMETHODS
Construction ofan in vitroexpression plasmid. A cDNA fragment covering the HCV ORF region from aapositions 722to 1908was constructedwith six overlappingcDNAs, Cl,C2, C3, C4,C5,andC6(Fig. 1). These cDNAfragments werefrom clones 6, 7, 8, and 9(23) and clones 6 and 18
(39),
respectively. A translation initiation codon in the5'-terminal end of the Cl fragment was constructed by inserting the 12-mer NcoI linker (5'-CAGCCATGGCTG-3') (Takara Shuzo) into the EcoRI site of pCl, which isarecombinantplasmid carrying the Cl fragment intheHincII site ofthe multicloning region of pTZ18Uvector(U.S. Biochemicals), afterfillinginthe cohesive endsof the restriction siteswith T4DNApolymerase. The 5'-terminal end of the ORF of the insert in the resultant clone, named pC1N, encoded four extra amino acids (methionine, alanine, glutamic acid, and
phenylalanine). The EcoRI-SphI fragment of the C2 frag-mentand theSphI-BamHI fragment of the C3fragmentwere subcloned into theEcoRI-BamHI site of the pTZ18Uvector
simultaneouslytoobtain the pC2-3 clone. Whennoadequate
restriction enzyme site was present in the overlapping
re-gion,weused thepolymerase chain reaction method for the
ligation
ofthetwo overlapping fragments (21).Theprimers
used for the
ligation
reactions were as follows: 5'-TITTG TACGGGCTCAGGGGCT-3', 5'-TCCATGTCAGAGAAG ACGAC-3', 5'-GCGGTGGCAGTAGAGCCCGTC-3', and5'-ACCCCTGCTCTCCAAAACTA-3' forligationofCl and C2 togivetheC1-2fragment; 5'-GTCGTCTTCTCTGACAT GGA-3',
5'-TGGTAGTCGAGTCAGTTGAGT-3',
5'-ACTCAACTGACTCGACTACCA-3', and 5'-TCAGCGGGCGTG AGCTCATA-3' for ligation of C2-3 and C4 to give the
C2-4 fragment; 5'-ACATGTGTCACCCAGACAGT-3',
5'-TCAGCGGGCGTGAGCTCATA-3', 5'-ATGACGCAGGCT GTGCIlIGG-3', and 5'-CTGCATTCCTTGCTCGATGT-3'
for ligation of C4 and C5 to obtain the C4-5 fragment;
5'-ATGACGCAGGCTGTGCTTGG-3',
5'-CTGCATTCCTT GCTCGATGT-3',5'-GTTCGATGAGATGGAAGAGT-3',
and
5'-CATCCACTGCACAGCCCCCTC-3'
for ligation of C5 and C6to obtain the C5-6fragment. The SphI-HindIII fragment of pClN was replaced by the SphI-Eco52Ifrag-ment ofC1-2 and theEcoS2I-HindIII fragment of the pC2
insert toyield pC1-2N. pC1-3Nwas obtained byreplacing
the BssHII-HindIII fragment of pCl-2N with the
BssHII-HindIIIfragment of pC2-3. TheBssHII-BamHIfragmentof
pC1-3N was replaced by the BssHII-BamHI fragment of
pC2-4toyield pCl-4N.TheBamHI-SacIfragment ofpC4-5
and theSacI-SmaI fragment of pC5-6wereinsertedbetween BamHI and the blunt-ended PstI sites ofpC1-4N toyield pC1-6N. The insert cDNA of pCl-6N encodes the amino acid sequencecorrespondingtotheregion fromaa722to aa
1908 of the HCV precursorprotein, which is thus termed
pHCN722-1908. The pCl-3Nfragmentwasinserted intothe
RsrII-HindIII site of pN340 (18)toyieldpHCN340-1325,the
insert of which encodes theregion from the putativesecond
HCVenvelope protein(gp7O) regiontothe serine proteinase
motif inthe HCV precursor polyprotein.
Construction of the deletion mutants and point mutants. Four 5'-terminal deletion mutants, pHCN729-1908, pHC
N898-1908, pHCN992-1908,andpHCN1117-1908,were con-structed frompHCN722-1908 bydeletingthe EcoRI-BssHII
fragment, theEcoRI-RsrII fragment, theEcoRI-SacIl
frag-ment, and the EcoR-BssHIIfragment, respectively, andby ligating with 12-mer (CAGCCATGGCTG [for
pHCN729-1908,pHCN898-1908,andpHCN1117-1908])and 8-mer(GC
CATGGC [for
pHCN992-19081)
NcoI linkers afterfilling in the reaction as described above. The 3'-terminal deletion mutants of pHCN722-1908 were constructed as follows.pHCN722-1647 was constructed by using the SacI-NsiI
fragmentofpC5-6instead of theSacI-SmaI fragmentduring
the construction ofpC1-6N. 1466 and
pHCN722-1325 werepCl4NandpCl-3N, respectively,intermediates
during construction of pHCN722-1908. pHCN722-1233, pHCN722-1174, and pHCN722-1019 were constructed by deleting the 3'-terminalportionofpHCN722-1325with exo-nuclease III after cutting the XbaI and PstI sites of the plasmid (17). To obtain pHCN898-1325 and pHCN992-1325, the AvrII-HindIII fragments of pHCN898-1908 and pHCN992-1908 werereplaced by theAvrII-HindIIIfragment ofpHCN722-1325.
The single-amino-acid mutations were introduced into
pHCN722-1908 by site-directed mutagenesis methods (21, 26). The mutation primers used, the name of the resultant
clones, and the amino acid substitutions were as follows:
5'-CACCAGCGTAGACGGTCCAA-3'wasused for produc-tion of pH1083A (His-1083 to Ala-1083); 5'-GACGAGGT
TCTGGTCTACAT-3'wasused forproductionofpD1107N (Asp-1107 to Asn-1107); and 5'-GTCCACCCGCGGAGCC CTTC-3' wasused forproduction ofpS1165A(Ser-1165 to J.VIROL.
ft,
on November 9, 2019 by guest
http://jvi.asm.org/
TWO PROTEINASES IN PROCESSING OF HCV POLYPROTEIN 4667
Ala-1165). The series of mutants pH932A, pH952A, pH1083A, pH1136A, pH1175A, pH1227A, and pH1229A,in
which a single histidine residue at a numbered amino acid position was changed to an alanine residue, were produced byusingthe primers5'-ATITGGACATAGGCGCCTCC-3',
5'-TGGAGTAAGAGCGTCATATAC-3', 5'-CACCAGCGT
AGACGGTCCA-3', 5'-GACATCGGCAGCCCTCGTGA-3',
5'-ATGCCTACAACGGCCCCCGA-3', 5'-AGCGTGTAAA
GCTGCCACTlTG-3, and 5'-AGTGGGAGCGGCTAAATG TG-3', respectively. The series of mutants pE972Q, pE980Q, pE1009Q, pE1058Q, pE1199Q, and pE1202Q, in which a single glutamic acid residue at anumbered amino acid position was changed to a glutamine residue, were produced by using the primers 5'-AAGACGACGGGCTG TACTGC-3', 5'-GTTlAGTCTGCATGTCAGAG-3', 5'-GA
AGTATCTGCTTCCCCCTT-3', 5'-ACCTGAACCTGCCC
ATCGAC-3', 5'-TCCATAGACTGAACGGGTAT-3', and
5'-ATGGTAGTITGCATAGACTC-3',
respectively.These-ries of mutants pC922A, pC993A, pC1123A, pC1125A,
pC1171A,andpC1185A, in whichasingle cysteine residueat anumbered amino acid position was changed to an alanine
residue, were produced by using the primers 5'-ATCAA CATGGCTGCACGGATG-3', 5'-ATGATGTCCCCAGCCG CCGC-3', 5'-CCGCAGGTGGCCGGTGTCAT-3',5'-GAGC
TGCCGGCGGTGGCCGGT-3', 5'-AAGGGGCAAGCAGT GGTCCA-3', and 5'-GGTGGCAGCAGCAGCCCGGAAG AT-3',respectively. The substituted nucleotidesequencesof
all of the clonesobtainedwereverified bysequencing.
Construction of in vivoexpression plasmids.Forexpression of HCV proteins in vivo, a eukaryotic cell expression plasmid bearing the enhancer and promoter ofthe human
cytomegalovirus immediate early gene was constructed as
follows. The 8-mer BamHI-linker was inserted into the
HindIlI site of pCMVcat (15)afterHindIIIdigestion and a
filling-in reaction. The
XbaI-BamHI
fragment, containingthe human cytomegalovirus enhancer-promoter, was in-serted into the
XbaI-BamHI
sites of pBluescript II KS(+) plasmid (Stratagene), and then theHincII-XhoI
fragment of the multicloning site of the plasmid was replaced by aBamHI-SalI fragment containing the simian virus40 polyad-enylation signal taken from pSVL (Pharmacia) after
filling
inofthe BamHI site. The resulting expression plasmid was named pKS(+)/CMV. The EcoRI-HindIII fragments of pHCN729-1908, pS1165A, and pH952A were inserted into theEcoRI-HindIIIsite ofpKS(+)/CMVtoconstructpCMV/ HCN729-1908,
pCMV/S1165A,
and pCMV/H952A,respec-tively.
Antibody. Apolyclonal antibodytothe HCV
p70
product was raised in rabbits byimmunization with the HCV viral protein 07, which was expressed in a fused form with3-galactosidase
inEscherichia
coli(31)
andwaspurified
by
sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE)
andelectrophoretic
elution.Invitrotranscription/translationandimmunoprecipitation. In vitro transcription/translation was performed with T7 RNA polymerase, a rabbit reticulocyte lysate, a canine
pancreatic microsomal membrane fraction (Promega), and
[35SJmethionine
(Amersham), as describedpreviously (18).All invitroexpression plasmidswerelinearizedby cleavage
of the unique HindIII site, located at the 3' ends of the
inserts,beforebeingused in in vitrotranscription. After in vitro translation at 30°C for 90 min, part of the translation product was treated with proteinase K (200 ,ug/ml) to deter-mine the location of the product in the lysate containing microsomal membranes, as described previously (18). In somecases, thetranslationproductwassuspended in 1 ml of
50 mM Tris (pH
7.2)-150
mM NaCl-1% Triton X-100-1% sodium deoxycholate-0.1% SDS (RIPA buffer), and the suspension was incubated with anti-p70 antibodies for 1 h. The immune complex was precipitated with Pansorbin (Cal-biochem). Theimmunoprecipitateswaswashed three timeswith RIPA buffer and suspended in 2x sample buffer(28).
The invitro translation productswere analyzed directlyor after immunoprecipitation by SDS-PAGE (19, 28, 36) and
thenby fluorography.
Proteinase inhibitorstudy. The effectsofvarious
protein-ase inhibitors were studied by their addition tothein vitro
translation mixture during the chase period ofpulse-chase analysis.Thein vitrotranscripts from pHCN722-1325were
translatedinvitroduringapulseperiod of 30 minat30°C in
the presence of
[35S]methionine.
The chase period was startedby theaddition ofcycloheximide (1 mg/ml) and cold methionine (1mM)fortermination ofthetranslation reaction in the presence or absence of a proteinase inhibitor. The proteinase inhibitors used were aprotinin (10 ,ug/ml), [(4-amidinophenyl)-methane sulfonyl fluoride] (APMSF) (0.2 mg/ml), E64 (1 mg/ml), bestatin (50 ,ug/ml), pepstatin (0.7 ,ug/ml), EDTA (2 mM), antipain (0.2 mg/ml), chymostatin (0.2mg/ml),
andphosphoramidon (0.2 mg/ml). All of these proteinase inhibitors were purchased from Boehringer Mannheim. Theproteinase(Cpro-1) activity
was measuredas anincrease in theamountofp21
radioactivity during
the chaseperiod byusing animaging
plate and BAS2000(Fuji
Photo Film Co.) (1), because the production of p21 is dependenton
Cpro-1
activityandbecause itwas oneofthe moststableof the invitro translationproducts of theHCV ORF(seeResults).
Transient expression assay and immunoblotting. COS-1 cells and Huh-7 cells were transfected by the calcium phosphate coprecipitation method (7). Samples of the
ex-pressionplasmids(10
pLg)
weretransfectedinto4 x105
cells in60-mmdishes. Cellswereharvested 48 h aftertransfection and resuspended in 2x sample buffer (28). Samples werefractionated on SDS-11% PAGE, and the proteins were
transferred electrophoretically to polyvinylidene difluoride
membranes(Millipore; Immobilon). After incubation for1h
at 25°C in 150 mM sodium phosphate (pH 7.2) with 3% bovine serum albumin for blocking, the membranes were incubated in the same buffercontaining
anti-p70
antibodies for2h at25°Candthen washedextensivelywith phosphate-buffered salinecontaining 0.05%Tween 20. Theantibodiesbound to the membranes were detected with biotinylated
goat anti-rabbit immunoglobulin G antibody and a
Vecta-stain ABC kit (Vector Laboratory) under the conditions specified by the supplier.
Nucleotide sequence accession number. The sequence of
the cDNA of pHCN722-1908 has been deposited in the
DDBJIEMBIVGenBank
DNA data bases under accessionnumber D11397.
RESULTS
Detection ofviral protein processingin in vitro transcrip-tion/translation products. To detect HCV polyprotein
pro-cessing and processed viral products, the
polypeptide
of HCV precursor protein from residue 340 to residue 1325(N340-1325) (Fig. 1),wasexpressedinanin vitro
transcrip-tion/translation system withpHCN340-1325asthe
template
for the transcription. This region of the precursor
protein
includes the amino acid sequences of the
putative
HCVenvelope protein, gp7O (18), and the
predicted
serine pro-teinase.Previously,
two products,gp7O
andp19,
were VOL.67, 1993on November 9, 2019 by guest
http://jvi.asm.org/
4668 HIJIKATA ET AL.
N340-1325
MM - + +
PK - - +
Mr S P S P S P
X103
200-11 p6w -ori
115-
98-
66-
45-30- t
20-VW ,o -..gp7O
*-t --"*-p55p
tfZ
---p33_-*-p21
a b c d e f
FIG. 2. Detection of proteinase activitycleavesinvitro transla-tion product. RNA transcribed from pN340-1325 in vitro was
translated in the absence(lanes aandb)orpresence(lanesc,d,e,
and f) of microsomal membranes (MM). After translation in the
presence ofmicrosomal membranes, the translate was incubated
with proteinase K(PK) (lanef).Thesupernatant(S) (lanesa, c,and
e) and particulate (P) (lanes b, d, and f) fractions of reaction mixtures obtained bycentrifugationwereanalyzed. Theamountof
supernatantanalyzedwasequivalent toone-fifth of theparticulate fraction.Thepositionof theoriginontheSDS-polyacrylamide gel is
indicated byori.Molecularsize markersareshownontheleft.
produced in a microsomal membrane-dependent manner
from the region spanningresidues 340to980(pN340) (18). In addition to gp70, three other polypeptides with molecular
masses of 55, 33, and 21 kDa made by N340-1325 in the presence of microsomal membranes (Fig. 2, lane d) were
detected, andwenamed these products p55, p33, and p21,
respectively. Product p55wasrevealedtobeaprecursorof
p33 and p21 by deletionmutantanalysis (see Fig. 4A, laneh) and by pulse-chase analysis (see Fig. 6A). Since gp7O is already known tobe produced from the N-terminal half of this polypeptide (18), p21 and p33werelikelytobeproduced from its C-terminal half. Production of p33 occurredevenin the absence ofmicrosomal membranes (Fig. 2, lanesa and
b). This result suggested the possibility that the translated polypeptide bears the proteinase activity which excises p33 from the precursor polypeptide, because no well-known
site-specific proteinase activitywas present in the reticulo-cyte lysate. It is supposed that p33 is located in the C-ter-minal regionof the translatedpolypeptide. The production of p21isknown torequiremicrosomal membranes. Therefore, if p33 islocatedonthe N-terminal side of p21, it would also
be expected to require microsomal membranes for its pro-duction. Theproduction of p33 in the reaction in the absence of microsomal membranes suggested that these three
pro-cessed products were likely to be arranged as follows:
NH2-gp7O-p21-p33-COOH. Theorder of thearrangement of these productswasfurtherconfirmed by deletion analysisas
described below. Translation products produced in the
re-action in the absence of microsomal membranes seemed to
be stacked at the origin of the polyacrylamide gel, as
previously reported (18) (Fig. 2, lane b). Unprocessed and unglycosylated polypeptides, including the gp7O region,
es-pecially its C-terminal hydrophobic region, were difficult to separate by the gel electrophoresis systems used in our
experiment(datanotshown).The weak p33signalin laneb of Fig. 2 may have been caused by the coexistence ofan aberrant form of unprocessed gp7Oregion.Suchadifference
was not observed in the cleavage reaction products of N722-1325, withmostofthe gp7Oregion deleted, asshown below(seeFig. 4A,lanes gandh).
The production ofp21 required the presence of
microso-mal membranes(Fig.2, laned).Cleavageof the region next to the gp7O C terminus yielded p19 (18), which was a C-terminal truncated form of p21. The supematant and
particulate fractionsof the translation reaction mixture con-tained p33; however,p21, like gp70, was present in only the
particulate fraction. Resistance toproteinase Kadded after the translation reaction (Fig. 2, lane f) suggested that p21
associated withmicrosomal membranes. As reported
previ-ously, p19 wassensitivetoproteinase K treatment underthe sameexperimentalconditions (18). Several other C-terminal deletion mutants of p21 showed different sensitivities to proteinase Kdigestion under the same experimental
condi-tions (data not shown). Proteins located in the lumen of microsomal membranes are known to be fully resistant to proteinase K treatment. The results obtained in the present
study, together with the above-mentionedunpublisheddata,
imply thatp21 is not located in thelumen of the microsomal membranes. More probably, the intact form of p21 may be buried in the lipid bilayer of microsomal membranes, in which it would be protected from proteinase Kdigestion.
Gene mapping of the N-terminalportion ofthe NS protein region of the HCVORF. Toassess the molecular nature of HCVproteinase,residues 722 to 1908 of the HCV precursor polyprotein and its N-terminal and C-terminal deletion mu-tants wereexpressed in an in vitro transcription/translation system. The template plasmids used for in vitro
transcrip-tion were pHCN722-1908, pHCN722-1647, pHCN722-1466,
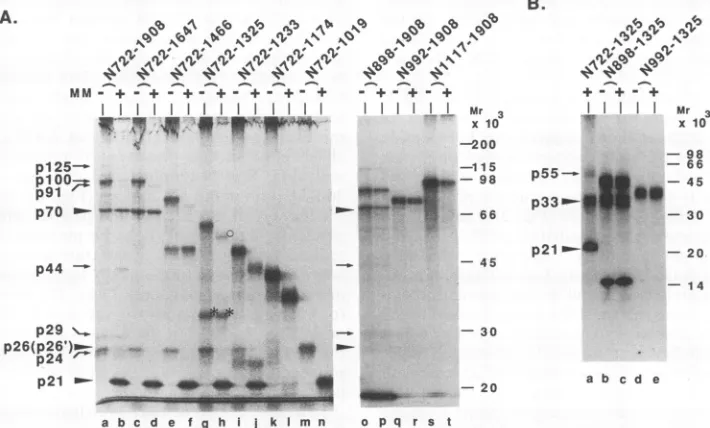
pHCN722-1325, pHCN722-1233, pHCN722-1174, pHCN 722-1019, pHCN898-1908, pHCN992-1908, and pHCN1117-1908 (Fig. 1). Two majorproducts withmolecular masses of 70 and 21 kDa and seven minor products with molecular masses of 125, 100,91, 44,29, 26, and 24 kDa were detected in thecleavage reaction products ofN722-1908 inthe pres-ence ofmicrosomal membranes and were named p70, p21,
p125, plOO, p91, p44, p29, p26, and p24, respectively (Fig.
3A, lane c). The products detected from this cleavage
reaction in the absenceofthe microsomal membraneswere the sameasthosedetected in thepresenceof the microsomal membranes, exceptthat p26 increased and p21 disappeared
(Fig. 3A, lane a). Theseobservationsindicated that the main proteolytic processing events in this reaction occurred in a microsomal membrane-independent manner. From the cal-culatedmolecular weightof the expressedpolypeptide, p125
seemed to be an intermediate that was processed in the
N-terminal small portion of the precursor only in the pres-ence ofmicrosomal membranes. A product slightly larger than p125,possibly theunprocessedprecursor, was found in thecleavage reaction products in the absence of microsomal membranes (Fig. 3A, lane a). All of the other minor
poly-peptides except p26 (shown in lanes a and cofFig. 3A) are likely torepresentintermediates, artificially processed
prod-ucts, or degradation products (25), because they decreased duringprolonged incubation. This contrasted withp21, p26,
and p70, which accumulated (data not shown) and were likely to be the processed products, although p26 was detected as afaint, smeared band. The precursorN722-1647,
which lacked the C-terminal 261 aa residues of the N722-1908 polypeptide, still yielded p21 and p70 after cleavage,
although p26 was not detected(Fig. 3B, lane c;Fig. 4A,lane J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.117.246.74.257.2]TWO PROTEINASES IN PROCESSING OF HCV POLYPROTEIN 4669
A. B. C.
N722-1 908
MM - +
Mr 3 T IP T IP
x10
200-11 5- _ .
8-
66-
45-I
aI
20- o
20-_
a b c d
pl25
*pl00
p91
-_p70 -p44
.,tp29
_..p26(p26')
p24
-._p21
N722-1647
MM - +
Mr3 T IP T IP
X
10,
200- 7 1
15-9 8
=
. pt9166- - p70
45-S1165A
MM
-Mr 3 T IP
x10
200-
115-98- rn
66-
45-
30-r _-.p26'
*'p24
20- _..Op21
3
0-
20-a b c d
I
a b
-T IP
.-pl25
..-p100
-.p26'
'*.p24 _.p2l
c d
FIG. 3. Detection of processedproducts of HCV polypeptides expressed in vitro byimmunoprecipitationanalysis. RNAs transcribedfrom pN722-1908 (A), pN722-1647 (B) and pS1165A (C)invitro weretranslated in the absence (lanesaandb) or presence (lanes c andd)of microsomalmembranes(MM). Translatedproducts(T) (lanesaandc)and productsimmunoprecipitated(IP)(lanes b andd) with antibody prepared against the polypeptide from residue 1308 to residue 1533 of HCV precursor(anti-p70 antibody) (see text) were analyzed by electrophoresison anSDS-11% polyacrylamidegel. The major processing products,p70,p26,andp21,areindicated byarrowheads.Proteins precipitated with the antibodyareidentifiedattheright. The possible processing intermediates or artificially processed products are indicated byarrows(see Results). The sizes of proteinmolecular mass markers are indicated at the left.
d). This result suggests that p26 originated from the C-ter-minal region of the N722-1908 polypeptide. The cleavage reaction products of N722-1647 lacked plOO, and p91 was
found asthelargest precursorprotein (Fig. 3B, lane c, and
Fig. 4A, lane d). These data suggested that plOO is an
intermediate form ofp70 and p26and that p91 is an
inter-mediatecomposedofp70andp21. The sizes of p21 and p70 were unchanged in the C-terminal deletion mutant,
N722-B. A.
A~4o%b~ 4b 41 %h 4A$
m
9m t
< 4
+-#
MM - +- - + - +" - + *t++-a?'
4"q
.I Mr 3
X 10
N'
sc+- +-9+9
Mr 3
v x 10
-200
p1 25-pl
0,07
w ** p70 0- - -_tIs,
.op44 - _
WVt
p29 _..
p26(p26')m-_ - @ _ t 7, * _
p24 r"
- 45
p21 0 _ _ _ Or -20
abcde .ijklh ml._...q-r
a b c d e f g h i j k I m n o p q r s t
FIG. 4. Genemapping oftheN-terminalregion of theputative NS protein-encoding region oftheHCVgenome.(A) Seriesof C-terminal (lanesc,d,e,f,g,h, i, j, k,1,m,and n)and N-terminal (laneso, p, q, r, s,andt)deletionmutant constructsof pN722-1908(lanesaandb)
wereusedastemplates for the in vitro transcription, and the resultant RNAsweretranslated in the absence (lanesa, c, e, g,i, k, and m)or
presence(lanes b, d, f, h, j,1, andn) of microsomal membranes (MM). (B) RNAs transcribed in vitro with the N-terminal deletionmutant constructsofpN722-1325 (lane a),pN898-1325 (lanes b and c), and pN992-1325 (lanes d and e)astemplatesweretranslated in the absence (lanesbandd)or presence(lanesa, c,and e) of microsomal membranes. Translation productswereanalyzed byelectrophoresison anSDS 11%-polyacrylamide gel (A) (28)or atricine-SDS-16% polyacrylamide gel (B) (19, 36). The products detected in the cleavage reaction of N722-1908in thepresenceof microsomal membranesaremarked byarrowsorarrowheads, with theirnames onthe left. ThepS5 and p33 shown inFig.2areindicatedbyanopencircle andasterisks, respectively.
-g8
-115As_ -66
on- -666 p3 - -30
p21 '-0 - 20
-14
- 30
a b c d e
VOL.67,1993
I
k
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.155.466.76.268.2] [image:5.612.132.487.425.639.2]4670 HIJIKATA ET AL.
1647
(Fig.
3B, lane c; Fig. 4A, lane d), supporting thispossibility.
The product ofthe 26-kDaprotein,
tentativelynamed
p26',
was detected incleavage
reactionproducts in the absence ofmicrosomalmembranes(Fig.
3A,lane a;Fig.
3B,
lanes a and c). The product p26' was possibly anN-terminal unprocessed form ofp21, because its decrease
occurred
simultaneously
with the appearance of p21 in reactions withmicrosomalmembranes(Fig.
3A,lane c;Fig.3B,
lanec),
and the molecular size ofp26'
wasnotaffectedby
the C-terminal deletions(Fig. 3B,
lanes a and c). The other C-terminaldeletions, N722-1466,
N722-1325, andN722-1233,
which lack theC-terminal 181, 322, and 414 aa residues of the N722-1647polypeptide,
respectively, re-sulted in decreased molecular sizes ofpossible
precursorpolypeptides,
from 91 kDato72,
55(corresponding
top55),
and45
kDa,
asexpected
(Fig. 4A,
lanesf, h,
andj).
IntheseC-terminal deletion mutants, the sizeofp70 alsodecreased
from 70 kDato
50,
33(corresponding
top33),
and 24 kDa,butthe
production
ofp21
was notaffected(Fig.
4A, lanes f,h,
andj).
These resultsindicated thatp70was located at aregion
downstreamofp21inthe HCV precursorpolyproteinand that
p33
is a C-terminal truncated form ofp70,
assuggested
above. Eachmolecularsize lost in the precursorpolypeptides
and derivatives ofp70by
the C-terminaldele-tions of the N722-1647
polypeptide
was in good agreement with that calculated from the deleted amino acid sequence of each deletionmutant.Therefore,
thecleavage
site betweenp70
andp26
(p70/p26)
seemed to be located at around aa residue 1650inthe HCVprecursorpolyprotein.
The cleav-agesiteofp21/p70
wasestimatedtobeataboutresidue1020 of the HCV precursorpolyprotein,
because a 21-kDapolypeptide
wasproduced
inthecleavage
reactionproducts
of N722-1019 in the presence of microsomal membranes
(Fig. 4A,
lanen).
Theproduct
withasize almost similarto that ofp26'
was observed in the translationproducts
of N722-1019 in the absence of microsomal membranes(Fig.
4A,
lanem).
From these results and calculations of the molecularweight,
theN-terminal end ofp21
was estimated to be located at about residue 800 of the HCV precursorpolyprotein.
The locations of these
processed
products
in the HCV precursorpolyprotein
werefurther assessedby
immunopre-cipitation
analysis
withantibody
against
thepolypeptide
fromaa1308to 1533 of the HCV precursor
polyprotein
(see
Materials and
Methods).
Themajor
product,
which wasimmunoprecipitated
from thecleavage
reactionproducts
of N722-1908by
thisantibody,
wasp70
(Fig. 3A,
lanesb andd).
Thepossible
processing
intermediates,
p125,
plO0,
andp91,
all of whichmight
includep70
sequence, were alsoimmunoprecipitated
(Fig. 3A,
laned).
Interestingly,
a prod-uctof the 26-kDaprotein
was found in thatimmunoprecip-itate
(Fig. 3A,
lanes b andd).
Since theantibody
usedin theimmunoprecipitation analysis
hadnocross-reactivity against
anyother precursor
polyprotein
region (18),
this resultmight
indicate that the 26-kDa
product
wasassociatedatleast withp70
in thereticulocyte
lysate
(see Discussion).
This 26-kDaproduct
wasnotdetected in theimmunoprecipitate
from thecleavage
reactionproducts
ofN722-1647,
which lacked the C-terminal 261 aa residues of the N722-1908polypeptide,
indicating
that p26 wasmapped
in the deleted C-terminalregion
of theN722-1908polypeptide
(Fig.
3B,lanes b andd).
Fromthese
results,
we concluded that themajor
processedproducts
from N722-1908 arep21, p70,
and p26 and that theseproducts
werearranged
inthe orderNH2-p21-p70-p26-COOH,
assummarized inFig.
8.Detection of the region required for HCV polyprotein
processing. Protein p21 was not produced in the cleavage
reactionproducts of N722-1174, althoughatranslation
prod-uctwithamolecularmassof 44 kDathatwasprocessedtoa 39-kDaproductin amicrosomalmembrane-dependent man-nerwasdetected(Fig.4A, lanes k and 1). This resultimplied
that processing of the p21/p7O site did not occur in this translatedpolypeptide. Inthecleavagereactionproductsof N898-1908,oneoftheN-terminal deletionmutantsof N722-1908 (Fig. 1) and all of the cleavage reaction products of N722-1908, except p21, were detected in the absence of membranes(Fig. 4A,lanesoandp). Thus, cleavagesatboth the p21/p70 and p70/p26 sites occurred independently of microsomal membranes. However, the cleavage reaction
products of N992-1908, both in the absence and in the presence of microsomal membranes, did not contain p70.
However, a product with a slightly larger size was found (Fig. 4A, lanes q and r). A faint bandmigrating at nearly the same position as p70, which may be a degradationproduct, wasoccasionally found(Fig. 4A, lane q). Since the unproc-essed 102-kDa polypeptide was barely detected, and since p26 was found intheN992-1908 cleavage reaction products, cleavage of thep7O/p26 site apparently occurred.Therefore, it was concluded that cleavage of the p21/p7O site did not occurin theN992-1908 polypeptide and that consequently a productcomposed of the C-terminal portion of p21 plus p70 migrated slightly slower, to a position of 70 kDa. Further-more, only asingle unprocessed polypeptide of the 90-kDa protein was found in the N1117-1908 cleavage reaction products, in which half of the serine proteinase motifwas deleted (Fig. 4A, lanes s and t), which suggested that the serineproteinase motif present in the HCV ORF was func-tional and was required at least forp7O/p26 site cleavage.
The loss of the p21/p70 site cleavage in this N-terminal deletion mutantwas further supported by the results from translations of the N898-1325 and N992-1325 polypeptides, whichareN-terminal deletion mutants of N722-1325(Fig. 1). Since p33 and a 15-kDa protein were produced from a 47-kDa precursorpolypeptide, the cleavage at the p21/p70 site of the N898-1325 polypeptide apparentlyoccurred in a microsomal membrane-independent manner (Fig. 4B, lanes b and c). However, only a single unprocessed 36-kDa translationproductwasfound, and p33was notproduced in the cleavage reaction products of N992-1325 either in the absence or in the presence of microsomal membranes,
indicating that N-terminal end cleavage of p33 from the 36-kDaprotein did not happen (Fig. 4B, lanes d and e).
Cleavage of the p21/p70andp70/p26sites occurred
inde-pendentlyand was likely to be mediated by the viral pro-teinase activity present in the translated polypeptide itself; however,cleavageof the p21 N terminuswasonly microso-mal-membrane dependent. Thus, the processing activities forcleavage of the p21/p70 site and the p70/p26 site were most likely located in the regions from aa 898to 1233 and fromaa992to 1908,respectively.This would be within the proposed serine proteinase motif of the HCV precursor
polyprotein (see Discussion).
Detection oftwo distinct proteinase activities. To confirm that the HCV serineproteinaseis actuallyfunctional and is
required for processing of viral proteins, the processing
potentials of three point mutants derived from N722-1908 were analyzed (Fig. 5). The point mutants used were
pH1083A,pD1107N,andpS1165A,inwhichHis-1083, Asp-1107, and Ser-1165, which were presumed to form the
catalytictriad of the serineproteinaseonthe basis of amino acid sequencehomologywithotherchymotrypsin-likeserine
proteinases(22),wereindependently replaced by Ala-1083,
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
TWO PROTEINASES IN PROCESSING OF HCV POLYPROTEIN 4671
A.
B.
909 + ,+ 959+
1 PLMZVLQAGMTRVPYFVRAQGLIRICMLVRXVAGGHYVQMAF KLAALTGTYVYDHL
2 I-
B---G----I-N--4 -AYL-KGAL--M---HA-L-M-TM--HL---R----VLLALGRW--
I----5 -AYL-KASLL-I---R---A-L-V-T--KHL--AR-I--LLI-IGRW
----I----6 --WI---SLLK---V---L-F-A-A--MI---VII--G---
N--,+ + + 1000 +
1 TPLRDIWAHAGLRDLAVAVE PVVFSDMETKI ITWGADTAACGDIISGLPVSARRGKE
2 ---PR ---
L---3 ---Q----T---L---5--- R-4 --MS5---AN---II--P--E-V-V---E---LE ---L-R-5 S--ST--AQ---I---P--K-V-V---X-V
----LH---L-R-6 ---
--N---Q----L---N---I-1050 +.
ILLGPADSFGEQGWRLLAPITAYSQQTRGLLGCIITSLTGRDKNQVDGEVQVISTA
2
---LEGR-L---E---V---3 ---LEGR--- A--- F--- ----E---V---4 V---GYTSK--S- --- A---T-VVSM---TEQA--I---V
5 V---GYTSK--1C---T---A-VV---EQA-Q---SV
6
---GGVSX---A---E----IV---# 1109
TQSFLATCVNGVCWTNV AGSXTLAGPXGPITQMYT - aDVGWPAPPGARSM6T
2 --- - A-
----3 - I- --- - _-- --- L
4 ---G TIS--L---- ---N--SR--V----Ss --SS---TX-LZ
5 --T--G-SIS--L----_ N V ___---- ----S---TK-LD
6 A----TR-I-S-I
T__---VI-Q-S--L-~~1159
1 PCTCGSSDLYLVTRADVIPVVReG2SLGGLLSPSPRPVSS G_LCP SGSVH
2 --________--__ll--A
3 ---
I--4 AV-- L-T--i-i ___
__--A-5 ---AV N--K---D-R--A---L-T w
--SR--A-6 ---_--
---A--A-**k-l- ~12+?+ , , +
I GtFRAA-VCTRGVAAVDFIVESMETTMRSPFTDNS SPPAVPQTFQVAHLHATG
3 _---V--- ---4
_-?J-S--
-- SI---TLDIVT---T-S---T---Y--GY---5 _L---SI--- LDVAT-T-S-S---T---
SY--GY---6 - ---NL---V--5S
---p125
pI00
p91 -l
p70
-~~~~~~~~~~~~~~~~~~~~~l
p26
[image:7.612.63.297.75.398.2]-p21
FIG. 5. Analysis for proteinasesCpro-1 and Cpro-2 withmutant
constructs.(A) The amino acidsequence(line 1) of the region from residue898to residue 1233 of the HCVprecursorpolyprotein, in
which Cpro-1 activity was detected, is alignedwith those of the corresponding regions of otherreported HCV clones,HCV-BK (line 2) (38),HCV-TW (line 3) (8), HC-J6 (line 4) (33),HC-J8 (line5) (32), andHCV-US (line 6) (11). Standard one-letter aminoacid abbrevi-ations areused. Hyphens indicate amino acid residues identicalto
those derived from the N722-1908sequence atthesamepositions.
Predicted domains homologous with that of chymotrypsin-like serineproteinaseareshaded (22). Conserved residuesin the serine
proteinase motifs of HCV, flaviviruses, and pestivirusesareboxed. #, the residuespredicted toforma catalytictriad, histidine 1083, asparticacid 1107, and serine 1165;*,residues predictedto
contrib-ute to substrate binding; +, histidine, glutamic acid, and cystein residues that are relatively conserved in these clones and were
independently substituted. (B) RNAs from eachof the 22 templates, which encoded a single-amino-acid mutant polypeptide of N722-1908,were translated in thepresence of microsomal membranes. Thecleavagereaction productswereanalyzedby electrophoresison an SDS-11% polyacrylamide gel. Intermediates and products are
indicated byarrowsandarrowheads, respectively, ontheleft.
Asn-1107, and Ala-1165, respectively. The processing
pat-terns of the cleavage reaction products of these mutants wereidentical. Productsp91, p70, and p26werenotfound in these cleavage reaction products, but the accumulation of plOO and p21 was observed (Fig. SB). These results
sug-gested thatthe proteinase activity required for cleavage of thep7O/p26 site was lost inthese mutants. Six other inde-pendent point mutations at amino acid residues in the
putative serine proteinase domain carried in mutants H1136A, H1175A, C1123A, C1125A, C1171A, and C1185A
did not essentially affect the cleavage at the p70/p26 site. Some reduction in cleavage activity was found in three of the four mutants of cysteine residues as described below. To confirm that the p70/p26 site cleavage did not occur in the mutant S1165A, processing of the mutant polypeptide was further examined by immunoprecipitation analysis with the anti-p70 antibody (see Materials and Methods). The major products immunoprecipitated from the S1165A cleavage reaction by this antibody were p125 andpl00 (Fig. 3C, lane d). However, p26 was not detected in that immunoprecipi-tate in either the absence or the presence of microsomal membranes (Fig. 3C, lanes b and d). These data confirm that the serine proteinase identified from the HCV ORF was actually functional and responsible for processing of the C-terminal end of p70. Cleavage of the p21/p70 site, how-ever, does not require the serine proteinase motif but the presence of this region as described above. Therefore, these proteinases, which cleave the p21/p70 site and the p7O/p26
site, weretentatively named HCV proteinase 1 (Cpro-1) and HCVproteinase 2 (Cpro-2), respectively.
Characterization of Cpro-1
activity.
The amino acid se-quence of the Cpro-1 region from aa 898 to 1233 did not show homology to any known proteinase structures except the serine proteinase motif. Therefore, Cpro-1 was character-ized bytesting the effects of known proteinase inhibitors on its activity during pulse-chase analysis of in vitro transla-tions (Fig. 6). During the chase period, the amount of radioactivity of p21 in the N722-1325 pulse-labelled transla-tion productsincreased, while those ofp55and p33 gradually decreased (Fig. 6A). Because the translation was terminated by the addition of cycloheximide and unlabelled methionine to the reaction mixture, theaccumulation of p21 during the chase period implied that p55 had been processed. An essentially identical result was seen during pulse-chase anal-ysis in the absence of microsomal membranes, in which an increasing amount ofradioactive p26' instead of that of p21 during the chase period was detected (data not shown). Product p21 should be stable in this reaction mixture, because this product was resistant toexogenous proteinase Ktreatment, as mentioned above (Fig. 2, lane f). However, p33, the C terminally truncated form of p70, might be unstable and thus degraded in the reticulocyte lysate during the chase period. Because thepattern of p26' increase was not linear with time, in contrast to that of p21 (data not shown), p26' was also likely to be slightly degraded during the chaseperiod. Therefore, Cpro-1 activity was measured bycounting the increase in the amount of relative radioac-tivity inp21 during the chase period. As shown in Fig. 6B, when the Cpro-1 activity in the absence of any proteinase inhibitor is considered to be 100%, it was found that the production ofp21wassuppressed to 22% of the control level only in the presence of EDTA (2 mM). Other proteinase inhibitors had no significant effect onp21 production. The inhibition of Cpro-1 activity by EDTA was decreased by the addition of MgCl2 or ZnCl2 (2 mM) (Fig. 6C). The Cpro-1 activity was increased to three times the control level by the addition ofZnCl2 but not ofMgCl2 (Fig. 6C). Other metal ions,Ni2'
and Co2+, also had no significant effects on Cpro-1 activity (data not shown). Furthermore, the amount ofradioactivep33, which decreased rather gradually without theaddition of ZnCl2 as shown in Fig. 6A, was also observed to beincreased by the addition of ZnCl2 (data not shown). These results indicated that production ofp21 during pulse-chase analysis was due to a site-specific cleavage reaction rather than to degradation of the precursorpolypeptide and suggested that Cpro-1 is azinc-dependentmetalloproteinase. VOL.67, 1993on November 9, 2019 by guest
http://jvi.asm.org/
4672 HIJIKATA ET AL.
A.
B.
C.
15 30 45 60 chaseperiod(min)
120
10(
2 e
0 0.
a-pulse:30min chase:60min
pulse:30min
40 chase:60min
i
°"
300-to--
%'-02 200
A.
0
0
EDTA(mM) 2 2 2 ZnCI2(mM) 2 2
MgCI2(mMM) 2 - 2
FIG. 6. Effects of various proteinase inhibitorsonCpro-1 activity. (A) Relativeradioactivities ofp55, p33,andp21in thechaseperiod.
RNAtranscribed frompN722-1325in vitrowastranslated invitrointhepresenceofmicrosomalmembranesand35S-labeledmethionine. The
chasereactionwasterminatedafterincubationat30°Cfor15, 30, 45,and 60min bythe addition of 2x samplebuffer(28).Allsampleswere
separatedbyelectrophoresisonanSDS-12%polyacrylamide gel. Relative radioactivitiesweremeasured withanimaging plateandBAS2000
(Fuji PhotoFilmCo.)andarerepresentedasphotostimulatedluminescence (PSL) (1).To confirm the termination oflabeling reaction,the translationreactionwascarriedoutin thepresenceofcycloheximideand nonradiolabeledmethionine in additiontothe standardradiolabeling condition. Open circles, p55; open squares, p33; closed circles, p21. (B) Cpro-1 activitywas measured as an increase in the relative
radioactivity of p21 during the 60-min chase. The increase intherelativeradioactivity of p21 in the control withoutanyproteinase inhibitors wasarbitrarilydesignatedas100% Cpro-1 activity. The proteinaseinhibitorsusedwereaprotinin, E64,pepstatin, EDTA, APMSF,antipain, bestatin, chymostatin, and phosphoramidon.Dataareaveragevalues fortriplicate experiments. (C) The chase reactionwascarriedoutin the absence (-)or presence(+)ofEDTA.Reactionmixtureswereincubatedat30°Cfor 60min in theabsenceorpresenceofZnCl2orMgCl2 asindicated.
MutationanalysisofCpro-1activity.Therewasnoamino acidsequence motif in theCpro-1 region thatwas homolo-gous with those found in the other known
metalloprotein-ases. Therefore, candidate residues coordinating with zinc
ion in the Cpro-1 region were determined through site-directed mutagenesis. Almost all metalloproteinases and metalloendopeptidases share common zinc-binding amino acids;histidine andglutamicacidresiduesarewellknownas
coordinators of zinc intheseenzymes(30, 41; forareview,
see reference 12). The cysteine residue is suggested to be essential for the catalytic properties of several metal
en-zymes and metal-binding proteins (41; for a review, see
reference12).Site-specificmutationsatzinc-binding ligands of a metalloproteaselead tocomplete abolition of the
pro-teaseactivity andconcomitant accumulation ofan inactive
precursorpolypeptide (6). Therefore, aseries ofpN722-1908
point mutants were constructed, in which the histidine, glutamic acid, and cysteine residues in the Cpro-1 region, normally conserved among all reportedHCV strains, were
independently replaced by alanine, glutamine, and alanine residues, respectively (Fig. 5A). The substituted amino acids
wereHis-932, -952, -1136, -1175, -1227, and -1229; Glu-972,
-980, -1009, -1058, -1199, and -1202; and Cys-922, -993, -1123, -1125, -1171, and -1185 in the HCV precursor
pro-teins; the translation products of these point mutantswere
named H932A, H952A, H1136A, H1175A, H1227A, H1229A, E972Q, E980Q, E1009Q, E1058Q, E1199Q, E1202Q, C922A, C993A, C1123A, C1125A, C1171A, and C1185A, respectively. Then, the catalytic properties of Cpro-1 and Cpro-2 of these mutants were compared with
those ofthewild-type construct(Fig. 5B). Since the Cpro-1 andCpro-2 activitieswerebothrequired for the production
ofp70,thepresenceofp70in thecleavage reaction products
of themutantRNAs indicated the retention of both protein-aseactivities inasinglemutantprotein.The absence ofp21 and p70 and the accumulation of the 91-kDa processing intermediatemayhaveimplied the lossofCpro-1activity. Of thesesingle point mutants, H952A and C993Awerefound
not to produce p21 and p70 but did accumulate p26 and 91-kDa products (Fig. 5B), which suggested that they had lostCpro-1 activitybut retainedCpro-2 activity. The same
result was obtained by in vitro translation of the point
mutant H952R, in which His-952was replaced by arginine (Fig. SB).Allmutantsinwhich glutamicacidresidueswere
substituted retained both proteinaseactivities, althoughthe Cpro-1 activityofE972Qwasreduced(Fig.5B).Allmutants ofcysteineresidues, exceptC993A, produced p21 andp7O, althoughin variousamounts.WithC1123A andC1171A, p70 was detectable only after prolonged incubation (data not shown). The main cleavage reaction product of C1123A, C1125A, and C1171Awas the 125-kDaprecursor polypep-tide, and no significant accumulation ofintermediates was
observed (Fig. SB). This indicated that the Cpro-1 and Cpro-2 activities were probablybothreduced in these
mu-tants and that cysteines 1123, 1125, and 1171, which are
located in the serineproteinasedomain of the HCV precur-sorpolyprotein(Fig.5A),areimportantbutnotessentialfor both Cpro-1 and Cpro-2 activity. From these results, the candidates for coordination with zinc ion in the Cpro-1 regionareproposed tobeHis-952andCys-993oftheHCV
precursor polyprotein, although further biochemical and structuralanalysesofCpro-1arerequiredtoprecisely deter-mine the metal coordinators.
Expression ofHCVproteins in cultured cells. Toconfirm that similarprocessingoccursineukaryotic cells, expression of the genes for polypeptides N729-1908, S1165A, and , 2
U) co 0
1
a
2 I
I
LI
J. VIROL.
6.. *
'436" 0.-"a'---43
-0
on November 9, 2019 by guest
http://jvi.asm.org/
[image:8.612.128.485.71.263.2]TWO PROTEINASES IN PROCESSING OF HCV POLYPROTEIN 4673
4-000 $l
IG
Mr x 10
o,,4?'
0 00 Mr _
200-Mr 3
x 10
-200
-115
115- 98
98- -; 4p0
6-4
~~-dP7)
_- 6645-45
COS-1 cells Huh-7 cells
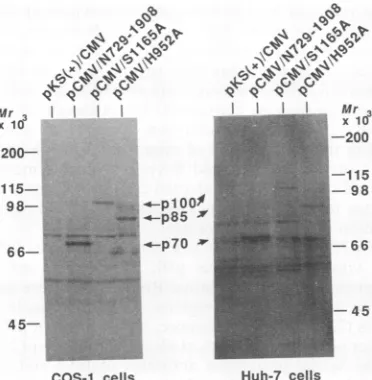
FIG. 7. Expression of HCV polypeptides in COS-1 and Huh-7 cells.Expression plasmidspCMV/N729-1908, pCMV/S1165A, and pCMV/H952Aweretransfected into COS-1orHuh-7cells,and the
proteins expressed bythesecell lineswere analyzed by immuno-blottingwith anti-p70antibody afterseparation by electrophoresis
on an SDS-11% polyacrylamide gelasdescribed in Materials and
Methods.
H952A in COS-1 cells and Huh-7 cellswas examined with
the expression vector pKS(+)/CMV (see Materials and Methods). Theexpression productswereanalyzed by immu-noblotting withanti-p70 antibody. As shown in Fig. 7, p70,
plOO, and an 85-kDa intermediate (p85) were detected in bothCOS-1 and Huh-7 cells transfected with pCMV/N729-1908,pCMV/S1165A, andpCMV/H952A, respectively, and scarcely any p91was found. The appearance ofp85 might havemeantthatfurtherprocessing of p21 occurred in vivo. Therefore, itwas concluded that theprocessing of the p70
precursor in the HCV ORF characterized in an in vitro
protein expression system reflected protein processing in
vivo.
DISCUSSION
Twoproteinases required for HCV precursor polyprotein
processing. Two distinct proteinases, Cpro-1 and Cpro-2, functioned inthe production of p70 from the putative non-structural region of HCV precursor polyprotein. Those activitieswere demonstrated by cleavage of clonedpartial HCV precursor polyprotein in an in vitro transcription/
translation system and by transient expression of cloned genes in cultured cells. The results suggested that both proteinase activitieswerelocated in the HCVpolypeptide. Thissuggestionmaybesupported by therecentobservation thatp70wasproduced from HCV polypeptide expressed in
E. colivia proteolytic processing identicaltothat found in the present paper (24). However, further biochemical and structural characterizations of purified enzymes will be
required for the final identification. It is possible that an
unknown proteinase(s) commonly present in the rabbit reticulocyte lysate,eukaryotic cultured cells, and E. colican
specifically recognize regions of the HCVprecursor poly-protein. The multiple proteinases encoded by thegenomesof someotherviruses, suchas poliovirus (40) and plant poty-virus(42), have been reported and characterized. Cpro-1 and Cpro-2 were responsible for the cleavages of the N- and
(722) -810 -1020
l1 1 serine protelnase-1650' (1908) aa I
I,
HD SNp70
AH C| Zn-dependent metalloprotelnase ?
[image:9.612.87.273.76.266.2]RTM_Cm C
FIG. 8. Summary ofprocessing and production of HCVp21and p70 products. The box indicates a part of the HCV precursor polyprotein. The N- andC-terminal sides of the precursor are shown byNand C on theleft and right ofthebox. Thep21and p26regions of the HCV precursor polyprotein are indicated by hatched and latticed boxes, respectively. Cleavages that have been shown or proposed to occur in a microsome-dependent manner and by pro-teinases encoded in the HCV genomeareindicatedbyanarrowhead and arrows,respectively. Theapproximatelocationsof the cleavage sitesareindicatedatthe top. Theputative Cpro-2 serine proteinase domain is shaded, and the predicted catalytic triad, His-1083, Asp-1107,andSer-1165, of the serineproteinase is indicatedby H, D, and S, respectively, above the box. The region required for detection ofCpro-1 activity is bracketed. Theproposed zinc (Zn)-coordinating residues,His-952 andCys-993,ofCpro-1areindicated by Hand C,
respectively,
below the box.C-terminal ends of p70, respectively. Both cleavage
reac-tions seemed to occur
mainly
intramolecularly(in
acis-dependent manner) because p70 production could not be
detectedin either in vitrotranslation reaction mixturesorin
cell lysates whichcoexpressed S1165A and H952A(datanot
shown). Although no inhibitor assay for Cpro-2 could be performed,point mutation analysis indicated that Cpro-2 isa
serineproteinase,aspredicted byasequence
alignment
and modeling study(22).Theresults of inhibitorassaysandpoint mutationanalysessuggested thatCpro-1
can beclassifiedas azinc-dependentmetalloproteinase.
Thecatalytic activities ofthese twoproteinasesare sepa-rable, because some mutants, such as S1165A andH952A,
retainedonly one of theseactivities(Fig.
5B).
However, theregionsrequiredfordetectionoftheseactivitiesintheHCV
precursor polyprotein overlapped. The Cpro-2 serine pro-teinase islikelytobelocatedin theregionfromresidue1075 to1185 of the HCVprecursorpolyprotein (22),
correspond-ingtotheN-terminalregion of
p70.
Atleasttheregion from residue898 to 1233 of the HCV precursorpolyproteinwasrevealedto be essential for the detection of Cpro-1 activity by analyses of a series of deletion mutants. The results suggestedthat thecandidatesfor thecoordinatorsof zinc ion intheCpro-1regionare
His-952
and Cys-993,bothof whichare located in the C-terminal region of
p21
(Fig. 8). How-ever, furtherexperimentation
with the authentic product, containing the Cpro-1 region, is required to confirm this suggestion, since thepossibility
that each point mutationcauses a change in the tertiary structure of
p21
and/oranother unknown type(s) of amino acid
residue(s)
which contributestozincbindingcannotbe completely ruledout. In addition, the C-terminal region just downstream of the serineresidue in theserineproteinasedomainwasessentialfor the cleavage mediated by
Cpro-1
activity, because theC-terminal deletion mutant, N722-1174, was notcleaved at the
p2l/p7O
site (Fig. 4A, lane 1). In thatmutant,only
the small regioncorresponding
to the C-terminalsubstrate-binding pocketof the serineproteinasestructure wasdeleted
(Fig. 5A).Largeramountsof
unprocessed
products
accumu-lated in thecleavagereactions ofN722-1477,N722-1325,
andVOL.67, 1993
on November 9, 2019 by guest
http://jvi.asm.org/
[image:9.612.334.548.77.157.2]4674 HIJIKATA ET AL.
N722-1233 than in that of N722-1908
(Fig.
4A),
indicating
that even a deletion in the
p70
C-terminalregion
reduced Cpro-1activity.
Furthermore,
the mutations ofcysteine
residues at aa 1123,
1125,
and 1171 to alanine residues reduced both theCpro-1 and theCpro-2
activities(Fig.
SB),
suggesting the
requirement
oftertiary
structure for full activity.There seemtobe atleasttwopossible
explanations
forwhyoverlapping regions
ofthe HCVprecursorpolypro-tein might be
required
for the detection of two distinct proteinase activities. Oneis that the entiretertiary
structureof thep21-p70precursor
protein
oratleasttheoverlapping
serine
proteinase
domain isrequired
foritsrecognition
as a substrateby the active domain ofCpro-1.
Cpro-1
might
belocated in the C-terminal half of
p21
or in the serineproteinase
structure. The otherpossibility
is that the se-quenceof theC-terminal half ofp21
plus
the active domain ofCpro-2 serineproteinase
functions as ametalloproteinase
with Cpro-1
activity.
In the former case,Cpro-1
would bethe smallestknown
metalloproteinase
thatspecifically
rec-ognizes
thetertiary
structure of alarge
polypeptide
as asubstrate. In the latter case, the structureof
Cpro-1
may bepartly constructed withtheserine
proteinase
structure, and its activity should be inactivated afterautocatalytic
ciscleavage,
implying
that twoproteinase
domainsof the HCV precursorpolyprotein
overlap
and form the enzyme in enzyme structure.Although
there has beennodirect report of aproteinase
like that described in the first case, adomain-overlapping
structure in the structural coreprotein
ofSindbis virus has been
recently
reported (9,
16).
Thecoreprotein was revealed to contain a C
terminally
truncated form ofachymotrypsin-like
serineproteinase
structure, the proteinaseactivity
of whichisresponsible
forreleasing
thecore
protein
from the precursorprotein by
autocatalytic
ciscleavage
(9, 16).
This indicated that thisviral coreprotein
hastwo
overlapping
functionaldomains,
onefor structural protein and another for enzyme. The serineproteinase
activity in the precursor of the core
protein
issuicidal,
becausethe
cleavage
site of theproteinase
islocated withinthe
substrate-binding
region
oftheproteinase
itself(9).
If theCpro-1
andCpro-2
domainsoverlap,
then theCpro-1
activity
may have a similar suicidal function that releases
Cpro-2
from the
Cpro-1
structure andthe HCVprecursorpolypro-tein.
PutativeNS
proteins
of HCV. Theregion
spanning
residue722 to residue 1908 of the HCV precursor
polyprotein
produced three
major
processed
products, p21, p70,
andp26
(Fig. 8).
Thisregion
may also haveproduced
smallerpro-cessed
polypeptides
thatwere notdetectableby
themethod used.Thefunctionalproperties
ofp21
andp70
asproteinases
were
suggested,
butthefunctionof thewild-type
formofp26
is still unknown. An
immunoprecipitation
experiment,
shown in
Fig. 3,
suggested
thatp26
waslikely
tobeassoci-ated with
p70
at least in the in vitro translation mixture. Recent in vitrotranscription/translation
experiments
with theentire NSprotein
region
from the HCVgenomesuggestthatp26 isaC-terminal truncated form of the31-kDa
(p31)
productwhich is
dependent
onCpro-2
activity
(20).
Invitro,
p31
is alsoassociatedwithp70,
although
thesignificance
of this molecular association is still unknown(20). Moreover,
p31
islikely
to be furtherprocessed
to smallerproducts
when
expressed
invivo,
suggesting
thepresence ofacellularproteolyticsite in
p31
(20).
The location and size of
p21
in the viral precursorpoly-protein
arelikely
tobe similartothose of flavivirusNS2aorNS2b
(37).
Theprotein
NS2aissuggested
tobeaproteinase
that cleaves the N-terminal end of its molecule(13),
andNS2b is
required
for thetrans-acting
activities of theflavi-virus serine
proteinase
NS3(3,
4,
14).
However,
thepro-cessing
oftheregion encoding p21
islikely
tobedifferent
from that ofthe
flavivirus
NS2region,
because theNS2a/
NS2band
NS2b/NS3
cleavages
areknowntobemediatedby
NS3 serineproteinase
activity (4,
14). Although
theprecise
function(s)
ofp21
is stillunknown,
itappeared
that itwaslocated in the
lipid bilayer
ofmicrosomal membranes. NSproteins
of the HCV-relatedflavivirus
form amembrane-associated
complex
in theinfected cell(35).
p21
mayplay
ananalogous
role intheprocessing,
assembly,
andmembraneassociation ofHCV NS
proteins.
HCV
p70
seems to be ahomolog
offlavivirus
NS3 andbovine viral diarrheal virus
p80,
because the motifs for serineproteinase
andaputative
RNAhelicasearedetected in the N- and C-terminalregions
of theserespective
viralproducts
(2,
22,
23).
Furthermore,
thelocation ofp70
in the precursorpolyprotein
is almostidenticaltothoseof NS3andp80.
The serineproteinase
activities of NS3 andp80
areessentialforthe
processing
of otherparts
oftheirrespective
NSproteins
(3,
4,
14).
The datapresented
heresuggested
that the
Cpro-2
serineproteinase
activity
wasrequired
forthe
processing
of HCV NSproteins.
Recently,
itwasshown thatprocessing
of the C-terminalportion
of the HCVpre-cursor
polyprotein
is abolishedby
apoint
mutation of serineataa1165toalaninein thedomainof
Cpro-2
proteinase (20).
ACKNOWLEDGMENTSThis work was
supported
in partby
agrant-in-aid
from theMinistry
of Health and WelfareComprehensive
10-YearStrategy
for CancerControl,
agrant-in-aid
for Scientific Research from theMinistry
ofEducation, Science,
andCulture,
Japan,
andaresearchgrantfrom the Princess Takamatsu CancerResearch Fund. REFERENCES
1.
Amemiya,
Y.,andJ.Miyahara.
1988.Imaging
plate
illuminates manyfields.Nature(London)
336:89-90.2. Bazan,J.F.,and R.J.
Flettericlk
1989. Detection ofatrypsin-like serine protease domain in flaviviruses and
pestiviruses.
Virology
171:637-639.3. Cahour, A.,B.
Falgout,
and C.-J. Lai. 1992.Cleavage
of thedengue
viruspolyprotein
at theNS3/NS4A
andNS4B/NS5
junctions
is mediatedby
viral proteaseNS2B-NS3,
whereas NS4A/NS4Bmay beprocessed by
acellularprotease.J.Virol.66:1535-1542.
4. Chambers,T.J.,A.Grakoui,and C.M. Rice.1991.
Processing
of theyellow
fevervirusnon-structuralpolyprotein:
acatalyti-cally
activeNS3proteinase
domain andNS2Barerequired
forcleavages
atdibasic sites. J.Virol.65:6042-6050.5. Chambers,T.J.,R.C.Weir,A.Grakoui,D.W.McCourt,J.F.
Bazan,R.J.Fletterick,and C. M. Rice.1990. Evidence that the N-terminal domain ofnonstructural
protein
NS3
fromyellow
fever virus is a serineprotease
responsible
forsite-specific
cleavages
intheviralpolyprotein.
Proc. Natl.Acad.Sci. USA 87:8898-8902.6.
Chang,
P.-C.,andY.-H. W. Lee. 1992. Extracellularautopro-cessing
ofametalloprotease
fromStreptomyces
cacaoi. J. Biol. Chem.267:3952-3958.7. Chen, C.,and H.
Okayama.
1987.High-efficiency
transforma-tion of mammalian cellsby
plasmid
DNA. Mol. Cell. Biol. 7:2745-2752.8. Chen, P.-J., M.-H. Lin, K.-F. Tai, P.-C. LAu, C.-J.
LAn,
and D.-S. Chen. 1992. The Taiwanesehepatitis
C virus genome: sequence determination andmapping
the 5' termini of viralgenomic
andantigenomic
RNA.Virology
188:102-113. 9.Choi,
H.-K.,L.Tong,
W.Minor, P. Dumas,U.Boege,
M.G.Rossmann, and G.
Wengler.
1991. Structure ofSindbis viruscore
protein
revealsachymotrypsin-like
serineproteinase
andthe