0022-538X/05/$08.00⫹0 doi:10.1128/JVI.79.23.14708–14718.2005
Copyright © 2005, American Society for Microbiology. All Rights Reserved.
The Neuronal Host Cell Factor-Binding Protein Zhangfei Inhibits
Herpes Simplex Virus Replication
Oksana Akhova,
1Matthew Bainbridge,
2and Vikram Misra
1*
Department of Veterinary Microbiology, Western College of Veterinary Medicine,1and Department of Computer Science,
University of Saskatchewan,2Saskatoon, Saskatchewan S7N 5B4, Canada
Received 12 July 2005/Accepted 9 September 2005
During lytic infection in epithelial cells the expression of herpes simplex virus type 1 (HSV-1) immediate-early (IE) genes is initiated by a multiprotein complex comprising the virion-associated protein VP16 and two cellular proteins, host cellular factor (HCF) and Oct-1. Oct-1 directly recognizes TAATGARAT elements in promoters of IE genes. The role of HCF is not clear. HSV-1 also infects sensory neurons innervating the site of productive infection and establishes a latent infection in these cells. It is likely that some VP16 is retained by the HSV-1 nucleocapsid as it reaches the neuronal nucleus. Its activity must therefore be suppressed for successful establishment of viral latency. Recently, we discovered an HCF-binding cellular protein called Zhangfei. Zhangfei, in an dependent manner, inhibits Luman/LZIP/CREB3, another cellular HCF-binding transcription factor. Here we show that Zhangfei is selectively expressed in human neurons. When delivered to cultured cells that do not normally express the protein, Zhangfei inhibited the ability of VP16 to activate HSV-1 IE expression. The inhibition was specific for HCF-dependent transcriptional activation by VP16, since a Gal4-VP16 chimeric protein was inhibited only on a TAATGARAT-containing promoter and not a on a Gal4-containing promoter. Zhangfei associated with VP16 and inhibited formation of the VP16–HCF– Oct-1 complex on TAATGARAT motifs. Zhangfei also suppressed HSV-1-induced expression of several cellular genes including topoisomerase II␣, suggesting that in addition to suppressing IE expression Zhangfei may have an inhibitory effect on HSV-1 DNA replication and late gene expression.
Herpes simplex virus type 1 (HSV-1) uses two complemen-tary strategies to avoid immune surveillance and to maintain itself in its host. Infection begins with virus replication and the lysis of infected epithelial cells. During this phase of lytic rep-lication the virus infects sensory nerves and establishes a latent infection in neuronal nuclei located in sensory ganglia (re-viewed in reference 6). The virus is thought to be almost completely quiescent during latent infection. Only one viral transcript and no readily detectible viral proteins are expressed. A variety of physiological and psychological stressors reactivate the latent viral genome to reenter the lytic phase. Following reactivation, the virus replicates in neurons and travels down the axons of sensory neurons to replicate again in epithelial cells causing a recrudescent lesion (reviewed in references 37 and 38).
During the lytic infection of epithelial cells the expression of viral genes is temporally regulated and the approximately 80 viral genes involved can be described as immediate-early (IE), early (E), or late (L) depending upon the order of their expres-sion. Initiation of the transcription of the five IE genes (ICP0, ICP4, ICP22, ICP27, and ICP47) is induced by the assembly of a multiprotein complex (VP16-induced complex [VIC]) made up of the virion protein VP16 and the two cellular proteins Oct-1 and host cellular factor (HCF) (reviewed in reference 21).
VP16 is synthesized in the late stages of lytic infection and is incorporated into the tegument of the virion. Upon infection,
VP16 is released into the cell, where it combines with HCF. This heterodimer recognizes Oct-1 bound to TAATGARAT motifs present in multiple copies in the promoters of all IE genes. VP16 interacts with HCF through a 4-amino-acid motif—EHAY, which is conserved as the sequence D/EHxY (where x can be any of several amino acids) in VP16 homo-logues in other herpesviruses and in some cellular HCF-bind-ing proteins. Transcriptional activation induced by VIC also depends on the VP16 activation domain located in the carboxyl terminus of the protein. This domain has binding sites for various components of the transcriptional complex as well as for the chromatin modification factors p300 and CBP (re-viewed in reference 11). The presence of the VP16 carboxy-terminal activation domain is not required for VIC formation (11) but is critical for HSV-1 IE gene expression (32).
HCF is a ubiquitously expressed chromatin-associated pro-tein. The mature form of HCF is derived from a 2,035-amino-acid precursor. Upon removal of internal repetitive domains the amino- and carboxy-terminal domains of HCF associate by noncovalent interactions (17, 45). These components of HCF play different roles in cell cycle progression and cytokinesis (14). Other functions of HCF such as chromatin structure modification and RNA splicing have recently been reported (2, 46). In addition to these cellular roles, HCF participates in the regulation of the IE gene expression of HSV-1, varicella-zoster virus (33), and possibly other alphaherpesviruses as well. VP16 binds to the amino-terminal component of HCF, which pos-sesses a-propeller-like structure formed by six Kelch repeats (12, 44). Mutations in the residues in the some of the Kelch repeats abolish binding to VP16 and the HCF-binding protein Luman (29). Mutations within the carboxyl terminus of the protein reduce transcription activity mediated by HCF (26).
* Corresponding author. Mailing address: Department of Veteri-nary Microbiology, Western College of VeteriVeteri-nary Medicine, Univer-sity of Saskatchewan, 52 Campus Drive, Saskatoon, Saskatchewan S7N 5B4, Canada. Phone: (306) 966-7218. Fax: (306) 966-7244. E-mail: [email protected].
14708
on November 8, 2019 by guest
http://jvi.asm.org/
Sequestering of HCF in the cytoplasm, or reducing its expres-sion with short interfering RNAs, disables its functional activ-ity and results in reduction of HSV-1 IE gene expression and viral yield (16, 33). Interestingly, while HCF-1 is an abundant nuclear protein in most cell types, it is found in the cytoplasm of sensory neurons from unstressed animals (18). This natural sequestering of the protein from the nuclei of cells that are the site for HSV-1 latency may contribute to the establishment of latency by the virus.
Several cellular proteins which, like VP16, interact with the amino terminus of HCF share the HCF-binding motif D/EHxY. These include E2F4 (27), Krox20 (27), Luman/LZIP/CREB3 (22, 24, 25), and Zhangfei (23). The motif is present in other pro-teins as well, but HCF binding by these propro-teins has not been examined (27).
Luman and Zhangfei are related basic leucine zipper (b-zip)-containing HCF-binding proteins. Luman activates promoters containing cyclic AMP and unfolded protein re-sponse elements (CRE and UPRE) (25, 30). In contrast, Zhangfei does not bind any of the known cognate sequences for b-zip proteins, nor does it activate promoters containing these sequences (23). However, Zhangfei has a profoundly repressive effect upon HCF-dependent transcription activation by Luman (30). HCF binding by both Zhangfei and Luman is required for efficient suppression by Zhangfei. Interestingly, while Zhangfei and Luman have incompatible b-zip domains and do not interact directly, Zhangfei does have the capacity to dimerize with other factors such as Xbp1 and ATF4 through their b-zip domains (34). The effect of these interactions has not been examined.
Following infection of sensory neurons, HSV-1 virions are transported in a retrograde manner to the neuronal nucleus, a process that involves cellular microtubules and components of the virion tegument (3, 28). Since it is possible that VP16 accompanies the other tegument proteins to the neuronal nu-cleus, its activation of lytic replication must be suppressed as a precondition for the establishment of viral latency. We had previously demonstrated that Zhangfei suppresses viral gene expression in HSV-infected cells, although the mechanism of this inhibition was not known (23). We have also demonstrated that Zhangfei suppresses the ability of Luman to activate tran-scription in an HCF-dependent manner. Since both Luman and VP16 bind HCF and require it for transcriptional activa-tion, we examined the ability of Zhangfei to suppress VP16. In this report we show that Zhangfei can indeed suppress HCF-dependent, VP16-mediated activation of the HSV-1 ICP0 pro-moter, resulting in a profound decrease in ICP0 RNA and protein, as well as in progeny virus yield from virus-infected cells. Zhangfei appeared to interact with VP16 and suppressed VIC formation. In addition, in HSV-1-infected cells, Zhangfei suppressed the expression of several cellular genes that are normally enhanced by the virus. While Zhangfei is not ex-pressed in most tissues, here we also show that it is present in sensory neurons, a location where its properties may be bio-logically relevant to the establishment of viral latency.
MATERIALS AND METHODS
Cells and plasmids.Vero and Hep2 cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM) with 10% newborn calf serum for Vero cells or 10% fetal bovine serum for Hep2 and HeLa cells. The medium also contained
1% penicillin-streptomycin. HEK 293 cells were maintained in RPMI 1640 sup-plemented with 10% fetal bovine serum, 2 mM glutamine, and 1% penicillin-streptomycin.
The plasmids encoding Zhangfei (pcZF); mutant Zhangfei which is unable to bind HCF (pcZF Y224A); LuS221Op, the truncated form of full-length Luman (pcLuS221Op); LuS221Op (N160G) with a mutation in the DNA binding do-main (pcLuS221Op N160G); and HCF (pSL7) have been previously described (19, 23, 30). The plasmid G5EC, a chloramphenicol acetyltransferase (CAT)
reporter which contains five copies of theSaccharomyces cerevisiaeGal4
up-stream activation signal (UAS), as well as the pM series of plasmids for con-structing Gal4 fusion proteins was a gift from Ivan Sadowski. Gal4-VP16 was constructed by transferring the entire VP16 coding sequence from pRG50 into pM. Plasmid RG50, which encodes VP16, and pAB5, a CAT reporter with a portion of the TAATGARAT-containing ICP0 promoter, were provided by Peter O’Hare. Plasmid CI-110, which encodes ICP0, was a gift from Roger Everett.
Viruses.Recombinant adenovirus vectors were constructed as described ear-lier (30). HSV-1 stock (KOS strain) was propagated in Vero cells. For infection of Hep2 cells, the HSV-1 stock was diluted in DMEM and cells were infected at a multiplicity of infection (MOI) of 0.1 PFU per cell (considered low) or 10 PFU/cell (high). Titration of serially diluted HSV-1 was performed in Vero cells overlaid with anti-HSV serum. After incubation for 48 h following infection, cells were stained with 0.5% crystal violet in 80% methanol and plaques were counted.
Antibodies, immunofluorescence, immunohistochemistry, and immunoblot-ting.Polyclonal serum against glutathioneS-transferase (GST)–Zhangfei was raised in rabbits as described previously (23). To deplete the serum of antibodies against GST or GST-Zhangfei, this serum was diluted 10-fold in phosphate-buffered saline and absorbed with either GST or GST-Zhangfei coupled to glutathione Sepharose beads (GE Healthcare). Monoclonal antibodies (LP1) against VP16 were a gift from Peter O’Hare, and ICP0 monoclonal anti-bodies were purchased from the Rumbaugh-Goodwin Institute. Rabbit
poly-clonal antibodies to human topoisomerase II␣(p170 form) were kindly provided
by TopoGen, Inc. (Mark T. Muller). Procedures for immunoblotting and immu-nofluorescence were as previously described (23). Occasionally, cells were fixed in 1% paraformaldehyde and permeabilized with 0.2% Triton X-100. Immuno-histochemistry of human cervical dorsal-root ganglion (provided by Anurag Saxena) or tissue arrays (BioChain, catalog no. T8235712-2) was performed on
5-m paraffin-embedded sections. The sections were dewaxed in CitriSolv
(Fisher) and cleared in graded alcohol. All washings were done in 1⫻
automa-tion buffer (Biomeda Corp.). After washing, the secautoma-tions were soaked in 1.5%
H2O2in methanol to inactivate endogenous peroxide activity. Antigen was
en-hanced by heating tissue sections in a microwave oven in 0.01 M sodium citrate buffer (pH 6.0). The sections were preincubated in blocker containing 4% goat serum in phosphate-buffered saline, 0.02% Tween 20, and 0.05% Brij 15 for 20 min at 42°C. Serum against Zhangfei (GST absorbed) or anti-Zhangfei-depleted serum (as a negative control), diluted 1:100 in 4% goat blocker, was applied overnight at 4°C. After thorough washing, the sections were incubated in biotin-ylated goat anti-rabbit antibodies (Molecular Probes) diluted 1:400 in 4% goat blocker for 20 min at 42°C. The sections were incubated in avidin-biotin complex reagent (ABC kit; Vector) for 30 min at 42°C. After being washed and sub-merged once in 10% methanol, the sections were stained with
3,3-diamino-benzidine (DAB) solution (0.1% DAB, 0.016% H2O2) for 5 min at 42°C. The
sections were counterstained with hematoxylin, processed through graded alco-hols, cleared in xylene, and embedded in balsam.
Transfection and reporter assay.Plasmid DNA was introduced into cells using calcium phosphate as described previously (31). Plasmids for transfection were purified by CsCl gradient centrifugation, and maximum amounts of transfected
DNA did not exceed 6g per 106cells. The procedure for determining CAT
activity was as described previously (30). When adenovirus vectors were used to introduce DNA into cells, virus was diluted in DMEM plus 10% fetal bovine
serum and added to Hep2 cells (1 ml for 1⫻106cells and 4 ml for 4⫻106cells)
at an MOI of 20 PFU/cell. Four hours later fresh medium was added to each well. For some experiments, 24 h after adenovirus infection, Hep2 cells were infected with HSV-1. Hep2-infected cells were harvested 6 h after HSV-1 infec-tion for examinainfec-tion by immunofluorescence and/or immunoblotting and 24 h later for HSV-1 titration.
Northern blotting. Total RNA from Hep2 cells was purified using Trizol reagent (Invitrogen) according to the manufacturer’s protocol. RNA samples (10
g per well) were separated on a 1% agarose-1.85% formaldehyde gel
sub-merged in morpholinepropanesulfonic acid (MOPS)-EDTA-sodium acetate buffer. RNA was transferred to a nylon membrane (Hybond-N; GE Healthcare). The probe for ICP0 was generated from coding sequence for ICP0 recovered
from pCI-110 and labeled with Redivue [␣-32P]dATP (GE Healthcare) using a
on November 8, 2019 by guest
http://jvi.asm.org/
random primer DNA labeling system (Invitrogen). The filters were prehybrid-ized at 45°C for 2 to 4 h in 0.05 M Tris, pH 7.5, containing 50% deionprehybrid-ized formamide, 1 M NaCl, 10% dextran sulfate, 1% sodium dodecyl sulfate (SDS),
and 100g salmon sperm DNA. Hybridization with probe was performed at
45°C for 16 h. After washing, membranes were exposed to autoradiography film (GE Healthcare) with intensifier screen at room temperature for 1 to 2 h.
Preparation of nuclear extracts and HSV-1 stock for electrophoretic mobility shift assay (EMSA).Nuclear extracts were prepared as described in reference 40.
Briefly, HeLa cells (0.1⫻108
to 1⫻108
) infected with adenovirus vectors
expressing Zhangfei, Zhangfei (Y224A), or-galactosidase were harvested 24 h
after infection. Cells were resuspended in 5 volumes of ice-cold buffer containing
10 mM HEPES-KOH (pH 7.9), 1.5 mM MgCl2, 10 mM KCl, 0.5 mM
dithio-threitol (DTT), and Complete Mini protease inhibitor cocktail (Roche) and incubated on ice for 10 min. Cells were recovered by centrifugation at 6,500 rpm for 20 s. Then buffer containing 0.1% NP-40 was added and cells were passed five times through a 22-gauge needle. Nuclei were pelleted by centrifugation and
resuspended in buffer containing 40 mM HEPES-KOH (pH 7.9), 4 mM MgCl2,
0.4 M KCl, 1 mM DTT, 25% glycerol, and Complete Mini protease inhibitor cocktail (Roche). NaCl was added to a final concentration of 300 mM. Nuclei were incubated on ice for 30 min, and proteins were recovered as a supernatant by centrifugation at 14,000 rpm for 10 min at 4°C. Protein samples were frozen
in aliquots at⫺70°C. The concentration of proteins was determined by the
Bio-Rad protein Assay Dye.
HSV-1 stock which was used for EMSA was purified on a Ficoll 400 gradient as described previously (7). Before being mixed with nuclear extracts, the concen-trated HSV-1 stock was incubated with 1% NP-40 for 30 min on ice to release VP16.
EMSA. Double-stranded, 32
P-labeled probe (5⬘-GATCCCGTGCATGCT
AATGATATTCTT, 3⬘-CTAGAAAGAATATCATTAGCATGCACGG) was
incubated in a reaction mixture containing 5g of HeLa nuclear extracts, 20 mM
HEPES-KOH, 0.2 M KCl, 0.5 mM DTT, 12.5% glycerol, 2 mM MgCl2, 5% Ficoll
400, 20 mg/ml bovine serum albumin, and 0.05 mg/ml poly(dI-dC) (Roche). In
some mixtures 5l of NP-40-treated HSV-1 lysate was added. Reaction mixtures
were incubated at 25°C for 20 min and resolved on 5% nondenaturing polyacryl-amide gels at constant voltage (200 V) for 3 h.
Immunoprecipitation analysis.Immunoprecipitation of VP16 with Zhangfei and Zhangfei (Y224A) was done according to the procedure described by Sto-janova et al. (42). Protein concentrations was measured with the Bio-Rad protein
assay dye, and in each reaction 500g of proteins was precipitated with LP1
antibodies diluted 1/100. Twenty percent protein A Sepharose CL-4B beads (GE Healthcare) in radioimmunoprecipitation assay buffer was used to collect the
immunoprecipitates. The beads were boiled with 40l of Laemmli buffer, and
after the beads were removed by brief centrifugation the proteins were separated by SDS-polyacrylamide gel electrophoresis. For two-dimensional gel
electro-phoresis the beads with immunoprecipitates were boiled in 140l of
two-dimensional buffer containing 9.8 M urea, 4% CHAPS {3-[(3-cholamidopropyl)-dimethylammonio]-1-propanesulfonate}, 65 mM of DTT, and 0.5% ampolyte mixture composed of 0.25% Bio-Lyte 3/10 and 0.25% Bio-Lyte 7/9 (Bio-Rad).
Two-dimensional gel electrophoresis.Seven-centimeter gel strips of pH 3 to 10 (Bio-Rad) were rehydrated at 50 V in two-dimensional buffer for 12 h. First-dimension separation of proteins, based on their isoelectric points, was carried out on the Bio-Rad Protean Isoelectrofocuser until 10,000 V · h was reached. The gel strips were incubated in equilibration buffer II containing 6 M urea, 2% SDS, 0.375 M Tris (pH 8.8), 20% glycerol, and 1% DTT for 15 min and equilibration buffer II containing 6 M urea, 2% SDS, 0.375 M Tris (pH 8.8), 20% glycerol, and 2.5% iodoacetamide for 20 min. A trace of bromphenol blue was added to the equilibration buffer II. The strips were washed in running buffer (0.25 M Tris, 1.92 M glycine, and 1% SDS), placed in a Bio-Rad Mini Protean gel apparatus, and overlaid with 0.5% agarose in running buffer. The proteins were separated according to their molecular weights on a 10% polyacrylamide gel at 200 V. Gels were stained with silver (41).
Microarrays.Each cDNA microarray from the Microarray Center (Ontario Cancer Institute, Canada) contains 19,000 human expressed sequence tag clones. RNA from Hep2 cells was treated as for Northern blot assays, except that HSV-1 infection was performed at an MOI of 1 PFU/cell. RNA samples were treated with 100 U of DNase I (Invitrogen) for 15 min. Reverse transcription of RNA and labeling of cDNA with Alexa Fluor 555 and 647 dyes were performed
according to the instructions supplied by Molecular Probes. Briefly, 10g of
RNA was mixed with anchored oligo(dT) primers (dT 20 VN) and heated at 70°C for 10 min. The reaction mixtures were then cooled on ice for 5 min. The reverse transcription of RNA was performed in a mixture containing dATP, dCTP, and dGTP (0.5 mM); dTTP (0.15 mM); aminoallyl-dUTP (0.30 mM); and 40 U of SSII at 42°C for 2 h. Hybridization and washing of microarrays were done as described earlier (30).
Array scanning and data analysis were carried out as described in reference 30.
RESULTS
Zhangfei is selectively expressed in human neurons.In this study we wished to explore the possibility that Zhangfei, by suppressing VP16-mediated initiation of the lytic cycle, might direct the viral genome into a latent state. However, to be of any biological relevance Zhangfei must be present in sensory neurons, the site of HSV-1 latency. While Zhangfei mRNA is present in many tissues, the protein is not expressed in cultured cells of epithelial and fibroblastic origin (22, 30). To identify tissues that contain the protein, we stained a human tissue array representing 31 human normal and tumor tissues with antiserum against Zhangfei. As a negative control we stained a parallel array with the anti-Zhangfei serum from which most of the antibodies against Zhangfei had been removed by absorp-tion to recombinant Zhangfei. Figure 1A shows that the anti-Zhangfei serum clearly recognized the protein on immuno-blots while binding by serum from which antibodies against Zhangfei have been removed by absorption was greatly re-duced. The only tissue in the array that specifically stained for Zhangfei was the parietal lobe of the cerebrum, the only rep-resentative of neuronal tissue in the array. In the cerebrum the protein was located in pyramidal neurons (Fig. 1B). The stain-ing was specific, as serum from which anti-Zhangfei antibodies had been removed stained poorly. Since the array did not contain examples of sensory ganglia, the main site of HSV-1 latency, we examined sections of formalin-fixed, paraffin-em-bedded human cervical dorsal-root ganglion. Here only the neurons stained for Zhangfei (Fig. 1C), and again, staining was reduced when anti-Zhangfei-depleted serum was used for de-tection. All neurons in the sections examined appeared to contain Zhangfei, although the intensity of staining varied. These results show that Zhangfei is selectively expressed in human sensory neurons and is thus well placed to influence HSV-1 latency.
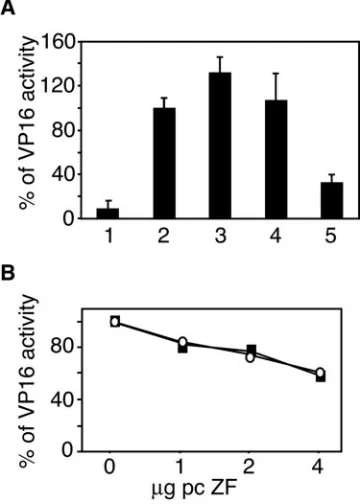
Zhangfei suppresses VP16-mediated expression of ICP0 promoter. We recently demonstrated that Zhangfei, in an HCF-dependent manner, inhibited transcriptional activation by Luman, another HCF-binding factor (30). Since Luman is considered a cellular analogue of VP16, we hypothesized that Zhangfei would suppress VP16-mediated transcription as well. A plasmid containing the coding sequences of the reporter protein CAT linked to a segment of the HSV-1 ICP0 promoter containing one copy of TAATGARAT was introduced into Vero cells along with optimal amounts of a vector expressing VP16. Some cultures were also cotransfected with increasing amounts of pcZF, a plasmid expressing Zhangfei. Zhangfei reduced VP16-mediated activation to basal levels (Fig. 2A). A plasmid expressing comparable levels (Fig. 2B) of a mutant of Zhangfei (ZF Y224A) that is unable to bind HCF (23) was less effective.
Zhangfei suppresses transcription and protein synthesis of ICP0 as well as progeny virus yield during HSV-1 infection.
Next we examined the effect of Zhangfei on ICP0 expression and virus replication in HSV-1-infected cells. Hep2 cells were infected with recombinant adenoviruses expressing either Zhang-fei, Zhangfei (Y224A), or-galactosidase. Twenty-four hours later the cells were infected with HSV-1 at an MOI of 0.1
on November 8, 2019 by guest
http://jvi.asm.org/
PFU/cell. Six hours postinfection we examined the cells for ICP0 RNA and protein. We also determined virus yield from the infected cultures 24 h after infection. Zhangfei, but not Zhangfei (Y224A), drastically reduced ICP0 RNA in infected cells (Fig. 3A). This decrease in ICP0 transcripts was also reflected in the number of ICP0-expressing cells (Fig. 3B and C, compare panels and lanes 1 and 3). As expected, there was also a corresponding decrease in the late protein VP16 pro-duced in Zhangfei-expressing cells (Fig. 3D) as well as progeny virus yielded by these cells (Fig. 3E). In contrast to Zhangfei, Zhangfei (Y224A) had little effect (compare lanes and panels 2 and 3). Although ICP0 RNA and protein yields were also suppressed in Zhangfei-expressing cells infected with HSV-1 at
a higher MOI of 10 PFU/cell (results not shown), this suppres-sion was not mirrored by a similar decrease in virus yields.
Zhangfei inhibitory effect is not due to the limiting quantity of HCF-1. A possible explanation for the inhibitory effect of Zhangfei is that Zhangfei deprives VP16 of HCF by sequestering the factor. Two experimental approaches were used to explore this possibility. First, VP16-mediated activation of the ICP0 pro-moter was tested in the presence of Zhangfei, as well as another HCF-binding factor, Luman. If Zhangfei binds all available cel-lular HCF, preventing activation by VP16, Luman, which also binds HCF, should also suppress the activation of the ICP0 pro-moter by VP16. Since Luman activates CRE-containing promot-ers, including that for ICP0 (22), we compared the effects of two mutant forms of Luman on VP16-induced activation: LuS221Op, a truncated form of Luman, which does not possess a transmem-brane domain, is not retained in the endoplasmic reticulum, and is therefore constitutively active, and LuS221Op (N160G), a trun-cated form of Luman that does not bind DNA (36) and is tran-scriptionally inactive. Both mutants bind HCF and would be ex-pected to compete for it with VP16 in cells in which HCF was limiting. We observed no inhibitory effect of LuS221Op or LuS221Op (N160G) on the VP16-induced activation of the ICP0 promoter (Fig. 4A, lanes 3 and 4). In contrast, Zhangfei inhibited activation of the ICP0 promoter, reducing it by 70% (Fig. 4A, lane 5).
[image:4.585.43.283.64.477.2]In an alternative approach to determine if Zhangfei sup-pressed VP16 by depriving it of HCF, we attempted to coun-teract the effects of Zhangfei by supplementing functional
[image:4.585.318.522.73.315.2]FIG. 1. Zhangfei is expressed in neurons. Zhangfei was detected in human tissues using antiserum against Zhangfei (anti-ZF). Serum from which antibodies against Zhangfei had been removed (absorbed) was used as a negative control. (A) Lysates of cells transfected to express Zhangfei (lanes 1) or a negative control (lanes 2) were sepa-rated by SDS-polyacrylamide gel electrophoresis and immunoblotted with anti-ZF or absorbed serum. Sections of a parietal lobe (B) and sensory ganglion (C) stained with anti-ZF or absorbed serum are shown. Bar, 50m.
FIG. 2. Zhangfei inhibits VP16 activation of the ICP0 promoter. (A) Vero cells were transfected with 100 ng of pAB5 (CAT reporter with ICP0 promoter) and with 50 ng of a plasmid expressing VP16 and increasing amounts (1 to 4g) of plasmids expressing Zhangfei (cir-cles) or Zhangfei (Y224A) (squares). The results are expressed as a percentage of CAT activity in cells cotransfected with plasmid express-ing VP16 and the CAT reporter pAB5. (B) Expression of Zhangfei was confirmed by immunoblotting using polyclonal serum against Zhangfei.
on November 8, 2019 by guest
http://jvi.asm.org/
HCF in the cell. A plasmid expressing HCF was introduced into cells along with VP16, the ICP0 reporter, and increasing amounts of pcZF. Expression of additional HCF in cells had no effect on the ability of Zhangfei to suppress VP16 (Fig. 4B), suggesting that Zhangfei did not exert its effect by depriving VP16 of HCF.
[image:5.585.80.508.69.405.2]Zhangfei suppresses VP16-mediated promoter activation only when VIC formation is required.VP16 possesses a potent activation domain and can activate reporter genes that do not contain TAATGARAT motifs as long as it can bind their promoters. For instance, chimerical molecules comprising the DNA-binding domain of yeast Gal4 and the entire VP16, or just its activation domain, can bind and activate promoters containing Gal4 response elements. In contrast to VP16-me-diated activation of TAATGARAT-containing promoters, this activation does not rely on HCF or Oct-1. To determine if the suppression by Zhangfei was specific for interactions that involve the formation of VIC, we examined its effects on promoters containing Gal4 UAS or TAATGARAT motifs.
Figure 5 shows that, while Zhangfei suppressed the activation of the TAATGARAT-containing ICP0 promoter by both VP16 and VP16, it had relatively little effect on the Gal4-VP16-mediated activation of pG5EC, a promoter containing five Gal4-binding elements.
Zhangfei may reduce VIC-DNA binding by sequestering VP16.Since the previous experiment suggested that the effect of Zhangfei was limited to interactions where promoter rec-ognition by VP16 required HCF and the formation of VIC, we determined if Zhangfei could reduce VIC formation. HeLa cells were mock infected or infected with adenoviruses express-ing-galactosidase, Zhangfei, or Zhangfei (Y224A) at an MOI that would ensure infection of all cells in the culture. After this, nuclear extracts were prepared from these cultures and incu-bated with a radioactively labeled double-stranded oligonucle-otide containing TAATGARAT. These extracts (Fig. 6, lanes 1, 3, 4, and 5) formed the Oct-1-containing complex, but since they did not contain VP16, they did not retard the migration of the probe to the extent corresponding to the formation of VIC.
FIG. 3. Zhangfei inhibits ICP0 transcript and protein expression and reduces the yield of progeny virus in HSV-1-infected cells. Hep2 cells were infected with adenovirus vectors expressing-galactosidase (lanes and panel 1), Zhangfei (Y224A) (lanes and panel 2), or Zhangfei (lanes and panel 3). Twenty-four hours later the cells were also infected with HSV-1. (A) Northern blot analysis of total RNA, which was harvested from cells probed with radiolabeled ICP0 DNA. The lower panel shows the intensity of ethidium bromide-stained 18S and 28S rRNA bands and indicates that equal amounts of RNA were loaded for each sample. (B) Hep2 cells were stained for immunofluorescence using a monoclonal antibody specific to ICP0. Total cells as well as fluorescent cells in four 400⫻microscope fields were counted in four independent experiments. (C) Cells expressing ICP0 are shown as percentage of total cells. Bars represent standard deviations of means. (D) VP16, ICP0, and Zhangfei in the samples were detected by immunoblotting. (E) HSV-1 yields by cells expressing-galactosidase, Zhangfei (Y224A), or Zhangfei 24 h after infection with HSV-1.
on November 8, 2019 by guest
http://jvi.asm.org/
All extracts led to equivalent amounts of Oct-1-containing complex. In contrast, nuclear extracts from HSV-1-infected HeLa cells induced the Oct-1 complex as well as VIC (lane 2). When a virion extract containing VP16 was added to the nu-clear extracts, they all led to the formation of the VIC (lanes 6, 7, and 8). However, lysates of cells expressing Zhangfei or Zhangfei (Y224A) contained significantly less VIC or Oct-1 complex (Fig. 6, compare lanes 7 and 8 with lane 6). These results suggested that Zhangfei-expressing cells contained an activity that could suppress VP16 and Oct-1 interactions with the TAATGARAT sequence.
[image:6.585.301.542.70.215.2]To determine if Zhangfei suppressed VIC formation by binding to VP16, we immunoprecipitated VP16 from lysates of cells expressing either VP16 alone or VP16 in conjunction with Zhangfei or Zhangfei (Y224A). All three lysates contained equivalent amounts of VP16, as determined by immunoblot-ting (not shown). The immunoprecipitates were then examined
[image:6.585.73.253.72.324.2]FIG. 4. Zhangfei does not deprive VP16 of functional HCF. (A) Vero cells were transfected with 100 ng of pAB5 (CAT reporter with ICP0 promoter) (lanes 1 to 5) or 50 ng of a plasmid expressing VP16 (lanes 2 to 5) and cotransfected with plasmids expressing either LuS221Op (lane 3), LuS221Op (N160G) (lane 4), or Zhangfei (lane 5). Results are expressed as percentages of CAT activity without LuS221Op, LuS221Op (N160G), or Zhangfei. (B) Vero cells were transfected with 100 ng of pAB5 and 50 ng of a plasmid expressing VP16. Increasing amounts of pcZF (0 to 4g) were cotransfected along with pcDNA (squares) or a plasmid expressing HCF (circles). Results are expressed as percentages of CAT activity with no Zhangfei.
FIG. 7. Zhangfei forms a complex with VP16. Lysates from Vero cells transfected as indicated in chart above (lanes 1 to 6) were immu-noprecipitated with antibodies against VP16. The immunoprecipitates were separated by SDS-polyacrylamide gel electrophoresis and trans-ferred to a nitrocellulose membrane. Zhangfei on the membrane was detected by immunoblotting using anti-Zhangfei serum. Lanes 7 and 8 (boxed) contain lysates of cells expressing Zhangfei or Zhangfei (Y224A) electrophoresed on the same gel as controls.
[image:6.585.51.278.477.640.2]FIG. 5. Zhangfei suppresses VIC-dependent but not VIC-indepen-dent activation by VP16. Vero cells were transfected with pAB5 or pG5EC, a reporter with five Gal4 UASs. Cells were cotransfected with increasing amounts of pcZF (1, 2, and 4g) and activator plasmids encoding either VP16 or Gal4-VP16. Squares represent the effect of Zhangfei on pG5EC activated by Gal4-VP16. The effect of Zhangfei on the activation of the ICP0 promoter by VP16 is represented by diamonds, and the effect on activation by Gal4-VP16 is represented as circles.
FIG. 6. Effect of Zhangfei on VP16-induced complex (VIC). Five micrograms of HeLa nuclear extracts was incubated with 5 ng of
32P-labeled DNA probe representing an ICP0 TAATGARAT motif.
Samples were separated on a 5% nondenaturing polyacrylamide gel. To some nuclear extracts an HSV-1 lysate containing VP16 was added (lanes 6, 7, and 8).
on November 8, 2019 by guest
http://jvi.asm.org/
[image:6.585.303.544.510.649.2]by immunoblot assays using anti-Zhangfei serum. Figure 7 shows that, in cells expressing both VP16 and Zhangfei, or its mutant Zhangfei (Y224A), VP16 was bound to Zhangfei (Fig. 7, lanes 5 and 6).
The VP16-Zhangfei complex contains additional cellular proteins.To determine if Zhangfei and VP16 formed a complex, we synthesized the proteins, either on their own or together, in an in vitro transcription-translation-coupled system. VP16 was then precipitated from this mixture, and Zhangfei was detected in the immunoprecipitate by immunoblotting with anti-Zhangfei serum. While the anti-VP16 antibody precipitated VP16 from lysates that contained it, we were unable to detect Zhangfei in the immuno-precipitate from a mixture of the two proteins (data not shown). This suggested that VP16 and Zhangfei do not interact directly with each other. To determine if other cellular proteins partici-pated in the interaction, we separated immunoprecipitates from cells expressing VP16 alone or VP16 plus Zhangfei by two-di-mensional electrophoresis. The gels were stained for peptides using a sensitive silver stain. In the immunoprecipitates from cells expressing both proteins we consistently observed three spots that were not present when the immunoprecipitates lacked Zhangfei (Fig. 8).
Microarray analysis.While the expression of most cellular genes is reduced in HSV-1-infected cells, the virus increases the expression of cellular components that assist in the repli-cative process. To determine if Zhangfei has an effect on any of these genes, we compared the levels of transcripts for 19,000 genes in cells expressing either-galactosidase, Zhangfei, or Zhangfei (Y224A). Hep2 cells were infected with adenovirus vectors expressing one of the three proteins followed a day later by HSV-1. Total RNA was extracted from the cultures 6 h later and analyzed on microarrays. Each experiment was re-peated twice.
A significance analysis of microarrays (43) revealed that levels of transcripts from 16 genes were significantly reduced in Zhangfei-expressing cells compared with -galactosidase- or
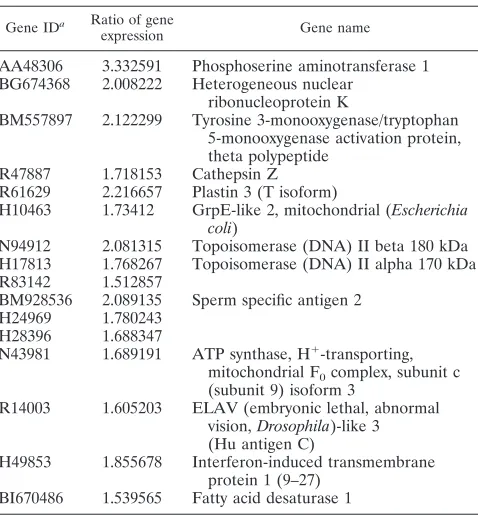
[image:7.585.87.501.71.272.2]Zhangfei (Y224A)-expressing HSV-1-infected cells. A list of these genes is shown in Table 1. A search of the literature showed that one of these genes, DNA topoisomerase II␣(p170 form), directly participates in HSV-1 DNA replication and late gene expression (1, 5). To confirm that topoisomerase II␣was indeed reduced in Zhangfei-expressing HSV-1-infected cells, we stained cells expressing -galactosidase, Zhangfei, or
FIG. 8. The VP16-Zhangfei complex contains other cellular proteins. Results of two-dimensional gel analysis of VP16 immunoprecipitated from cells expressing either VP16 (A) or VP16 as well as Zhangfei (B) are shown. Circles in panel B are spots seen only in immunoprecipitates of cells that expressed both VP16 and Zhangfei (two replicates). Numbers at left are molecular masses in kilodaltons.
TABLE 1. Genes down-regulated in Zhangfei-expressing HSV-infected cells
Gene IDa Ratio of gene
expression Gene name
AA48306 3.332591 Phosphoserine aminotransferase 1 BG674368 2.008222 Heterogeneous nuclear
ribonucleoprotein K
BM557897 2.122299 Tyrosine 3-monooxygenase/tryptophan 5-monooxygenase activation protein, theta polypeptide
R47887 1.718153 Cathepsin Z R61629 2.216657 Plastin 3 (T isoform)
H10463 1.73412 GrpE-like 2, mitochondrial (Escherichia coli)
N94912 2.081315 Topoisomerase (DNA) II beta 180 kDa H17813 1.768267 Topoisomerase (DNA) II alpha 170 kDa R83142 1.512857
BM928536 2.089135 Sperm specific antigen 2 H24969 1.780243
H28396 1.688347
N43981 1.689191 ATP synthase, H⫹-transporting, mitochondrial F0complex, subunit c
(subunit 9) isoform 3
R14003 1.605203 ELAV (embryonic lethal, abnormal vision,Drosophila)-like 3 (Hu antigen C)
H49853 1.855678 Interferon-induced transmembrane protein 1 (9–27)
BI670486 1.539565 Fatty acid desaturase 1
aID, identification.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:7.585.302.541.458.716.2]Zhangfei (Y224A) for topoisomerase II␣. Figure 9A shows that Zhangfei reduced the number of cells in the culture ex-pressing topoisomerase II␣. In contrast, Zhangfei (Y224A) had no effect. We also transfected cells to express Zhangfei or Zhangfei (Y224A) and stained cells for Zhangfei as well as topoisomerase II␣. In four 20⫻fields examined few Zhangfei-expressing cells also stained for topoisomerase II␣ while
Zhangfei (Y224A) was also present in many cells that also stained for topoisomerase II␣(Fig. 9B).
DISCUSSION
[image:8.585.98.488.66.584.2]Lytic infection in HSV-1-infected epithelial cells proceeds according to a well-characterized sequence of gene expression
FIG. 9. Effect of Zhangfei on the expression of topoisomerase II␣. (A) Hep2 cells were infected with adenovirus vectors expressing -galac-tosidase (Adeno-gal), Zhangfei (Adeno-ZF), or Zhangfei (Y224A) [Adeno-ZF(Y224A)]. The cells were then fixed and stained for topoisomerase II␣. The percentage of total cells expressing topoisomerase II␣is shown, with standard deviations from the means of two experiments indicated. (B) Cells were transfected with plasmids expressing either Zhangfei or Zhangfei (Y224A) and stained for Zhangfei (green) and topoisomerase II␣(red).
on November 8, 2019 by guest
http://jvi.asm.org/
that begins with the initiation of transcription of IE genes by VP16 and the cellular proteins HCF and Oct-1. However, little is known about how this program is suppressed in neurons, leading to the establishment of latency.
We show that neurons in sensory ganglia (Fig. 1C), the site of HSV-1 latency, contain an HCF-binding protein that can suppress the activity of VP16 and also modulate the expression of other genes required for viral replication. We have previ-ously shown that this protein, Zhangfei, in an HCF-dependent manner, can suppress HSV gene expression (22) and can also suppress the ability of Luman/LZIP/CREB3 (30), a protein capable of activating IE gene expression (25).
Here we show that, in cells transiently expressing Zhangfei and VP16, Zhangfei inhibited the ability of VP16 to activate the promoter for the IE protein ICP0 (Fig. 2). HCF appeared to be required for optimal expression, since a mutant of Zhangfei impaired for binding HCF was less efficient. Depen-dence of the activity of a protein on HCF can be demonstrated by showing that there is a decrease in activity in the absence of
available HCF. This can be done either by using cells with a conditional lesion in HCF (8) or by sequestering the protein in the cytoplasm (16, 30). However, since Zhangfei’s target, VP16, also requires HCF, it is difficult to use this criterion for showing that Zhangfei requires HCF to suppress VP16. In any case the dependence of Zhangfei on HCF was not absolute since Zhangfei (Y224A), which has a mutant HCF-binding motif, also suppressed VP16, albeit to a lesser extent.
[image:9.585.111.475.66.430.2]Consistent with our results with cells transiently expressing Zhangfei and VP16, Zhangfei also suppressed the amounts of ICP0 RNA and protein in cells infected with HSV-1 (Fig. 3A and B). Possibly as a consequence, the expression of a late viral protein and progeny virus yields were also suppressed in these cells (Fig. 3D and E). Interestingly, while Zhangfei suppressed ICP0 expression in cells infected with HSV-1 at both low and high multiplicities of infection, it had no effect on virus yields when high multiplicities of infection were used. This is consis-tent with observations that at high multiplicities of infection the requirement for ICP0 decreases (4, 13, 39).
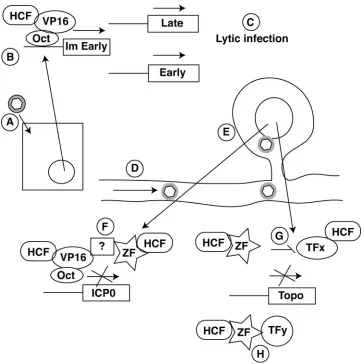
FIG. 10. Model for the potential role of Zhangfei in the establishment of HSV-1 latency in sensory neurons. (A) HSV-1 infects an epithelial cell. (B and C) Lytic infection (C) is initiated by the assembly of VIC comprising VP16, HCF, and Oct-1 on TAATGARAT elements on IE promoters. (D) The HSV-1 nucleocapsid along with tegument protein (gray halo around hexagon), possibly including VP16, is transported to the neuronal nucleus by retrograde transport. (E and F). In the neuronal nucleus (E), Zhangfei inhibits VIC formation by interacting with VP16 in an HCF-dependent manner (F). (G and H). Zhangfei also suppresses topoisomerase II␣expression either in an HCF-dependent manner (G) or in an HCF-independent manner (H) by suppressing cellular transcription factors.
on November 8, 2019 by guest
http://jvi.asm.org/
Zhangfei suppressed the ability of VP16 to form a complex with Oct-1 and an oligonucleotide containing the TAATG ARAT element from the ICP0 promoter (Fig. 6). A simple explanation for this might be that Zhangfei binds and seques-ters VP16, preventing it from assembling as VIC. Indeed, we found that an antibody against VP16 immunoprecipitated Zhangfei from cells expressing both Zhangfei and VP16 (Fig. 7). However, we were unable to show that VP16 and Zhangfei interact directly with each other, at least when they are syn-thesized together in vitro. In addition, since Zhangfei inhibited only the ability of Gal4-VP16 to activate a TAATGARAT-containing promoter while having no effect on its ability to activate a Gal4 UAS-containing promoter (Fig. 5), it is possible that Zhangfei influences only VP16 (or Gal4-VP16), and indi-rectly associates with it, when it is a component of VIC. This is consistent with our observation that, while Zhangfei had no effect on the Oct-1 and TAATGARAT complex in nuclear extracts that did not contain VP16 (Fig. 6, compare lanes 3 and 4), it reduced both VIC and the Oct-1 complex when VP16 was present (Fig. 6, compare lanes 6 and 7). It is also consistent with the presence of other cellular proteins in the immunopre-cipitate (Fig. 8).
Finally, we show that, in addition to inhibiting the ability of VP16 to activate the transcription of ICP0, Zhangfei also sup-pressed the expression of cellular proteins that are normally induced following HSV-1 infection (Table 1) (Fig. 9). At least one of these, topoisomerase II␣, is required for HSV-1 DNA replication and late gene expression. Several topoisomerase II␣binding sites were identified at HSV-1 DNA (5), and the enzyme has been shown to participate in HSV-1 DNA repli-cation and late gene expression (1).
Following replication in epithelial cells, HSV-1 capsids are delivered by retrograde transport to the cell bodies of sensory neurons. While it is not clear which of the tegument proteins make it to the neuronal nucleus, transport of the viral capsid along axonal microtubules does require at least some of the tegument proteins (3, 28). In the neuron the decision to pursue lytic or latent infection appears to be made at an early stage, with latency as a consequence of suppression of viral IE gene expression (reviewed in references 6 and 35). If VP16 indeed accompanies the viral nucleocapsid to the neuronal nucleus, mechanisms must be in place to suppress its activity. Several hypotheses have been proposed to account for the suppression of VP16-mediated IE gene expression. The neuron appears to have a high Oct-2-to-Oct-1 ratio (9, 10, 15, 20). While both proteins can bind TAATGARAT motifs, only Oct-1 has struc-tural elements required for associating with the VP16-HCF complex. It has been suggested that Oct-2 sterically hinders occupation of the TAATGARAT sites by Oct-1, thereby pre-venting IE gene expression. A lack of available HCF in the neuronal nucleus has also been suggested as a mechanism for the suppression of VP16-mediated IE gene expression. Kristie and others (18) demonstrated that, unlike in other cells, HCF is present in the cytoplasm of unstressed neurons. However, such scenarios, involving the lack of appropriate Oct-1 and HCF, would be relevant only for suppressing IE gene expression if VP16 was delivered to the neuron along with the viral genome.
Based on our observations we propose a model for the ability of Zhangfei to suppress HSV-1 replication in the neu-ron (Fig. 10). Zhangfei is expressed in sensory neuneu-rons that are
the site of HSV-1 latency. The HSV-1 capsid, in association with VP16 and other tegument proteins, is delivered to the neuronal nucleus where Zhangfei actively suppresses the abil-ity of VP16 to activate IE gene expression. In neurons where lytic replication proceeds beyond IE expression, Zhangfei has a further suppressive effect on subsequent stages of the viral lytic cycle by inhibiting cellular proteins such as topoisomerase II␣that are required for these steps.
Our results for interactions between Zhangfei and VP16 lead us to hypothesize that Zhangfei is an important regulator of the stress response in neurons. An understanding of how it influences viral and cellular molecules will not only tell us how herpesviruses use host pathways to suppress their replication and establish latency but also shed light on how neurons might respond to stress.
ACKNOWLEDGMENTS
This study was supported by an operating grant from the Natural Sciences and Engineering Research Council (NSERC) of Canada to V.M. O.A. was supported by a postdoctoral fellowship from the Saskatchewan Health Research Foundation.
REFERENCES
1.Advani, S. J., R. R. Weichselbaum, and B. Roizman.2003. Herpes simplex virus 1 activates cdc2 to recruit topoisomerase II alpha for post-DNA
syn-thesis expression of late genes. Proc. Natl. Acad. Sci. USA100:4825–4830.
2.Ajuh, P., J. Chusainow, U. Ryder, and A. I. Lamond.2002. A novel function for human factor C1 (HCF-1), a host protein required for herpes simplex
virus infection, in pre-mRNA splicing. EMBO J.21:6590–6602.
3.Bearer, E. L., X. O. Breakefield, D. Schuback, T. S. Reese, and J. H. LaVail.
2000. Retrograde axonal transport of herpes simplex virus: evidence for a single mechanism and a role for tegument. Proc. Natl. Acad. Sci. USA
97:8146–8150.
4.Cai, W., and P. A. Schaffer.1992. Herpes simplex virus type 1 ICP0 regulates expression of immediate-early, early, and late genes in productively infected
cells. J. Virol.66:2904–2915.
5.Ebert, S. N., S. S. Shtrom, and M. T. Muller. 1990. Topoisomerase II cleavage of herpes simplex virus type 1 DNA in vivo is replication dependent.
J. Virol.64:4059–4066.
6.Efstathiou, S., and C. M. Preston.2005. Towards an understanding of the
molecular basis of herpes simplex virus latency. Virus Res.111:108–119.
7.Elliott, G., and P. O’Hare.1999. Live-cell analysis of a green fluorescent
protein-tagged herpes simplex virus infection. J. Virol.73:4110–4119.
8.Goto, H., S. Motomura, A. C. Wilson, R. N. Freiman, Y. Nakabeppu, K. Fukushima, M. Fujishima, W. Herr, and T. Nishimoto.1997. A single-point mutation in HCF causes temperature-sensitive cell-cycle arrest and disrupts
VP16 function. Genes Dev.11:726–737.
9.Hagmann, M., O. Georgiev, W. Schaffner, and P. Douville.1995. Transcrip-tion factors interacting with herpes simplex virus alpha gene promoters in
sensory neurons. Nucleic Acids Res.23:4978–4985.
10.He, X., M. N. Treacy, D. M. Simmons, H. A. Ingraham, L. W. Swanson, and M. G. Rosenfeld.1989. Expression of a large family of POU-domain
regu-latory genes in mammalian brain development. Nature340:35–41.
11.Herrera, F. J., and S. J. Triezenberg.2004. VP16-dependent association of chromatin-modifying coactivators and underrepresentation of histones at immediate-early gene promoters during herpes simplex virus infection. J.
Vi-rol.78:9689–9696.
12.Hughes, T. A., S. La Boissiere, and P. O’Hare.1999. Analysis of functional domains of the host cell factor involved in VP16 complex formation. J. Biol.
Chem.274:16437–16443.
13.Jordan, R., and P. A. Schaffer.1997. Activation of gene expression by herpes simplex virus type 1 ICP0 occurs at the level of mRNA synthesis. J. Virol.
71:6850–6862.
14.Julien, E., and W. Herr.2003. Proteolytic processing is necessary to separate and ensure proper cell growth and cytokinesis functions of HCF-1. EMBO J.
22:2360–2369.
15.Kemp, L. M., C. L. Dent, and D. S. Latchman.1990. Octamer motif mediates transcriptional repression of HSV immediate-early genes and
octamer-con-taining cellular promoters in neuronal cells. Neuron4:215–222.
16.Khurana, B., and T. M. Kristie.2004. A protein sequestering system reveals control of cellular programs by the transcriptional coactivator HCF-1.
J. Biol. Chem.279:33673–33683.
17.Kristie, T. M., J. L. Pomerantz, T. C. Twomey, S. A. Parent, and P. A. Sharp.
1995. The cellular c1 factor of the herpes simplex virus enhancer complex is
a family of polypeptides. J. Biol. Chem.270:4387–4394.
on November 8, 2019 by guest
http://jvi.asm.org/
18.Kristie, T. M., J. L. Vogel, and A. E. Sears.1999. Nuclear localization of the C1 factor (host cell factor) in sensory neurons correlates with reactivation of
herpes simplex virus from latency. Proc. Natl. Acad. Sci. USA96:1229–1233.
19.La Boissiere, S., T. Hughes, and P. O’Hare.1999. HCF-dependent nuclear
import of VP16. EMBO J.18:480–489.
20.Lillycrop, K. A., C. L. Dent, S. C. Wheatley, M. N. Beech, N. N. Ninkina, J. N. Wood, and D. S. Latchman.1991. The octamer-binding protein Oct-2 re-presses HSV immediate-early genes in cell lines derived from latently
in-fectable sensory neurons. Neuron7:381–390.
21.Liu, Y., W. Gong, C. C. Huang, W. Herr, and X. Cheng.1999. Crystal structure of the conserved core of the herpes simplex virus transcriptional
regulatory protein VP16. Genes Dev.13:1692–1703.
22.Lu, R., and V. Misra.2000. Potential role for Luman, the cellular homologue
of herpes simplex virus VP16 (␣genetrans-inducing factor), in herpesvirus
latency. J. Virol.74:934–943.
23.Lu, R., and V. Misra.2000. Zhangfei: a second cellular protein interacts with herpes simplex virus accessory factor HCF in a manner similar to Luman and
VP16. Nucleic Acids Res.28:2446–2454.
24.Lu, R., P. Yang, P. O’Hare, and V. Misra.1997. Luman, a new member of the CREB/ATF family, binds to herpes simplex virus VP16-associated host
cel-lular factor. Mol. Cell. Biol.17:5117–5126.
25.Lu, R., P. Yang, S. Padmakumar, and V. Misra. 1998. The herpesvirus transactivator VP16 mimics a human basic domain leucine zipper protein,
Luman, in its interaction with HCF. J. Virol.72:6291–6297.
26.Luciano, R. L., and A. C. Wilson.2002. An activation domain in the C-terminal subunit of HCF-1 is important for transactivation by VP16 and
LZIP. Proc. Natl. Acad. Sci. USA99:13403–13408.
27.Luciano, R. L., and A. C. Wilson.2003. HCF-1 functions as a coactivator for
the zinc finger protein Krox20. J. Biol. Chem.278:51116–51124.
28.Luxton, G. W., S. Haverlock, K. E. Coller, S. E. Antinone, A. Pincetic, and G. A. Smith.2005. Targeting of herpesvirus capsid transport in axons is coupled to association with specific sets of tegument proteins. Proc. Natl.
Acad. Sci. USA102:5832–5837.
29.Mahajan, S. S., and A. C. Wilson.2000. Mutations in host cell factor 1 separate its role in cell proliferation from recruitment of VP16 and LZIP.
Mol. Cell. Biol.20:919–928.
30.Misra, V., N. Rapin, O. Akhova, M. Bainbridge, and P. Korchinski.2005. Zhangfei is a potent and specific inhibitor of the host cell factor-binding
transcription factor luman. J. Biol. Chem.280:15257–15266.
31.Misra, V., S. Walker, S. Hayes, and P. O’Hare.1995. The bovine herpesvirus
␣genetrans-inducing factor activates transcription by mechanisms different from those of its herpes simplex virus type 1 counterpart VP16. J. Virol.
69:5209–5216.
32.Mossman, K. L., and J. R. Smiley.1999. Truncation of the C-terminal acidic transcriptional activation domain of herpes simplex virus VP16 renders
ex-pression of the immediate-early genes almost entirely dependent on ICP0.
J. Virol.73:9726–9733.
33.Narayanan, A., M. L. Nogueira, W. T. Ruyechan, and T. M. Kristie.2005. Combinatorial transcription of herpes simplex virus and varicella zoster virus immediate early genes is strictly determined by the cellular coactivator
HCF-1. J. Biol. Chem.280:1369–1375.
34.Newman, J. R., and A. E. Keating.2003. Comprehensive identification of
human bZIP interactions with coiled-coil arrays. Science300:2097–2101.
35.Preston, C. M.2000. Repression of viral transcription during herpes simplex
virus latency. J. Gen. Virol.81:1–19.
36.Raggo, C., N. Rapin, J. Stirling, P. Gobeil, E. Smith-Windsor, P. O’Hare, and V. Misra.2002. Luman, the cellular counterpart of herpes simplex virus VP16, is processed by regulated intramembrane proteolysis. Mol. Cell. Biol.
22:5639–5649.
37.Roizman, B.1995. Herpesviridae, p. 2221–2230.InB. N. Fields, D. M. Knipe, and P. M. Howley (ed.), Fields virology, 3rd ed., vol. 2. Lippincott-Raven, Philadelphia, Pa.
38.Roizman, B., and A. E. Sears.1995. Herpes simplex viruses and their
repli-cation, p. 2231–2296.InB. N. Fields, D. M. Knipe, and P. M. Howley (ed.),
Fields virology, 3rd ed., vol. 2. Lippincott-Raven, Philadelphia, Pa. 39.Samaniego, L. A., L. Neiderhiser, and N. A. DeLuca.1998. Persistence and
expression of the herpes simplex virus genome in the absence of
immediate-early proteins. J. Virol.72:3307–3320.
40.Sambrook, J., and D. W. Russell.2001. Molecular cloning: a laboratory manual, 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
41.Shevchenko, A., M. Wilm, O. Vorm, and M. Mann.1996. Mass spectrometric
sequencing of proteins silver-stained polyacrylamide gels. Anal. Chem.68:
850–858.
42.Stojanova, A., C. Caro, R. J. Jarjour, S. K. Oster, L. Z. Penn, and R. J. Germinario.2004. Repression of the human immunodeficiency virus type-1
long terminal repeat by the c-Myc oncoprotein. J. Cell. Biochem.92:400–
413.
43.Tusher, V. G., R. Tibshirani, and G. Chu.2001. Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci.
USA98:5116–5121.
44.Wilson, A. C., R. N. Freiman, H. Goto, T. Nishimoto, and W. Herr.1997. VP16 targets an amino-terminal domain of HCF involved in cell cycle
pro-gression. Mol. Cell. Biol.17:6139–6146.
45.Wilson, A. C., K. LaMarco, M. G. Peterson, and W. Herr.1993. The VP16 accessory protein HCF is a family of polypeptides processed from a large
precursor protein. Cell74:115–125.
46.Wysocka, J., M. P. Myers, C. D. Laherty, R. N. Eisenman, and W. Herr.2003. Human Sin3 deacetylase and trithorax-related Set1/Ash2 histone H3-K4 methyltransferase are tethered together selectively by the cell-proliferation
factor HCF-1. Genes Dev.17:896–911.