upregulate the costimulatory molecule CD154 is at the core of this dysregulation. To test the hypothesis that increased expression of CD154 on infected CD4ⴙT cells could modulate immune function, we constructed a replication-competent simian immunodeficiency virus (SIV) vector that expressed CD154. We found that this recombinant vector directed the expression of CD154 on the surface of infected CD4ⴙ T cells and that expression of CD154 resulted in activation of B cells present in the same cultures. Experimental infection of rhesus macaques resulted in very low viral loads for the CD154-expressing virus and the control virus, indicating that expression of CD154 did not result in increased viral replication. Analyses of the anti-SIV immune responses and the phenotype of lymphocytes in blood and lymphoid tissues showed changes that occurred during the acute phase of infection only in animals infected with the CD154-expressing SIV, but that became indistinguishable from those seen in animals infected with the control virus at later time points. We conclude that the level of expression of CD154 in itself is not responsible for affecting the immune response to an attenuated virus. Considering that the CD154-expressing SIV vector and the virus control did not carry an activenefgene, our results suggest that, in CD4ⴙT cells infected with wild-type virus, Nef is the viral factor that interferes with the immune mechanisms that regulate expression of CD154.
The CD154 protein, also known as CD40 ligand (CD40L), is a type II membrane glycoprotein of 39 kDa with an extracel-lular domain homologous to tumor necrosis factor alpha (TNF-␣) and -(1). This protein is expressed transiently on CD4⫹T cells after activation in vitro or in vivo and weakly on CD8⫹T cells after in vitro activation with anti-CD3 or phorbol myristate acetate (PMA)-ionomycin (2). CD154 binds to CD40, a 50-kDa member of the TNF receptor superfamily expressed on several cells of the immune system, including immature and mature B lymphocytes, monocytes, interdigitat-ing dendritic cells (IDCs) in the T-cell areas of secondary lymphoid organs, follicular dendritic cells (FDCs), and thymic epithelium (45). CD154-CD40 interactions are required for the priming and expansion of antigen-specific CD4⫹T cells and CD8⫹cytotoxic T lymphocytes (CTLs) and in the induc-tion of costimulatory activity on antigen-presenting cells (APCs) (4–6). CD154-CD40 interactions are also critical in B-cell proliferation, immunoglobulin (Ig) production, Ig class switching, rescue of B cells from apoptotic death, germinal center formation, and generation of B-cell memory (7). Cell-cell contact via CD40-CD154 is required for macrophages to be activated and produce nitric oxide and interleukin-12 (IL-12) (8). The importance of CD154 expression in vivo was revealed by studies of patients suffering from hyper-IgM syn-drome (HIGM1), a human X-linked immunodeficiency caused
by mutations in the CD154 gene that result in lack of func-tional expression of CD154 on the surface of activated T cells (9). HIGM1 patients suffer from recurrent upper respiratory tract infections and certain opportunistic infections such as cryptosporidial diarrhea andPneumocystis cariniipneumonia. Studies of CD154-deficient mice have shown that their CD4⫹ T cells proliferate very poorly in vivo in response to protein antigens; however, in vitro proliferation in response to poly-clonal activators was not affected (23).
CD154-CD40 contacts are important for the development of immune responses to viral infections, although innate defense mechanisms, some CD8⫹CTL responses, and short-lived neu-tralizing antibody responses can be induced in a CD154-CD40-independent fashion (22). However, there is in vivo and in vitro evidence that CD154 signaling is dysregulated during human immunodeficiency virus (HIV) infection (9, 29). For example, early in HIV-1 infection, B-cell responses to T-cell-dependent antigens are impaired, and this deficiency has been associated with a defective T-cell function (36, 48). Among the APC functions that have been reported to be defective in HIV infection is a marked impairment of production of IL-12 by peripheral blood mononuclear cells (PBMCs) and macro-phages from HIV-infected patients (7), which can be corrected by addition of soluble trimeric CD154 (10, 42). Similarly, re-duced pathogen-specific CD154 expression on CD4⫹T cells has been observed in HIV-infected individuals (41). Finally, there are striking similarities between opportunistic infections and hypergammaglobulinemias seen in HIGM1 and HIV-in-fected patients. The association of HIV gp120 with CD4 has been implicated in the reduced expression of CD154 and the consequent reduced costimulation activities of the APCs (20, 21).
* Corresponding author. Mailing address: Southwest Foundation for Biomedical Research, 7620 NW Loop 410, San Antonio, TX 78227. Phone: (210) 258-9603. Fax: (210) 670-3310. E-mail: lgiavedo@sfbr .org.
† Present address: Centro Nacional de Rehabilitacio´n, Xochimilco, C.P.14389, Mexico.
4679
on November 8, 2019 by guest
In this study, we report for the first time the construction of a replication-competent SIV vector that expresses a biologi-cally active CD154 molecule. We demonstrate that CD154 is presented on the surface of infected CD4⫹T cells and that it can stimulate neighboring B cells. Finally, we show that infec-tion of rhesus macaques with this virus results in early and transient phenotypic changes that do not affect, in the long term, the outcome of the antiviral immune response.
MATERIALS AND METHODS
Cloning of the human and rhesus CD154 genes.The CD154 gene was cloned from RNA from human or rhesus macaque PBMCs stimulated by plating cells on plastic flasks coated with anti-CD3 antibodies (clone UCHT1 from Beckman-Coulter for human PBMCs, and clone FN-18 from U-Cytech for rhesus PBMCs). After 6 h of stimulation, cells were harvested and lysed with 1 ml of Trizol in order to extract intact RNA cellular. A first-strand cDNA was prepared with oligo(dT) (Amersham, Piscataway, N.J.) and Superscript II reverse transcriptase (Invitrogen, Carlsbad, Calif.). Two sets of primers were designed based on the
human CD154 sequence: (i) primers FCD154 (5⬘-AGAAGATACCATgTCGA
CTTTAACACAGCATG-3⬘) and RCD154 (5⬘-ACCACAGCCTGCAAGTCgaC
ACTGTTCAGA-3⬘), which bind outside the coding sequence, were used to
amplify the full-length gene; and (ii) primers sFCD154 (5⬘-ATGATTGGGTcga
CACTTTTTGCTaTGTAT-3⬘) and RCD154 were used to amplify the
extracel-lular domain of the CD154 protein. These primers contained mutations (in italic lowercase) that introduced a SalI cloning site (underlined). The PCR products were isolated from agarose gel and cloned into the pCR 2.1 plasmid (Invitrogen), and three different clones were sequenced. Finally, the human CD154 gene was purified as a SalI DNA fragment and cloned into the SalI site of pSIVHy (18).
Cells and viruses.SIVmac239, which contains anefgene with an open reading frame, was generously provided by R. Desrosiers (New England National Pri-mate Research Center, Southborough, Mass.). CEM-x-174 cells, rhesus PBMCs, and lymph node (LN) cells were used for SIV isolation and propagation. These cells were maintained in RPMI 1640 supplemented with 10% fetal calf serum, 2 mM glutamine, 0.1 mg of streptomycin per ml, and 100 U of penicillin per ml (Cellgro Mediatech, Herndon, Va.) (RPMI 10). The herpesvirus saimiri-trans-formed rhesus T-302 cell line was maintained in RPMI 10 with 25 IU of IL-2 per ml.
To prepare the recombinant viruses, the CD154 gene, as a SalI fragment, was cloned into the SalI cloning site of the plasmid pSIVHy, which contains the
whole SIVmac239 proviral genome with a deletion of thenefgene and a unique
SalI cloning site (18). Plasmids containing the CD154 gene in the sense (pSIVCD154) and antisense (pSIVCNTL) orientation were electroporated into CEM-x-174 cells to generate SIVCD154 and SIVCNTL, respectively. Virus rep-lication was monitored by the SIV core antigen capture enzyme-linked immu-nosorbent assay (ELISA) (Coulter Corporation, Hialeah, Fla.), and expression of CD154 was monitored by flow cytometry with the phycoerythrin (PE)-labeled antibody TRAP1 (Beckman-Coulter, Miami, Fla.). Viral stocks were prepared and titers were determined in CEM-x-174 cells.
In vitro analysis of retroviral infection.CEM-x-174, rhesus T-302 cells, and unstimulated PBMCs from uninfected rhesus macaques were infected at a mul-tiplicity of infection of 0.1 with SIVCD154, SIVmac239, or SIVCNTL. Expres-sion of CD154 was monitored by flow cytometry with direct staining with anti CD154-PE (clone TRAP1; Beckman-Coulter). Other markers detected were CD3 (clone FN-18, Biosource, Camarillo, Calif.), CD4 (clone OKT4; Ortho Diagnostic Systems, Raritan, N.J.), CD8 (clone 3B5; Caltag, Burlingame, Calif.), and CD20 (clone B1; Beckman-Coulter). Samples were acquired in a FACScan flow cytometer (Becton Dickinson, San Jose, Calif.), and data were analyzed with CellQuest software (Becton Dickinson Immunocytometry Systems).
Infection of rhesus macaques.Eight colony-bred, weight- and age-matched
adult, Indian female rhesus macaques (Macaca mulatta) were used in this
ex-periment. Animals were seronegative for simian type D retroviruses, simian T-cell leukemia virus, and SIV. The animals were used and cared for in accor-dance with the Association for the Assessment and Accreditation of Laboratory Animal Care, International Guidelines, and with the approval of all protocols by the Southwest Foundation for Biomedical Research Institutional Animal Care and Use Committee (IACUC). Macaques 35, 37, 42, and 44 were inoculated
intravenously (i.v.) with 1 ml of RPMI 1640 containing 104
50% tissue culture
infective doses (TCID50s) of SIVCD154. Animals 36, 38, 40, and 41 received 104
TCID50s of the control virus SIVCNTL i.v. Animals were scheduled to be
euthanized if they showed three or more of the following clinical symptoms: (i)
weight loss greater than 10% in 2 weeks or 30% in 2 months; (ii) chronic diarrhea that was unresponsive to treatment; (iii) infections that were unresponsive to antibiotic treatment; (iv) inability to maintain body heat or fluids without sup-plementation; (v) persistent, marked hematological abnormalities, including lym-phopenia, anemia, thrombocytopenia, or neutropenia; and (vi) persistent, marked splenomegaly or hepatomegaly. None of the animals involved in this experiment showed any of these signs. Peripheral LNs, axillary and/or inguinal, were obtained by transcutaneous biopsy under ketamine hydrochloride anesthe-sia (10 mg/kg of body weight injected intramuscularly; Parke-Davis, Morris Plains, N.J.) prior to and at 2, 4, and 12 weeks postinfection. Lymphocyte suspensions were obtained by mechanical teasing of tissues.
Cytokines and viral antigens in plasma.Levels of IL-12, IL-18, and alpha
interferon (IFN-␣) in plasma were determined with commercially available
ELISA kits (Cytoscreen monkey IL-12 from BioSource; Hu-IL-18 ELISA from
MBL, Nagoya, Japan; and Hu-IFN-␣ELISA from PBL, New Brunswick, N.J.).
The limits of detection were 4 pg/ml for IL-12, 12.5 pg/ml for IL-18, and 10 pg/ml
for IFN-␣. Plasma IFN (IFN-␣/) activity was also determined by measuring
inhibition of the cytopathic effect caused by encephalomyocarditis virus infection in A549 cells (23).
Cell-associated viral loads and virus isolation.Cell-associated virus, latent or productive, was measured by limiting dilution assay of cocultures of LN cells and CEM-x-174 cells in 24-well plates (24). Twice weekly, culture media were assayed for the presence of the SIV major core protein (p27) by ELISA (33). Cultures were recorded as positive for virus when p27 antigen was detected at two consecutive time points. End-point cultures were maintained and tested for 4 weeks before being scored as negative. Virus levels were calculated according to
the method of Reed and Muench (38) and expressed as TCID50/10
6
cells. Virus
isolation was performed by cocultivation of 106PBMCs with 105CEM-x-174
cells. Virus production, length of cultivation, and criterion for positivity were determined as described above for cell-associated viral loads.
Plasma viral RNA loads.The determination of viral RNA loads in heparinized plasma was performed by Quality Biological, Inc. (Gaithersburg, Md.), utilizing a nucleic acid sequence-based amplification (NASBA) assay that has a limit of detection of 2,000 genome equivalents.
Isolation of proviral DNA.Proviral DNA was extracted from LNC–CEM-x-174 cell cocultures that were positive by SIV Gag ELISA. DNA was amplified by
PCR with primers specific for the SIV 3⬘region CCGTCTGGAGATCTGCGA
CAGAGACT (forward) and GACTGAATACAGAGCGAAATGC (reverse) (27). Nucleic acids were denatured at 94°C for 1 min, annealed at 65°C for 1 min, and extended at 72°C for 2 min; this cycle was repeated 35 times. DNA fragments containing the intact CD154 gene insert were 1,160 nucleotides (nt) in length, whereas wild-type SIV originated 820-nt DNA fragments.
Lymphocyte phenotyping.Phenotypic characterization of lymphocytes in pe-ripheral blood (PB) and LNs was performed by flow cytometry with three-color direct immunofluorescence. Surface staining was performed by incubating whole blood or LN cells with monoclonal antibodies conjugated to fluorescein isothio-cyanate (FITC), phycoerythrin (PE), or Tricolor (PE-Cy5) for 30 min at room temperature. The antibodies used were anti-monkey CD3 (clone FN-18), CD4
(clone OKT4), CD8␣(clone 3B5), CD8␣(clone 2St8.5H 7), CD14 (clone
MY4), CD16 (clone 3G8), CD20 (clone B1), CD40 (clone mAb89), CD69 (clone CH/4), CD86 (clone FUN1), CD154 (clone TRAP1), and HLA-DR (clone BU63). Samples were acquired in a FACScan flow cytometer, and data were analyzed with CellQuest software. Lymphocytes were gated based on their char-acteristic forward versus side scatter pattern, and a second gate was established
by using CD3 fluorescence. CD16⫹NK cells and CD20⫹B cells were determined
in the CD3⫺lymphoid population, whereas the activation markers CD25, CD69,
and CD154 were determined in the CD4⫹and CD8⫹CD3⫹lymphoid cells.
Absolute values for cells in whole blood were obtained by combining the per-centages obtained by flow cytometry with the values of total leukocyte count per microliter and the differential formula for each animal at each time point.
Anti-SIV antibody detection.Plasma samples were analyzed for the presence of antibodies reactive to SIV envelope glycoproteins and the p27 core protein. Antigens for ELISA plates were obtained from a viral preparation of SIVmac239 concentrated by 20% sucrose cushion centrifugation. The protein content of the viral preparation was determined with the CBQCA Protein Quantification kit (Molecular Probes, Eugene, Oreg.). Anti-gp160 antibodies were quantitated with 96-well ELISA plates coated with concanavalin A and SIV gp160 as previously described (11, 39). For the anti-p27 antibody ELISA, the disrupted, envelope-depleted viral preparation was added to ELISA plates previously coated with anti-p27 antibodies (33) and incubated overnight at 4°C.
IFN-␥ELISPOT.Rhesus PBMCs were seeded at 2⫻105cells in 150l of
RPMI 10 in IFN-␥enzyme-linked immunospot (ELISPOT) plates (U-Cytech,
Utrecht, The Netherlands), and incubated with pools of 15-mer peptides that
4680 HODARA ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
encompassed the SIVmac239 Gag protein (obtained from the NIH AIDS
Re-search and Reference Reagent Program) at 2g/ml. After 18 h of incubation at
37°C, cells were lysed and plates were developed according to the manufacturer’s instructions. Spots were counted, and the difference between the number of spots for the reactive peptide minus the number of spots for the peptide control was
adjusted by the percentage of CD8⫹T cells in the samples and expressed as
spot-forming cells (SFC)/106CD8⫹T cells.
Statistical analyses.The two-samplettest for samples with equal variances was performed with Excel (Microsoft Corporation). Statistical significance was
given forP⬍0.05.
RESULTS
In vitro characterization of recombinant viruses.We cloned the human and rhesus CD154 genes from stimulated T cells. Our studies of the kinetics of expression of CD154 on rhesus lymphocytes, stimulated with plate-bound anti-CD3 antibod-ies, showed that maximum expression on CD4⫹T cells was achieved after 4 to 6 h of stimulation. The levels of expression were maintained up to 12 h poststimulation, after which they diminished (data not shown). This regulated expression is sim-ilar to the one observed for the human CD154 gene (17). Comparison of the nucleotide sequences coding for the extra-cellular region of human (GenBank accession no. L07414) and rhesus CD154, amplified with PCR primers sFCD154 and RCD154, showed only two amino acid differences among 218 residues (Fig. 1). Although we were able to amplify the full-length human CD154 gene with primers FCD154 and RCD154, we did not succeed with the rhesus CD154 gene, which indicates that there may be more differences in nucleo-tide sequences for the 5⬘ end region between the two genes. Considering the 99% homology between the extracellular por-tions of human and rhesus CD154 proteins, we decided to use the human CD154 gene for all our constructs. The CD154 gene was then inserted in place of nef into the genome of SIVmac239. Viruses carrying the CD154 gene in the sense (SIVCD154) and antisense (SIVCNTL) orientations were gen-erated in CEM-x-174 cells. Only cells infected with SIVCD154 expressed CD154 on their surface, as detected by flow cytom-etry (data not shown).
We analyzed the effects of SIV infection on the level of expression of CD154 on the cell surface of a continuous T-cell line derived in our laboratory from rhesus macaque PBMCs by
infection with herpesvirus saimiri. Infection of T-302 cells with viruses that lacked nef, such as SIVCD154 and SIVCNTL, resulted in a slight CD4 downregulation, whereas infection with SIVmac239 induced marked downregulation of both CD3 and CD4 proteins (data not shown). These data combined indicated that the recombinant viruses SIVCD154 and SIVCNTL indeed do not have a functionalnefgene.
We also studied the phenotypic changes induced by infection of rhesus PBMCs with SIV. Because we were interested in studying the capacity of the different viruses to induce cell activation, we infected unstimulated PBMCs with SIVmac239, SIVCD154, or SIVCNTL and cultured them in the presence of IL-2. Analysis of cells on day 9 showed that infection with SIV reduced the level of expression of the IL-2 receptor␣ chain (CD25) on T cells; however, PBMCs infected with SIVCD154 had significantly higher levels of CD25 than cells infected with SIVCNTL or SIVmac239 (Fig. 2A). Analysis of expression of CD154 also showed that only T cells infected with SIVCD154 had detectable levels of this surface protein (Fig. 2B). Identi-fication of which T cells expressed CD154 revealed that the CD8⫺ population, that is the CD4⫹ T cells, were the ones presenting the protein on their surface (Fig. 2C): the few CD8⫹T cells that expressed CD154 were CD4⫹CD8⫹, which are regularly present in macaques (data not shown). This ex-periment indicated that SIVCD154 was able to infect rhesus CD4⫹T cells and directed the expression of CD154 on the surface of the infected cells.
Because B cells are one of the cell types that interact with CD154-expressing CD4⫹T cells, we also analyzed activation markers and costimulatory molecules on these cells in the infected PBMC cultures. The percentage of B cells expressing CD40, the ligand of CD154, was significantly augmented only in cultures infected with SIVCD154 (Fig. 3A). Similarly, only cultures infected with SIVCD154 had elevated percentages of B cells expressing the early activation marker CD69 (Fig. 3B). Finally, only B cells from cultures infected with SIVCD154 showed upregulation of CD86, a costimulatory molecule whose expression is increased after CD40-CD154 interaction. These data clearly showed that the CD154 molecule expressed on CD4⫹T cells infected with SIVCD154 interacted with CD40 on B cells and that this CD40-CD154 contact induced
activa-FIG. 1. Comparison of the amino acid sequence of the extracellular domain of the human (GenBank accession no. L07414) and rhesus CD154 gene products. The first methionine (M in boldface) residue in both sequences was introduced by PCR. Differences in sequence are underlined and in boldface italic.
on November 8, 2019 by guest
[image:3.585.112.478.70.220.2]tion and up-regulation of other costimulatory molecules on B cells. Thus, SIVCD154 induced expression of CD154 on in-fected cells, and this newly synthesized molecule was biologi-cally active.
Infection of rhesus monkeys with recombinant SIV vectors.
After demonstrating the in vitro capacity of SIVCD154 to induce a biologically active CD154 on the surface of infected CD4⫹ T cells, we designed an experiment to study the real relevance of this CD154 expression within the context of a retroviral infection. Therefore, we inoculated two groups of four rhesus macaques each with a high dose of SIVCD154 or SIVCNTL. Measurement of viral loads in plasma or in LNs indicated low levels of replication during the acute phase of infection but did not show any significant difference between the two groups of animals. In general, virus in plasma was detectable by 2 weeks postinfection, and it peaked then or at 4 weeks postinfection. By 12 weeks postinfection, virus in plasma was below the limit of detection (2,000 SIV copies) in almost all of the animals (Fig. 4A). Cell-associated viral loads in LNs, on the other hand, peaked by 4 weeks postinfection and were still detectable, although at low levels, by 12 weeks postinfec-tion (Fig. 4B). Virus isolapostinfec-tion from rhesus lymphocytes by long-term culture demonstrated that some animals were able to control infection efficiently after primary infection (rhesus macaques 37, 44, 38, 40, and 41), whereas other animals had persistent, low-level viremia (rhesus macaques 35, 36, and 42);
however, both patterns were seen in either group (data not shown). Thus, infection of rhesus macaques with an SIV that expressed CD154 did not result in higher viral loads when compared with animals infected with an attenuated SIV that did not express this molecule.
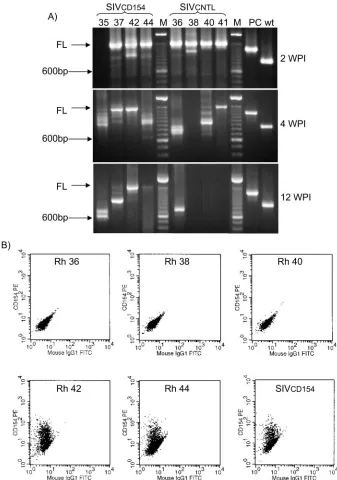
The stability of the CD154 gene in virus isolated from the infected macaques was analyzed by PCR and flow cytometry. As seen in other experiments with recombinant retroviruses, the genetic stability varied from animal to animal, and it was similar for SIVCD154 and SIVCNTL. For example, the CD154 gene was lost as early as 2 to 4 weeks postinfection in rhesus macaques 35 and 36, whereas the full-length CD154 gene was still detectable at 12 weeks postinfection in rhesus macaques 42 and 44 (Fig. 5A). Confirming the presence of the full-length CD154 gene, virus isolated from rhesus macaques 42 and 44 at 12 weeks postinfection was still able to direct expression of CD154 on the surface of the infected cells (Fig. 5B). By 16 weeks postinfection, we were not able to detect full-length CD154 inserts in any of the animals. These measurements demonstrated that the stability of the CD154 gene in the re-combinant vectors was not dependent on the orientation of the insert with respect to the SIV genome; they also showed that SIVCD154 directed the expression of CD154 on infected cells in vivo.
Phenotypic changes in lymphocytes from rhesus monkeys infected with recombinant SIV vectors.We analyzed the
phe-FIG. 2. Analysis of rhesus PBMCs after infection with SIVs. Unstimulated rhesus PBMCs were infected with SIVCD154, SIVmac239 (nef
open), or SIVCNTL at a multiplicity of infection of 0.1; uninfected cells were included as controls. After 9 days of infection, cells were harvested; stained with fluorescent antibodies against CD3, CD8, CD25, and CD154; and analyzed in a FACScan. (A) CD3 versus CD25, gated on lymphocytes; (B) CD3 versus CD154, gated on lymphocytes; (C) CD8 versus CD154, gated on CD3⫹lymphocytes. The experiment was repeated with three different donors with similar results, and a representative example is provided.
4682 HODARA ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:4.585.115.473.68.367.2]notypic changes induced by infection with SIVCD154 and SIVCNTL in LN and blood lymphocytes during the acute phase of the viral infection. Particularly in LNs, we studied expression of CD154 on CD4⫹T cells, and activation markers and costimulatory molecules that are present on B cells and
that linked to CD40-CD154 interactions. Variations in the percentages of CD4⫹T, CD8⫹T, and B cells were not statis-tically different between both groups. LN lymphocytes from all animals showed a gradual, slight reduction in the percentage of CD4⫹T cells and an increase in the number of CD8⫹T cells;
[image:5.585.138.447.68.342.2]FIG. 3. Analysis of costimulatory molecules and activation markers on rhesus B cells after infection with SIVs. Unstimulated rhesus PBMCs were infected with SIVCD154, SIVmac239 (nefopen), or SIVCNTL at a multiplicity of infection of 0.1; uninfected cells were included as controls. After 9 days of infection, cells were harvested; stained with fluorescent antibodies against CD20, CD40, CD69, and CD86; and analyzed in a FACScan. (A) CD20 versus CD40, gated on lymphocytes; (B) CD20 versus CD69, gated on lymphocytes; (C) CD20 versus CD86, gated on lymphocytes. The experiment was repeated with three different donors, and the same representative example is provided.
FIG. 4. Viral loads in plasma and LN cells in rhesus macaques after infection with SIV vectors. Rhesus macaques were infected with 104
TCID50s of SIVCD154 (solid symbols, straight lines) or SIVCNTL (open symbols, dotted lines). (A) SIV RNA loads in heparinized plasma were
determined by NASBA; the limit of detection was 2,000 SIV copies/100l of plasma. (B) Cell-associated viral loads were determined in LN cells by limited dilution and cocultivation with CEM-x-174 cells.
on November 8, 2019 by guest
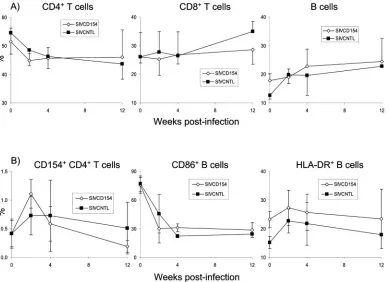
a similar increase was observed for LN B cells (Fig. 6A). Levels of expression of CD154 on LN CD4⫹T cells were similar for both groups before infection (0.42%⫾0.23% for animals in the SIVCD154 group versus 0.41%⫾0.26% for animals in the SIVCNTL group), but increased to higher levels in animals infected with SIVCD154 by 2 weeks postinfection (1.11% ⫾ 0.25% for animals in the SIVCD154 group versus 0.73% ⫾
[image:6.585.122.459.71.551.2]0.34% for animals in the SIVCNTL group); however, this difference was not statistically significant (P⫽0.16; Fig. 6B). Analysis of costimulatory molecules and activation markers in LN B cells did not reveal differences that were statistically significant between the two groups either. However, it was interesting to observe in all animals a transient increase in the percentage of HLA-DR⫹B cells, whereas the percentage of
FIG. 5. Characterization of virus isolated from macaques infected with SIVCD154 or SIVCNTL. (A) Stability of the CD154 insert. The 3⬘long terminal repeat region of the viruses isolated from infected rhesus macaques was amplified by PCR as described in Materials and Methods, using primers that encompassed the region where the CD154 gene had been inserted. PCR-amplified DNA fragments were separated by agarose electrophoresis. FL, full-length CD154 insert, 1,500 bp; M, 100-bp-ladder molecular weight marker; PC, positive control for extended insert, 1,340 bp; wt, wild-type SIVmac239, 880 bp. WPI, weeks postinfection. (B) Expression of CD154 by virus isolated from infected macaques. CEM-x-174 cells used for isolation of SIV from SIVCD154- or SIVCNTL-infected macaques were stained with an anti-CD154 antibody and analyzed by flow cytometry. SIVCD154 represents CEM-x-174 cells directly infected with SIVCD154.
4684 HODARA ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
CD86⫹B cells dropped significantly to low and stable levels after infection (Fig. 6B).
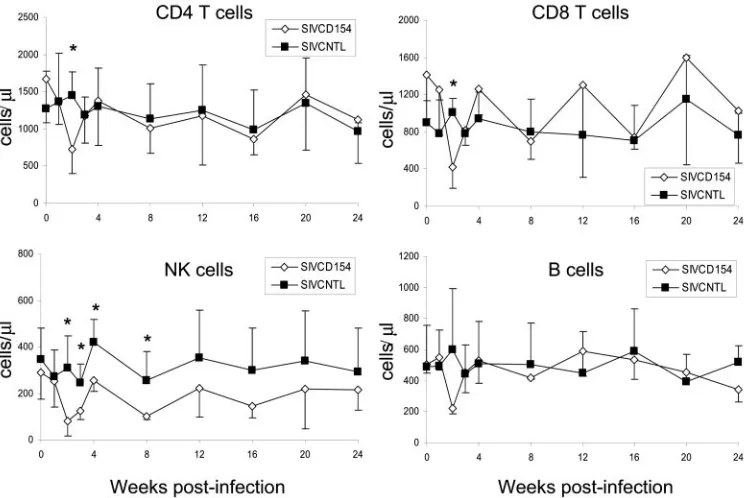
Analysis of peripheral blood lymphocytes (PBLs) showed some transient differences between animals infected with SIVCD154 or SIVCNTL that were statistically significant. For example, animals infected with SIVCD154 had a drastic reduc-tion in the number of CD4⫹and CD8⫹T cells, B cells, and NK cells at 2 weeks postinfection. The reduction in absolute num-ber of NK cells was even more pronounced, lasting until 8 weeks postinfection (Fig. 7). This reduction in absolute num-bers at 2 weeks postinfection in animals infected with SIVCD154 was also accompanied by a reduced percentage of activated (cells expressing CD69) CD4⫹T, B, and NK cells (Fig. 8). Interestingly, only CD8⫹T cells from animals infected with SIVCD154 had a peak of activation by 8 weeks postin-fection. In general, after the acute phase of infection, absolute numbers of blood lymphocytes in all animals remained stable, but the general state of activation of CD4⫹and CD8⫹T cells remained higher than the preinfection levels.
Immune response of rhesus macaques to infection with re-combinant SIV vectors.We studied changes in plasma cytokine concentration induced by infection with SIVCD154 and SIVCNTL, as well as the cellular and humoral immune re-sponse elicited by these viruses. In contrast to what is observed after infection with pathogenic SIV (19), analysis of levels of IFN-␣, IL-12, and IL-18 in plasma before and after infection with these recombinant SIV vectors did not show consistent
and significant changes for any of these cytokines in either group (data not shown).
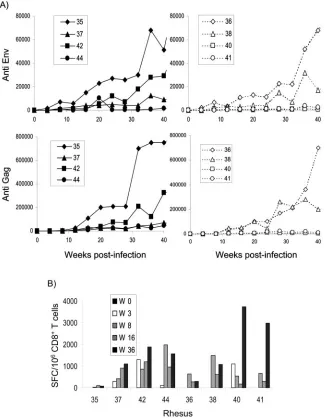
The antibody response against SIV Gag and Env was deter-mined by specific ELISA. Similar to what was observed throughout this study, there were no statistically significant differences between the levels of antibodies produced by ani-mals infected with SIVCD154 and those produced by aniani-mals infected with SIVCNTL. The patterns were similar for both groups of animals and consisted of very low antibody titers for those animals that controlled viremia efficiently, whereas vire-mic animals such as rhesus macaques 35 and 36 had high anti-SIV gp160 and Gag antibody titers (Fig. 9A).
The cell-mediated immune response was studied by IFN-␥ ELISPOT using 15-amino-acid peptides that encompassed the SIV Gag protein; these SFC were identified as being CD8⫹T cells by IFN-␥ intracellular staining (data not shown). Al-though by 3 weeks postinfection we were able to identify SIV Gag-specific SFC in three out four animals in the SIVCD154-infected group, as compared to one out of four for the SIVCNTL-infected animals, these differences disappeared by 8 weeks postin-fection (Fig. 9B). Interestingly, the pattern that was observed for the cell-mediated immune response was the opposite one to that seen for the humoral response; that is, animals that controlled the infection efficiently had high levels of SIV Gag-specific SFCs, whereas viremic animals had very low numbers.
[image:7.585.102.491.71.353.2]In summary, both SIVCD154 and SIVCNTL behaved like attenuated viruses, and there was no evidence that the level of
FIG. 6. Changes in LN cell phenotype after infection with SIV vectors. Rhesus macaques were infected with SIVCD154 (open diamonds) or SIVCNTL (solid squares); LN biopsies were performed before and after infection, and LN cells were analyzed by flow cytometry. (A) Analysis of lymphocyte subsets. LN cells were stained with antibodies to CD3, CD4, CD8, and CD20. (B) Analysis of costimulatory molecules and activation markers on LN CD4⫹T cells and B cells. LN cells were stained with antibodies to CD3, CD4, and CD154 or CD20, CD86, and HLA-DR. Values represent the mean of four animals with the standard deviation.
on November 8, 2019 by guest
expression of CD154 induced by SIVCD154 altered the devel-opment of the immune response to the virus.
DISCUSSION
It has been observed that infection with HIV leads to dys-regulation of CD154 signaling in CD4⫹T cells and impairment of the generation of T-cell-dependent immune responses in infected patients (28, 29, 42). It was first suggested by Chirmule et al. (8) that interaction between HIV gp120 and the CD4 receptor might play a role in the mechanisms underlying CD154 dysregulation. Recently, Zhang et al. (50) demon-strated that, indeed, interactions between native, virion-asso-ciated HIV gp120 and the CD4 receptor were responsible for reducing CD154 mRNA and protein levels on T cells. Conse-quently, it has been suggested that this reduced CD154 expres-sion on CD4⫹T cells, in turn, impairs the ability of the APCs to express cytokines and deliver other costimulatory signals, leading to the deficiencies in both APC and CD4⫹T-cell func-tion observed during HIV and SIV infecfunc-tion. As a result of these findings, we hypothesized that this vicious cycle could be broken with a recombinant SIV expressing its own CD154, because SIV-infected CD4⫹T cells would have the ability to express CD154 even if the association of gp120 with CD4 blunted the normal expression of CD154.
We engineered a recombinant SIV that lacked thenefgene and had the capacity to replicate and express CD154 on the surface of the infected cells. It could be argued that all the studies performed in HIV-infected individuals that suggest a CD154 dysfunction upon infection have been performed with individuals infected withnef-expressing viruses. In fact,
[image:8.585.104.476.70.319.2]infec-tion of macaques withnef-deficient SIV resulted in mild im-mune alterations and lack of disease progression during the first several years of infection (26), with similar outcomes found in humans infected withnef-deficient HIV (13). How-ever, long-term studies in infected humans and rhesus ma-caques have demonstrated thatnef-deficient HIV and SIV are still pathogenic and able to induce immune dysfunction and AIDS, although in a protracted manner (reviewed in reference 47). Thus, considering thatnef is not the only viral gene in-volved in immune dysfunction of the host and that experimen-tal evidence suggests that gp120-CD4 interactions impair CD154 expression, our experimental approach was valid. We then demonstrated in vitro that this virus, SIVCD154, could infect rhesus PBMCs, induce the expression of CD154 (Fig. 2), and stimulate the B cells present in the same culture (Fig. 3). For these experiments, we had to infect rhesus PBMCs that were not stimulated before infection. Mitogenic stimulation of PBMCs prior to infection is a routine technique employed for the amplification of retroviral cultures; however, the resulting global upregulation of activation and stimulation markers on lymphocytes would mask any effect due to the expression of CD154 by SIVCD154. Thus, our in vitro PBMC studies re-sulted in a low number of CD4⫹T cells being infected, but the unique outcome of B-cell activation and upregulation of CD154 seen after infection with SIVCD154 was clearly distin-guishable from what was observed for SIVmac239 and SIVCNTL. For final proof of our hypothesis, we performed in vivo experiments, infecting rhesus macaques with SIVCD154. As a control, we infected a second group of animals with SIVCNTL, a virus that underwent similar genetic manipula-tions as SIVCD154 but did not express CD154. Surprisingly,
FIG. 7. Changes in absolute values of peripheral blood lymphocytes after infection with SIV vectors. Rhesus macaques were infected with SIVCD154 (open diamonds) or SIVCNTL (solid squares). Blood lymphocytes were stained with antibodies to CD3, CD4, CD8, CD16, and CD20 and analyzed as described in Materials and Methods. Values represent the mean of four animals with the standard deviation. Asterisks indicate a significant difference between both groups (P⬍0.05; unpairedttest).
4686 HODARA ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
CXCR4-dependent viruses (3). Similarly, increased levels of HIV infection have been reported after CD154-mediated ac-tivation of Langerhans cell-like DCs (25) and plasmacytoid DCs (15). In the context of our experiments, both SIVCD154 and SIVCNTL behaved like attenuated viruses that induced low viral loads and, depending on the animal, transient viremic episodes (Fig. 4). Although virus isolated from SIVCD154-infected macaques was shown to direct expression of CD154 during the acute phase of the infection (Fig. 5), viral titers were similar to those seen for the macaques infected with the con-trol virus. Thus, expression of active CD154 during acute SIV infection with anef-deficient virus did not result in increased viral replication. The higher viral loads observed in animals 35 and 36, which were infected with SIVCD154 and SIVCNTL, respectively, are a consequence of the loss of the CD154 gene at a very early time point. This loss of “superfluous” genetic material results in viral species more fitted for replication and is a phenomenon that has been observed before, both in vivo
[image:9.585.99.480.449.684.2]markable and long-lasting decrease in CD86⫹LN B cells may be the consequence of an unidentified antiviral immune re-sponse, since it has been shown that activated B cells can induce stimulation of CD4⫹T cells via CD86-CD28 interac-tion and lead to increased HIV replicainterac-tion (31). The only differences that were statistically significant occurred in periph-eral blood at 2 weeks postinfection, when absolute numbers of lymphocytes and activation levels were reduced in animals infected with SIVCD154 (Fig. 7 and 8). This phenomenon cannot be explained by levels of viral replication, because viral loads were low and similar for both groups of macaques. One possible explanation is that expression of CD154 on SIVCD154-infected T cells resulted in the activation of CD40-expressing endothelial cells, and activated endothelial cells have been shown to regulate leukocyte trafficking into inflam-matory sites by secreting the chemokines IL-8, MCP-1, and RANTES (44, 46). Thus, this reduction in absolute numbers and activation levels for blood lymphocytes may be a reflection
FIG. 8. Changes in activation state (expression of CD69) of peripheral blood lymphocytes after infection with SIV vectors. Rhesus macaques were infected with SIVCD154 (open diamonds) or SIVCNTL (solid squares). Blood lymphocytes were stained with antibodies to CD3, CD4, CD8, CD16, CD20, and CD69 and analyzed as described in Materials and Methods. Values represent the mean of four animals with the standard deviation. Asterisks indicate a significant difference between both groups (P⬍0.05, unpairedttest).
on November 8, 2019 by guest
of increased extravascular trafficking of activated lymphocytes, which may be rapidly controlled by homeostatic mechanisms once viral loads decline.
As mentioned before, CD154-CD40 interactions are critical for the development of T-cell-dependent immune responses. Nevertheless, analysis of the anti-SIV immune responses in-duced in macaques infected with SIVCD154 or SIVCNTL did not show any significant difference, except for an early anti-SIV Gag cell-mediated response in SIVCD154-infected monkeys (Fig. 9B). We believe that these similarities in immunoge-nicity are a consequence of SIVCD154 and SIVCNTL both being negative for Nef expression. SIV Nef has been shown to downmodulate cell surface expression of CD4 (16), CD3 chain (4, 24), major histocompatibility complex (MHC) class I (37), and CD28 (5, 43) molecules, and to induce
expression of FAS ligand (49) on infected cells. All these events have the potential to affect the ability of the infected CD4⫹T cell to interact with an APC via the T-cell receptor (TCR) or the delivery of costimulatory signals. Thus, it is possible that, in the absence of Nef, the disturbances to the CD154-CD40 axis associated with the interaction between gp120 and CD4 (8, 50) are not of a magnitude such as to prevent the development of a strong immune response. An-other factor that favors the host immune response over the virus replication, and that has been observed before (1, 18, 20), is that both SIVCD154 and SIVCNTL are recombinant SIV⌬nef viruses that have an additional DNA sequence (the CD154 gene), which reduces the kinetics of viral replication and allows the generation of an early immune response. Nevertheless, this immune response is not completely
effec-FIG. 9. Analysis of the anti-SIV humoral and cellular immune response of rhesus macaques after infection with SIV vectors. Rhesus macaques were infected with SIVCD154 (rhesus macaques 35, 37, 42, and 44) or SIVCNTL (rhesus macaques 36, 38, 40, and 41). (A) Anti-SIV Env and Gag antibody titers were determined in rhesus plasma by antigen-specific ELISA as described in Materials and Methods. (B) SIV Gag-specific IFN-␥ SFC were determined by ELISPOT using rhesus PBMCs stimulated with overlapping 15-mer SIV Gag peptides.
4688 HODARA ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:10.585.127.452.72.490.2]genic infection. However, our studies do not preclude the development of immunotherapeutic approaches that target CD154, but they reinforce the concept that the CD154-related immune defects seen in HIV infection may be caused by events upstream of the signals that lead to the expression of CD154.
ACKNOWLEDGMENTS
This study was funded by NIH grants R01 AI41923, R21 AI44363, and R51 RR013986. NASBA determinations were possible thanks to the sponsorship of the Reagent Resource Support Program for AIDS Vaccine Development, National Institute of Allergy and Infectious Diseases, National Institutes of Health.
The authors thank Kathy Brasky, Robert Geiger, and personnel from the Department of Comparative Medicine of the Southwest Na-tional Primate Research Center for assistance with the animal studies. We also thank A. Hopstetter for editing the manuscript.
REFERENCES
1.Alexander, L., R. S. Veazey, S. Czajak, M. DeMaria, M. Rosenzweig, A. A. Lackner, R. C. Desrosiers, and V. G. Sasseville.1999. Recombinant simian immunodeficiency virus expressing green fluorescent protein identifies
in-fected cells in rhesus monkeys. AIDS Res. Hum. Retrovir.15:11–21.
2.Baba, T. W., V. Liska, A. H. Khimani, N. B. Ray, P. J. Dailey, D. Penninck, R. Bronson, M. F. Greene, H. M. McClure, L. N. Martin, and R. M. Ru-precht.1999. Live attenuated, multiply deleted simian immunodeficiency
virus causes AIDS in infant and adult macaques. Nat. Med.5:194–203.
(Erratum,5:590.)
3.Bakri, Y., A. Mannioui, L. Ylisastigui, F. Sanchez, J. C. Gluckman, and A. Benjouad.2002. CD40-activated macrophages become highly susceptible to X4 strains of human immunodeficiency virus type 1. AIDS Res. Hum.
Ret-rovir.18:103–113.
4.Bell, I., C. Ashman, J. Maughan, E. Hooker, F. Cook, and T. A. Reinhart.
1998. Association of simian immunodeficiency virus Nef with the T-cell receptor (TCR) zeta chain leads to TCR down-modulation. J. Gen. Virol.
79:2717–2727.
5.Bell, I., T. M. Schaefer, R. P. Trible, A. Amedee, and T. A. Reinhart.2001. Down-modulation of the costimulatory molecule, CD28, is a conserved ac-tivity of multiple SIV Nefs and is dependent on histidine 196 of Nef. Virology
283:148–158.
6.Bergamini, A., F. Bolacchi, C. D. Pesce, M. Carbone, M. Cepparulo, F. Demin, and G. Rocchi.2002. Increased CD4 and CCR5 expression and human immunodeficiency virus type 1 entry in CD40 ligand-stimulated
mac-rophages. J. Infect. Dis.185:1567–1577.
7.Chehimi, J., S. E. Starr, I. Frank, A. D’Andrea, X. Ma, R. R. MacGregor, J. Sennelier, and G. Trinchieri.1994. Impaired interleukin 12 production in
human immunodeficiency virus-infected patients. J. Exp. Med.179:1361–
1366.
8.Chirmule, N., T. W. McCloskey, R. Hu, V. S. Kalyanaraman, and S. Pahwa.
1995. HIV gp120 inhibits T cell activation by interfering with expression of
costimulatory molecules CD40 ligand and CD80 (B71). J. Immunol.155:
917–924.
9.Chougnet, C.2003. Role of CD40 ligand dysregulation in HIV-associated
dysfunction of antigen-presenting cells. J. Leukoc. Biol.74:702–709.
10.Chougnet, C., E. Thomas, A. L. Landay, H. A. Kessler, S. Buchbinder, S. Scheer, and G. M. Shearer.1998. CD40 ligand and IFN-gamma synergisti-cally restore IL-12 production in HIV-infected patients. Eur. J. Immunol.
28:646–656.
11.Cole, K. S., J. L. Rowles, B. A. Jagerski, M. Murphey-Corb, T. Unangst, J. E.
11041.
16.Foster, J. L., S. J. Anderson, A. L. Frazier, and J. V. Garcia.1994. Specific suppression of human CD4 surface expression by Nef from the pathogenic
simian immunodeficiency virus SIVmac239open. Virology201:373–379.
17.Gauchat, J. F., J. P. Aubry, G. Mazzei, P. Life, T. Jomotte, G. Elson, and J. Y. Bonnefoy.1993. Human CD40-ligand: molecular cloning, cellular distribu-tion and reguladistribu-tion of expression by factors controlling IgE producdistribu-tion.
FEBS Lett.315:259–266.
18.Giavedoni, L., S. Ahmad, L. Jones, and T. Yilma.1997. Expression of gamma interferon by simian immunodeficiency virus increases attenuation and re-duces postchallenge virus load in vaccinated rhesus macaques. J. Virol.
71:866–872.
19.Giavedoni, L. D., M. C. Velasquillo, L. M. Parodi, G. B. Hubbard, and V. L. Hodara.2000. Cytokine expression, natural killer cell activation, and phe-notypic changes in lymphoid cells from rhesus macaques during acute
infec-tion with pathogenic simian immunodeficiency virus. J. Virol.74:1648–1657.
20.Giavedoni, L. D., M. C. Velasquillo, L. M. Parodi, G. B. Hubbard, and V. L. Hodara.2002. Expression of IL-18 by SIV does not modify the outcome of
the antiviral immune response. Virology303:327–337.
21.Giavedoni, L. D., and T. Yilma.1996. Construction and characterization of replication-competent simian immunodeficiency virus vectors that express
gamma interferon. J. Virol.70:2247–2251.
22.Grewal, I. S., and R. A. Flavell.1997. The CD40 ligand. At the center of the
immune universe? Immunol. Res.16:59–70.
23.Grewal, I. S., J. Xu, and R. A. Flavell.1995. Impairment of antigen-specific
T-cell priming in mice lacking CD40 ligand. Nature378:617–620.
24.Howe, A. Y. M., J. U. Jung, and R. C. Desrosiers.1998. Zeta chain of the T-cell receptor interacts with nef of simian immunodeficiency virus and
human immunodeficiency virus type 2. J. Virol.72:9827–9834.
25.Kawamura, T., M. Qualbani, E. K. Thomas, J. M. Orenstein, and A. Blau-velt.2001. Low levels of productive HIV infection in Langerhans cell-like dendritic cells differentiated in the presence of TGF-beta1 and increased viral replication with CD40 ligand-induced maturation. Eur. J. Immunol.
31:360–368.
26.Kestler, H. W., III, D. J. Ringler, K. Mori, D. L. Panicali, P. K. Sehgal, M. D. Daniel, and R. C. Desrosiers.1991. Importance of the nef gene for
mainte-nance of high virus loads and for development of AIDS. Cell65:651–662.
27.Kirchhoff, F., H. W. Kestler III, and R. C. Desrosiers.1994. Upstream U3 sequences in simian immunodeficiency virus are selectively deleted in vivo in
the absence of an intactnefgene. J. Virol.68:2031–2037.
28.Kornbluth, R. S.2000. The emerging role of CD40 ligand in HIV infection.
J. Leukoc. Biol.68:373–382.
29.Kornbluth, R. S. 2002. An expanding role for CD40L and other tumor necrosis factor superfamily ligands in HIV infection. J. Hematother. Stem.
Cell Res.11:787–801.
30.Kornbluth, R. S., K. Kee, and D. D. Richman.1998. CD40 ligand (CD154) stimulation of macrophages to produce HIV-1-suppressive beta-chemokines.
Proc. Natl. Acad. Sci. USA95:5205–5210.
31.Krzysiek, R., E. A. Lefevre, C. Legendre, D. Treton, D. Dormont, P. Ga-lanaud, G. Gras, and Y. Richard.1998. B cell-driven HIV type 1 expression in T cells: an essential role of CD86 costimulatory molecule. AIDS Res.
Hum. Retrovir.14:989–997.
32.Lee, M. E., S. Z. Bucur, T. W. Gillespie, J. W. Adams, A. T. Barker, E. K. Thomas, J. D. Roback, and C. D. Hillyer.1999. Recombinant human CD40 ligand inhibits simian immunodeficiency virus replication: a role for
inter-leukin-16. J. Med. Primatol.28:190–194.
33.Lohman, B. L., J. Higgins, M. L. Marthas, P. A. Marx, and N. C. Pedersen.
1991. Development of simian immunodeficiency virus isolation, titration, and neutralization assays which use whole blood from rhesus monkeys and an antigen capture enzyme-linked immunosorbent assay. J. Clin. Microbiol.
29:2187–2192.
34.Marzio, P. D., B. Sherry, E. K. Thomas, G. Franchin, H. Schmidtmayerova, and M. Bukrinsky.2003. Beta-chemokine production in CD40L-stimulated
on November 8, 2019 by guest
monocyte-derived macrophages requires activation of MAPK signaling
path-ways. Cytokine23:53–63.
35.McDyer, J. F., M. Dybul, T. J. Goletz, A. L. Kinter, E. K. Thomas, J. A. Berzofsky, A. S. Fauci, and R. A. Seder.1999. Differential effects of CD40 ligand/trimer stimulation on the ability of dendritic cells to replicate and transmit HIV infection: evidence for CC-chemokine-dependent and
-inde-pendent mechanisms. J. Immunol.162:3711–3717.
36.Morris, L., J. M. Binley, B. A. Clas, S. Bonhoeffer, T. P. Astill, R. Kost, A. Hurley, Y. Cao, M. Markowitz, D. D. Ho, and J. P. Moore.1998. HIV-1 antigen-specific and -nonspecific B cell responses are sensitive to
combina-tion antiretroviral therapy. J. Exp. Med.188:233–245.
37.Piguet, V., O. Schwartz, S. Le Gall, and D. Trono.1999. The downregulation of CD4 and MHC-I by primate lentiviruses: a paradigm for the modulation
of cell surface receptors. Immunol. Rev.168:51–63.
38.Reed, L. J., and H. Muench.1938. A simple method of estimating fifty
percent endpoints. Am. J. Hyg.27:493–497.
39.Robinson, J. E., D. Holton, J. Liu, H. McMurdo, A. Murciano, and R. Gohd.
1990. A novel enzyme-linked immunosorbent assay (ELISA) for the detec-tion of antibodies to HIV-1 envelope glycoproteins based on immobilizadetec-tion of viral glycoproteins in microtiter wells coated with concanavalin A. J.
Im-munol. Methods132:63–71.
40.Stahl-Hennig, C., B. R. Gundlach, U. Dittmer, P. ten Haaft, J. Heeney, W. Zou, D. Emilie, S. Sopper, and K. Uberla.2003. Replication, immunogenic-ity, and protective properties of live-attenuated simian immunodeficiency
viruses expressing interleukin-4 or interferon-gamma. Virology305:473–485.
41.Subauste, C. S., M. Wessendarp, J. A. Portilllo, R. M. Andrade, L. M. Hinds, F. J. Gomez, A. G. Smulian, P. A. Grubbs, and L. A. Haglund.2004.
Patho-gen-specific induction of CD154 is impaired in CD4⫹T cells from human
immunodeficiency virus-infected patients. J. Infect. Dis.189:61–70.
42.Subauste, C. S., M. Wessendarp, A. G. Smulian, and P. T. Frame.2001. Role of CD40 ligand signaling in defective type 1 cytokine response in human
immunodeficiency virus infection. J. Infect. Dis.183:1722–1731.
43.Swigut, T., N. Shohdy, and J. Skowronski.2001. Mechanism for
down-regulation of CD28 by Nef. EMBO J.20:1593–1604.
44.Thienel, U., J. Loike, and M. J. Yellin.1999. CD154 (CD40L) induces human endothelial cell chemokine production and migration of leukocyte subsets.
Cell Immunol.198:87–95.
45.van Kooten, C., and J. Banchereau.2000. CD40-CD40 ligand. J. Leukoc.
Biol.67:2–17.
46.van Kooten, C., J. S. Gerritsma, M. E. Paape, L. A. van Es, J. Banchereau, and M. R. Daha.1997. Possible role for CD40-CD40L in the regulation of
interstitial infiltration in the kidney. Kidney Int.51:711–721.
47.Whitney, J. B., and R. M. Ruprecht.2004. Live attenuated HIV vaccines:
pitfalls and prospects. Curr. Opin. Infect. Dis.17:17–26.
48.Wolthers, K. C., S. A. Otto, S. M. Lens, R. A. Van Lier, F. Miedema, and L. Meyaard.1997. Functional B cell abnormalities in HIV type 1 infection: role
of CD40L and CD70. AIDS Res. Hum. Retrovir.13:1023–1029.
49.Xu, X. N., B. Laffert, G. R. Screaton, M. Kraft, D. Wolf, W. Kolanus, J. Mongkolsapay, A. J. McMichael, and A. S. Baur.1999. Induction of Fas ligand expression by HIV involves the interaction of Nef with the T cell
receptor zeta chain. J. Exp. Med.189:1489–1496.
50.Zhang, R., C. J. Fichtenbaum, D. A. Hildeman, J. D. Lifson, and C.
Choug-net.2004. CD40 ligand dysregulation in HIV infection: HIV glycoprotein 120
inhibits signaling cascades upstream of CD40 ligand transcription. J.
Immu-nol.172:2678–2686.
4690 HODARA ET AL. J. VIROL.