0022-538X/79/02-0705/11$02.00/0
Vaccinia
Virus
Replication
I.
Requirement for the
Host-Cell Nucleus
DENNIS E. HRUBY,l*LINDA A. GUARINO,2 ANDJOSEPH R. KATES'
DepartmentsofMicrobiology'andPharmacology,2HealthSciences Center, State University of New York,
Stony Brook,New York 11794
Received for publication 29 August 1978
Using cytochalasin B-induced enucleation techniques,weexamined the ability ofvaccinia virus toreplicate in the absence ofthe host-cellnucleus in several
mammaliancell lines.Itwasfound that virus-infected enucleatedcells(cytoplasts) prepared fromBSC-40, CVC, and Lcellswereincapable ofproducing infectious
progeny virus. The natureof this apparent nuclearinvolvementwas studied in
detail in BSC-40 cells. Modulations designedtomaximize cytoplastintegrity and longevity, suchasreductionof thegrowth temperatureand initialmultiplicityof
infection, did notimprove virus growth in cytoplasts. Sodium dodecyl
sulfate-polyacrylamide gel analysis of the[35S]methionine pulse-labeledproteins
synthe-sized in vaccinia virus-infected cytoplasts demonstratedthat bothearly and late
viral gene products were being expressed at high levels and with the proper
temporalsequence. Vaccinia virus cytoplasmic DNA synthesis, asmeasured by [3H]thymidine incorporation, peaked at 3hpostinfection andwas 70 to90% of control levels in cytoplasts. However, in the cytoplasts this DNA was not
convertedto aDNase-resistant form late in infection, whichwasconsistent with the failure to isolate physical particles from infected cytoplasts. Treatment of vaccinia virus-infectedcells with100
,jg
of rifampin/ml from0 to 8h toincrease thepools of viralprecursors,followed by subsequent removal of the drug, resulted inathreefold increase virusyield.This treatmenthadnoeffectonvirus-infected cytoplasts. Finally, vaccinia virus morphogenesis wasstudiedunderanelectron microscope in thin sections of virus-infectedcellsandcytoplasts which hadbeenpreparedatvarious timesduringasingle-step growthcycle. Itwasapparentthat, although early virus morphogenetic forms appeared, there was no subsequent DNAcondensationorparticle maturation in thecytoplasts. These resultssuggest
that vaccinia virus requiressomefactororfunction from thehost-cellnucleus in ordertomatureproperly and produce infectiousprogenyvirus.
Vacciniavirus,alargecomplex DNA-contain-ingpoxvirus, has beenthoughttoreplicatesolely in the cytoplasm ofsusceptible host cells (23). This ideawasbasedonseveral lines of evidence: vaccinia virus containsorcodes formany ofthe
enzymes necessary for DNA synthesis,
tran-scription, and modification (18); virus replicative factoriesarelocatedexclusivelyinthecytoplasm (11); and DNAsynthesis,asmeasuredby
auto-radiography, occurs in vaccinia virus-infected enucleated L cells (24). However,other
experi-ments have shown that pretreatment of host
cellswith eithermitomycin CoractinomycinD
toinhibit hostnuclear functionscauses amarked
diminutioninsubsequentvaccinia virusgrowth
(15, 26). Also, there have been indications that some virus-specific DNA synthesis may occur
within the nucleus ofpoxvirus-infected cells(9,
13,29). PenningtonandFollett(22) studied the
replication ofvaccinia virus inenucleated
BSC-1 cells and found that the production of infec-tious progeny virus was completely blocked. Thiswasaccompanied byaconsiderable reduc-tion in the levels ofvirus-specificDNAsynthesis, protein synthesis, and mature particle forma-tion, and also in the size and number ofvirus
factories. Similar observations have been made concerningtheinabilityofaniridovirus,African swine fevervirus,toreplicatein Verocytoplasts (20). It should bepointedoutthatreplicationof cytoplasmic RNA viruses suchasvesicular
sto-matitus virus and SemlikiForestvirus is
essen-tiallynormal inenucleated cells(8).
With therecentevidencedemonstratingthat
anothergroupofcytoplasmicDNAviruses,the
icosahedralvirusessuchasfrogvirus3,require
thenucleus forviral DNAreplicationand
tran-scription(10),itwasof interesttoreexamine the
so-calledcytoplasmicmode ofpoxvirus
replica-tionand therelationshipofthe apparent nuclear
705
on November 10, 2019 by guest
http://jvi.asm.org/
706
involvement. Intheexperiments reportedhere,
wehave carefullyexaminedtheability of
vacci-niavirustoreplicateinthe absence of the host-cellnucleus. Our resultsconfirmand extend the
previous observationthatvaccinia virus willnot
grow in enucleated cells. Furthermore,
experi-mental conditions have been defined under
which the levels of early and late viral gene
expression in enucleatedcellswerenearly
equiv-alent tothose in controlcells, yetno infectious
virus was produced. This apparently resulted
fromaberrantpackagingand maturation of virus
particles late in the infectious cycle, indicating
that some host-cell nuclear function was
re-quiredtoobtainpropervaccinia virus
morpho-genesis.
MATERIALS AND METHODS
Radioisotopes and chemicals. [3S]methionine
(586.4 Ci/mmol) and [methyl-3H]thymidine (55.6
Ci/mmol)werepurchasedfromNewEnglandNuclear
Corp. Cytosine arabinoside and rifampin were from
Sigma Chemical Co. Cytochalasin B was obtained
from Aldrich Chemical Co. All gel electrophoresis
reagentscamefrom Bio-Rad Laboratories.
Cells and virus. BSC-40 cells,obtained from M.
Ensinger,whoselectedforaderivative of BSC-1cells
which grew well at 400C, were maintained in Eagle
minimalessential medium(MEM) plus 10%
heat-in-activatedfetal calfserum (AFCS).Lcells(I. Tamm)
were grown in MEMplus 5% AFCS. CVC cells (P.
Tegtmeyer) weremaintained in basal Earle medium
(BME) plus10%AFCS.
Vacciniavirus,WRstrain, wasobtained from the
American Type CultureCollection and twiceplaque
purifiedonLcells.Purifiedvesicularstomatitisvirus,
Indianastrain,wasdonatedbyL.A. Ball.
Growth and purification of vaccinia virus.
Crude stocks of vaccinia virus were maintained by
low-multiplicitypassageonL cells.Crude virus stock
wastrysinizedwithanequalvolumeof 0.25%trypsin
(GIBCO) at37°Cfor15minwithfrequent shakingin
a Vortex mixer and then was diluted with ice-cold
adsorptionmedium(1part Puck'ssaline,1partMEM
plus5% AFCS). Confluentmonolayersof L cells (107
to3 x 107cells/100- by20-mmdish)wereinfected at
amultiplicityofinfection(MOI)of 0.01to0.1 at250C
for 3 h. The virus inoculumwasthenremoved,MEM
plus5%AFCS wasadded,and the disheswere
incu-batedat370Cfor 2to4days(oruntil themicroscopic
cytopathiceffect[CPE]wascomplete).Infectedcells
wereharvestedbycentrifugationand thenwere
resus-pended in 1 ml of MEM (minus serum)/dish and
storedat-70°C.The titer of crude stockspreparedin
thisway was109to3x 109PFU/ml.
Purified vaccinia virus stocks were prepared by
infecting log-phase spinner L cells (about 5 x 105
cells/ml) ataninitialMOI of1with crude virus stock
and thenincubatingthe infected culture at370Cfor 48 h. Although one mightnot expect allcells to be
initially infectedat thisMOI, itin fact consistently
gavethehighestviralyields.Viruswaspurifiedfrom
the infected cellsby two cycles ofsucrose gradient
centrifugation as previously described (1) except that
virusdispersion by sonic treatment wastotally
elimi-nated andreplaced with gentlehomogenization in a
Duallhomogenizer. Assuming1unit of optical density
at 260 nm equals 1.2 x 1010particles, purified viral
pelletswereresuspendedin1mMTris-hydrochloride
(pH 9) to a concentration of1mg/ml(2x 1011
parti-cles/ml) andstored insmall portionsat-70°C. Virus
yields from this procedure were 150 to 300 PFU/cell,
and the resultantpurifiedvirushad a PFU-to-particle
ratio of 1:8 to 1:20.
Plaque assay. Infectious vaccinia virus was
as-sayed induplicateonconfluent60-by 15-mmdishes
of BSC-40 cells. Ten-fold serial dilutions of purified
virus were made inphosphate-buffered saline
contain-ing1 mMMg2".Crude virus stocksweretrypsinized
with anequal volume of 0.25% trypsin at370Cfor 15
min and then diluted with adsorption medium. In
either case,washed-cellmonolayers were infected with
0.5ml of virus inoculum for3h at25°C with occasional
rocking to prevent drying and to obtain a uniform
distribution of virus. Virus inocula were removed after
3h andreplaced with MEMplus 5% AFCS and 1%
agar (Noble). The assays were incubated at 37°C
under 5%C02for2 to3daysand then were stained
with 0.01% neutral redfor2 to 4h to visualize and
enumerateplaques.
Enucleation ofcell monolayers. Cell monolayers
wereenucleated on 60-by 15-mm tissue culture dishes
byuseof thedrugcytochalasinB (3). Slight
modifi-cations ofpreviously published procedures (25)were
necessary to achieve high-yield mass enucleation of
the variouscelllines(specific proceduresfor eachcell
line aregivenbelow). Subconfluent monolayers ofcells
wereinvertedasepticallyinbottles of 37°C
cytochal-asin B-containing medium plus serum and
preincu-bated at 37°C. The bottles were then placed in a
prewarmed GSA rotor and centrifuged in a Sorvall
RC2-B centrifuge at 37°C. After centrifugation, the
cytochalasin B-containing medium was replaced with
mediumplus serum, and the monolayers were put at
370C
untilthey returned to their normal morphology.Control cellsweretreatedsimilarly except that they
were notcentrifuged. The efficacy of enucleation was
estimated by Giemsa staining the monolayers and
counting the percentage ofcellswhichstillretained a
nucleus. Thecellloss was estimated by both
micro-scopicinspectionandcountingcells,before and after
enucleation, byuseofahemocytometer. Specific
de-tails of theprocedure used for each cellline are as
follows: for L cells, MEM + 5% AFCS + 10 jAg of
cytochalasin B/ml, 15-minpreincubation,
centrifuga-tionat8,500 rpm for45min,1-hrecovery; for BSC-40
cells,MEM+10%AFCS+10
Mlg
ofcytochalasin B/ml,nopreincubation, centrifugation at 8,500 rpm for 20
min, 1-h recovery; for CVCcells, BME + 10%AFCS
+50,ugofcytochalasin B/ml, 1-hpreincubation,
cen-trifugationat10,500 rpm for45min,30-minrecovery.
Single-step growth experiments. After the
re-covery interval of the enucleation procedure,
mono-layerswerewashed twice with
370C
medium(minusserum) and then infected with 0.5 ml ofanappropriate
dilution of purified vaccinia virus in
phosphate-buffered salinecontaining 1mMMg2'.After 30 min
at 25°C, the virusinoculum was removed, and the
on November 10, 2019 by guest
http://jvi.asm.org/
monolayers were washed twice with 370C medium
(minusserum).Mediumplusserum(4ml)wasadded,
and thedisheswere put at370Cand 5%C02.Atthe
indicatedtimesduplicatedishes ofvirus-infectedcells
andcytoplasts were put at -70°C to stop further virus
development. The following day the dishes ofcells
were thawed, andalldebris was loosened with the aid
of a rubber policeman and an additional 1 ml of
medium. The entireinfected-cell lysate wascollected
and refrozen. Infectious virus titers were then deter-mined as described earlier. Experiments measuring virus macromolecular synthesis were carried out
sim-ilarlyexceptthat themonolayerswerewashed twice
with medium(lacking serum and the appropriate
me-tabolite) prior tobeing labeled with the radioactive
precursor.
SDS-PAGE. Sodium dodecyl
sulfate-polyacryl-amidegel electrophoresis (SDS-PAGE) performed on
12.5%slabgelsby the methods of Studier (28). Small
samples ofradioactively labeled infected cellswere
acetone precipitated, dried, resuspended in sample
buffer,boiled for 5min,and thenanalyzedon
1-mm-thick slab gels. Autoradiography of dried gels was
carriedoutwith Kodak RPRoyalOmat medical
X-rayfilm,and the films were later developed in a Kodak
Rapid-Processautomaticdevelopingunit.
Electronmicroscopy. Multipledishesof BSC-40
cells andcytoplasts (106 cells/dish,98.6%enucleation,
2% cell loss) were infected with vaccinia virus at a
MOI of 10. At 0, 3, 6, 9, and 12 hpostinfection,cells
wereremoved from the dishes withtrypsin-EDTA and
harvestedby low-speed centrifugation.Thecellswere
suspended in 1 ml of 2.5%glutaraldehyde in 0.15 M
cacodylate buffer (pH 7.4) containing 2 mM CaCl2.
After 2 min at250C,thesuspensionwascentrifuged,
the supernatant fluid was decanted, and the
undis-turbedpelletswerefixed in the aboveglutaraldehyde
for30minat250C.Thepelletswerepostfixedin 1%
osmium tetroxide in 0.15 Mcacodylate (pH7.4) for 30
minand then stained en bloc with 0.5%uranylacetate
in acetatebuffer (pH 5.0) for 30 min,dehydratedin
acetone, and embedded inEpon812(14).Thin sections
were cut on a Porter-Blum MT-2 ultramicrotome,
stained withuranylacetateand leadcitrate(27),and
examinedin a JEOL lOOBelectronmicroscopeat 60
kV.
RESULTS
Lackofvaccinia virusreplicationin
enu-cleated mammalian cells.Cytochalasin B-in-ducedmassenucleation ofmonolayersof animal cells isawidelyusedtechniquewhichhasproven
useful in the study of various virus-host cell interactions (25). We have utilized this meth-odologytoenucleate several different cell lines
to testtheeffects of thisprocedureontheability
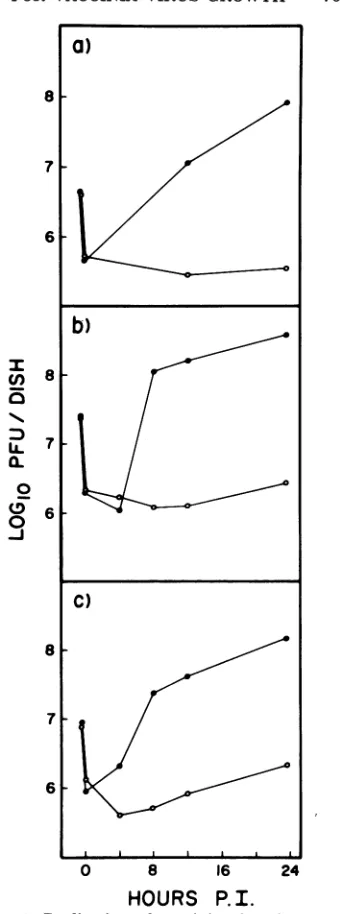
of thecellstosupportasubsequent infectionby vacciniavirus. Figure 1 shows the results
ob-tained by infecting CVC, L, and BSC-40 cells
andcytoplastswith vaccinia virus at low multi-plicity and monitoring infectious virus growth for 24 h postinfection. Low-multiplicity infec-tions, 5 to 10 PFU/cell, were chosen for two
I
U)
5
0
0-j
0 8 16 24
[image:3.505.277.449.64.521.2]HOURS P. I.
FIG. 1. Replication ofvaccinia virus in control
and enucleated mammalian cells. Subconfluent
monolayersofintactcells(0)orcytoplasts(0)were
infectedwithvacciniavirus at a MOIof 5. Duplicate
dishes were removed at theindicated times,and virus
washarvested and titered on BSC-40 monolayers as
described. (a) CVC cells (9 x 105 cells/dish, 99.8%
enucleation, 5% cell loss); (b) L cells (5 x 106
cells/dish,99.5%enucleation, 8% cell loss); (c)
BSC-40cells(1.5x 10'cells/dish,98.4%enucleation, 3%
cell loss).
reasons.First,itwasfoundthat theseconditions
gave the highest relative virus yield in 24 h.
Second, high-multiplicity infections elicitedan
acute destructive CPEwhich cytoplasts might
on November 10, 2019 by guest
http://jvi.asm.org/
708 HRUBY, GUARINO, AND KATES
have beenless abletotolerate andrepair than wholecells. It is obvious inFig. 1that,although vaccinia virusgrewquite well inallthreecontrol cells (80to 105PFU/cell),itgrewpoorly ifatall
in thecognate cytoplasts (0.4to 1.2 PFU/cell).
The small increment of increase in infectious virus titer inthecytoplastpreparationscould be accounted for by the 1 to 3% remaining
nu-cleated cells,soit isdoubtful whether enucleated cells are capable of producing any infectious
progenyvaccinia virus. Also, noplaquesorfoci
of infection could be observedonmonolayersof cytoplastsinfected with50to100PFU of virus
perdish andstained with neutral redatvarious timespostinfection. As acontrol,the above
ex-periments wererepeatedwith vesicular
stoma-titis virus whichwasabletogrowto30to100% ofcontrol titers, depending on initial MOI, in each of the variouscytoplasts,inagreementwith previous work (7). One interesting experiment would have been to enucleate vaccinia virus-infected cells at various times postinfection to
ascertain when the nucleuswasrequiredin the viralgrowthcycle.Unfortunately, the combina-tion of virus-induced CPE and centrifugation caused the cellstofall off the dishduring
enu-cleationattimes after3 h,and thus the above experimentwas nottechnicallyfeasible.
Furtherstudies into theapparentnuclear
re-quirement for vacciniavirusgrowthwerecarried
outwithBSC-40 cells. Thissystemwas chosen foravarietyofreasons:itaffordedease,rapidity, andreproducibilityof theenucleationprotocol; the biosynthetic capabilities of BSC-40
cyto-plasts were very high, as will become evident later; BSC-40 monolayers demonstrated the highest plaquing efficiency of several cell lines
thatwere tested, soexperiments would be
car-ried out and titered on homologous cells; and, finally,results could becomparedtoearlier work whichusedasimilar cellline,BSC-1 (22).
Experiments designedtoruleoutpossible
ar-tifactual results duetoside effects from thedrug cytochalasinBonvacciniavirusreplicationwere
carriedout.Pretreatmentof BSC-40monolayers
with 10 ,ug ofcytochalasin B/mlorpresence of
thedrugthroughoutthe courseof infectiononly reduced thevirusyieldby 21 and32%,
respec-tively.Thisagrees withearlierstudies in which 4 ,ug of cytochalasin B/ml was used (6). Note thatthe1-hpretreatmentregimenwasthe nor-mal control used in most of the experiments reportedhere. Underamicroscopeitwas
appar-ent that BSC-40 cells rapidly extruded their
nucleusinresponsetocytochalasinBto apoint
whereitwasconnectedtothe mainbodyof the
cellbyonlyaslender threadofcytoplasm.Even
in this grossmorphological state, thecells
sup-ported vacciniavirusgrowth,whereasbreakage
of the cytoplasmic stalk withcentrifugal force
during the enucleation procedure resulted in a
total loss of infectious virus production (Fig. 1).
Onetrivial explanation for the failure of
vac-cinia virus to replicate in enucleated BSC-40
cells is that during the period of timerequired
for virusdevelopment (9to 10h) the cytoplasts deteriorate too rapidly for mature virus to be
formed. Several lines of evidenceargue against
this. First of all, other viruses with generation
timessimilartothatofvaccinia virus growwell inenucleated BSC-1cells(8),althoughthe
com-plex morphogenetic pathway ofvaccinia virus may complicate this sort of analogy. Second, BSC-40cytoplastsareabletotakeup and retain
the vitaldye neutral red for 36 to 48 h. Third,
infectingBSC-40cellsand cytoplastsatseveral
MOI (1, 10, or 100) to minimize or maximize vaccinia virus-inducedcellularCPEdidnot raise orlower the ability of cytoplaststosupport virus
growth, whereas there was aninverse relation-ship between multiplicity and virus yield in
whole cells. Microscopic observations
demon-strated that whatever virus function elicited CPE (rounding up and clumping), it was
ex-pressed with the same timing and severity in
cytoplastsasin wholecells.Finally, carryingout
experiments at
310C,
acondition knowntopro-long the integrity and pro-longevity of cytoplasts
(25), didnotallow vaccinia virusreproduction in
cytoplasts but it did lower the virus yield in
wholecells50% (datanotshown).
Expression of both early and late virus functions in enucleated cells. The ability of vaccinia virustodirect itsownmacromolecular synthesis andtoinhibit host-specificevents was next examined. Figure 2 shows the results of
pulse-labeling vaccinia virus-infected BSC-40
cellsandcytoplasts with
[3H]thymidine
to mea-sureDNAsynthesis. Incorporation of the radio-active label showed virtually identical kineticsin bothcases,withthemaximumrateoccurring at 3 h postinfection. At this time, the virus-infected cytoplastsweresupporting synthesis of
vaccinia virus DNA at about 80% of control
levels,althoughthiscapability dropped off more
rapidlyatlatertimes(3.5 to 5 h) in the cytoplast
preparations. To ensure that what was being
measuredwas, infact,vaccinia virus DNA
syn-thesis, an identical experiment wascarried out
except that cytoplasmic extracts were made at
each time point andcounted. The results were
identical to thosein Fig. 2, indicating that
vir-tually all of the radioactive counts were being incorporated in the cytoplasmic fraction, i.e., into viral DNA. The fact that vaccinia virus
DNA wassynthesizedincytoplastsstrongly
im-plies that the virus-coded early functions for DNAbiosynthetic enzymes and core uncoating
on November 10, 2019 by guest
http://jvi.asm.org/
12 0
8
100
0
4I 1
I
-J 0
I-O 2 4 6
HOURS P.I.
FIG. 2. Vaccinia virus DNAsynthesis ininfected
BSC-40 cells. Intact BSC-40 cells (0) orcytoplasts
(0) (2.75x 106cells/dish, 98.7%enucleation,2%cell
loss) wereinfectedwith vacciniavirusat aMOIof
10.Atthe indicated times, disheswerepulsedwith
[3H]thymidine (2,uCi/ml) for 10min, and then the
trichloroacetic acid-precipitable radioactivity per
dishwasdetermined.(a)Pulse-labeled vaccinia virus
DNAsynthesis.(b)Cumulativeplot ofvaccinia virus
DNA synthesis. Note: at 3 h, dishes of uninfected
BSC-40 cells andcytoplasts incorporated154,000and
1,200 cpm,respectively.
had been expressed (24). The question ofthe
newlysynthesized DNAbeing transcriptionally active in enucleates was next examined. RNA
synthesis was not studied directly, but rather
theviruspolypeptides beingcoded forwere
re-solved by SDS-PAGE. In the absence of any
stringent translationalcontrol,theviralproteins being translated should reflect theamountand
types of vaccinia virus mRNA species being
transcribedatvarioustimespostinfection. Figure3shows the results of[35S]methionine pulse-labeling of BSC-40 cells and cytoplasts
infected athigh multiplicity(50PFU/cell) with
vaccinia virus. The top panel exhibits some of
the kineticconsiderations. The first point to be
made is that BSC-40 cytoplasts, whether
in-fected ornot,initiallysynthesized protein at 60
to 70%of the rate of controlcellsand then lost
their protein synthetic capabilities slowly
throughout the duration of the experiment. In both cells and cytoplasts, a rapid inhibition of
host-cellprotein synthesis occurred during the
first1.5h, but thiswasrapidly obscuredas
virus-specific proteins began to be made (3 to 8 h).
This effect was even more dramatic when the
FIG. 3. Vaccinia virus protein synthesis in BSC-40
cells. IntactBSC-40 cells(0)or cytoplasts(0) (2.4 x
106 cells/dish, 98.0% enucleation, 2% cell loss) were
infectedwith vaccinia virusataMOIof 50. At the
indicatedtimes,disheswerepulsedfor20minwith 5
,uCiof[3S]methionine/ml. Cells were harvested into
0.5mlof ice-cold 1 mMTris-hydrochloride (pH 9).
Uninfected cellsandcytoplastswerealso monitored
(dashed lines). (a)Samples of100Il were processed
todeterminehottrichloroacetic acid-precipitable
ra-dioactivity. (b)Amountsof50,ul of the infected
sam-pleswere analyzed by SDS-PAGE andsubsequent
autoradiography (lanes 1-5, vaccinia virus-infected
BSC-40 cells at 0, 1.5, 3, 5, and 8 hpostinfection;
lanes6-10,vacciniavirus-infected cytoplastsat0, 1.5,
3, 5,and 8 hpostinfection).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.505.73.218.66.331.2] [image:5.505.252.447.132.522.2]infectionswerecarriedout at alow multiplicity
(5 PFU/cell), in which case virus protein
syn-thesis was only a smallpercentage of the total.
Figure 3b is an autoradiograph of an
SDS-polyacrylamide gel analysisofthe
[35S]methio-nine-labeledproteins synthesizedinthe
experi-ment shown in Fig. 3a. Invirus-infected whole
cells (lanes 1-5) and enucleated cells (lanes
6-10), the patternofgeneexpressionwas
iden-tical:at 0hprimarily host proteinswerelabeled,
at 1.5 hearly vacciniavirus proteins appeared,
at 3h (coincident withpeak viralDNA
synthe-sis) therewerebothearlyandlate vaccinia virus proteins beingmade, andat 5and8hlate viral proteins predominated.Also, itwasobviousthat
the same proteins in the same relative ratios werebeing synthesizedinthe enucleatedcellsas
in the whole cells, albeit in somewhat reduced
amounts. The early andlate vaccinia proteins
wereidentifiedby comparisonto standard vac-cinia virus proteins labeled in the presence or
absence of vaccinia virus DNA synthesis (21;
Fig. 4). It has previously been shownthat
vac-cinia virusproteinsundergo normalcleavagesin
enucleated cells (22), andwe were also ableto
substantiate this (datanot shown). The status
of otherproteinmodifications in vaccinia virus-infected enucleatedcells, suchasacetylationor
phosphorylation, isnotyetknown.
Itwasof interest to seehowlong enucleated cellswerecapable ofsynthesizing vacciniavirus
proteins. Therefore, infectionswerecarried out
in the presence of cytosine arabinoside, which
inhibits DNA synthesis and hence late gene
expression. Figure 4 showsthat, although host protein synthesiswasrapidly shut off, early
vac-cinia proteins continued to be synthesized in
enucleated and whole cellsformore than24h.
Thus, it wasobvious that enucleatedcellshave
thecapabilityofmaking largeamountsof virus protein forlongperiods of time. It was
interest-ing to note that in the absence of late gene
expression there was a great reduction in
ob-served morphological CPE, thereby implying that this effect was the result of a late gene function.
DNA packaging and viral
morphogene-sis.Theassemblyof vaccinia virus constituents
into matureinfectiousparticlesis acomplicated
multistepprocess which has beenonlypartially
characterized(17). It waspossiblethat,although
vaccinia virus-infected enucleated cells
pro-duced all of the necessary viral components,
criticalcatalytic concentrations of one or more
may not have beenreached, thereby inhibiting
morphogenesis. To address this question, we
used the drugrifampin, which has been shown
to completely block vaccinia virus assembly
while allowing continued expression of both
.. X \.
HOU.8 P
R E.
H0iP' PT
b...
FIG. 4. Vaccinia virus protein synthesis in the
presenceof cytosine arabinoside(CAR).BSC-40 cells
(0) or cytoplasts (0) (2.3 x 106 cells/dish, 97.6%
enucleation, 3%o cellloss)wereinfected with vaccinia
virusataMOIof50.CAR waspresentat25pg/ml
throughout the infection. Disheswerepulsed with2
,ICiof[35S]methionine/mlatthe indicatedtimes and
analyzed as described in the legendofFig. 3. (a)
Vaccinia virus protein synthesis. (b) SDS-PAGE
analysis of [35S/methionine-labeled vaccinia virus
proteins (lanes 1-5, 50,ul ofvirus-infected BSC-40
cellsat0, 4,8, 12, and24hpostinfection; 6-10, 100
1lofvirus-infected cytoplastsat0, 4, 8, 12, and24h
postinfection.
early and late virus functions (19). Upon drug
removal this effect israpidly reversed, and
infec-tiousparticles arequicklyformed. Table 1 shows
the results ofsuch a treatmentonvaccinia
virus-infected BSC-40cells and cytoplasts. The rifam-pin block-reversal step elicited a 3.5-fold increase
of virusgrowthinwhole cells. No similar
ampli-fication was observed in the cytoplast
prepara-tions.
Earlier it was demonstrated that vaccinia
vi-rus-specific DNA synthesis occurred at a high
rateininfectedcytoplasts (Fig. 2),butthe
sub-sequent fate of this DNA was not examined.
Table 2 shows the results obtained when the
viral DNAsynthesized from2to4h
postinfec-tion was labeled with [3H]thymidine and then
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.505.269.458.69.378.2]NUCLEAR REQUIREMENT FOR VACCINIA VIRUS GROWTH
TABLE 1. Effectonrifampinonthe yield of
vaccinia virus from infected BSC-40 cellsa Timeof Control Enucleatedcells rifampin
removal PFU/dish PFU/cell PFU/dish PFU/cell (h)
0 3.4 x 107 50.3 3.5 x 105 0.5
8 1.2 x108 177.8 3.7x 105 0.6
24 3.6 x105 0.5 3.3 x 105 0.5
aBSC-40
cells
and cytoplasts (7 x 105cells/dish,
99.5% enucleation, 1% cell loss) were infected with
vaccinia virus at a MOI of5inthe presence of 100,tg
of rifampin/ml. Drug-containing medium was
moved at the indicated times postinfection and
re-placed with medium alone. At 24 h, infections were
stopped and titers were determined as usual. All assays
weredone induplicate.
evaluatedatvarious timesforpackaging,as
as-sayed by conversion of the labeled DNA froma
DNase-sensitivetoaDNase-resistant form(19).
Whereasmorethan50% of theviralDNAmade
inwhole cells became resistanttotheactionof
DNase, therewas no change in the DNase
sen-sitivity of cytoplast viral DNA in 20 h. This
observation was supported by the failure ofus
andothers (22)toisolatephysical particles from
vaccinia virus-infectedenucleated cells. Vaccinia
virus infections of BSC-40 cells and cytoplasts
were carried outin thepresence of
[3H]thymi-dine. The labeledlysates, plusunlabeledcarrier
virus, werethensubjectedtothevirus
purifica-tion scheme described in Materials and
Meth-ods. Less than 5% of the radioactivity banding
with maturevirus from controlcellswasfound
ata corresponding position from the cytoplast
preparation (data notshown). Taken together,
thesedataimplythat viral DNAsynthesizedin
the absence of a host-cell nucleus was either
neverpackaged orwas packagedin an
osmoti-callyfragile form thatwasdisrupted during the
DNase assays and particle purification
proce-dures.The exact nature ofmorphogeneticevents
happening in vaccinia virus-infected cells and
cytoplasts was then examined in an electron
microscope.
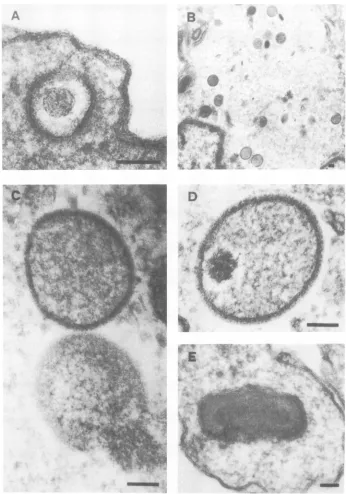
Figure 5 displays typical examples of virus
developmentalforms observed in vaccinia
virus-infected BSC-40 cells during a single growth
cycle experiment.Fromuncoatingtoproduction
ofnewinfectious progenyvirus 9 to 12 hlater,
an orderly set of morphogenetic steps can be
seen,similartothosepreviously described (17).
Vacciniaviruswastakenupanduncoated(Fig.
5A), the cores subsequently broke down and
became "viroplasm" or "virus factories" (Fig.
5B), membraneswithprominent spicules
enve-loped "viroplasm" (Fig. 5C), andDNA
conden-sationbeganatoneend(Fig. 5D) andproceeded
to the formation ofmature particles (Fig. 5E).
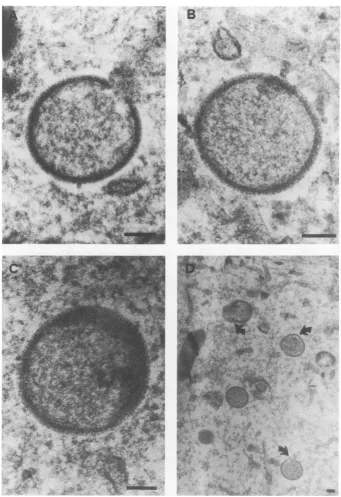
Figure 6 demonstrates that thiswasnot true
in virus-infectedenucleatedcells.Although the
earlystepsofmorphogenesisuptoformation of
immature particles (Fig. 6A and B) appeared
normal, beyond this development went awry.
DNA condensation, ifseenatall,wasaberrant,
giving a mottled appearance to the immature
particles (Fig. 60). At later times (6 to 12 h
postinfection), membrane aberrations and bi-zarreviral formswereapparent(Fig. 6D). Virus
development seemed to be arrested ata point
analogoustotheclass Jmutants of Dalesetal.
(5).Also, virus factories of similar size and
den-sitywereobserved in both whole and enucleated
cells.
DISCUSSION
Pennington and Follett originally observed
that vaccinia virus wouldnotgrowinenucleated
BSC-1 cells (22). These experiments were
car-riedout atahigh MOI under conditions where
viral yields in control cells wererelatively low
andoverallviralgeneexpression and DNA
rep-lication in enucleated cells were depressed. It
could beargued that this resultwasduetosome
factors other than enucleation per se. For
ex-ample, the high MOI could cause severe CPE
whichwould result in aninhibition ofthe
pro-duction of critical concentrations of viral
com-ponentsrequired forpropermaturation in
BSC-1cells.However,wehavebeen abletoshowthat
inseveralcell lineslow-multiplicity vaccinia
vi-rus infectionsresulting in good viral growthin
cellsdidnot elicit any viralgrowth in the
cor-responding cytoplasts (Fig. 1). Byallthe criteria
tested, both early and late viral functionswere
expressed properly andatrelatively high levels
in enucleated BSC-40 cells (Fig. 2 and 3), but
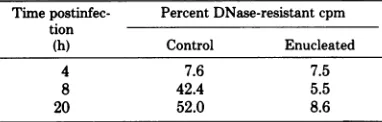
TABLE 2. Conversionofvaccinia viruscytoplasmic
DNAtoaDNase-resistantforma
Timepostinfec- Percent DNase-resistant cpm tion
(h) Control Enucleated
4 7.6 7.5
8 42.4 5.5
20 52.0 8.6
aBSC-40
cells
andcytoplasts (3.5x 106cells/dish,
98%enucleation,3%cellloss)wereinfected with
vac-ciniavirus at aMOI of10.From2 to 4hpostinfection,
thedisheswerelabeledwith 2,uCiof[3H]thymidine.
Atthe indicatedtimes,duplicate plateswereremoved,
cells were harvested, and acytoplasmic extract was
prepared (19). One half of theextractwasprecipitated
withtrichloroacetic acid and counted; theother half
wasmade50yg/mlinpancreaticDNaseand incubated
at37°C for30niinpriortotrichloroacetic acid
precip-itation.
71
VOL. 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.505.263.454.511.572.2]*~~~~'F- F# = m
FIG. 5. Electron micrographs ofthin sectionsofvacciniavirus-infectedBSC-40 cells. Displayed are the
typicalvaccinia virusmorphogenetic formsthatwereobservedattheindicated timespostinfection. (A)Virus
particle shortlyafterpenetration, Oh. (B)Virusfactory,3h.(C)Immatureparticlesintheprocess of enveloping
DNA, 6h. (D)Partiallycondensed DNAwithin immatureparticle,9h. (E)Mature virusparticle, 12h. Bar
=0.1
ALM.
the viral DNA synthesized in enucleated cells the maturation of the viral components into
wasnotconvertedtoaDNase-resistant formas matureprogenyparticleswasaberrantorabsent
itwasin controlcels(Table 2).Amoredetailed inenucleatedcells (Fig. 5and6). Incontrast to
electron microscopic study demonstrated that PenningtonandFollett (22),wefoundno
differ-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.505.92.438.78.572.2]B
A&
i-,-4.. ,,ll.. t
-A. I 1
FIG. 6. Electronmicrographs of thin sections of vaccinia virus-infected BSC-40 cytoplasts. The
predomi-nantforms observedatthe indicated timespostinfectionare shown. (A)Immature particle enveloping DNA,
3h. (B) Completely enveloped particle,6h.(C) Immature particle exhibiting faultyDNAcondensation, 12 h.
(D)Immatureparticleslateininfection withaberrant membranes and noapparent DNAcondensation,12 h. Bar=0.1,im.
encein the number or size of virus factories in
the enucleated cells. From the data reported
here and from other studies of virus-host cell
interactionsusingenucleatedcells(25), it is
un-likelythat this result is due to the enucleation
procedure itself. Therefore,ourdatasupport and
extend earlier work (22) and indicate that the
hostnucleusplays some importantrole in
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.505.81.422.74.571.2]cinia virus growth. This also agreeswith other studiesusing metabolic inhibitors (15, 26). Thus, it would seen that, like frog virus 3, vaccinia viruscan nolonger be consideredacytoplasmic
virus in thestrictsenseof the word (23).
Ifone acceptsthe notion that vaccinia virus
requiresthe host-cell nucleus in orderto
repli-cate, then several interesting questions are
raised. First, does vaccinia virus merely require the physical environment of the nuclear body,
or mustthenucleus be active? Previous studies
with phytohemagglutinin-activated leukocytes suggested an active nuclear requirement (16). However, because of the quiescentstate of the protein synthetic machinery in resting
leuko-cytes,theseresultsmustbe viewed with caution.
Asecondquestionis whether theneeded nuclear functionisacomponent or an enzymeand, sim-ilarly, whether this function is normally
ex-pressed inuninfected cellsoris inducedby the vaccinia virusinfection. Onefinding whichmay
bepertinent is that of LaColla and Weissbach (13), whoshowed thatsomevacciniavirus DNA replication occursin the nucleus. Itis possible that replication of vaccinia virus DNA in the nuclear environment is required for expression of the needed nuclear function. However, the observed absence of UV-enhanced reactivation ofsubsequent vaccinia virus growth under
con-ditions which stimulate the growth of a bona fide nuclear virus such asherpes simplex virus (2) questions this hypothesis.
Whatever itsnature,therequirement for
nu-clear function becomes apparentduring the
as-sembly of vaccinia virus. Itmaybe asituation analogoustothegroElocus of Escherichiacoli, where a host-coded function is required for
properphage morphogenesis (4). Itisalso
pos-sible thatone or moreof the virioncomponents
(DNA,RNA,orprotein)aresynthesizedor mod-ifiedimproperly in the absence of the nucleus. Experimentally, thismay notbe detected until
this component is unable to interact
produc-tively during assembly,hencefaulty morphogen-esis.
It is interesting to consider how these data
compare with earlierresearch on the effectsof
theanti-vaccinia virusdrugVirazole orribavirin
(1-fi-D-ribofuranosyl-
1,2,4-triazole-3-carboxam-ide) (12). This drug allows expression ofboth
earlyand late viralfunctionsbutgreatlyinhibits
production of infectious progeny. It has been
speculated that Virazole inhibits some specific
late viral function. An alternate explanation is
that the drug inhibitstherequiredhostnuclear
function.
Obviously, much more work remains in the
studyof vaccinia virusreplication and howthe
virus interacts with both the cytoplasmic and
nuclear compartments of the cell. Current
re-search efforts in this laboratoryarecenteredon
using intact cells to investigate the molecular
nature of the apparent nuclear requirement.
Thisareaof research wouldseem tobe
impor-tant in understanding vaccinia virus-host cell interactions and possibly in elucidating previ-ously undescribed host-cellfunctions.
ACKNOWLEDGMENTS
Thiswork wassupported by Public Health Service training grantCA-09176-04 from the National Cancer Institute and by Public Health Service grantAI-11839-01 from the National Institute of Allergyand Infectious Diseases to J.R.K.
Wethank D. L.Lynn for excellent technical assistance and R. C. Condit for many valuable discussions and suggestions. We alsoacknowledge the members of J. Lucas's laboratory forsharing theirexpertisein cellenucleation procedures.
LITERATURE CITED
1.Becker, Y., and W. K.Joklik.1964.Messenger RNA in cells infected with vaccinia virus. Proc. Natl. Acad. Sci. U.S.A.51:577-585.
2. Bockstahler, L.E., andC. D.Lytle. 1977. Radiation enhanced reactivation of nuclearreplicating mamma-lian viruses. Photochem. Photobiol.25:477-482. 3. Carter, S. B. 1967. Effects of cytochalasins on mammalian
ceLs.Nature(London)213:261-264.
4.Casjens, S.,and J.King.1975.Virus assembly. Annu. Rev. Biochem. 44:55-611.
5. Dales, S., V. Milovanovitch, B. G. T. Pogo, S. B.
Weintraub, T. Haima, S. Wilton, and G.
Mc-Fadden.1978.Biogenesisof vaccinia: isolation of con-ditional lethalmutantsand electronmicroscopic char-acterizationof theirphenotypicallyexpressed defects. Virology 84:403-428.
6. Deitch,A.D.,S.G.Sawicki,G. C.Godman, and S. W.Tanenbaum.1973.Enhancement of viral infectiv-ity bycytochalasin. Virology 56:417-428.
7. Foilett,E.A.C.,C. R.Pringle,W.H. Hunner, and J. J. Skehel. 1974.Virus replication in enucleatecells:
vesicular stomatitis virus and influenza virus. J. Virol. 13:394-399.
8. Follett,E.A.C., C.R.Pringle,and T. H.Pennington. 1975.Virusdevelopment in enucleate cells: echovirus,
poliovirus, pseudorabies virus,reovirus, respiratory syn-cytial virus and semliki forest virus. J. Gen. Virol. 26: 183-196.
9. Gafford,L.G.,andC. C. Randall.1976.Virus-specific
RNAand DNA in nuclei of cells infected with fowlpox virus.Virology 69:1-14.
10.Goorha, R.,D. B.Willis,and A.Granoff.1977. Mac-romolecularsynthesisin cells infected by frog virus3. VI.Frog virus 3replicationisdependentonthecell
nucleus. J. Virol. 21:802-805.
11.Joklik,W.K.,and Y. Becker.1964.Thereplicationand uncoating of vacciniaDNA.J. Mol. Biol.10:452-474. 12.Katz,E.,E.Margalith,and B. Winer.1976.Inhibition of vaccinia virusgrowth bythenucleosideanalogue
1-,8-D-ribofuranosyl-1,2,4-triazole-3-carboxamide
(vira-zole, ribaviran).J. Gen. Virol.32:327-330.
13.LaColla, P., and A. Weissbach. 1975. Vacciniavirus infection of HeLa cells.I.Synthesis ofvaccinia DNA in hostcellnuclei. J. Virol.15:305-315.
14.Luft, J. H. 1961.Improvementsinexpoxyresin embed-dingmethods. J.Biophys. Biochem. Cytol. 9:409-412. 15.Magee, W. E., and 0. V. Miller. 1968. Initiation of vaccinia virusinfection in actinomycin D-pretreated
cells.J. Virol.2:678-685.
16. Miller,G., andJ.F. Enders.1968.Vaccinia virus
on November 10, 2019 by guest
http://jvi.asm.org/
cationand cytopathic effect in cultures of phytohem-agglutin-treated human peripheral bloodleukocytes. J. Virol.2:787-792.
17. Morgan, C. 1976.Vaccinia virus reexamined: develop-mentandrelease. Virology73:43-58.
18.Moss, B. 1974.Reproduction of poxviruses,p.405-474.In
H.Fraenkel-Conrat and R. R. Wagner (ed.), Compre-hensivevirology, vol. 3. Plenum Press, New York. 19. Moss, B., E. N.Rosenblum, E. Katz, and P. M.
Grim-ley. 1969.Rifampicin: aspecificinhibitor ofvaccinia virus assembly. Nature(London) 224:1280-1284. 20. Ortin, J., and E. Vinuela. 1977. Requirement of thecell
nucleus for African swine fever virusreplication in Vero cells. J.Virol. 21:902-905.
21. Pelham, H. R. B., J. M. M.Sykes, and T. Hunt.1978. Characteristics ofacoupled cell-free transcription and
translationsystemdirectedby vaccinia cores.Eur. J.
Biochem. 82:199-209.
22. Pennington, T. H., and E. A. C. Follett. 1974. Vaccinia virusreplication in enucleate BSC-1cells:particle
pro-duction andsynthesis of viral DNA and proteins. J.
Virol. 13:488-493.
23. Portner, A., and C. Pridgen. 1975. Viral replication,p.
27-42.In A. Notkins (ed.), Viral immunology and im-munopathy. AcademicPress Inc., New York. 24. Prescott,D.M.,J.Kates,and J.B. Kirkpatrick. 1971.
Replication of vaccinia virus DNA in enucleated L-cells. J. Mol. Biol. 69:505-508.
25. Pringle, C. R. 1977. Enucleationas atechnique in the
study of virus-host interactions. Curr. Top. Microbiol. Immunol. 76:49-82.
26. Reich, E., and R. M. Franklin.1961.Effect ofmitomycin C onthe growth ofsomeanimalviruses.Proc.Natl.
Acad. Sci.U.S.A. 47:1212-1217.
27. Reynolds, E. S. 1963. Theuseof lead citrateathigh pH
as anelectronopaquestain in electronmicroscopy. J.
Cell Biol. 17:208-213.
28. Studier, F. W.1973.Analysis of bacteriophage T7 early RNAs and proteins on slab gels. J. Mol. Biol. 79:
237-248.
29. Walen, K. H. 1971. Nuclear involvement in poxvirus infection. Proc. Natl. Acad. Sci. U.S.A. 68:165-168.
on November 10, 2019 by guest
http://jvi.asm.org/
![Figure 3infectedpulse-labeling shows the results of [35S]methionine of BSC-40 cells and cytoplasts at high multiplicity (50 PFU/cell) with](https://thumb-us.123doks.com/thumbv2/123dok_us/1519655.104555/5.505.252.447.132.522/figure-infectedpulse-labeling-shows-results-methionine-cytoplasts-multiplicity.webp)