0022-538X/93/074442-06$02.00/0
Copyright ©D 1993,AmericanSociety for Microbiology
Analysis of the Terminal Repeat Binding Abilities of Mutant
Adeno-Associated
Virus Replication Proteins
QICHENGYANGANDJAMES P. TREMPE*
Departmentof Biochemistry and Molecular Biology, MedicalCollege of Ohio, Toledo, Ohio 43699-0008
Received 8February 1993/Accepted 19 April 1993
The adeno-associated virus(AAV) Rep78andRep68 proteins playessential roles in viral DNAreplication,
trans activation of viral geneexpression, and suppression ofoncogene-mediated cellular transformation. By
usinganextensivesetof linker insertionanddeletion mutations in thereplicationgene, wemappedtheregions ofthe Rep78 protein that mediatebindingtothe AAVoriginofreplication invitro. Deletions thatremoved amino acid codons 25to62,88to113,125to256,and346to400abolishedbinding.Alterations in several other regions ofthe protein affected the binding affinityof the mutant proteins. All of the mutant proteins that supportAAV DNAreplicationorp40transactivation boundtothe terminal repeatsequence,thusverifyingthe
importance ofbindingfor thesefunctions. Severalmutantrepgenesthat failedtosuppressoncogene-mediated cellular transformationproduced proteinsthatwerecapableofbindingtothe AAV terminal repeatsequences.
Adeno-Associated Virus (AAV) is a defective human
parvovirus that efficiently reproduces in cells coinfected with a helper virus, such as adenovirus orherpesvirus (2).
Replicationof thesingle-strandedAAVgenomerequirestwo
elements: the terminal repeats at both ends, which form T-shaped hairpins thatserve asorigins ofAAV DNA repli-cation (4),andthereplication (rep)gene(2, 6, 18). None of
the functions provided by the helper virus are absolutely
requiredfor AAV DNAreplication (2, 16, 23).Therepgene
encodes at least four overlapping Rep proteins, Rep78, Rep68, Rep52,andRep40,derivedfrom alternate transcrip-tion initiation at either the p5 or the p19 promoter and mRNA splicing. The two largest Rep proteins, Rep78 and Rep68, bind specifically to the AAV terminal repeats and have site-specific and strand-specificendonuclease activity which resolves the covalently joined ends of DNA replica-tion intermediates (1, 7-9, 17). Theyalsotrans activate the AAV capsid (cap) gene promoter (p40) (10, 19). Several cis-actingelements in thevicinityof the AAV transcription
promotershave been identifiedasnecessaryforRep protein-mediated p40 promotertrans activation (11). The terminal repeat sequences can also serve as cis-acting elements for this phenomenon. The smaller Rep proteins, Rep52 and Rep4O,arenotinvolved in AAV DNAreplicationand donot bind to the AAV terminal repeat sequences (3, 9). This suggests that the amino-terminal portion of Rep78 and Rep68,which isabsent inRep52andRep 40,isimportantfor DNA binding. Since carboxyl-terminal truncatedrep gene mutantssupportAAV DNAreplication (6, 18, 20)andRep68
canbindtothe terminal repeats(8),thecarboxylterminusof theRep proteins isnotnecessaryfor DNAbinding. Others have documented a preliminary screening of regions in
Rep78 that are essential for binding to the terminal repeat
sequences(12).Inthisstudy,weexpandedonthatreportby
assayingourpreviously published (25) comprehensive series
ofmutantRep78 proteinsfor theabilitytobindtotheAAV originofreplication.
Preparationof Rep proteinextractsfrom COS-1 cells.All of
our rep gene mutants (Fig. 1) were subcloned into the
pCDM8 expressionvector(InvitrogenCorp.), which utilizes
*Correspondingauthor.
the cytomegalovirus early promoter and a simian virus 40 (SV40) origin of DNA replication, to produce high-level expression of cloned DNAs in COS-1 cells. The wild-type
construct,pCDMrep,containstherepgenefromnucleotides 263to2233 in the AAVgenome.Tomeasurethe levelofRep
protein production, COS-1 cells (4 x 105/60-mm-diameter dish) were transfected with wild-type or mutant rep gene
plasmids. Two days after transfection, the cellswere
har-vestedby scraping andwashed inphosphate-buffered saline. Thecellswerelysedin STM-NPbuffer(10mMTris
hydro-chloride [pH8.0], 0.25mMsucrose, 10 mMMgCl2, 1.0mM
phenylmethylsulfonyl fluoride, 0.1 mM dithiothreitol, 0.5% NonidetP-40), and nucleiwerepelleted bycentrifugationat
2,000 xg.Thesupernatantfractionswerediscarded,except for those from cells transfected withpdl1684-1878,oneofthe
putative nuclear localization mutants, whichwas used for theDNA-bindingassays.Thenucleiwereresuspendedin30 ,ulofSTM-NPcontaining200 mMNaCl and incubatedat4°C for30min.Thenucleiwerepelletedat10,000 xgfor15min, and theresulting supernatants (nuclear extracts)were used in DNA-binding assays. Figure 2 shows a Western blot
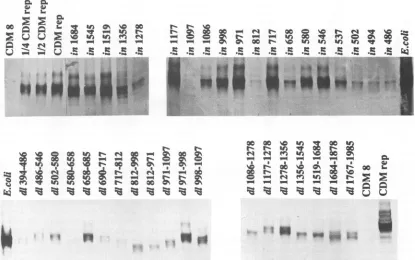
(immunoblot) analysisofaliquotsofthenuclearextractsthat hadbeenseparatedby sodium dodecyl sulfate-12.5% poly-acrylamide gel electrophoresis and probed with antiserum against Rep protein producedinEscherichiacoli (21). Most of theRep protein producedfromthisconstructwasRep78;
however,weoccasionallydetectedverylow levels ofRep52 butno Rep68orRep4O production. Densitometric scanning
analysisindicated thatmostof themutantsproduced levels ofprotein comparable to that from thewild-typerep gene;
however,someof themutantsproduced proteinatalevelas
lowas20%of thewild-typelevel. Low-levelexpressionmay
result from the differential stability of mutant proteins. MutantpinlO97 directs thesynthesis ofRep protein that is detectable via indirectimmunofluorescence,butwehavenot been ableto detect itbyWestern blot assays(26).
Rep78fromrepgene-transfected COS-1 cells binds
specifi-cally to the AAV terminal repeat sequences. To determine
whether the mutant Rep proteins produced in plasmid-transfected COS-1cellswerecapableofbindingtothe AAV terminal repeat sequences, gel mobility shift assays were
performed.To obtainradiolabeled AAVhairpinDNA, plas-midpsub201 (15)wasdigestedwithPvuIIandXbaI and the
4442
on November 9, 2019 by guest
http://jvi.asm.org/
321 502 690
486 546 658j 812
494
H8
6851
Insertion Mutants
993
98 1897 1278
7F
9112
971 1086 1177 1356
537 717
Deletion Mutants
dl
394-486 di486-546dl502-580 dl580-658
dl658-685 dl690-717
di717-812
dl
812-971 d1812-91di92
dl971-1097 dl998-1097
dl1086-1278
dl
1177-1278dl
1278-1356dl
1356-1545 dl1519-1684dl
1684-187898
dl
1767-1985
FIG. 1. Map of the AAV rep gene openreadingframe ofthe 621-amino-acidRep78proteinand locationsofmutations.The vertical lines indicate thelocations oftheXhoI linker insertions, and the horizontal linesatthe bottom indicate the locations of thedeletionmutants indicated. Thenumbers above theinitiation and termination codons refertonucleotide locations inthe AAV genome. The ATG initiation codon for theRep52 andRep4Oproteinsisindicatedat nucleotide 993.
190-bp termini of the AAVgenome were isolated by 1.5% agarose gel electrophoresis in Tris-acetate-EDTA running
buffer andGENECLEAN (Bio101,Inc.)in accordance with the manufacturer's recommendations. The isolated termini were denatured at 100°C and chilled on ice to promote
self-annealing of the hairpin. The hairpin was radiolabeled with the Klenow fragment of DNA polymerase, [c-32P]dCTP, and unlabeled dGTP, TTP, and dATP. The radiolabeled hairpin was separated from unincorporated
nucleotides byelectrophoresisthrougha1.5%
agarose-Tris-w~ .w
C.
CD~~~~0
~~~ne~~~
nM ,NeaI
" MN~~~~~~~10 4 ~ m e iI
""sU~~~~"
M4
Nil *nwuw
anij
ootoo u)
o~~t
- o Fo*-tw > ^ 4
i
^ tF >CDoQ )
oo oo % tn 3 m n
t-
t-n0 O At cc
- - -
-n
V-V V _ voo "Noin
_3
_ _ __3
_ _of
FIG. 2. Production ofmutantRep proteinsfromCOS-1 cells. Westernblotanalysisofnuclear extractsfrom COS-1 cells transfectedwith
thewild-typerepgene(pCDMrep),expressionvectorpCDM8,and various rep gene mutants.Lane E. colicontainedRep protein produced fromanE. coliexpressionvector(21).
1884
1519 1684
1545
2184
URR
1878
C. 'a
t-0z
u
U
I m I I
I n
I
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.81.543.74.284.2] [image:2.612.106.521.426.686.2]A.
B.
1 2 3 4 5
.... .j- .
1 2 3 4 5 6 7 8
.* _ O
_ B -_
-_IF _ii
1 2 3 4 5 6 7 8 9 10 11 12 13 14
B-10
11,1
ww
15 16 17
1i
18
is
Z
_~~~~.
FIG. 3. (A) Binding of wild-type Rep protein from COS-1 cell extracts to theAAVterminal repeatsequences. The bindingreaction containednoextract(lane 1)or1plfroma30-,ul nuclearextractfrom 106 COS-1 cellstransfected withpCDMrep (lane 4)orpCDM8 (lane
5). Lanes 2 and 3 containedtwodifferentpreparations of Rep78 producedfrom recombinantbaculoviruscontainingtherepgene(19). (B) Supershiftassaywith 0.1(lane 2),0.25(lane 3),0.5(lane 4),or1(lanes5to7)plofpCDMrep-or1plofpCDM8 (lanes1and8)-transfected COS-1 nuclearextractintheabsence(lanes1to5)orpresenceof 1(lane 6)or2(lanes7and8)plof ammoniumsulfate-fractionatedantiserum againsttheRepprotein (21). Bindingassays wereperformedwithSV40 origin-containing plasmidsin the reaction mixture. (C)Removal of background binding. Gel retardation assays weredone withnonuclearextract(lanes5 and16)or0.1(lane 4),0.25(lane 3),0.5(lane 2),or
1(lanes 1, 8, 10, 15, and '8)p,lofpCDMrep-or1plofpCDM8 (lanes 6, 7, 9,11to14,and17)-transfectedCOS-1 cell nuclearextractinthe
presenceofnoplasmid DNA (lanes1to6, 11,and 14to15),0.5p,gof linearizedpGEM3Z (lanes9 and10),0.5p,gof linearizedpCDM8 (lanes 7 and8),0.5(lane 13)or1.0(lane 12)pgofpGEMSV,or0.5 p,geach ofpCDM8andpGEMSV (lanes16to18)in thebindingreaction. *,
nonspecificbackground band; B, bound hairpin; F, free hairpin.
acetate-EDTAgel and transferred onto DEAEpaper. The hairpinwaseluted from theDEAEpaperin 10 mM TrisHCI
(pH7.5)-l mMEDTA-1.0 MNaClat65°C.We used5,000
cpmof theradiolabeled hairpin ineachbindingreaction. Forgel mobility shift experiments, 1 pul of nuclearextract wasincubatedwith radiolabeledhairpinDNAandtheassays were performed by electrophoresis through 4%
polyacryl-amidegels essentiallyaspreviously described (7). Figure 3A
showsanautoradiogram of suchanassaywithextractsfrom COS-1 cells that had been transfected with wild-type rep gene plasmid pCDMrep (lane 4) or expression vector plas-mid pCDM8 (lane 5). Lanes 2 and 3 contained nuclear extractsfromtwodifferent preparationsof the Rep78protein produced from recombinant baculovirus (14). Asshown by others(1, 7, 9, 14), three shifted bandscanbeseenwith the
Rep proteinproduced from the baculovirusvector(Fig. 3A, lane2). The samethree bandswerealso detected in pCDM
rep-transfected COS-1 cellextractsuponshorterexposures
(26)orinvarious amountsofextracts(Fig. 3B, lanes4and 5).
To verify the identity of the bound hairpin, supershift
assayswere performed with Rep protein-specific antibody
(21).Rep-specific antibodywasaddedtothebindingreaction after 20 min of incubation of the Rep-containing extracts with the hairpin. The Rep protein-bound hairpinwas
con-verted toa slower-mobility complex thatwas incapable of enteringthegelwhen specific antibodywasincluded in the incubation(Fig. 3B,lanes 6 and7). However, additionofthe
antibodyto extracts frompCDM8-transfected cells did not alter themobilityof thehairpin (lane 8). Theapparentlarge size of the supershifted complex is due to the polyclonal
nature of the antibody preparation that was used in this
experiment. Supershift assays using an anti-oligopeptide
antibody (13) produced slower-mobility complexesthat
mi-grated as distinct bands in the gel (26). As an additional verification ofthe Rep78-hairpin complex, addition of
ex-cess,unlabeledAAVhairpinDNAabolishedbinding(26).
Insomereactions, backgroundbands with fastermobility than the Rep-hairpin complex wereobserved inboth pCD Mrep- and pCDM8-transfected cell nuclear extracts (Fig.
C.
40
;I
W-171-..
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.130.491.71.393.2]x- u. E: r. r x c X
a- Xd v *t *E- *m *t
rz
N - X X r. x m x it_ I- Un m
V" e._ * a r)
I4
1
*i "w wZ "-F_[vi4-B-"
- - - N
o o& XI I%
tz A
-_- B_-i
t
;
x. '. t.
-oi
t
-4
t .,t.I&
--*-F-go-N N U)
'k
nN
'9t-0
t
Idmw
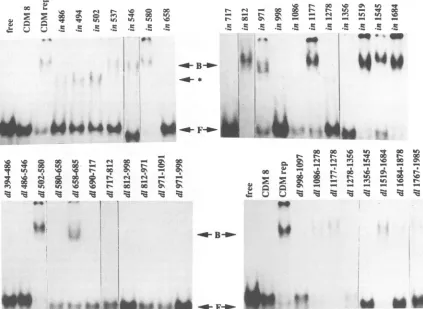
FIG. 4. Bindingof mutant AAVRepproteinsto AAVterminal repeats. Equivalent amounts of mutant Rep proteins were used in the gel retardation assay with linearizedpCDM8 and pGEMSV plasmids in the binding reaction. *, nonspecific background band; B, bound hairpin;
F, freehairpin.
3C, lanes 1 and 6, respectively). These background bands werereduced, however, when the amount of extract in the
binding reactionwas reduced (Fig. 3C, lanes 1 to 4). Anti-bodies against the Rep protein failed to supershift the
faster-mobility complex (26).To eliminate these bands, we used several other types ofcompetitorDNAsinthebinding reaction. Addition of 0.5 ,ug of linearized pCDM8 to the
bindingreactions eliminated thebackgroundbandproduced
by nuclear extracts from both pCDM8- and pCDMrep-transfected COS-1 cells(Fig. 3C,lanes 7 and8).Addition of 0.5 ,g oflinearizedpGEM3ZtopCDM8-transfected (lane 9)
andpCDMrep-transfected (lane 10) COS-1 cell nuclear ex-tractsdidnotreduce theamountof thebackgroundband. A third plasmid, pGEMSV, which contains the entire SV40 genomeinserted into theBamHI site ofpGEM3Z(26), was also capable of reducing the background band in binding reactions with extractsfrompCDM8-transfected cells (Fig.
3B; compare lanes 12 and 13 with lane 14). When both
pGEMSVandpCDM8wereaddedtothebinding reactions,
thebackgroundband waseliminatedentirely. Therefore,we usedthis combination ofcompetitorDNAsin ourassaysto eliminate thebackground bands.
Binding ofmutantRep proteinsto the AAV terminalrepeat sequences. Becauseof the differentialproductionofsomeof themutant Repproteins,weuseddensitometric analysis of Western blots(Fig. 2)toestimate theamountof Repprotein in the extracts. This enabled us to use nearly equivalent amounts of Rep protein in the binding assays. However,
when higher concentrations of extract were used, the amount ofbackground binding increased, but this did not prevent us from making a definitive determination of the binding abilities of the mutant Rep proteins.
In a previous report, we demonstrated that five of our
XAhoI linker insertion mutants, pin685, pin690, pinll77, pin1878, and pin1884, and one of our deletion mutants,
pd1658-685,
supported AAV DNA replication in vivo (25).Since Rep78 or Rep68 binding to the terminal repeat se-quences is believed to be essential for AAV DNA
replica-tion,weassumed that thesemutant Repproteinswould also bind to thehairpin. Two such replication-positive mutants,
pinll77 andpd1658-685, were tested in thegel shift assays andwere foundtobindtothe hairpin efficiently (Fig. 4).
Arepresentative sampling ofgel shift assays is shown in Fig. 4, and Table1summarizes theDNA-binding abilities of the mutant Rep proteins from three or more independent
assays.Most oftheinsertionmutants producedproteinthat wascapable ofbindingtothehairpin.Mostof thelesions in the Rep78 protein that abolished bindingare located in the amino-terminal half of the protein. Onlytwo mutants with deletions in the amino terminus of the protein,
pd1502-580
and
pd1658-685,
arepositive forbindingtotheAAVhairpin(Fig. 4). Mutants with insertions in those two regions,
pin537, pin546, pin580, pin685, andpin690,caneitherbindto
the AAV hairpin or support AAV DNA replication (25).
Thus,twoamino-terminalregionsin theRep78protein,from
a No a Qo va
$ v tUm in
,eS U) %sv
00 a
w
to
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.104.527.76.385.2]TABLE 1. DNAbinding of AAV rep gene mutants'
Mutant Codons affected AAV DNA Hairpin
replication binding
in486 56-57 -
-in494 58-59 -
-in502 61-62 -
-in537 73-74 - +
in546 76-77 - +
in580 87-88 - +
in658 113-114 -
-in685b 122-123 + ND
in690b 124-125 + ND
in 717 133-134 -
-in 812 164-165 - +
in971 217-218 - +
in998 226-227 -
-in1086 256-257 -
-in 1097C 259-260 - ND
in 1177 286-287 + +
in 1278 320-321 -
-in 1356 346-347 -
-in 1519 400-401 - +
in 1545 409-410 - +
in 1684 455-456 - +
in 1878b 520-521 + ND
in 1884b 522-523 + ND
dl394-486 25-56 -
-dl 486-546 57-76 -
-dl502-580 62-87 - +
dl580-658 88-113 -
-dl 658-685 114-122 + +
dl690-717 125-133 -
-dl717-812 134-164 -
-dl 812-971 165-217 -
-dl812-998 165-226 -
-dl971-998 218-226 -
-dl971-1097 218-259 -
-dl998-1097 227-259 -
-dl1086-1278 257-320 - +
dl1177-1278 287-320 - +
dl1278-1356 321-346 - +
dl1356-1545 347-409 -
-dl 1519-1684 401-455 - +
dl1684-1878d 456-520 - +
dl1767-1985d 482-556 - +
a Aplus-or-minus sign indicatesthat the mutantproteinbound to the AAV
terminal repeatswithweakaffinity. ND,notdetermined.These
determina-tionsarebaseduponthreeor moreindependentplasmidtransfection
exper-imentsandgelshiftassays.
bThesemutantssupportedAAV DNAreplication.
cNoDNA-bindingassay was done with this mutantbecausenoRepprotein
wasdetectedinWestern blot assays.
dLocatedmainlyincytoplasm.
codons 62 to 87 and 114 to 124, are dispensable for DNA binding.
Inthecentralportion of the Rep78protein, three mutants,
pd11086-1278, pd11177-1278,
andpd11278-1356,
producedRep78 proteins capable of weak binding to DNA. This suggests that thisregion in Rep78 stabilizes the protein-DNA complex. Other published findings indicate the importance ofthis region for hairpin binding because small deletions in this region abolished binding (12). Our ability to detect limited hairpin binding with these mutants may be due to a higher level of Rep78 protein in our crude nuclear extracts than in the invitro translation products used by McCarty et al. (12).
Mutations in the carboxyl terminus of the Rep protein
from amino acid residues 401to 621may haveonlyaslight
effect onbinding. Deletions in theseregions do not abolish binding but alter the Repprotein'saffinity for the viralorigin
ofreplication.Thetwoputative nuclear localizationmutants pd11684-1878 and pd11767-1985 can also bind to DNA, al-though with weak affinity. One of the mutants that binds
tightlytothehairpin, pinlS19,trans activates the AAVp40
promoter butdoesnotsupport AAV DNAreplication(25).
ThebindingofpinlS19toAAVterminal repeats supports the model in which DNA binding may be necessary for Rep
protein to trans activate the p40 promoter.
Themajor functions of Rep78 in the AAV life cycleare to replicateviral DNAandtransactivate thep40transcription
promoter. The findings presented here indicate that these functionsrequire Repproteins that are capable of bindingto the viralorigin of replication. An additional function of the AAV Rep78 protein is suppression of oncogene-mediated cellular transformation (5, 25). It is interesting that the DNA-binding capacity of the Rep protein (as measured in these AAVhairpingel shift assays) is apparently not essen-tial for this function. Thefollowing mutants failtosuppress oncogene-mediatedtransformation in mouse fibroblast cells orprimary BRK cells but demonstrate measurable bindingto the AAV hairpin: pin537, pin812, pd11086-1278, pdlll77-1278, pd11278-1356, pd11684-1878, and pd11767-1985. This suggeststhat DNA binding is not required for transformation suppression. Alternatively, the domains in the Rep78 protein that are essential forbindingtothe AAVhairpin are different from those regions that mediate nonspecific DNA binding (9), and these other regions may be responsible for suppres-sion of cellular transformation.
Whenweincluded SV40origin-containing plasmids inour gel mobility shift assays, background binding to the AAV hairpin was reduced dramatically (Fig. 3), whereas other nonspecific plasmid DNA did not alter the background. It has been reported that the AAV terminal repeat sequence, when cloned into a plasmid, serves as areplicationorigin in carcinogen-treated CHO cells in the absence of AAV-en-coded genes (24). Therefore, the AAVorigin ofreplication is recognized by the cellular replication machinery. It has also been reported thatunidentified cellular proteinscanbindto sequences in the AAV terminal repeats and that they show comparable affinity to the SV40 origin core sequence (22). Thus, when another origin of replication that is recognized by similarcellular factors is included in thegel shift assays, it effectively competes with the AAV origin for binding to the cellularcomponents.
This work expands the analysis of ourcollection of rep gene mutants by defining four regions thatarenecessary for binding to the AAV terminal repeat sequences: amino acid residues 25 to 62, 88 to 113, 125 to 256, and 346 to 400. Although removal of amino acids 125 to 256 abolished binding, insertions between amino acids164and 165(pin812) and 217 and 218 (pin971) did not disrupt binding. It should also be noted that aninsertion between amino acids 400 and 401 (pinl519) did not affectbinding, even though adeletion betweenresidues 347 and 409(pd11356-1545)abolished bind-ing. Because of the large sizes of someof thesedeletions, the conformation of these mutant proteins is probably drasti-cally affected, which in turn abolishes hairpin binding. However, when these regions were disruptedvia twoamino acid insertions (as in pin812, pin971, and pinl519), mutant Rep78 was still capable of forming a complex with the hairpin. Therefore, a definitive appraisal of how these do-mainsparticipate in hairpin binding awaits further investiga-tions. Relatively large regions of AAV hairpin DNA are
on November 9, 2019 by guest
http://jvi.asm.org/
protected by Rep protein binding (1, 7), and at least three different protein-DNA complexes are present in gel shift assays (1, 7, 9, 14; this report), suggesting that the larger Rep proteins form oligomers or interact with cellular proteins during interaction with the AAV termini. Identification of cellular proteins that interact withthe AAVorigin of repli-cation will lendinsight into howRep78 performsits numer-ousfunctions.
Wethank R. A. Owens forprovidinguswithunpublished data,R. J. Samulski for thepsub201 plasmid, RuchikaGuptafor excellent technicalassistance, andJian Zhu forhelpinpreparing thefigures for themanuscript.
The work performed in this study was supported by the Ohio Cancer ResearchAssociation, Columbus, Ohio,and The March of Dimes Birth Defects Foundation(1-FY92-0527),WhitePlains, N.Y.
REFERENCES
1. Ashktorab, H., and A. Srivastava. 1989.Identification of nuclear proteins that specifically interact with the adeno-associated virus 2 inverted terminal repeathairpin DNA. J. Virol. 63:3034-3039.
2. Berns, K. I., and R. A. Bohenzky. 1987. Adeno-associated viruses: an update. Adv. Virus Res. 32:243-306.
3. Chejanovsky, N., and B. J. Carter. 1989. Mutagenesis of an AUG codon in the adeno-associated virus rep gene: effectson viral DNAreplication. Virology 173:120-128.
4. Hauswirth, W. W., and K. I. Berns. 1977.Origin and termina-tion of adeno-associated virus DNA replication. Virology 78: 488-499.
5. Hermonat, P. L.1991. The adeno-associated virus Rep78gene inhibits cellulartransformationby bovinepapillomavirus. Virol-ogy172:253-261.
6. Hermonat, P. L., M. A. Labow, R.Wright, and K. I. Berns. 1984.Genetics ofadeno-associatedvirus: isolation and prelim-inary characterization ofadeno-associatedvirustype 2 mutants. J.Virol. 51:329-339.
7. Im,D.-S., and N. Muzyczka. 1989. Factors that bind to adeno-associated virus terminal repeats. J. Virol. 63:3095-3104. 8. Im, D.-S., and N. Muzyczka. 1990. The AAV origin binding
protein Rep68 is an ATP-dependent site-specificendonuclease with DNA helicaseactivity. Cell 61:447-457.
9. Im,D.-S., and N. Muzyczka. 1992. Partialpurification of adeno-associatedvirusRep78,Rep52, andRep4Oand their biochemi-cal characterization. J. Virol. 66:1119-1128.
10. Labow, M. A., P. L. Hermonat, and K. I. Berns. 1986.Positive and negative autoregulation of adeno-associated virus type 2 genome.J. Virol. 60:251-258.
11. McCarty, D. M., M. Christensen, and N. Muzyczka. 1991. Sequences required for coordinate induction of adeno-associ-ated virus p19 and p40 promoters by Rep protein. J. Virol. 65:2936-2945.
12. McCarty, D. M., T.-H. Ni, and N. Muzyczka. 1992.Analysis of mutations inadeno-associated virus Rep protein in vivo and in vitro.J. Virol. 66:4050-4057.
13. Mendelson, E., J. P. Trempe, and B. J. Carter. 1986. Identifi-cation of thetrans-acting Rep proteins of adeno-associated virus byantibodies to asyntheticoligopeptide.J.Virol. 60:823-832. 14. Owens, R. A., J. P.Trempe, N.Chejanovsky,and B.J. Carter. 1991. Adeno-associated virus Repproteins produced in insect andmammalianexpression systems: wild-type and dominant-negative mutant proteins bind to the viral replication origin. Virology 184:14-22.
15. Samulski, R. J., L.-S. Chang, and T. Shenk 1987. A recombi-nantplasmid from which aninfectious adeno-associated virus genome can be excised in vitro and its use to study viral replication. J. Virol. 61:3096-3101.
16. Schlehofer, J. R., M.Ehrbar, and H. ZurHausen. 1986.Vaccinia virus, herpes simplex virus, and carcinogens induce DNA amplification in ahuman cell line andsupportreplication ofa helper virus dependentparvovirus. Virology152:110-117. 17. Snyder, R. O., R. J.Samulski, and N. Muzyczka. 1990. In vitro
resolution ofcovalentlyjointed AAV chromosome ends. Cell 60:105-133.
18. Tratschin, J. D., I. L.Miller, and B. J. Carter. 1984. Genetic analysis of adeno-associated virus: properties of deletion mu-tantsconstructed in vitro and evidence foranadeno-associated virusreplication function.J. Virol.51:611-619.
19. Tratschin, J. D., J. Tal, and B. J. Carter. 1986. Negativeand positive regulation in trans of gene expression from adeno-associated virus vectors in mammalian cells. Mol. Cell. Biol. 6:2884-2894.
20. Tratschin, J. D., M. H. P. West, T. Sandbank, and B. J. Carter. 1984. Ahumanparvovirus, adeno-associated virus,as a eukary-otic vector: transient expression andencapsidationof the pro-caryoticgeneforchloramphenicol acetyltransferase.Mol.Cell. Biol. 4:2072-2081.
21. Trempe, J. P., E. Mendelson, and B. J. Carter. 1987. Charac-terization ofadeno-associated virusRep proteins in human cells by antibodies raised againstRep expressedinEscherichiacoli. Virology 161:18-28.
22. Walz, C., and J. R. Schlehofer. 1992. Modification of some biologicalproperties of Hela cellscontaining adeno-associated virus DNA integrated into chromosome 17. J. Virol. 66:2990-3002.
23. Yacobson, B., T. Koch, and E. Winocour. 1987.Replicationof adeno-associated virus insynchronized cells without the addi-tionofahelpervirus. J. Virol. 61:972-981.
24. Yalkinoglu, A.O.,J.Zentgraf, and U.Hubscher.1991.Origin of adeno-associated virus DNAreplication is atargetof carcino-gen-inducibleDNAamplification.J. Virol. 65:3175-3184. 25. Yang, Q., A. Kadam, and J. P. Trempe. 1992. Mutational
analysis ofthe adeno-associated virus rep gene. J. Virol. 66: 6058-6069.
26. Yang, Q., and J. P. Trempe.Unpublisheddata.