Copyright © 1998, American Society for Microbiology
Identification of a Domain within the Human T-Cell Leukemia
Virus Type 2 Envelope Required for Syncytium Induction
and Replication
BETTY POON
1ANDIRVIN S. Y. CHEN
1,2*
Department of Microbiology and Immunology
1and Division of Hematology-Oncology,
2UCLA School of Medicine, Los Angeles, California 90095
Received 12 May 1997/Accepted 13 November 1997
In vitro infection by human T-cell leukemia virus type 1 and 2 (HTLV-1 and HTLV-2) can result in syncytium
formation, facilitating viral entry. Using cell lines that were susceptible to HTLV-2-mediated syncytium
formation but were nonfusogenic with HTLV-1, we constructed chimeric envelopes between HTLV-1 and -2 and
assayed for the ability to induce syncytia in BJAB cells and HeLa cells. We have identified a fusion domain
composed of the first 64 amino acids at the amino terminus of the HTLV-2 transmembrane protein, p21, the
retention of which was required for syncytium induction. Construction of replication-competent HTLV genomic
clones allowed us to correlate the ability of HTLV-2 to induce syncytia with the ability to replicate in BJAB
cells. Differences in the ability to induce syncytia were not due to differences in the levels of total or cell
membrane-associated envelope or in the formation of multimers. Therefore, we have localized a fusion domain
within the amino terminus of the transmembrane protein of HTLV-2 envelope that is necessary for syncytium
induction and viral replication.
Human T-cell leukemia virus types 1 and 2 (HTLV-1 and
HTLV-2) are type C retroviruses that have been associated
with a variety of human malignancies. HTLV-1 is the
etiolog-ical agent of adult T-cell leukemia as well as a degenerative
neurological disorder, HTLV-1-associated myelopathy/tropical
spastic paraparesis (28, 40, 58, 60, 83). Recent reports have
also implicated HTLV-1 infection with arthropathy (42, 65),
polymyosis (23, 37), and uveitis (48, 49, 51). HTLV-2 has been
associated with a rare form of atypical hairy cell leukemia (62,
63, 68) as well as some cases of neuropathy (33, 39). It is
estimated that between 10 million and 20 million individuals
worldwide are infected with HTLV, with an overall risk of 5%
of disease progression in infected individuals (14). HTLV is
endemic in southern Japan, the Caribbean Basin, and Central
and South America. In the United States, recent reports have
identified a high proportion of HTLV, especially HTLV-2,
infection in intravenous-drug abusers (44, 61, 64).
Cell-to-cell contact is considered critical for the in vivo and
in vitro transmission of HTLV-1 and HTLV-2, as infection by
cell-free HTLV virus is inefficient in vitro and in vivo. By
analogy with other enveloped viruses, HTLV infection of
sus-ceptible cells is likely mediated by the envelope glycoprotein.
Antibodies against HTLV envelope are protective against
in-fection in vivo (71, 80), and multiple epitopes that elicit
neu-tralizing antibodies have been identified throughout the
pro-tein (31, 34, 56). Initially synthesized as a precursor propro-tein,
gp61, HTLV envelope is subsequently modified by
glycosyla-tion and cleaved into two subunits, gp46 and p21. The external
surface glycoprotein, gp46, is anchored to the cell surface by
noncovalent association with the transmembrane envelope
gly-coprotein, p21. Interaction of envelope with the as yet
uniden-tified cellular receptor leads to cell-to-cell fusion and can result
in syncytium formation.
We were interested in identifying the molecular
determi-nants of HTLV involved in syncytium formation and viral
entry. Our laboratory has several cell lines that are permissive
to HTLV-2- but not HTLV-1-mediated cell fusion. Therefore,
we constructed recombinants between the HTLV-1 and -2
envelope genes and assayed for the loss of syncytium induction
in BJAB cells and HeLa cells. Loss of a 64-amino-acid (aa)
domain located at the amino terminus of the HTLV-2
trans-membrane protein, p21, correlated with a loss in the ability of
the envelope chimera to induce cell fusion. When the chimeric
envelopes were expressed in the context of
replication-compe-tent genomic clones, there was a good correlation between
syncytium induction and the ability to replicate in permissive
cells. Present within the identified fusion domain is a
hydro-phobic region and a heptad repeat resembling a leucine zipper.
We examined the contribution of the fusion domain to the
structural integrity of the HTLV-2 envelope by using a vaccinia
virus expression system. None of the recombinants affected the
synthesis, transport, or oligomer formation of the HTLV
gly-coprotein complex.
MATERIALS AND METHODS
Cells.BJAB cells were grown in RPMI 1640 medium with 10% fetal bovine serum (FBS) (Gemini, Calabasas, Calif.). HeLa cells were grown in Dulbecco modified Eagle medium containing 10% calf serum. 729ph6neo and SLB1, which express infectious HTLV-2 and HTLV-1, respectively, were grown in Iscove’s medium with 20% FBS.
Oligonucleotide-directed mutagenesis and DNA manipulation.The HTLV-2 envelope sequences from nucleotide (nt) 5123 to 7392 (72) from BCHTLV (30) and the HTLV-1 envelope sequences from nt 5126 to 7480 (69) from Env1A (kindly provided by D. Slamon, University of California, Los Angeles) were subcloned between the SphI and MluI restriction sites of CDM7 (kindly provided by D. Camerini, University of Virginia), replacing a 2.6-kb fragment and creating CDM7-II and CDM7-I, respectively. Following preparation of single-stranded DNA using M13 K07 phage, mutagenesis was performed according to the pro-tocol for the T7-GEN in vitro mutagenesis kit (United States Biochemical). The nucleotide positions of the HTLV-1 sequences in clones NH, NK, KH, KM, MH, and NKM are indicated in parentheses. Clones NH (nt 5179 to 6680), NK (nt 5179 to 6118), KH (nt 6118 to 6680), KM (nt 6118 to 6290), MH (nt 6290 to 6680), and NKM (nt 5179 to 6118 and 6290 to 6680) were constructed by
* Corresponding author. Mailing address: 11-934 Factor, Division of
Hematology-Oncology, Department of Medicine, UCLA School of
Medicine, Los Angeles, CA 90095-1678. Phone: (310) 825-4793. Fax:
(310) 794-7682. E-mail: [email protected].
1959
on November 9, 2019 by guest
http://jvi.asm.org/
Downloaded from
on November 9, 2019 by guest
http://jvi.asm.org/
Downloaded from
on November 9, 2019 by guest
http://jvi.asm.org/
introducing compatible restriction sites within HTLV-1 and -2 envelope. A novel
HindIII site was introduced at the ends of both genes. KpnI and MstI sites,
already present within the HTLV-1 envelope, were introduced into the same locations within the HTLV-2 envelope. All clones were confirmed by DNA sequencing. Oligonucleotides containing base pair substitutions that created the unique restriction sites (underlined) were as follows: for HTLV-2, 59CTG CTA TTG GTA CCG CAC GGC GGC G 39(KpnI site 6105), 59GCT GCA AAG CTT GCA GGT CTA 39(HindIII site 6650), and 59CGT CTA TTT TGC GCA GCA TAC TGT GC 39(MstI site 6292); for HTLV-1, 59GGC TGG GGA AGC TTG AGG CGA TGA G 39(HindIII site 6682).
Constructs BM and KB were made by using overlapping oligonucleotides that consisted of nt 6118 to 6200 (sense) and 6195 to 6310 (antisense) of HTLV-2 and HTLV-1 envelope and nt 6116 to 6195 (sense) and 6184 to 6300 (antisense) of HTLV-1 and HTLV-2 envelope, respectively. The oligonucleotides were an-nealed by heating in extension buffer (40 mM Tris [pH 7.4], 20 mM MgCl2, 50
mM NaCl) at 65°C for 5 min and allowing the reaction to cool to 25°C. Extension occurred at 25°C for 10 min by the addition of 1ml Sequenase (United States Biochemical). The resulting 200-bp fragment, containing a unique BamHI re-striction site at nt 6200, was subsequently cloned into CDM7-II between the newly created KpnI and MstI restriction sites.
The genomic clones were obtained by inserting the chimeric envelopes from CDM7 between the SphI and MluI restriction sites of H6H11. H6H11 contains the HTLV-2 genomic sequences fromlH6 (8) inserted between the HindIII site of pBR322.
The vaccinia virus-driven expression vectors containing the HTLV envelopes were obtained by PCR amplification of the envelope sequences from CDM7 into the pTM-3 expression vector (kindly provided by B. Moss, National Institute of Allergy and Infectious Diseases). The HTLV-2 envelope was amplified by using forward primer (59GCG GAA TTC TTT TCT TCC TAC TTT TAT TC 39) and reverse primer (59GAA TCG AGT TAG GGC TGG 39). The HTLV-1 envelope was amplified by using forward primer (59GGC GAA TTC TTC TCG CCA CTT TGA TTT 39) and reverse primer (59CGC AGA TCT TAT CGG CGG GAG CGG GAT CC 39). The resulting 1.7-kb PCR fragments were digested with
EcoRI and PstI and inserted downstream of the bacteriophage T7 promoter of
pTM-3.
p24 assay.Supernatants from transfected BJAB cells were collected and clar-ified by centrifugation at 3,000 rpm for 5 min. Analysis for the presence of HTLV p24 antigen was performed by enzyme-linked immunosorbent assay (ELISA), using Coulter kit 6604252 according to the manufacturer’s procedure.
Protein analysis.Cells were lysed on ice for 10 min in OGL lysis buffer (100 mM Tris [pH 8.0], 100 mM NaCl, 1 mM CaCl2, 250 mM octyl-glucosidase). For
detection of denatured proteins, samples were heated at 100°C for 5 min in 13
loading buffer (100 mM Tris [pH 6.8], 20% glycerol, 0.02% bromophenol blue) with the addition of 4% sodium dodecyl sulfate (SDS) and 5%b -mercaptoetha-nol, and separated on SDS–10% polyacrylamide gels. For detection of native proteins, samples were diluted in 13loading buffer and separated on 4 to 20% gradient acrylamide gels in the presence of 0.01% SDS in the running buffer (79). Western analysis was performed with anti-HTLV-1 gp46 antibody (clone 65/ 6c2.2.34; Cellular Products Inc., Buffalo, N.Y.) and developed by using the Amersham enhanced chemiluminescence assay.
Transfections and blue syncytium assay.Transient transfection assays were performed by resuspending 5 3106 BJAB cells in electroporation medium
(RPMI 1640 with 20% FBS) and electroporating with 15mg of DNA at 230 V and 960mF.
The blue syncytium assay was performed as previously described (45), with the following modifications. HeLa cells were infected with wild-type vaccinia virus (WR) or VTF7-3 (multiplicity of infection [MOI]51.0) for 2 h at 37°C. Cells were subsequently transfected with 15ml of Lipofectin (Gibco BRL) and 10mg of DNA/105cells in serum-free Dulbecco modified Eagle medium as specified by
the manufacturer. Cells infected with VTF7-3 were transfected with vaccinia virus-driven HTLV envelope constructs. Cells infected with WR were trans-fected with pEM-ClacZbgAn (45). At 16 h posttransfection, cells expressing the
HTLV envelope clones were harvested by treatment with phosphate-buffered saline-EDTA and washed twice with medium. Then 105cells from each of the
two sources, WR infected and VTF7-3 infected, were mixed in the presence of actinomycin D (1mg/ml) to prevent vaccinia virus spread. After a 7-h incubation at 37°C, cultures were fixed and stained in situ for 1 h at 37°C with solution containing 4 mM potassium ferrocyanide, 2 mM MgCl2, and 0.5 mg of
5-bromo-4-chloro-3-indolyl-b-D-galactopyranoside (X-Gal) per ml. Giant blue syncytia
were counted microscopically.
Flow cytometry.Cells were stained with 1mg of anti-HTLV-1 gp46 antibody clone (65/6c2.2.34; Cellular Products) diluted in 100ml of FACS (fluorescence-activated cell sorting) buffer (phosphate-buffered saline with 2% FBS and 0.01% sodium azide) at 4°C for 20 min. After one wash with FACS buffer, cells were incubated at 4°C, with fluorescein isothiocyanate-conjugated sheep anti-mouse antibody (diluted 1/200; Cappel) for an additional 20 min. Cells were resus-pended in FACS buffer, and data were acquired on a FACScan (Becton Dick-inson) and analyzed with the Lysis 2 software program.
RESULTS
The amino terminus of the HTLV-2 transmembrane
pro-tein contains sequences required for syncytium formation in
BJAB cells.
HTLV-1 and -2 can induce syncytium formation
upon infection of a variety of cell types, including T- and
non-T-cell lines (35, 47, 80). We had previously observed that
an Epstein-Barr virus-negative human B-cell line (32), BJAB,
formed numerous multinucleated cells upon cocultivation
with 729ph6neo, an HTLV-2-producing cell line. In contrast,
cocultivation with HTLV-1-producing cells, such as SLB1 cells,
did not result in visible syncytia. We took advantage of the
differential ability of HTLV-1 and -2 to form syncytia in BJAB
cells to define the regions of the HTLV-2 genome involved in
syncytium formation.
Previous experiments have mapped the fusion phenotype to
the HTLV envelope protein, as expression of envelope alone
was sufficient to induce syncytia (18, 45, 57). HTLV-1 and -2
show 75% amino acid homology between the envelope
pro-teins, allowing construction of recombinants which should
maintain the functionality of the protein. The chimeric
enve-lope genes were subsequently substituted into H6H11, an
in-fectious HTLV-2 genomic clone (Fig. 1). These full-length
clones containing HTLV-1 envelope sequences allowed us to
study not only the syncytium-inducing phenotype but also the
contributions of the envelope domains in viral replication. The
genomic clones were transfected into BJAB cells and, at 3 days
posttransfection, analyzed microscopically for syncytium
for-mation. At this time, the ability of the clones to cause fusion is
due to the transient expression of viral proteins from the
trans-FIG. 1. The amino terminus of HTLV-2 p21 is necessary but not sufficient for syncytium formation in BJAB cells. BJAB cells (53105) were transfected withgenomic constructs containing chimeric envelopes (shown) as described in Ma-terials and Methods. At day 3 posttransfection, cells were analyzed for syncytia by microscopic analysis.1and2indicate that greater than or less than 10% of the cell population were undergoing cell fusion, respectively. HTLV-2 sequences are depicted by open boxes; HTLV-1 sequences are depicted by dark boxes.
on November 9, 2019 by guest
http://jvi.asm.org/
fected DNA. Syncytia were defined as giant cells greater in
diameter than three single cells.
Transfection with the genomic HTLV-2 clone, H6H11,
pro-duced syncytia in 10 to 30% of the cell population. In contrast,
a genomic clone in which the entire HTLV-2 envelope had
been substituted by the HTLV-1 envelope (NH) did not cause
any visible cell fusion. These results are in agreement with our
previous observation that HTLV-2- but not
HTLV-1-produc-ing cells can induce syncytia in BJAB cells. These results also
confirmed the role of the envelope gene in syncytium
forma-tion.
The HTLV envelope is composed of the external surface
glycoprotein, gp46, and the transmembrane anchoring protein,
p21. We studied the contributions of these domains in
syncy-tium induction by substituting these individual domains from
HTLV-1 into the HTLV-2 envelope. A clone (NK) that had
the gp46 domain substituted by HTLV-1 envelope sequences
but retained the p21 transmembrane domain of HTLV-2 was
capable of inducing syncytia in BJAB cells. The reciprocal
clone (KH), containing a replacement of the p21 domain with
HTLV-1 sequences, lost the fusogenic phenotype. These
re-sults appear to map the syncytium induction phenotype in
BJAB cells to the HTLV-2 p21 transmembrane protein.
Additional chimeras were constructed to further define the
regions within p21 necessary for cell fusion. Replacement of
the carboxyl portion of HTLV-2 p21 with HTLV-1 sequences
(MH) did not affect syncytium formation. However,
substitu-tion of the amino terminus of HTLV-2 p21 with HTLV-1
sequences (KM) resulted in loss of cell fusion. Therefore, this
64-aa region within the HTLV-2 p21 transmembrane protein
appeared to be necessary for syncytium induction.
We next substituted either the amino or carboxyl portion of
these 64 aa from HTLV-2 p21 with HTLV-1 envelope
se-quences. These smaller substitutions within the amino
termi-nus of HTLV-2 p21 revealed that a region from HTLV-2
envelope comprised of aa 330 to 365 was required for fusion.
Substitution of these sequences with the corresponding
HTLV-1 sequences (BM) correlated with a loss of syncytium
induction. This conclusion is further supported by the
reten-tion of these sequences from HTLV-2 p21 in all clones that
were able to cause syncytia in BJAB cells (NK, MH, and KB).
Therefore, we have defined a region essential for HTLV-2
fusion in BJAB cells to a 34-aa domain located at the amino
terminus of the HTLV-2 envelope transmembrane protein.
Loss of this 34-aa domain from HTLV-2 envelope negatively
affected syncytium induction, as seen by the inability to fuse
BJAB cells. We next tested whether the sequences from the
amino terminus of HTLV-2 p21 were sufficient for syncytium
induction in the context of a recombinant where all other
envelope sequences were derived from HTLV-1. A clone
con-taining the first 64 aa of HTLV-2 p21 in the context of HTLV-1
envelope (NKM) did not induce syncytia in BJAB cells. A
substitution of the smaller 34-aa domain from HTLV-2 also
did not result in syncytium induction by the HTLV-1 envelope
(data not shown). Therefore, the fusion domain identified in
HTLV-2 was not sufficient on its own to confer the syncytium
induction phenotype upon the HTLV-1 envelope.
The amino terminus of HTLV-2 p21 is required for HTLV-2
replication in BJAB cells.
Cell fusion is believed to be the
major route of HTLV viral spread, as cell-free transmission is
highly inefficient both in vivo and in vitro (6, 46, 52, 59).
Therefore, we examined the ability of the recombinants to
replicate in BJAB cells and correlated it with their ability to
cause syncytia. The replicative potential of the chimeras in
BJAB cells was determined by ELISA for the production and
secretion of viral core antigen, p24, over several weeks (Fig. 2).
All of the clones were p24 positive on day 3, indicating that all
of the chimeras were capable of viral protein production from
the transfected DNA (data not shown). As expected, the
genomic HTLV-2 clone, H6H11, replicated to high levels over
this time period, as shown by an increase in the production of
p24 in the supernatant. The increased p24 production
corre-lated with increased cell fusion with syncytia in over 80% of the
FIG. 2. HTLV-2 replication in BJAB cells requires sequences at the amino terminus of p21. BJAB cells (53105) were transfected with genomic constructscontaining chimeric envelopes as described in Materials and Methods. Every 3 days, the medium was changed by allowing the cells to settle and replacing half of the medium. At 3, 7, 14, and 21 days posttransfection, supernatants were collected and cell debris was removed by centrifugation at 3,000 rpm for 5 min as described in Materials and Methods. The amount of secreted HTLV p24 was quantitated by ELISA as described in Materials and Methods. (A) Clones that induced syncytia in BJAB cells (Fig. 1) which included full-length HTLV-2 envelope (II), as well as clones NK, MH, and KB. (B) Clones that did not induce syncytia in BJAB cells (KH, KM, BM, and NKM). (C) Results from the full-length HTLV-2 envelope and the full-full-length HTLV-1 envelope (NH). The data are representative of three independent experiments. OD550, optical density at 550 nm.
on November 9, 2019 by guest
http://jvi.asm.org/
cells during this time period. All clones that were capable of
causing cell fusion (NK, MH, and KB) were also able to
rep-licate in BJAB cells, producing p24 at levels similar to those of
H6H11 (Fig. 2A). Similarly, clones that did not induce syncytia
(KH, KM, BM, and NKM) did not replicate, producing an
initial burst of p24 due to the transfected DNA that was not
sustained over time (Fig. 2B). These results support the
premise that HTLV-2 spread is dependent on the cell fusion
activity, as only the clones capable of syncytium induction were
also able to replicate in BJAB cells. As the
replication-com-petent clones (NK, MH, and KB) also retained the amino
terminus of the HTLV-2 p21 domain, we conclude that the
same 34-aa sequences within the transmembrane protein p21
confer both the syncytium induction phenotype and the
repli-cative capability in BJAB cells.
Interestingly, the clone containing the entire HTLV-1
enve-lope sequences (NH) was able to replicate to the same degree
as the wild-type HTLV-2 (Fig. 2C), albeit at a slightly lower
rate and without detectable cell fusion. Cocultivation of BJAB
cells with lethally irradiated SLB1 cells, the original source of
the HTLV-1 envelope clone, did not result in productively
infected BJAB cells, as evidenced by the eventual decline in
p24 production over time (data not shown). Therefore, there
may be interactions occurring between the HTLV-1 envelope
and other HTLV-2 viral components that allow replication in
BJAB cells.
Fusion of HeLa cells by HTLV-2 envelope alone requires the
amino terminus of p21.
The ability of the construct containing
the HTLV-1 envelope to replicate in BJAB cells suggested that
interactions of envelope with other viral proteins may be
oc-curring. Therefore, we investigated the contributions to
syncy-tium formation of the HTLV envelope protein alone in the
absence of other viral components. We next constructed
vac-cinia virus/T7 promoter-driven vectors that expressed only the
HTLV chimeric envelopes. HTLV envelope expressed by the
vaccinia virus/T7 polymerase expression system has been
re-ported to be produced in a properly processed and folded form
(3).
We also used a modification of a sensitive blue syncytium
assay to quantitate the amount of cell fusion caused by the
envelope chimeras (45). Briefly, cells were transfected with a
construct containing the
b-galactosidase gene under the
con-trol of the T7 RNA polymerase promoter. A second set of cells
were transfected with the HTLV envelope construct, as well as
infected with vaccinia virus expressing the T7 RNA polymerase
(VTF7-3). Upon mixing of the two populations, cell fusion
would allow the T7 RNA polymerase expressed from VTF7-3
to activate the
b-galactosidase gene, resulting in a blue syncytia.
In these experiments, HeLa cells were used, as they
pro-duced the highest level of expression of HTLV envelope (data
not shown). In addition, syncytia were easier to score in HeLa
cells due to the adherent nature of the cells. HeLa cells, similar
to BJAB cells, also showed a differential ability to be fused by
HTLV-1 and -2 envelope (Fig. 3). HTLV-2 envelope gave rise
to 780 blue syncytia, compared to the 9 syncytia produced by
the HTLV-1 envelope (NH). Clones that were capable of
in-ducing syncytia in BJAB cells (NK, MH, and KB) also resulted
in significant amounts of blue syncytia. The majority of the
clones that were unable to cause fusion in BJAB cells (KH,
KM, and NKM) were also unable to cause fusion in HeLa cells.
In general, the blue syncytium assay in HeLa cells, using
vac-cinia virus-expressed envelope, appeared to confirm the fusion
results of the genomic clones in BJAB cells, with one exception.
Although loss of the first 64 aa at the amino terminus of
HTLV-2 p21 (KM) resulted in loss of syncytia in both HeLa
cells and BJAB cells, cell fusion could be induced in HeLa cells
by clones that retained either the amino or carboxyl portion of
this region (BM or KB, respectively). In contrast, in BJAB
cells, clone KB was fusogenic whereas clone BM did not form
visible syncytia. Therefore, syncytium induction in HeLa cells
required two discrete adjacent domains within p21 that
in-cluded the 34-aa fusion domain identified in BJAB cells. It is
possible that clone BM caused a low level of fusion in BJAB
cells that was undetectable microscopically and caused a
de-tectable amount of fusion in HeLa cells due to the greater
sensitivity of the assay. Alternatively, there may be differences
in the interaction of regions of envelope with cellular
compo-nents from these two cell lines. There is also the possibility that
BJAB cells and HeLa cells differ in their cell surface
require-ments for fusion, akin to the different coreceptors in the case
of human immunodeficiency virus type 1 (HIV-1) cell fusion.
The syncytium-inducing phenotype of the chimeric
enve-lopes is not due to differences in the levels of intracellular
envelope.
The ability of viral envelope proteins to induce
syn-cytia is influenced by several factors, including intracellular
expression levels of the envelope, density of the fusion protein
on the cell surface, and correct protein configuration. We were
unable to examine the envelope expressed by transfection of
nonproductive genomic clones, as the levels of envelope
ex-pression were below detection by both
radioimmunoprecipita-tion and flow cytometry (data not shown). We therefore used
the high levels of HTLV envelope produced by the vaccinia
virus system to analyze expression of the recombinants.
We first examined the intracellular levels of envelope in the
vaccinia virus-infected cells, using a monoclonal antibody that
recognizes gp46 from HTLV-1 and HTLV-2 (Fig. 4). By
West-ern analysis on SDS-PAGE, approximately equal amounts of
precursor gp61 envelope were detected from all of the
chi-meric envelope clones. The majority of the envelope protein
existed as uncleaved precursor, similar to findings for infected
cells (data not shown). These results indicate that the
differ-FIG. 3. The amino terminus of HTLV-2 p21 is required for syncytium for-mation in HeLa cells. HeLa cells (106) were infected with wild-type vaccinia virus(WR) or VTF7-3 (MOI51.0) for 2 h at 37°C. HeLa cells that were infected with WR were transfected with pEM-ClacZbgAn, a plasmid in which the Escherichia coli lacZ gene has been linked to the T7 promoter. Cells that were infected with
VTF7-3 were transfected as described in Materials and Methods with vaccinia virus constructs expressing chimeric HTLV envelopes. At 16 h posttransfection, 105cells from each of the two sources were mixed and scored for blue syncytia
as described in Materials and Methods. These data represent the average of two duplicate wells and are representative of two independent experiments.h, HTLV-2;■, HTLV-1.
on November 9, 2019 by guest
http://jvi.asm.org/
ence in the ability of the clones to cause fusion in HeLa cells
was not attributable to differential expression levels of the
chimeric envelopes.
The chimeric envelopes are expressed at similar levels on
the cell surface of infected cells.
We next compared the levels
of expression of the recombinant envelopes on the cell surface
by flow cytometric analysis (Fig. 5). Compared to uninfected
cells, both wild-type HTLV-1 and -2 envelopes were efficiently
expressed by the vaccinia virus expression system. Similar
lev-els of HTLV envelope were detected on the surface of cells
infected with vaccinia virus expressing the various envelope
chimeras regardless of their ability to induce syncytia. The
anti-gp46 antibody was specific for the HTLV envelope, as we
detected a significant level of HTLV envelope on the cell
surface of the transfected cells over an isotype control antibody
(data not shown). The expression by all the recombinant
en-velope constructs of comparable levels of gp46 on the cell
surface suggested that the loss of syncytium induction was not
due to a defect in the transport of gp46 to the membrane.
The HTLV envelope chimeras are capable of forming
mul-timers.
Oligomerization of envelope proteins is necessary for
their proper function and viral infectivity. HTLV-1 envelope
has been previously reported to be capable of forming
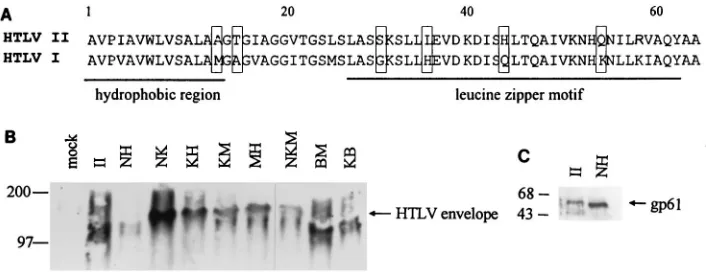
multim-ers (55). The 64-aa domain in HTLV-2 envelope that we have
identified as important in cell fusion and viral replication in
BJAB cells contains a region that bears similarities to a leucine
zipper motif (Fig. 6A) (7). The different syncytium-inducing
phenotypes may therefore be due to effects on the
multimer-ization of the envelope protein. We examined the
oligomer-ization potential of the chimeric envelopes by Western analysis
of native protein complexes separated on nondenaturing
pro-tein gels (Fig. 6B). The wild-type HTLV-1 migrated at
approx-imately 125 kDa, confirming that the HTLV-1 envelope exists
as a multimer. Similarly, the HTLV-2 envelope also migrated
at 125 kDa, indicating that the HTLV-2 envelope may also
function as a multimer. Analysis of the chimeric envelopes
revealed that all of the recombinant envelopes also migrated as
multimers. As an additional control, we first denatured the
HTLV protein complexes at 100°C and then separated them
on the same nondenaturing protein gels as specified above.
These proteins migrated at 61 kDa, the expected size for the
monomeric form of the HTLV envelope (Fig. 6C). There was
no correlation between the ability of the chimeric envelopes to
induce syncytia and their ability to form oligomers. Therefore,
the inability of some of the chimeric envelopes to induce
syn-cytia was not due to a failure to multimerize.
DISCUSSION
The amino terminus of HTLV-2 p21 encodes a fusion
do-main required for HTLV-2 syncytium induction and
replica-tion.
We have analyzed the requirements for syncytium
induc-tion by the HTLV-2 envelope protein. Our results have
identified a 64-aa domain located within the amino terminus of
the transmembrane protein as necessary for
HTLV-2-medi-ated cell fusion of BJAB cells and HeLa cells. This region was
not sufficient by itself to confer the syncytium-inducing
pheno-type, as the presence of this domain within the HTLV-1
enve-lope did not permit fusion. The ability of HTLV-2 genomic
clones to replicate in permissive cells correlated with the ability
to induce syncytia. We were unable to attribute the loss of the
syncytium-inducing phenotype to total cell levels, levels of
mem-brane associated, or oligomerization of the envelope protein.
In this study, we used two cell lines, BJAB and HeLa, that
were permissive for HTLV-2- but not HTLV-1-mediated cell
fusion. However, other groups have previously reported on the
ability of HTLV-1 to induce syncytia in HeLa cells (16, 18).
Agadjanyan et al. have also reported that several subclones of
a BJAB cell line were fusogenic upon cocultivation with both
HTLV-2 and HTLV-1 (1). Our inability to observe fusion with
HTLV-1 envelope in BJAB cells or HeLa cells may be a
prop-FIG. 4. The chimeric HTLV envelopes produce equivalent amounts ofin-tracellular gp61. HeLa cells (105) were infected with VTF7-3 (MOI51) for 2 h
at 37°C and transfected with vaccinia virus constructs expressing chimeric HTLV envelopes as described in Materials and Methods. Cells were harvested 16 h posttransfection. Total cell lysates (10mg) were separated on SDS–10% poly-acrylamide gels and analyzed for the presence of HTLV envelope by Western blot analysis as described in Materials and Methods. Sizes (in kilodaltons) of the molecular weight standards are indicated on the left; gp61 is indicated by the arrow on the right.
FIG. 5. The chimeric HTLV envelopes are expressed efficiently on the cell surface of infected cells. HeLa cells (105) were infected with VTF7-3 for 2 h at
37°C and transfected with vaccinia virus constructs expressing chimeric HTLV envelopes as described in Materials and Methods. Cells were harvested 16 h posttransfection, and 53103cells were analyzed by flow cytometry for
expres-sion of envelope on the cell surface as described in Materials and Methods. The histograms indicate relative cell number (y axis) as a function of relative amount of gp46 on the cell surface (x axis). Mock-infected cells are represented by the white histogram area, and cells transfected with chimeric HTLV envelopes are represented by the dark histogram area.
on November 9, 2019 by guest
http://jvi.asm.org/
erty of the cell lines upon propagation. Distinct properties may
arise in cell lines upon long-term culture, as illustrated by the
isolation of subclones of BJAB cells that have lost the ability to
form syncytia with HTLV-2 (1). Alternatively, the HTLV-1
envelope used in our studies may have cell tropisms distinct
from those of other HTLV-1 envelope clones. Sequence
anal-ysis between the Env1A envelope from SLB1 cells and the
envelope derived from a replication-competent HTLV-1 clone
(41) revealed no amino acid differences within the putative
fusion domain (data not shown). However, there are a total of
seven amino acid substitutions elsewhere in the envelope
pro-tein which may account for the differences observed.
Previous work in our lab has suggested that the
transmem-brane protein of HTLV-2 is required for cell fusion, as
expres-sion of gp46 alone was not sufficient to induce syncytia (45). In
addition, large substitutions within the HTLV-1
transmem-brane protein by murine leukemia virus envelope sequences
abolished HTLV-1-mediated cell fusion (17, 18). These studies
localized the fusion domain within the extracellular portion of
the transmembrane protein. We have further defined the
fu-sion domain in HTLV-2 to the amino terminus of p21. It is
likely that the corresponding region in HTLV-1 envelope
serves a similar role since linker insertion mutations within the
hydrophobic stretch in the N-terminal part of HTLV-1 p21
resulted in loss of syncytium induction (57).
Potential mechanisms of viral entry mediated by the
HTLV-2 fusion domain.
Fusion domains have been localized to
the transmembrane protein, and specifically to the amino
ter-minus, of other retroviral envelopes, including HIV-1 and
sim-ian immunodeficiency virus (25, 81). These domains are highly
hydrophobic with a predicted
a-helical structure. It has been
hypothesized that these domains form sided helixes with most
of the bulky hydrophobic residues on one side of the helix (15).
Located adjacent to these hydrophobic sequences are heptad
repeat sequences with nonpolar residues at the first and fourth
positions, similar to a leucine zipper motif (7). This region has
been postulated to play a role in stabilizing the oligomeric form
of these molecules. However, mutations in the HIV-1 leucine
zipper, while abolishing syncytium formation with CD4
1cells
and impairing infectivity, did not affect the ability of the
enve-lope protein to form oligomers (10, 22). Mutagenesis of the
HIV-1 envelope point to a critical role of the leucine zipper
motif in HIV-1 membrane fusion and virus entry, likely at a
post-CD4 binding step (82).
The HTLV-2 fusion domain that we have identified contains
a hydrophobic domain adjacent to a region with homology to
a leucine zipper motif. In BJAB cells, the syncytium-inducing
phenotype appeared to map to the leucine zipper region.
How-ever, substitution of the HTLV-2 leucine zipper domain did
not ablate fusion in HeLa cells, indicating that in some cell
types this domain alone may not be necessary for fusion. The
mechanism by which the HTLV-2 fusion domain contributes to
syncytium induction remains to be clarified. There was no
apparent correlation between the ability of the HTLV
enve-lope chimeras to induce syncytia and the intracellular or cell
surface expression, nor was there an apparent effect of the
recombinants on the formation of multimers, similar to the
HIV-1 results. One possible hypothesis is that the fusion
do-main in HTLV-2 p21 is required for receptor-mediated
con-formational changes. It is known that concon-formational changes
occur upon HIV-1 envelope binding to CD4. Post-CD4
bind-ing events included enhanced antibody bindbind-ing and cleavage by
an exogenous proteinase of the V3 loop, a major neutralizing
determinant in gp120 (13, 66), as well as exposure of previously
cryptic epitopes (67, 77). One indicator of overall changes in
the HIV-1 envelope protein is the observed enhanced
shed-ding of gp120 upon CD4 binshed-ding (4, 76, 78). Introduction of
prolines into highly conserved leucine or isoleucine residues of
the HIV-1 leucine zipper affected secretion of gp120 (9),
sug-gesting the involvement of this region in modifying the
enve-lope tertiary structure. Other viral fusion proteins similarly
undergo conformational changes in order to acquire their
fu-sion potential (29, 81). Therefore, the loss of syncytium
induc-tion and infectivity by our chimeric envelope constructs may be
due to an effect on the ability of the envelope to undergo the
appropriate conformational changes required for HTLV viral
entry.
[image:6.612.123.476.68.204.2]A second hypothesis for the role of the fusion domain in
HTLV-2 in syncytium induction may be at the level of receptor
utilization. Precedence for this possibility exists for HIV-1,
where syncytium formation can be mediated by sequences
which include the V3 loop in gp120 (13, 26, 54). The V3 loop
has also been implicated in the cell tropism observed for
var-ious strains of HIV-1. Changes in the V3 loop can influence the
FIG. 6. The chimeric HTLV envelopes are similar in the ability to form multimers. HeLa cells (106) were infected with VTF7-3 for 2 h at 37°C and transfected withvaccinia virus constructs expressing chimeric HTLV envelopes as described in Materials and Methods. Cells were harvested 16 h posttransfection and lysed in 500ml of OGL buffer. (A) Sequence comparison of the amino terminus of HTLV-2 and HTLV-1 p21 proteins. The first 64 aa of HTLV-2 and HTLV-1 p21 are represented. The nonconservative amino acid changes between HTLV-2 and HTLV-1 are boxed. (B) Western analysis of native envelope proteins. Total-cell lysates (20ml) were separated on a 5 to 20% gradient nondenaturing acrylamide gel with 0.01% SDS in the running buffer as described in Materials and Methods. HTLV envelope was detected by Western blot analysis as described in Materials and Methods. Sizes (in kilodaltons) of the molecular weight standards are indicated on the left; the multimeric forms of HTLV envelope are indicated by the arrow on the right. (C) Western analysis of denatured envelope proteins. Total-cell lysates (20ml) were heated to 100°C for 5 min and separated on a 5 to 20% gradient nondenaturing acrylamide gel with 0.01% SDS in the running buffer as described in Materials and Methods. HTLV envelope was detected by Western blot analysis as described in Materials and Methods. Sizes of the molecular weight standards are indicated on the left; gp61 is indicated by the arrow on the right.
on November 9, 2019 by guest
http://jvi.asm.org/
efficiency of entry of different HIV-1 strains into different cell
types, such as T-cell lines, macrophages, and microglial cells
(11, 36, 38, 50, 70, 73, 74). Although the primary receptor for
HIV-1 binding to cells is CD4, recent reports have identified
secondary receptors, CCR-5 and CXCR-4, that modulate viral
entry. CCR-5 and CXCR-4 are both members of the
chemo-kine receptor family and mediate infection at an early stage by
macrophage-tropic and T-cell-tropic viruses, respectively (2, 5,
19–21, 24, 43). The ability of HIV-1 to utilize CCR-5 or
CXCR-4 has been mapped to the V3 loop (12, 53), previously
implicated in syncytium formation and viral entry. Using a
highly sensitive syncytium assay, we have described cell lines
that display differential abilities to be fused by HTLV-1 and -2.
Yet, based on receptor interference studies, HTLV-1 and -2
have been postulated to share a common receptor on the tk-1
region of human chromosome 17 (27, 75). Therefore, it is
conceivable that, analogous to HIV-1, the fusion capability of
HTLV-1 and -2 in BJAB and HeLa cells is mediated by
inter-action of the fusion domain with accessory molecules that
determine cellular tropism.
ACKNOWLEDGMENTS
We thank Qi-Xiang Li, Jia-Qi Zhao, and Marilee Greenwald for
assistance in preparation of vaccinia virus stocks, and we thank
Yi-ming Xie for assistance with the blue syncytium assay.
This work was supported by NIH grant CA38597 and the Leukemia
Society of America. B.P. was supported by Public Health Service
train-ing grant GM07185, and I.S.Y.C. was a Scholar of the Leukemia
Society of America.
REFERENCES
1. Agadjanyan, M. G., K. E. Ugen, B. Wang, W. V. Williams, and D. B. Weiner. 1994. Identification of an 80-kilodalton membrane glycoprotein important for human T-cell leukemia virus type I and type II syncytium formation and infection. J. Virol. 68:485–493.
2. Alkhatib, G., C. Combadiere, C. C. Broder, Y. Feng, P. E. Kennedy, P. M. Murphy, and E. A. Berger.1996. CC CKR5: a RANTES, MIP-1alpha, MIP-1beta receptor as a fusion cofactor for macrophage-tropic HIV-1. Sci-ence 272:1955–1958.
3. Arp, J., M. LeVatte, J. Rowe, S. Perkins, E. King, C. Leystra-Lantz, S. K. Foung, and G. A. Dekaban.1997. A source of glycosylated human T-cell lymphotropic virus type 1 envelope protein: expression of gp46 by the vac-cinia virus/T7 polymerase system. J. Virol. 70:7349–7359.
4. Berger, E. A., J. R. Sisler, and P. L. Earl. 1992. Human immunodeficiency virus type 1 envelope glycoprotein molecules containing membrane fusion-impairing mutations in the V3 region efficiently undergo soluble CD4-stim-ulated gp120 release. J. Virol. 66:6208–6212.
5. Berson, J. F., D. Long, B. J. Doranz, J. Rucker, F. R. Jirik, and R. W. Doms. 1996. A seven-transmembrane domain receptor involved in fusion and entry of T-cell-tropic human immunodeficiency virus type 1 strains. J. Virol. 70: 6288–6295.
6. Cann, A. J., and I. S. Y. Chen. 1997. Human T-cell leukemia virus types I and II, p. 1856. In B. N. Fields, D. M. Knipe, and P. M. Howley (ed.), Fields virology, 3rd ed. Lippincott-Raven Publishers, Philadelphia, Pa.
7. Chambers, P., C. R. Pringle, and A. J. Easton. 1990. Heptad repeat se-quences are located adjacent to hydrophobic regions in several types of virus fusion glycoproteins. J. Gen. Virol. 71:3075–3080.
8. Chen, I. S., J. McLaughlin, J. C. Gasson, S. C. Clark, and D. W. Golde. 1983. Molecular characterization of genome of a novel human T-cell leukaemia virus. Nature 305:502–505.
9. Chen, S. S. 1994. Functional role of the zipper motif region of human immunodeficiency virus type 1 transmembrane protein gp41. J. Virol. 68: 2002–2010.
10. Chen, S. S., C. N. Lee, W. R. Lee, K. McIntosh, and T. H. Lee. 1993. Mutational analysis of the leucine zipper-like motif of the human immuno-deficiency virus type 1 envelope transmembrane glycoprotein. J. Virol. 67: 3615–3619.
11. Chesebro, B., J. Nishio, S. Perryman, A. Cann, W. O’Brien, I. S. Chen, and K. Wehrly.1991. Identification of human immunodeficiency virus envelope gene sequences influencing viral entry into CD4-positive HeLa cells, T-leukemia cells, and macrophages. J. Virol. 65:5782–5789.
12. Choe, H., M. Farzan, Y. Sun, N. Sullivan, B. Rollins, P. D. Ponath, L. Wu, C. R. Mackay, G. LaRosa, W. Newman, N. Gerard, C. Gerard, and J. Sodroski.1996. The beta-chemokine receptors CCR3 and CCR5 facilitate infection by primary HIV-1 isolates. Cell 85:1135–1148.
13. Clements, G. J., M. J. Price-Jones, P. E. Stephens, C. Sutton, T. F. Schulz, P. R. Clapham, J. A. McKeating, M. O. McClure, S. Thomson, and M. Marsh.1991. The V3 loops of the HIV-1 and HIV-2 surface glycoproteins contain proteolytic cleavage sites: a possible function in viral fusion? AIDS Res. Hum. Retroviruses 7:3–16.
14. Delahunty, M. D., I. Rhee, E. O. Freed, and J. S. Bonifacino. 1996. Muta-tional analysis of the fusion peptide of the human immunodeficiency virus type 1: identification of critical glycine residues. Virology 218:94–102. 15. Delamarre, L., C. Pique, D. Pham, T. Tursz, and M. C. Dokhelar. 1994.
Identification of functional regions in the human T-cell leukemia virus type I SU glycoprotein. J. Virol. 68:3544–3549.
16. Denesvre, C., C. Carrington, A. Corbin, Y. Takeuchi, F. L. Cosset, T. Schulz, M. Sitbon, and P. Sonigo.1996. TM domain swapping of murine leukemia virus and human T-cell leukemia virus envelopes confers different infectious abilities despite similar incorporation into virions. J. Virol. 70:4380–4386. 17. Denesvre, C., P. Sonigo, A. Corbin, H. Ellerbrok, and M. Sitbon. 1995.
Influence of transmembrane domains on the fusogenic abilities of human and murine leukemia retrovirus envelopes. J. Virol. 69:4149–4157. 18. Deng, H., R. Liu, W. Ellmeier, S. Choe, D. Unutmaz, M. Burkhart, P. Di
Marzio, S. Marmon, R. E. Sutton, C. M. Hill, C. B. Davis, S. C. Peiper, T. J. Schall, D. R. Littman, and N. R. Landau.1996. Identification of a major co-receptor for primary isolates of HIV-1. Nature 381:661–666.
19. de The, G., and R. Bomford. 1993. An HTLV-I vaccine: why, how, for whom? AIDS Res. Hum. Retroviruses 9:381–386.
20. Doranz, B. J., J. Rucker, Y. Yi, R. J. Smyth, M. Samson, S. C. Peiper, M. Parmentier, R. G. Collman, and R. W. Doms.1996. A dual-tropic primary HIV-1 isolate that uses fusin and the beta-chemokine receptors CKR-5, CKR-3, and CKR-2b as fusion cofactors. Cell 85:1149–1158.
21. Dragic, T., V. Litwin, G. P. Allaway, S. R. Martin, Y. Huang, K. A. Na-gashima, C. Cayanan, P. J. Maddon, R. A. Koup, J. P. Moore, and W. A. Paxton.1996. HIV-1 entry into CD41cells is mediated by the chemokine
receptor CC-CKR-5. Nature 381:667–673.
22. Dubay, J. W., S. J. Roberts, B. Brody, and E. Hunter. 1992. Mutations in the leucine zipper of the human immunodeficiency virus type 1 transmembrane glycoprotein affect fusion and infectivity. J. Virol. 66:4748–4756. 23. Evans, B. K., I. Gore, L. E. Harrell, T. Arnold, and S. J. Oh. 1989.
HTLV-I-associated myelopathy and polymyositis in a US native. Neurology 39: 1572–1575.
24. Feng, Y., C. C. Broder, P. E. Kennedy, and E. A. Berger. 1996. HIV-1 entry cofactor: functional cDNA cloning of a seven-transmembrane, G protein-coupled receptor. Science 272:872–877.
25. Freed, E. O., D. J. Myers, and R. Risser. 1990. Characterization of the fusion domain of the human immunodeficiency virus type 1 envelope glycoprotein gp41. Proc. Natl. Acad. Sci. USA 87:4650–4654.
26. Freed, E. O., D. J. Myers, and R. Risser. 1991. Identification of the principal neutralizing determinant of human immunodeficiency virus type 1 as a fusion domain. J. Virol. 65:190–194.
27. Gavalchin, J., N. Fan, P. G. Waterbury, E. Corbett, B. D. Faldasz, S. M. Peshick, B. J. Poiesz, L. Papsidero, and M. J. Lane.1995. Regional local-ization of the putative cell surface receptor for HTLV-I to human chromo-some 17q23.2-17q25.3. Virology 212:196–203.
28. Gessain, A., F. Barin, J. C. Vernant, O. Gout, L. Maurs, A. Calender, and G. de The.1985. Antibodies to human T-lymphotropic virus type-I in patients with tropical spastic paraparesis. Lancet ii:407–410.
29. Gilbert, J. M., L. D. Hernandez, J. W. Balliet, P. Bates, and J. M. White. 1995. Receptor-induced conformational changes in the subgroup A avian leukosis and sarcoma virus envelope glycoprotein. J. Virol. 69:7410–7415. 30. Green, P. L., Y. M. Xie, and I. S. Chen. 1990. The internal methionine codons
of human T-cell leukemia virus type II rex gene are not required for p24rex
production or virus replication and transformation. J. Virol. 64:4914–4921. 31. Hadlock, K. G., C. J. Goh, P. A. Bradshaw, S. Perkins, J. Lo, J. E. Kaplan, R. Khabbaz, and S. K. Foung.1995. Delineation of an immunodominant and human T-cell lymphotropic virus (HTLV)-specific epitope within the HTLV-I transmembrane glycoprotein. Blood 86:1392–1399.
32. Hall, W. M., H. Takahashi, C. Liu, M. H. Kaplan, O. Schneewind, S. Ijichi, K. Nagashima, and R. Gallo.1992. Multiple isolates and characteristics of human T-cell leukemia virus type II. J. Virol. 66:2456–2463.
33. Harrington, W. J., W. Sheremata, B. Hjelle, D. K. Dube, P. Bradshaw, S. K. Foung, S. Snodgrass, G. Toedter, L. Cabral, and B. Poiesz.1993. Spastic ataxia associated with human T-cell lymphotropic virus type II infection. Ann. Neurol. 33:411–414.
34. Horal, P., W. W. Hall, B. Svennerholm, J. Lycke, S. Jeansson, L. Rymo, M. H. Kaplan, and A. Vahlne.1991. Identification of type-specific linear epitopes in the glycoproteins gp46 and gp21 of human T-cell leukemia viruses type I and type II using synthetic peptides. Proc. Natl. Acad. Sci. USA 88:5754–5758.
35. Hoshino, H., M. Shimoyana, M. Miwa, and T. Sugimura. 1983. Detection of lymphocytes producing a human retrovirus associated with adult T-cell leu-kemia by syncytia induction assay. Proc. Natl. Acad. Sci. USA 80:7337–7341. 36. Hwang, S. S., T. J. Boyle, H. K. Lyerly, and B. R. Cullen. 1991. Identification of the envelope V3 loop as the primary determinant of cell tropism in HIV-1. Science 253:71–74.
on November 9, 2019 by guest
http://jvi.asm.org/
37. Ishii, K., K. Yamato, Y. Iwahara, T. Eguchi, Y. Uemura, N. Takehara, Y. Ohtsuki, H. Taguchi, and I. Miyoshi.1991. Isolation of HTLV-I from muscle of a patient with polymyositis. Am. J. Med. 90:267–269.
38. Ivanoff, L. A., D. J. Looney, C. McDanal, J. F. Morris, F. Wong-Staal, A. J. Langlois, S. R. Petteway, and T. J. Matthews.1991. Alteration of HIV-1 infectivity and neutralization by a single amino acid replacement in the V3 loop domain. AIDS Res. Hum. Retroviruses 7:595–603.
39. Jacobson, S., T. Lehky, M. Nishimura, S. Robinson, D. E. McFarlin, and S. Dhib-Jalbut.1993. Isolation of HTLV-II from a patient with chronic, pro-gressive neurological disease clinically indistinguishable from HTLV-I-asso-ciated myelopathy/tropical spastic paraparesis. Ann. Neurol. 33:392–396. 40. Jacobson, S., V. Zaninovic, C. Mora, P. Rodgers-Johnson, W. A. Sheremata,
C. J. Gibbs, C. Gajdusek, and D. E. McFarlin.1988. Immunological findings in neurological diseases associated with antibodies to HTLV-I: activated lymphocytes in tropical spastic paraparesis. Ann. Neurol. 23(Suppl.):S196– S200.
41. Kimata, J. T., F. H. Wong, J. J. Wang, and L. Ratner. 1994. Construction and characterization of infectious human T-cell leukemia virus type 1 molecular clones. Virology 204:656–664.
42. Kitajima, I., K. Yamamoto, K. Sato, Y. Nakajima, T. Nakajima, I. Ma-ruyama, M. Osame, and K. Nishioka. 1991. Detection of human T cell lymphotropic virus type I proviral DNA and its gene expression in synovial cells in chronic inflammatory arthropathy. J. Clin. Invest. 88:1315–1322. 43. Lapham, C. K., J. Ouyang, B. Chandrasekhar, N. Y. Nguyen, D. S. Dimitrov,
and H. Golding.1996. Evidence for cell-surface association between fusin and the CD4-gp120 complex in human cell lines. Science 274:602–605. 44. Lee, H., P. Swanson, V. S. Shorty, J. A. Zack, J. D. Rosenblatt, and I. S.
Chen.1989. High rate of HTLV-II infection in seropositive i.v. drug abusers in New Orleans. Science 244:471–475.
45. Li, Q. X., D. Camerini, Y. Xie, M. Greenwald, D. R. Kuritzkes, and I. S. Chen.1996. Syncytium formation by recombinant HTLV-II envelope glyco-protein. Virology 218:279–284.
46. Miyamoto, K., N. Tomita, A. Ishii, T. Nishizaki, K. Kitajima, T. Tanaka, T. Nakamura, S. Watanabe, and T. Oda. 1984. Transformation of ATLA-negative leukocytes by blood components from anti-ATLA-positive donors in vitro. Int. J. Cancer 33:721–725.
47. Nagy, K., P. Clapham, R. Cheingsong-Popov, and R. Weiss. 1983. Human T-cell leukemia virus type I: induction of syncytia and inhibition by patients’ sera. Int. J. Cancer 32:321–328.
48. Nakao, K., and N. Ohba. 1993. Clinical features of HTLV-I associated uveitis. Br. J. Ophthalmol. 77:274–279.
49. Nakao, K., N. Ohba, and M. Matsumoto. 1989. Noninfectious anterior uve-itis in patients infected with human T-lymphotropic virus type I. Jpn. J. Ophthalmol. 33:472–481.
50. O’Brien, W. A., Y. Koyanagi, A. Namazie, J. Q. Zhao, A. Diagne, K. Idler, J. A. Zack, and I. S. Chen.1990. HIV-1 tropism for mononuclear phagocytes can be determined by regions of gp120 outside the CD4-binding domain. Nature 348:69–73.
51. Ohba, N., M. Matsumoto, M. Sameshima, Y. Kabayama, K. Nakao, K. Unoki, F. Uehara, K. Kawano, I. Maruyama, and M. Osame.1989. Ocular manifestations in patients infected with human T-lymphotropic virus type I. Jpn. J. Ophthalmol. 33:1–12.
52. Okada, M., Y. Koyanagi, N. Kobayashi, Y. Tanaka, M. Nakai, K. Sano, K. Takeuchi, Y. Hinuma, M. Hatanaka, and N. Yamamoto. 1984. In vitro infection of human B lymphocytes with adult T-cell leukemia virus. Cancer Lett. 22:11–21.
53. Oravecz, T., M. Pall, and M. A. Norcross. 1996. Beta-chemokine inhibition of monocytotropic HIV-1 infection. Interference with a postbinding fusion step. J. Immunol. 157:1329–1332.
54. Page, K. A., S. M. Stearns, and D. R. Littman. 1992. Analysis of mutations in the V3 domain of gp160 that affect fusion and infectivity. J. Virol. 66:524–533. 55. Paine, E., R. Gu, and L. Ratner. 1994. Structure and expression of the human
T-cell leukemia virus type 1 envelope protein. Virology 199:331–338. 56. Palker, T. J., E. R. Riggs, D. E. Spragion, A. J. Muir, R. M. Scearce, R. R.
Randall, M. W. McAdams, A. McKnight, P. R. Clapham, and R. A. Weiss. 1992. Mapping of homologous, amino-terminal neutralizing regions of hu-man T-cell lymphotropic virus type I and II gp46 envelope glycoproteins. J. Virol. 66:5879–5889.
57. Pique, C., T. Tursz, and M. C. Dokhelar. 1990. Mutations introduced along the HTLV-I envelope gene result in a non-functional protein: a basis for envelope conservation? EMBO J. 9:4243–4248.
58. Poiesz, B. J., F. W. Ruscetti, A. F. Gazdar, P. A. Bunn, J. D. Minna, and R. C. Gallo.1980. Detection and isolation of type C retrovirus particles from fresh and cultured lymphocytes of a patient with cutaneous T-cell lymphoma. Proc. Natl. Acad. Sci. USA 77:7415–7419.
59. Popovic, M., G. Lange-Wantzin, P. S. Sarin, D. Mann, and R. C. Gallo. 1983. Transformation of human umbilical cord blood T cells by human T-cell leukemia/lymphoma virus. Proc. Natl. Acad. Sci. USA 80:5402–5406. 60. Popovic, M., P. S. Sarin, M. Robert-Gurroff, V. S. Kalyanaraman, D. Mann,
J. Minowada, and R. C. Gallo.1983. Isolation and transmission of human retrovirus (human t-cell leukemia virus). Science 219:856–859.
61. Robert-Guroff, M., S. H. Weiss, J. A. Giron, A. M. Jennings, H. M. Ginzburg, I. B. Margolis, W. A. Blattner, and R. C. Gallo.1986. Prevalence of anti-bodies to HTLV-I, -II, and -III in intravenous drug abusers from an AIDS endemic region. JAMA 255:3133–3137.
62. Rosenblatt, J. D., J. C. Gasson, J. Glaspy, S. Bhuta, M. Aboud, I. S. Chen, and D. W. Golde.1987. Relationship between human T cell leukemia virus-II and atypical hairy cell leukemia: a serologic study of hairy cell leukemia patients. Leukemia 1:397–401.
63. Rosenblatt, J. D., D. W. Golde, W. Wachsman, J. V. Giorgi, A. Jacobs, G. M. Schmidt, S. Quan, J. C. Gasson, and I. S. Chen.1986. A second isolate of HTLV-II associated with atypical hairy-cell leukemia. N. Engl. J. Med. 315:372–377.
64. Sandler, S. G. 1986. HTLV-I and -II. New risks for recipients of blood transfusions? JAMA 256:2245–2246.
65. Sato, K., I. Maruyama, Y. Maruyama, I. Kitajima, Y. Nakajima, M. Higaki, K. Yamamoto, N. Miyasaka, M. Osame, and K. Nishioka.1991. Arthritis in patients infected with human T lymphotropic virus type I. Clinical and immunopathologic features. Arthritis Rheum. 34:714–721.
66. Sattentau, Q. J., and J. P. Moore. 1991. Conformational changes induced in the human immunodeficiency virus envelope glycoprotein by soluble CD4 binding. J. Exp. Med. 174:407–415.
67. Sattentau, Q. J., J. P. Moore, F. Vignaux, F. Traincard, and P. Poignard. 1993. Conformational changes induced in the envelope glycoproteins of the human and simian immunodeficiency viruses by soluble receptor binding. J. Virol. 67:7383–7393.
68. Saxon, A., R. H. Stevens, and D. W. Golde. 1978. T-lymphocyte variant of hairy-cell leukemia. Ann. Intern. Med. 88:323–326.
69. Seiki, M., S. Hattori, Y. Hirayama, and M. Yoshida. 1983. Human adult T-cell leukemia virus: complete nucleotide sequence of the provirus genome integrated in leukemia cell DNA. Proc. Natl. Acad. Sci. USA 80:3618–3622. 70. Sharpless, N. E., W. A. O’Brien, E. Verdin, C. V. Kufta, I. S. Chen, and M. Dubois-Dalcq.1992. Human immunodeficiency virus type 1 tropism for brain microglial cells is determined by a region of the Env glycoprotein that also controls macrophage tropism. J. Virol. 66:2588–2593.
71. Shida, H., T. Tochikura, T. Sato, T. Konno, K. Hirayoshi, M. Seki, Y. Ito, M. Hatanaka, Y. Hinuma, and M. Sugimoto.1987. Effect of the recombinant vaccinia viruses that express HTLV-I envelope gene on HTLV-I infection. EMBO J. 6:3379–3384.
72. Shimotohno, K., Y. Takahashi, N. Shimizu, T. Gojobori, D. W. Golde, I. S. Chen, M. Miwa, and T. Sugimura.1985. Complete nucleotide sequence of an infectious clone of human T-cell leukemia virus type II: an open reading frame for the protease gene. Proc. Natl. Acad. Sci. USA 82:3101–3105. 73. Shioda, T., J. A. Levy, and C. Cheng-Mayer. 1991. Macrophage and T
cell-line tropisms of HIV-1 are determined by specific regions of the enve-lope gp120 gene. Nature 349:167–169.
74. Shioda, T., J. A. Levy, and C. Cheng-Mayer. 1992. Small amino acid changes in the V3 hypervariable region of gp120 can affect the T-cell-line and mac-rophage tropism of human immunodeficiency virus type 1. Proc. Natl. Acad. Sci. USA 89:9434–9438.
75. Sommerfelt, M. A., B. P. Williams, P. R. Clapham, E. Solomon, P. N. Goodfellow, and R. A. Weiss.1988. Human T cell leukemia viruses use a receptor determined by human chromosome 17. Science 242:1557–1559. 76. Stamatatos, L., and C. Cheng-Mayer. 1993. Evidence that the structural
conformation of envelope gp120 affects human immunodeficiency virus type 1 infectivity, host range, and syncytium-forming ability. J. Virol. 67:5635–5639. 77. Stamatatos, L., and C. Cheng-Mayer. 1995. Structural modulations of the envelope gp120 glycoprotein of human immunodeficiency virus type 1 upon oligomerization and differential V3 loop epitope exposure of isolates dis-playing distinct tropism upon virion-soluble receptor binding. J. Virol. 69: 6191–6198.
78. Thali, M., C. Furman, E. Helseth, H. Repke, and J. Sodroski. 1992. Lack of correlation between soluble CD4-induced shedding of the human immuno-deficiency virus type 1 exterior envelope glycoprotein and subsequent mem-brane fusion events. J. Virol. 66:5516–5524.
79. Tyagi, R. K., B. R. Babu, and K. Datta. 1993. Simultaneous determination of native and subunit molecular weights of proteins by pore limit electrophore-sis and restricted use of sodium dodecyl sulfate. Electrophoreelectrophore-sis 14:826–828. 80. Weiss, R. A., P. Clapham, K. Nagy, and H. Hoshino. 1985. Envelope prop-erties of human T-cell leukemia viruses. Curr. Top. Microbiol. Immunol. 115:235–246.
81. White, J. M. 1990. Viral and cellular membrane fusion proteins. Annu. Rev. Physiol. 52:675–697.
82. Wild, C., J. W. Dubay, T. Greenwell, T. J. Baird, T. G. Oas, C. McDanal, E. Hunter, and T. Matthews.1994. Propensity for a leucine zipper-like domain of human immunodeficiency virus type 1 gp41 to form oligomers correlates with a role in virus-induced fusion rather than assembly of the glycoprotein complex. Proc. Natl. Acad. Sci. USA 91:12676–12680.
83. Yoshida, M., M. Seiki, K. Yamaguchi, and K. Takatsuki. 1984. Monoclonal integration of human T-cell leukemia provirus in all primary tumors of adult T-cell leukemia suggests causative role of human T-cell leukemia virus in the disease. Proc. Natl. Acad. Sci. USA 81:2534–2537.