Protein
Composition
of the
Structural
Components
of
Vesicular Stomatitis Virus
ROBERT R.WAGNER, TERRY C. SCHNAITMAN,RUTH M.SNYDER,ANDCARL A.SCHNAITMAN
Department ofMicrobiology, The University of Virginia School of Medicine, Charlottesville, Virginia 22901
Received forpublication 24 February 1969
Digitonin, a sterol glycoside which complexes with cholesterol, stripped off
the envelope of vesicular stomatitis (VS) virions and liberated two viral
struc-turalproteins, 83% ofP6 and 53% ofP4. Deoxycholate alsodisruptedVSvirions
but released nucleocapsidcoreswhichcould beidentifiedby higher buoyant density,
ratio ofincorporated 3H-uridineto "C-protein, and electron microscopy.Themajor
nucleocapsid protein was P5 but varying amounts of the minor protein aggregate
P2 were present, depending on the concentration of urea used for extraction. P2
appeared tobe a polymer of P5. Two otherminor structural proteins, P1 and P3,
couldnot be located in the virion. From these data, we conclude that the three
microscopically identifiablestructuresof VS virionsareeachcomposed primarilyofa
single major protein,asfollows:P6 = envelopeprotein,P4 = proteinofunderlying
"shell," and P5 = nucleocapsid protein.
In thepreceding paper (10) the structural
pro-teins of vesicular stomatitis (VS) viruses were
characterizedbypolyacrylamide gel
electrophore-sis afterdissolution ofpurified virionswith acetic
acid, sodium dodecyl sulfate (SDS), 0.5 M urea,
and 2-mercaptoethanol. Three major proteins
wereidentifiedand designatedP4, P5, and P6. In addition, two consistent minor stainable bands andcoincidentpeaksofradioactivity (P2andP3)
appeared to be aggregates ofthe major proteins on the basis oftheir susceptibility to 8 M urea.
Anotherminorprotein, P1, was resistant to
frac-tionation by8Murea.
This report describes experiments designed to
locate these VS viral proteins in the
micro-scopically identifiable structural components of thevirion.
MATERIALS AND METHODS
The materials and basicprocedures have been de-scribed in detail in theprecedingreport (10) andare
outlined below.
Production and puriication oflabeled virus. Stock preparations of the Indiana serotype of VS viruswere
grownin 10monolayer cultures ofLcells, -1.8 X 107
cells per monolayer. Virus yields in 17 hr were ap-proximately S X 1010 plaque-forming units in100 ml ofmedium. Viralproteinswere labeled by adding to special basal medium, Eagle's (BME) 10,pc/ml each
of3H-leucine (10.3to14.7c/mmole) and3H-tyrosine
(28.2 to 43 c/mmole) or 1.13 ,c/ml of uniformly labeled 'IC-amino acids (54 mc/matom). In several
experiments in which the proteins were labeled with
"4C-aminoacids,viral RNA waslabeled with 10uc/ml
of3H-uridine (20.0c/mmole) addedto the same cul-ture media butcontainingactinomycin (0.33,g/ml). Batches (100 ml)oflabeled crude virus were partially purified by differential centrifugation andwere
con-centratedto0.5or 1.0ml.The concentratedviruswas
thenpurified byratezonalcentrifugation in0to40% linear sucrose gradients and then by isopycnic sedi-mentation in CsCl (see 10).
Electron microscopy. Specimens were examined and photographed in Siemens-Halske Elmiskop I
microscopes by John W. Greenawalt and Glenn Decker at JohnsHopkins University or by Carl A. Schnaitman at the University of Virginia. Purified virions or fractionated viral components were nega-tively stained with phosphotungstic acid (PTA), as
previously described (3). Insome casesthe viral
com-ponents were dialyzed overnight against2% PTA in distilled water before being placed on
Formvar-coveredgrids reinforced with carbon. Pelletedvirions werefixed in glutaraldehyde andOS04,embedded in epoxy resin, sectioned with a diamond knife, and stained withuranylacetateandlead citrate.
Polyacrylamide gel electrophoresis. The method of Maizel (5) was used with only minor modifications.
Inbrief, viral proteins (2.5to 3.5 mg/ml) were solu-bilizedby addition ofatenthvolumeof glacial acetic acid and then made 0.5or8 M with respecttoureaand 1% withrespect to SDS. After incubation at 37 C for 1 hr, thesuspension was then dialyzed at room tem-peraturefor16 to 18hragainst200to400ml of 0.01 M phosphate buffer (pH 7.2) containing 0.1% SDS, 0.5
or 8 M urea, and 0.1% 2-mercaptoethanol. Protein samples of 0.1 to0.2mlwith added bromophenol blue were layered with sucrose on 7.5% polyacrylamide gelscontaining0.1 Mphosphate (pH 7.2),0.5 M urea, and 0.1% SDS. Electrophoresis was carried out at 611
on November 11, 2019 by guest
http://jvi.asm.org/
then slicedtransversely into sections of 1.25mmeach; thegel slices weredepolymerizedwith30% H202, dis-solved in N-chlorosuccinimide (NCS), and diluted in toluene-based fluors. The radioactivity was counted in ascintillation spectrometer (Packard Instrument Co., Inc., Downers Grove, lll.).
RESULTS
Structure of VS virions. Several groups of
in-vestigators have described the structural
com-ponents of VS virions in somedetail (3, 6, 7, 9).
Allmicroscopists appear to agree on the presence
of a surface membranous structure or envelope
with protruding spikes. In addition, an internal
component,appearingascloselyspacedstriations
in intact virions, can be released as a coiled or
partially uncoiled helix, which probably
repre-sents the nucleoprotein core. There is some
dis-agreement aboutthelength and origin of the coils
and about the presenceorabsence of a structure
between thecoreand theenvelope.
Figure 1A shows the structural components of
negatively stainedVSvirionsatvarious stages of
disorganization. Three distinct structures can be
seen. Figure1Balso revealsatleastthree concen-tric layers in virions sectioned transversely and
stained with uranyl acetate and lead. These
ob-servations leadus toconfirm that VSvirions are
cylinders composed of (i)an outerenvelope with
spikes, (ii) a rigid "shell" underneath the
sur-rounding envelope, and (iii) an inner, packed
nucleoprotein coil.
Fractionation of VS virions with digitonin. VS
virions mature at the cell surface, bud from the
cytoplasmic membrane, and appear to have a
high content of lipid, particularly cholesterol,
which is almost undoubtedly associated with the
envelope (7, 9). JamesMcSharry of this
depart-ment (unpublished data) confirmed the presence
oflargeamountsof cholesterol andobservedthat
3H-cholesterol is rapidly incorporated during
growth ofVS virus; the 3H label was retained by
VS virions after extensive purification. These
findings ledus tobelieve that the envelope and its
associatedproteinscould bestrippedoff by
treat-ment with the sterol glycoside, digitonin.
Digi-tonin complexes with cholesterol (2) and has
been used successfully by Schnaitman et al. (8)
to dissociate the outer and inner membranes of
mitochondria. A similar technique was used by
de-The (1) to remove the envelope of Rauscher
leukemia virus from itsnucleocapsidcore.
Infectious B virions of VS virus labeled with
4C-amino acids were purified by rate zonal and
viral suspension in a ratio of 0.4 mg of digitonin
to 1 mgof viralprotein. After incubation at 4 C
for 10 min, the virus-digitonin mixtures were
centrifuged at 40,000 rev/min (-130,000 X g)
for 90 min in 1 -ml tubes specially designed to fit
the SW 65 rotor. The supernatant fluids were
drawnoff, the tubesweredrained, and thevisible
pellets were suspended to their original volumes in
distilledwater orEarle'sbalancedsaltsolution.
Figure 2A shows an electron micrograph of
digitonin-treated pelleted virus stained with PTA.
Viral particles can be readily identified but show
considerable distortion, flattening, and lack oJ PTA penetration compared with relatively intact
virions (Fig. 1A). A prominent feature is that
these "shells" of VS virions appear to be devoid
ofenvelopes and characteristic spikes.
Samples of100,liters of supernatant fluids or
pellets of digitonin-treated VS virions labeled
with 'IC-amino acids were mixed with an equal
amount of intact purified VS virions that had been labeled with 3H-leucine and 3H-tyrosine to provide viral protein markers. Proteins were
ex-tractedfromthesemixtureswithacetic acid,0.5 M
urea, and 1% SDS; the solubilized I"C- and
3H-proteins werethensubjected tocoelectrophoresis on 7.5% polyacrylamide gels. The gels were stained for scanning and were sectioned for counting of bothisotopes.
The resultsof one of these experiments are
re-corded inFig. 3. As shown in Fig.3A,sixpeaks of
14Cradioactivity can be identified in pelleted virus
aftertreatmentwithdigitonin; each of thesepeaks corresponds exactly to the six peaks of optical density (OD)at610nmandtothesix3H-protein
markers extracted from intact virions. Only P6
wasappreciablyreduced in amount.
Figure 3B illustrates the results of
electropho-reticanalysis of thesupernatant-fluid proteins
re-leased from the same digitonin-treated virions
labeled with
"4C-amino
acids. Only P6 and P4were present in the supernatant fluid in
appre-ciableamounts.Table 1 summarizes these databy
showing the relative proportions of I4C counts
present in proteins at each gel position
corre-sponding to the six marker proteins. As
noted,
digitonin caused the release of 83% of P6 and
53% ofP4 fromVS virions but
only
3.5% ofP5andinsignificant amountsofP1, P2, and P3.
These data suggest that the primary
en-velope-associated proteinisP6but thatdigitonin
also dissociates about 50% of P4, presumably
from the surface of the virion.
Release of nucleocapsid from VS virions by
deoxycholate. Deoxycholate disrupts VS virions
on November 11, 2019 by guest
http://jvi.asm.org/
FIG. 1. Structure ofVS virions. (A) Purifiedandconcenitrated VS virionsat variousstagesofdisorganization after incubationindistilledwaterfor24hrat4 Cwerenegativelystainedwith PTA.Notetlheexternallayerwith spikes, the dense underlyinglayeror"shell",atd theinternal striationsrepresentingnucleoproteincoils. X 320,000.
(B) Transverse section of pelleted, fixed, andepoxy-embedded VS virions stainedwith uranylacetate and lead
citrate. Notethefringe of spikes,anunderlyingelectron-dense ring, and internalstructure,presumably
represenf-ing sectioned coils. X 400,000.
pow"
zi
on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.481.56.434.56.578.2]FIG. 2. DegradedVSvirionis. (A) ShellsofVSvirionstreated withdigitoniin,pelletedat130,000X g,andstailed
with PTA.Note thedenseshellsbuttherelativepaucityofsurroundingspikes. X 90,000. (B) Coilsof'VS virions
released bytreatmenit withdeoxycholate, separatedbyban1dingin a CsCl gradient, anddialyzedagainst PTA in
distilledwater.X 160,000.
on November 11, 2019 by guest
http://jvi.asm.org/
~~~~~~~~~~~~~~~~~5
.C 4
o02 0 4 0 6 o 2 0 4 0 6
~ ~ ~ Mlimtr ofglMlieeso e
Ps~~ ~~~
3~~~~~~~~~
2~~~~~
0pi~~~ ~ ~ ~ ~ ~ ~ ~ ~~~~F
slice gel were anlye fo 4-ad3-rt akrsetatdfo natvron.()Plee rcino
>20
20-~~ ~ ~ ~ ~ ~ ~ ~ ~
14
0 15I
digitonin-degrade virions, 6,152 coun0,c
dgdvro,,80~ cuDip3H
O~ ~ ~ ~
me rs of g-ueliirn ta ofr puef
virionsdegradedwithdigitoninandseparated by centrifugationat130,000X gfor90miin. Thetoppanelsrepresenit
tracings of gels stained with Coomassieblue andscannedat 610 nm in a Gilford spectrophotometer. Thesame
slicedgelswereanalyzed for "4C- and 31H-proteinimarkers extractedfrom intact virions. (A) Pelletedfraction of' digitonin-degraded virions, 6,152 "4C counts/mmnper100a~iters. (B) Supernatant fraction ofthesame
digitonlin-degraded virions,4,308 14Ccounts/minlper 100 jAtliters.
and releases internal coils (6, 7). We adopted
this method for controlled degradation of VS virions. For purification of the nucleocapsid
component, we took advantage of the fact
that intact VS virions have a relative buoyant
density in CsCl of approximately 1.20 g/ml (6,
10). We reasoned, correctly, that the lipid-free
nucleocapsid should havea higher buoyant
den-sity and, therefore, should be readily separable
from the enveloped virion by isopycnic sedimen-tation.
VS virionswere growninLcells in 1:50 BME without serum but containing 3H-uridine (10
lAc/ml)
and 'IC-amino acids (1.13 ,c/ml). The cells were exposed to actinomycin (0.334g/ml)
inthe mediumthroughout the period of infection.
Afterincubation for 8 hrat37C, equalvolumes of
Eagle's minimal essential medium with cold
amino acids and 2% calf serum were added to
TABLE 1. Percenttage of total14C radioactivity
re-covered frompolyacrylamideproteini fractiontsof
VS virionstreatedwith digitonini andcenztrifuged
at130,000 X ga
Percentageof total 14C Protein
Supernatantfluid Pellet
P1 0.2 3.5
P2 0.3 1.3
P3 0.2 0.9
P4 9.7 8.6
P5 1.1 29.8
P6 27.7 5.5
Total 39.2 49.6
aThe supernatant-fluid
14C
radioactivityap-plied to the gel contained 4,308 counts/min per
100 ,uiters, and the pellet radioactivity contained
6,152counts/minper 100,1iters.Dataareabstracted fromexperiment shown inFig. 3.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:5.481.49.445.69.366.2]WAGNER ET AL.
each culture. Media were harvested and pooled
17 hrafter infection, and the virions were
con-centrated and purified. The procedure adopted
for controlled degradation of VS virions was
toadd 1.25 mg of sodium deoxycholate
(Mathe-son,Colemanand Bell, Cincinnati, Ohio) to 0.25
ml ofthe labeled viral suspension at a protein concentration of approximately 1.5 mg/ml.
Higher concentrations of deoxycholate resulted
in lower yields of nucleocapsids. The viral
sus-pension was stirred constantly for 2 hr at room
temperature,then wasmixed with 4.5 ml of CsCl
in 0.01 M tris(hydroxymethyl)aminomethane (Tris) buffer, pH 7.8 (startingspecific gravity, 1.3
g/ml), and was centrifuged for 72 hr at 38,000
rev/min in theSW 50 rotor. Two clearly visible
and sharp light-scattering bands could be seen
consistently, but onlyasingleband could be seen
in control viral suspensions not exposed to
de-oxycholate.
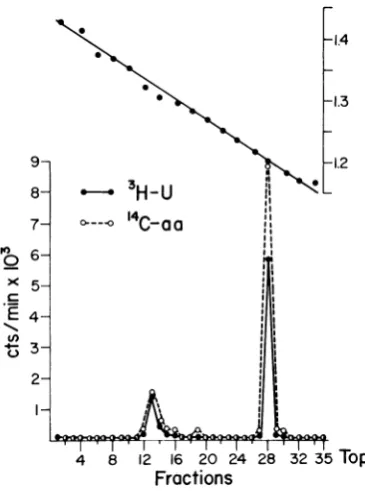
Figure4shows thedistributionofradioactivity in arepresentativeCsClgradient after
centrifuga-tion of deoxycholate-degraded purified virions
labeled with 3H-uridine and "4C-amino acids.
Coincident sharp peaks of 3H and 14C
disintegra-tionswere evident atdensities equivalent to 1.32
and 1.20g/ml. As noted, only 20% of the counts
in RNAand 14% of the counts in protein were
present in the high-density peak, suggesting that
most of the nucleoprotein was not dissociated fromlipid-containing virions. On repeated
deter-minations, the amount of
deoxycholate-dissoci-able3H-RNAranged from 10 to 30%.
Neverthe-less, the3H:14C ratio (RNA:protein) wasalways
greaterinthehigh-density region of thegradient.
Aminor error isimplicit in these calculations
be-cause some
"4C-amino
acidsareincorporatedinto RNA.The nucleoprotein releasedfrom VSvirionsby deoxycholate was completely resistant to
ribo-nuclease (40 ,ugof crystalline pancreatic
ribonu-cleaseper 100,uliters ofdegraded virion
suspen-sion).
Ribonuclease treatment did not affect thebuoyant density ofthe nucleoprotein or its
3H-uridine and
"4C-protein
content.Thehigh-densitynucleoprotein fraction was not infectious when
platedon Lcells.
Fraction 13 (p _ 1.32 g/ml) from the CsCl
gradient shown in Fig. 4 was dialyzed overnight against 2% PTA in distilled water and was
examinedbyelectronmicroscopy.Figure2Bshows
onerepresentativefield of thispreparation, which
reveals the characteristic coiled nucleoprotein of
VS virus (7, 9). Other fields and other similar
preparations often revealedaggregated coils, but
only rare intact
virions, always
markedly
dis-torted,wereencountered. The top
band,
fraction28
(Fig. 4),
p 1.20g/ml,
containedlarge
num-bersofrecognizable intactvirions, usually
aggre-gated and distorted, but only rare coils which
seemed to be attached tofragments ofviral debris.
Identification of nucleocapsid protein. The
high-and low-density fractions from CsCI gradients after isopycnic centrifugation of
deoxycholate-degraded VSvirionslabeledwith"4C-aminoacids
wereanalyzed for their protein constituents.
3H-proteins extracted from intact virions were used
asmarkers.
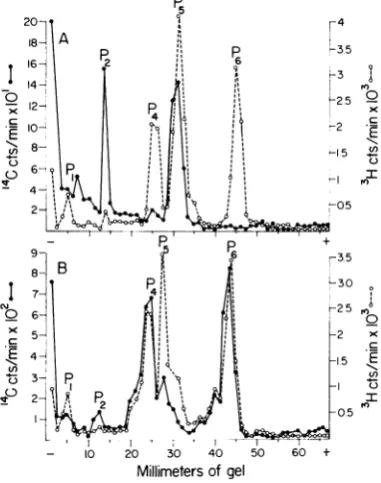
Figure 5 shows the electrophoretic profiles of
"C-proteins of fraction 13 (p _ 1.32) and 28
(p_1.20) of the CsClgradientdepicted in Fig. 4.
These proteins were extracted with and dialyzed
against0.5M urea aswell asacetic acid, SDS,and
2-mercaptoethanol. The nucleoprotein coils
col-lected from fraction 13 contained two primary
proteins that migrated in SDS-gels to the same
positionasP2 and P5ofthe3H-proteinsused as
9- 8- 7-la 6-)( 5-c .L) 2-
I-- -n-w
0----O
14C-aa
,~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
-- 2 t S i~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~I
-1.4 -1.3 -1.2 II II II II II II
4
8
12
16 20
2428 32 35
Top
Fractions
FIG. 4. Fractionation inCsCIdensitygradient of VS
virioncomponentsdegraded bydeoxycholate. Thevirus
was grown in L cells in the presence of3H-uridine
(10p.c/ml)and14C-aminoacids(1.13pAc/ml),purifiedby
differential, ratezonal andequilibrium centrifugation,
then disrupted with sodium deoxycholate, and mixed
with 4.5 ml of CsCl (startingspecific gravity, 1.30
mg/ml). Centrifugation was carried outfor 72 hrat
38,000 rev/minintheSW50rotor. Fractionsofabout 0.15ml each werecollectedfrom thebottom, and
50-,uliter samples were assayedfor density and
radioac-tivity. Symbols: 0, 3H(RNA); 0, 14C (protein).
616 J. VIROL.
3W-1I I'll
on November 11, 2019 by guest
http://jvi.asm.org/
[image:6.481.256.439.286.532.2]markers(Fig. 5A). There was no P6 and less than
107%
of P4 in the nucleocapsid fraction. Theslowest moving 14C peak could not be identified
and may be a polymer. In addition, a large
amount of "4C-protein did not enter the gel. These
results suggested that a considerable amount of
nucleocapsidproteinaggregated under the
condi-tions oftheexperiment.Previous experiments (10)
had shown that P2 is a polymer or aggregate of
oneorseveral ofthemajorproteins.
Figure5B reveals that, in contrast to the
nucleo-capsid, P4and P6 were thepredominant proteins
in deoxycholate-degraded virions that band at a
densityof 1.20g/ml. Although there was some P5 in the low-density fraction, it was markedly
re-duced compared to P5 present in intact virions
labeledwith 3I-amino acids.
Extraction with 8 M urea provided a more
critical test of the identity of the nucleocapsid
protein. The above experiments were repeated
with virions labeled with "4C-amino acids,
de-I
0
X
.E
I
4
0
n
CY,
0
C-9
to
X
.E_
In
II ,I
20 30 40 50
Millimeters ofgel
FIG. 5. Electropherograms of14C-proteins extracted
from VSvirions degradedwith deoxycholateand
frac-tionatedbyequilibrium centrifugation. The14C-proteins ofeach fraction were extracted withacetic acid, 0.5
mturea,SDS,and2-mercaptoethanolalong
with3H-pro-tein markersfrom intactpurified virions andwere
co-electrophoresed on 7.5% polyacrylamide gels in the presence of0.5 mt urea and 0.1% SDS. (A)
Deoxy-cholatefraction 13 (p 1.32g/ml) shown inFig. 4.
(B) Deoxycholatefraction 28 (p-1.20 g/ml) shown inFig.4.
0 .E
e0
- 10 20 30 40 50
Millimeters ofgel
60 +
FIG. 6. Electropherograms of 14C-proteins extracted
with 8mrureafromVSvirion componentsdegradedwith
deoxycholate andfractionated by CsCl density
cein-trifugationunderthesameconditionsasinFig.5except for theuseof 8Mureafor extraction ofproteins from
the two virion fractions. Each fraction was
electro-phoresedconcurrently on separate gels. Tlhe
positiolns
ofthepeaks
(P1,
P2, P4, P5,and P6) correspondto3H-proteins extractedfrom intact VS virions and run onthe same gels.Symbols: *,deoxycholatefraction of p-1.32g/ml; 0, deoxycholatefraction of p _ 1.19
g/ml.
gradedwithdeoxycholate, and
separated
byCsCI centrifugationinto twofractions ofdensities 1.32and 1.19 g/ml. Both fractions were extracted
with and dialyzed against 8 M urea as well as acetic acid, SDS, and
2-mercaptoethanol prior
to electrophoresis on separategels
along with 3H-protein markers extracted from intact virions.Figure 6 compares the electrophoretic profiles
of the two
deoxycholate-degraded
virionfractionssolubilized with 8 M urea.
Clearly,
P5 was thepredominant polypeptide in the
nucleocapsid
fraction, although about
15%l
ofrecoverable 'ICcounts still migrated with P2. Noother
proteins
weredetectedin thenucleocapsid. Incontrast, P4
andP6predominatedin the
deoxycholate
product that bandedatdensity1.19g/ml; residualP5wasalso present in the low-density fraction in an
amount equivalent to that in the
nucleocapsid
fraction but considerably less than that expected
inintact virions.
DISCUSSION
As previously reported (10), the molecular
weights of Indiana serotype VS virion proteins
were estimated as follows: P6-34,500, P5
59,500,P4_81,500, P3 140,000, P2
186,000,
and
P1
__ 275,000.
Similar values for four ofP6
on November 11, 2019 by guest
http://jvi.asm.org/
[image:7.481.253.446.67.244.2] [image:7.481.48.239.300.542.2]adifferent molecular weight, a finding which led to the suggestion that P6 is the surface protein (antigen) responsible for neutralization of each of the two viruses with only type-specific
antiserum (10). The studies reported herein lend
support tothis contention. P6 is the primary
pro-tein split off VS virions exposed to digitonin,
which presumably complexes with cholesterol in the virion envelope. Virions pelleted after
digi-tonin treatment had a correspondingly reduced
content ofP6 and appeared by electron
micros-copy tobe devoidofenvelopes and spikes.
How-ever, about 50 % ofP4 wasalso solubilized after
exposure of virions to digitonin. It seems
prob-able, therefore, that P4 is the structural protein nextmost accessible todigitonin and may
repre-sent the shell protein underneath the envelope.
Only insignificant amounts of P5 and the three
minor proteins were released from virions by
action ofdigitonin.
The experiments on degradation of VS virions
with deoxycholate revealed that 10 to
30%C
ofribonucleocapsid was released as determined by banding of fractionated virions in CsCl gradients
centrifuged toequilibrium. The nucleocapsid was identified on the basis of its high buoyant density,
its increasedratio of 3H-RNA to 14C-protein, and
visualization of its characteristic coils by
elec-tronmicroscopy.Essentially two proteins, P5 and
P2, were present in the nucleocapsid fraction
afterextraction in 0.5Murea.Acomparison of the
molecular weights of these two proteins raises
the question whether P2 is a polymer of P5,
possibly a trimer. Even after extraction with 8 M
urea, about 15 MC of the nucleocapsid protein
migrates inSDS-gels containing0.5 Mureain the
region of P2. The most likely interpretation of
these findingsis that the nucleocapsid proteinis
composed ofasinglepolypeptide,P5, which hasa
marked tendency to aggregate into polymeric
forms, particularly after purification. Such an
hypothesis is consistent with our earlier finding
thatP2present in intactvirionswassubstantially
reduced in amount by extraction in 8 M urea.
KangandPrevec (4) also identifiedthe same
pro-three structural components: an envelope with
spikes, an underlying "shell", and an internal
coiled nucleocapsid core. The studies on partial
degradation with digitonin and deoxycholate
in-dicate that the envelope protein isP6 and the core
protein is P5. The partialcontamination of both
solubilized envelope andseparated cores with P4
is consistent with the hypothesis that P4 is the
shellprotein which lies between the envelope and
thenucleocapsid. Data presented previously (10)
suggest that the minor proteins P2 and P3 are
aggregates or polymers of the major proteins.
The position and function ofthe large
polypep-tide,P1, areunclear.
ACKNOWLEDGMENTS
This investigation was suppor-ted by Public Health Service grant CA-10387 fr-om the National Cancer Institute and grant GB-6537Xfromthe National Science Foundation.
LITERATURECITED
1. de-Th6, G., 1967. Action de la digitonine sur les virions leucemogenesmurins. C. R. Acad.Sci.S6r.D264:2347-2349. 2. Devlin,T. M.,and A. L. Lehninger. 1958. Thepreparation
ofphosphor-ylatingsubfragmentsofratliver mitochondria
withdigitonin.J.Biol. Chem.233:1586-1588.
3. Huang, A. S.,J. W. Greenawalt,and R. R. Wagner. 1966. Defective Tparticlesof vesicularstomatitisvirus.1. Prepa-ration, morphology andsome biologic properties. Virol-ogy 30:161-172.
4. Kang, C.Y.,and L. Prevec. 1969. Proteins ofvesicular stomra-titis virus. I. Polyacrylamide gelanalysisof viralantigens. J. Virol.3:404-413.
5. Maizel, J. V., Jr. 1966. Acrylamide-gel electropherograml-s
by mechanical fractionation: radioactive adenovirus pro-teins. Science151:988-990.
6. McCombs, R. M., M. Benyesh-Melnick, and J. P. Brun-schwig. 1966. Biophysical studies of vesicular stomatitis virus. J. Bacteriol. 91:803-812.
7. Nakai,T.,and A. F. Howatson. 1968. The finestructureof
vesicularstomatitisvirus.Virology35:268-281.
8. Schnaitman, C., V.G. Erwin,and J. W.Greenawalt. 1967. Thesubmitochondrial localizationof monoamine oxidase. J. Cell Biol. 32:719-735.
9. Simpson, R. W.,aindR. E. Hauser. 1966. Structural com-ponentsof vesicularstomatitis virus.Virology 29:654-667. 10. Wagner, R.R.,T.A.Schnaitman,andR. M.Snyder. 1969. Structuralproteinsof vesicularstomatitis viruses. J. Virol. 3:395-403.