JOURNAL OF VIROLOGY, Mar. 1994, p. 1675-1681 Vol. 68,No.3 0022-538X/94/$04.00+0
CopyrightC) 1994,AmericanSocietyforMicrobiology
Mutations Which Affect
the
Inhibition of
Protein
Phosphatase
2A
by Simian Virus
40
Small-t
Antigen
In
Vitro Decrease
Viral Transformation
SHUBHANGEE MUNGRE,' KRISTINE
ENDERLE,'
BEN TURK,1 ANALIAPORRAS,'
YUN-QIWU,2 MARC C. MUMBY,2ANDKATHLEEN RUNDELL'*Departmentof Microbiology-Immunology and The CancerCenter, Northwestern University, Chicago, Illinois 60611-3008,l andDepartmentof Pharmacology, University of Texas
Southwestern Medical Center,Dallas, Texas 75235-90412 Received10September1993/Accepted3 December 1993
Threeindependent point mutationswithin residues 97 to103 ofthesimian virus 40 small-tantigen (small-t)
greatly
reduced theability
of purified small-t to inhibit protein phosphatase 2A invitro. These mutations affected the interaction ofsmall-tantigen withtheprotein phosphatase 2AAsubunit translatedinvitro,and apeptide fromtheregionidentified by these mutations released theAsubunit from immunecomplexes. When introduced into virus, the mutations eliminated the ability of small-t to enhance viral transformation of growth-arrested rat Flll cells.Incontrast, the mutant small-tantigens wereunimpairedinthetransactivation of theadenovirusE2promoter,anactivity which was reduced byadoublemutationinsmall-t residues43 and 45.The simian virus 40 (SV40) small-t antigen (small-t) en-hances the ability of the virus to transform many cell types. This is particularly evident when growth-arrested cells are assayed(3, 14). The mechanisms through which small-t affects cells were obscure for many years, but two biochemical ap-proaches for small-t have now been reported. The first involves theinteraction ofsmall-t withproteinphosphatase 2A(PP2A).
Cellularproteinswhich coimmunoprecipitate with small-t
an-tigenhave been studied for several years (10,21, 28, 32), and theidentification of theseasthecatalytic C and regulatory A subunits of PP2Aprovidedanessential cluetosmall-t function
(16, 29, 30). Byusing purifiedsmall-t and PP2A, it has been shown that this interaction results in an inhibition of PP2A activity against several substrates in vitro (31). Significantly, these substrates include the SV40 large-T antigen and the cellularp53protein (22).
It has also been shown that small-t can transactivate poly-merase II- and polymerase 111-directed transcription from a variety of promoters when cotransfected with reporter con-structs(13).The relevance of this observation to infection has been called intoquestion by the failure of small-t expressed from an adenovirus vector to activate transcription of the adenovirus E2 promoter (19). Small-t transactivation appears to require E2F-binding sites in the E2 promoter (12). One
possibility is that transactivation reflects alterations in the phosphorylation stateof E2F. PP2A-mediated inhibition of at least one transcription factor, CREB (27), has been docu-mented recently.
Mutational analysesof small-t antigen have focusedonthe
unique C-terminal half of the protein (1, 9, 15). Deletion analyses have clearly shown that this region is essential to small-t function. The C-terminal 102-amino-acid region is
cysteinerich and containstwoCysXCysXXCysclustersaswell
*Correspondingauthor. Mailingaddress: Departmentof
Microbi-ology-Immunology, Northwestern University, Olson 8450, 303 E. Chicago Ave., Chicago, IL 60611.Phone: (312)503-5923. Fax: (312) 503-1339.
as four isolated Cys residues (6, 25). Manypoint mutations introduced into theCys residues of thetwoclusters have been shown to produce highly unstable proteins (9), a property which correlates with the ability of small-ttobind zincions(7,
26). The severe instability of these mutant proteins made it impossible todetermine roles forindividual residuesin PP2A
inhibition, transactivation,ortransformation. We show in this report that mutations in the Cys clusters were unable to preventinteraction of small-t translated in vitro with the PP2A A subunit but that mutation of two isolated Cys residues upstreamfrom theCys clusterssignificantlyreduced
coprecipi-tation of theAsubunit with small-t. Inaddition, 50%inhibition of theactivityof the AC form of PP2Arequired5to10times more of the purified mutant proteins than of the wild-type
(WT) protein. These mutant small-tantigenswereunable to enhancetransformation when assayed by usingreconstructed viruses.
MATERIALSANDMETHODS
Cellsandviruses.African greenmonkeykidney (CV1)cells and Fischer rat F111 cells were propagated in Dulbecco modified Eagle medium containing 10% fetal bovine serum. All virusstockswereharvested fromlysates initiated with a low
multiplicity ofvirus, and thetiters weredetermined on CV1 cells.Intransformation assays, virusstockswereconcentrated
by centrifugation in an SW41 rotor for 3 h at 40,000 rpm. Pellets were suspended in a small volume of serum-free medium orphosphate-buffered saline, sonicated, and treated with chloroformto sterilize them.
Construction of mutant viruses and plasmids. Mutations were constructed in plasmid pw2t asdescribed previously (7,
17). In most cases, a derivative ofpw2t in which the HindIII site at SV40nucleotide5171 was convertedtoBglII wasused
(26). To constructviruses,mutantsequenceswerethen trans-ferred into pMK/SV40, a plasmid which contains the entire WTSV40 genome cloned via its EcoRI site into the
kanamy-cin-resistant plasmid pMK. ReplacementofVVT with mutant sequenceswas
accomplished
by using uniqueBglII
andBstXI 1675on November 9, 2019 by guest
http://jvi.asm.org/
1676 MUNGRE ET AL.
(nucleotide 4759) sites. Virus was isolated following digestion with EcoRI, ligation, and transfection of CV1 cells using DEAE-dextran. All plasmids and viral DNAs were sequenced through the transferred region to confirm the presence of the desired point mutation. For expression in bacteria, mutant small-t sequences were cloned into plasmid ptrc99A or pKK223-3 as described previously (7, 26). Full-length small-t antigen was expressed from ptrc99A, which also expressed minor amounts (20%) of a 14-kDa protein derived from internal initiation at amino acid 52 in bacteria. Only the 14-kDa formof small-t was expressed from pKK223-3, because small-t sequences were too far downstream from the plasmid ribosome binding site for the natural initiation codon to be used.
Purification of small-t from bacteria. Small-t was purified from solubilized bacterial debris fractions in the presence of urea andzinc ions as described previously (7, 26).
Invitrotranscription and translation. Following transfer of mutant sequences into pGEM3 (Promega), RNAs were tran-scribed by usingT7polymerase and translated by using reticu-locyte lysate kits (Promega). In some experiments, the TNT coupled translation system (Promega) wasused. When unla-beled small-t was made by in vitrotranslation, parallelreaction mixtures containing
[35S]methionine
were used to determine the quantities of small-t synthesized. Mutant small-t antigens were translated as efficiently as the WT in these experiments. Transcription vectors which express the PP2A A subunit (clone1P16)and its mutant derivative 514,which lacks the final two C-terminal repeats, were kindly provided by G. Walter
(Uni-versity of California at SanDiego)(20). Small-t purified from bacteria (10 to 200 ng per reaction mixture) or unlabeled small-t translated in vitro (5 to 10 ng) was incubated with
35S-labeledAsubunit(5 to10ng).After incubationonicefor 60min, immunecomplexeswerecollectedbyusing preformed
complexes of the monoclonal antibody PAb430 (8) bound to formalin-treated and heat-inactivated Staphylococcus aureus. Immune complexes werecentrifuged and washed three times RIPA buffer (20). Synthetic peptides supplied by the North-western University Biotechnology Service were used in com-petition experiments.Inthese, 10 ,ug of each peptide in200
RI
of RIPA bufferwas incubated with the washedcomplexesfor 30 min on ice. Peptides and any released A subunit were removed by three additional washes inRIPAbuffer.Radioac-tivity remainingwith small-t in immune complexes was mea-sured.
Transformationandtransactivation assays.Transformation of rat Flll cells to anchorage-independent growth was per-formed as described previously (9). Virus concentrated as described abovewasusedtoachievemultiplicitiesof infection of 100 to 200 PFU per cell. Transactivation assays were
performed by using pw2t-based plasmids and pE2CAT re-porter plasmids as described previously (9), except that
lipo-fectinwasused for transfection.
Inhibition of PP2Aby purified small-t antigen. Theeffects ofWT andmutant small-t onthe activity of the AC form of PP2A were determinedasdescribed previously (31). Purified PP2A(1nM)wasincubated for 30 min at4°C with the various concentrations ofpurified bacterially expressed small-t. PP2A activity was then measured by using phosphorylated myosin light chains (2 ,uM final concentration) as the substrate.
RESULTS
Coimmunoprecipitation of Asubunit with small-t antigen. Becauselarge-T antigen doesnotbindcellular PP2A A and C
subunits,it seemedlikely that the interaction of small-t antigen
a
b
c
. ;;
a:S iS ASS0:j:? 0 ff;t Sk:.00:0-:
00t:0:;C;tV:.t:-0-00000.t0:070.000...t0_,
00 tV REWog';:3.beX
sn 0oSSRtt
AiVE.;,S00Sf,A0000070A,AtS7S'f';
ff';A4:00j0000 ;'0f'; i. idAD'. i''''U'S,': 0,,,-S''0,,0: 'V0t'V00t' it00 ,;0",0"ff'','S,l0g; iV_L ;0 ;0:;; 000 0iS. 0000
_ .0f,'g' t: 000 '- ft '.E'i'',,5f',4
0X_r
:',foS:L.'00-'00i'1.''X0-at'd'
70 iV0,7 00000000'S'V0S't tV ,00".'_ 0000000: 0'S.0E','X'0e ta',, ft f.0;07X'''"f)
tt,,S,S,0i,',.':,.',l
've 72:jl,,l0fi.*0,00
ff0'S.'l.0.y' 0t,00 ;:
*',-v='r;-. ffXr..f;^.^ 00 :
': XiV 'd UR7
0. A.00:
X,;; ff
d
4:
:::
:E :~
4,:0 A:
FIG. 1. Translationproducts of small-t and the A subunit. Linear-izedplasmidsweretranscribed with T7polymerase invitro,and then theRNAwasused for translation inareticulocytelysate(Promega). Plasmid pGEM3t was linearized with BamHI outside the small-t coding regiontoproduce full-length 17-kDa small-t (lane a)orwith NdeItoproduceatruncatedderivativewhichcontainsamino acids1to
111of small-tantigen(lane b).The Asubunitsused in thisstudywere
provided by Gernot Walter in a Bluescript vector. The WT parent plasmid p16is shown inlane c,andthe deletion mutantis shown in lane d.
with PP2A is mediatedby thecysteine-rich unique region of small-tantigen.We(9)and others(1, 15) havestudied small-t
proteinswith mutations in this region,particularly those with mutations in thetwo CysXCysXXCys clusters.We have mon-itored PP2Ainteractionbyimmunoprecipitationwith infected cell extracts or by coimmunoprecipitation of the cellular
proteinswith small-tpurified from bacteria (2), but the pro-nouncedinstability of many of the mutantproteins of interest
(9)made itimpossibletodefine the effects of themutationson PP2Abinding.
In an attemptto eliminate theproblem ofprotein stability,
weturned to anassayusing proteins synthesized in vitro. The 61-kDaA subunit of PP2A orits mutantderivative (20) was transcribed from a Bluescriptvector and translatedby using
[35S]methionine.
Small-t WT and mutant sequences weretranscribed and translated from pGEM vectors. Products obtained from in vitroreactionsareshown inFig.1.Ingeneral,
50 ,ul of translation reactionsyielded 5 to 10 pmol of small-t
(morethan 80ng),while translation of theAsubunitwasless
efficient, yieldingabout 20ng per50-plA reaction mixture. Ininitialexperiments, quantitiesof small-tantigenrequired
forefficient interaction with translatedAsubunitwere deter-mined by using small-t antigen purified from bacteria as described previously (7, 26). Various amounts of purified
small-twereincubated onicewith 5 to 10ng of35S-labeledA subunit and then incubated withpreformedantibody-S.aureus complexes, washed withRIPAbuffer, and counted. Asshown inFig. 2, theAsubunitwasefficiently immunoprecipitated by small-tantigenover awideconcentration range.Aportion (20
to30%)of theAsubunit translated in vitrowasassociated with small-t antigen in a form which was resistant to stringent
washes with RIPA buffer. Although not shown here, gel
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
SMALL-t TRANSFORMATION AND PP2A INHIBITION 1677
E
QL
6000
5000
4000
30001
2000[
1000
200 66 33 20 10
[image:3.612.98.254.76.212.2]ng
FIG. 2. Coprecipitation of full-length and truncated PP2A A sub-units with small-tantigen. Plasmid 1P16 and its truncated derivative 514weretranscribedand translated in vitro by using[35S]methionine asdescribedin Materials and Methods. The translation mixtureswere
thendivided, andvarious concentrations of unlabeled small-t purified from bacteriawereaddedtoeachportion.Afterincubationonice, the
mixtureswereimmunoprecipitated with monoclonal antibodyPAb430.
Approximately 25% of the total translated A subunits was able to
interact with small-t antigen in theseassays.Backgroundradioactivity
(counts per minute precipitated in theabsence of small-t) was
sub-tracted from each sample. This was between 600 and 800 cpm in
variousexperiments. Closed symbols indicate precipitations with the full-length A subunit; open symbols indicate those with themutant
proteinwhich lacks the 14th and 15threpeatsof the A subunit.
electrophoresis confirmed that the radioactivity measured
re-flected theamountsofAsubunitpresent.
Reticulocyte lysate is known to contain PP2A activity (4). Consequently, it was possible that the interaction of small-t
withAsubunittranslated in vitrowasinfluenced by C subunit
presentin translation mixtures. Toexamine the role of the C subunit in our assay, we used a derivative of the A subunit
which lacks two of the C-terminal repeats known to be required for binding of the A subunit to the C subunit. As shown inFig. 2, precipitationofthemutantAsubunit required muchhigherconcentrations ofsmall-tantigen, suggesting that theinteraction of small-twiththeAsubunitis stabilized by the Csubunit when concentrations of small-t arereduced. Thus,
the affinity of the interaction between small-t and the A subunit alone may be relatively low, requiring either high
concentrations of small-t to drive the reaction equilibrium toward the complexed form or, alternatively, stabilization by
the C subunit.
Sequencesfromresidue 111arenotrequiredforinteraction withtranslated Asubunit. From theresults showninFig. 2, it is clear that it should be possible to measure interaction of
mutantsmall-t antigens byusing the quantities obtained from in vitro translation. Various mutant small-t sequences were,
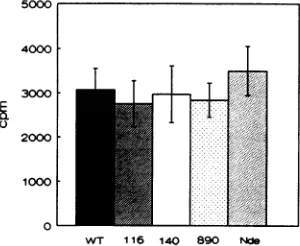
therefore, cloned into pGEM vectors for further study. Ini-tially,we were interested in mutations at Cys residues of the two Cysclusters. As shown in Fig. 3,mutationsat Cys-116or
Cys-140,in the first and secondclusters, respectively, didnot
affect the ability of small-tantigen to interact with A subunit translated in vitro. A similar result was obtained with the
in-frame deletion mutant d1890,which lackstwo Cys residues ofthefirstcluster (24).
ThepGEM plasmidwhich contained WT small-tsequences wasalsousedtoexpress atruncated small-tantigen.When the plasmid was cut at the NdeI site within small-t coding se-quences, invitro transcription-translation produced aprotein
which contained only the first 111 amino acids of small-t
5000
4000
3000
2000
1000
I
[image:3.612.361.511.78.201.2]WT 116 140 890 Nde
FIG. 3. Coprecipitation of A subunit withmutantsmall-tantigens. The PP2A A subunitwastranslatedin vitro by using[35S]methionine
and thendivided into portions. Translated small-t antigenswereadded
tothe radioactive Asubunit,andthen themixtureswereincubatedon
ice andprecipitated by using PAb430asdescribed in Materials and
Methods. Radioactivitypresentinimmunoprecipitates in the absence of small-t antigen (800 cpm)wassubtracted from the totalradioactivity
found in each sample. Small-t derivatives usedwere the WT,point
mutantCys-116- Ser (116), point mutantCys-140-->Ser(140), d1890 (890), and truncated small-t translated from pGEM3t cleaved with NdeIbeforetranscription (Nde). Bars,standard deviation.
antigen. Gel electrophoresis confirmed that no residual
full-length small-t could be detected followingtranslation(Fig.1). This truncated protein was fully capable of binding the A
subunit (Fig. 3), suggesting that sequences following amino acid 111 were not required in this assay. Missing sequences
containtwoCysresidues of the firstCys clusterand theentire second cluster.
Amino-terminal residues of small-t are not required. Ex-pression of small-t in bacteria results insynthesisofa 14-kDa
truncatedformresultingfrominternalinitiationatMet-52, in addition tothefull-length 17-kDaprotein.The 14-kDa deriv-ative is the only small-t-related protein expressed in some
derivativesofpKK223-3 in which theplasmidribosome bind-ingsite istoofarupstreamof the naturalinitiating codon (7). The truncated small-t has been purified by using the same
proceduresused topurifyWTsmall-t. As shown inFig. 4, the 14-kDaproteininhibits PP2A activityinvitroas efficientlyas
the full-length 17-kDa small-t expressed from the plasmid ptrc99Awhichwasused inmostofourstudies. Takentogether withresults obtainedby using the protein expressed following NdeI cleavage, these datasuggestthat small-t residues neces-saryfor PP2A interaction in the in vitro translation assayfall between amino acids 52 and 111.
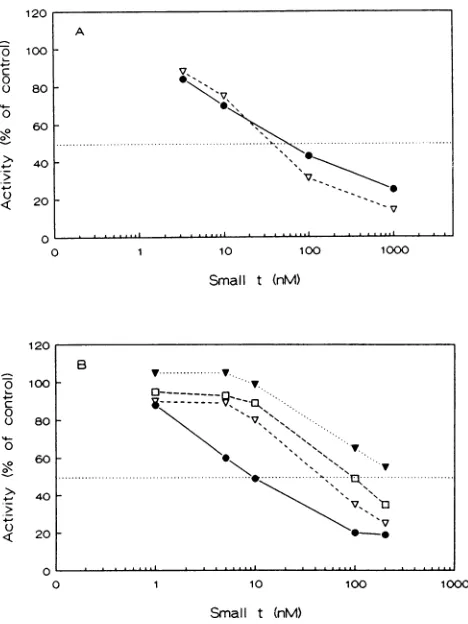
Mutations in the amino acid 97 to 103 region of small-t. Comparisonof the SV40small-tantigenwith those ofseveral relatedpapovaviruses(5, 6, 18)showedthattwoadditionalCys residues are conserved. These are at residues 97 and 103of SV40.In addition,aproline residue is locatedbetween these
two Cys residues in all papovaviruses. To study this region, individual mutationswereintroduced intoCys-97 (changedto Ser), Pro-101 (Ala), and Cys-103 (Ser). When these were
translated in vitro, the Cys-97 and Pro-101 mutant proteins
werefoundtobecompletelydefective in thebindingof theA subunit(Fig. 5A).TheCys-103 proteinshowedpartial binding, which was consistently less than that of the WT protein. Evidence that the amino acid 97 to 103 region (the 97-103 region) is directly involved in binding the A subunit in this
assaywas obtained by using synthetic peptides. As shown in Fig.SB,a peptidewithasequenceidentical toresidues 96 to 106 of the SV40 small-t antigen was able to release the A
--- ---
NNNNNNN
,I,
I
\I
VOL.68, 1994
-L
on November 9, 2019 by guest
http://jvi.asm.org/
0
c
0
00
>1
41
0
0
c 0 0
I.
>1
4J
120
100
80
60
40
20
0 10 100 1000
Small t (nM)
120 B
v.. V
100 v
0---80 V.
[image:4.612.63.297.74.386.2] [image:4.612.360.516.76.467.2]600
...,...
40-\ 'fV,
20 * -
--40
0 10
6000
5000
4000
a 3000 0
2000 .
1000
0
6000 5000
4000
E
Q 3000 0
2000
1000
0
6000
100 1000
Small t (nM)
FIG. 4. In vitro inhibition of PP2A by purified small-t antigen.
Bacterially expressed small-t antigens were purified as described
previously (7) and then used to analyze inhibition of phosphatase activity ofthe heterodimericAC form of PP2A (1 nM). Phosphory-lated myosin light chains were used as the substrate. Activity is
expressed as the percentage of the control activity obtained in the
absence of added small-t. (A) The activity ofthe 14-kDa derivative
(small-t amino acids 52 to 184) (open triangles) expressed from
pKK223-3 was compared to that of the full-length 17-kDa protein (closed circles) expressedfrom ptrc99A. (B) Full-length mutant
pro-teins expressed from ptrc99A were purified individually and then
compared with the WT protein (closed circles) in PP2A inhibition
assays. Mutant proteins used were Cys-97--Ser (open triangles),
Cys-103--Ser (open squares), andPro-101---Ala(closedtriangles).
subunit from immunoprecipitated complexes. The control peptides used contained residues 120to130 of small-tantigen (Fig. SB)oramutantformofthe 96-106 peptide in which Ser
wassubstituted for thetwoCys residues (Fig. SC). Thesewere
notabletoreleasethe A subunit.Althoughnotshownhere,a
peptidewhichcontained Cys-97but had aSer substitution at
103 also released Asubunit from immune complexes. This is consistent with the partial activity of the Cys-103 mutant in binding theAsubunit.
Effects of 97-103 mutations on PP2A activity. Purified
small-t antigen has been shown to inhibit PP2A in vitro (22, 31). To examine the effects of the mutations in the 96-106 region, each mutationwasintroduced into the bacterial
expres-sion vector ptrc99A. Following induction of bacteria and
solubilization ofsmall-t antigen usingurea in thepresenceof
zincions(7), monomeric small-twasobtained by gel filtration.
Small-t was further purified by using DEAE-cellulose and
hydroxylapatite chromatography as described previously (7,
26). As shown in Fig. 4, 10 nM WT small-t antigen inhibited
5000
4000
§ 3000
2000
1000
A
NT 103 97 101
- CYS SER
FIG. 5. Coprecipitation of A subunitby 97-103 mutant proteins
andeffectsofpeptidesfrom this region.Small-tmutantproteinsand
the PP2A Asubunitweretranslatedasdescribed in thelegendtoFig.
3and then used forcoprecipitationof the A subunitusingmonoclonal
antibodyPAb430. (A) 35S-labeled A subunitwas coprecipitatedwith
WT, mutantCys-103-Ser (103),mutant Cys-97--*Ser (97), and
mu-tant Pro-101-*Ala (101). (B) Washed immunecomplexes containing
radiolabeledAsubunit and WT small-tantigenwereincubated alone
(-),with 10,ugofpeptide96-106(96),orwithpeptide120-130(120)
for30min onice and thenwashed twice with RIPA buffer.
Radioac-tivityremainingwith the immunecomplexwasmeasured.(C)Immune
complexeswereincubated alone(-),withthe 96-106peptide (CYS),
orwith apeptideinwhich Cysresidueswere replacedby Ser(SER).
PP2Aactivity by 50%; 50% inhibition required 5to 10times
moreof the Cys-97 and Cys-103 mutantproteins. The small-t
mutant at Pro-101 was even more affected in its ability to
inhibit PP2A. None ofthe mutants istotally unableto inhibit PP2A, however, suggestingthat otherregions ofsmall-t may
interact with either the A orCsubunits of the enzyme.
Effect of 97-103 region mutations on transformation. The various mutations in the 97-103 regionwere also introduced
into intact virusesby using thepMK/SV40plasmid. Incontrast tomutations inCys residuesofthetwoCys clusters,themutant
small-t antigens produced in a short pulse appeared quite A
-v
... ...
--v
- 96 120
T
F
F
on November 9, 2019 by guest
http://jvi.asm.org/
SMALL-t TRANSFORMATION AND PP2A INHIBITION 1679
[image:5.612.105.253.77.129.2]a
b
cd
e
f
g
FIG. 6. Accumulated levels of small-t in mutant infections. F111 cellswereinfected with 20 total PFU per cell of WTvirus,amixture of WT anddl888, dl888 alone, or viruses with point mutations in small-t antigen. Cellswerelabeledwith
[135SJmethionine
for 20 hbeginningat30 h postinfection. Cell extracts were prepared by using nonionic detergent Nonidet P-40, and then small-t was recovered by immuno-precipitation. Patterns shown are the accumulated levels of small-t in cellsinfected withd1888(lane a), WT SV40 (lane b),WTSV40-d1888 (1:3) (lane c), mutant Cys-97-Ser (lane d), mutant Pro-101--sAla (lane e),mutant Cys-103-Ser (lane f), and WTSV40-dl888 (1:2) (lane
g).
stable in 1- to 3-h chase periods (not shown). However, the mutant proteins are slightly less stable than the WT, and the accumulated levels of small-t recovered from Cys-103 and Ala-101 aftera24-hlabelingperiodwerereducedabout three-tofivefold compared with theWTvirus.Toaccuratelymeasure the effects of these mutations on transformation ofrat Fll cells, it was important to define conditions in which similar large-T and small-t levelswere achieved following infections with themutant and WTviruses. Asshownin Fig. 6,levelsof accumulatedsmall-tantigenaregreaterin WTinfections than in mutant infections. However,when WTviruswasdiluted in stocks ofd/888 to maintain the total overall titer, equivalent amounts of small-t could be achieved in mutant and WT infections. The mixed infection maintains equivalent levels of large-Tantigen (not shown) while varying the levels ofsmall-t and allows the mutant viruses to be compared with the WT under identical conditions.
The mutantviruses were concentrated bycentrifugation to increase titers and then compared with WT and small-t deletion mutant viruses in transformation of rat cells to anchorage-independent growth.Asshown in Table 1,all three mutants were completely defective intransformation of these cells. Thedefectcannot beattributed tothe reduced levels of small-tantigen expressed by these viruses, because dilution of the WTvirus in thepresence ofthe deletion mutant did not reduce itsabilitytotransformratcellsdramatically.Also, when aliquots of the FIll cells used forplatingin agar were grown inculture for2days,levelsof mutant small-t antigens detected following anovernight pulsewere atleast equaltothose found inthe d1888-WT(3:1) mixed infections (not shown).
Effect of the 97-103 region mutations on transactivation.
TABLE 1. Transformation ofratFl 1 I cellsbymutantviruses
Virus(es) (ratio)" (meanColonies/dish±SD)"
WT... 1355 17
WT:d/888(1: 1) ... ... 99±13
WT:d1888 (1:3) ... ... 52 ± 25
WT:dl888 (1:5)... 45 ± 18
d1888...0 ±0
Cys-97...1 ± 1
Pro-l1l...1 ±I
Cys-103 ...0 ±0
a b c d e f
* 0s0;Ej* *
wt
FIG. 7. Transactivation bymutantsmall-tantigens.CVlcellswere
cotransfectedwith pw2tconstructs and pE2CAT by using lipofectin.
Cells were extracted and analyzed for chloramphenicol
acetyltrans-feraseactivityasdescribedpreviously (9).Thechromatograph shown
is representative of several experiments. The pw2t-based plasmids used for cotransfections with pE2CATexpressedWT small-t antigen (lane a) or one of the mutants Cys-103-->Ser (lane c). Cys-97-Ser (lane d), and Pro-101-Ala (lane e).A d1888 derivative ofpw2t(lane
f)wasused forasmall-tnegativecontrol. Theplasmidused for lane b
contained small-t antigen with a double mutation which changed
Pro-43to Leuand Lys-45toAsn.
Another known function of small-t antigen is its ability to
transactivate the adenovirus E2 promoter (12, 13, 19). To determine whether the Cys 97-103 region playedarole in this
activity, the mutations were cloned into plasmid pw2t for
cotransfectionwithaplasmidinwhichchloramphenicol
acetyl-transferase wasexpressed under the control of the adenovirus
E2promoter as in earlier studies (9, 19). As shownin Fig. 7,
mutations at Cys-97, Pro-101, and Cys-103 did not affect the ability of small-t antigen totransactivate the adenovirus
pro-moter,while mutation oftwoamino acidsatresidues 43 and 45
of small-t antigen reduced transactivating activity. The assays
shownin Fig. 7were runbeyond the linearrange tocompare
the 43-45 region mutation to the control lacking small-t. In experiments not shown here inwhich the total acetylation of chloramphenicol was less than 15%, transfection of plasmids
encoding the 97-103 regionmutantproteins resultedinatleast WTlevels oftransactivation. Inseveral experiments, levels of transactivation caused by the 97-103 region mutant proteins exceededeven that of WT small-t.
Mutations in the 43-45 regionwere made initially because
this region contains a hexapeptide which is completely
con-served in allpapovaviruses (5, 25). More recently, it has been shown thata4-amino-acidsequenceinthisregionisalso found inonePP2A B subunit (23) and that mutations in this region eliminatesmall-t transactivation of humanpapillomavirustype 16. Itisworthnoting that the first 51 aminoacidsof small-tare
absentina 14-kDa derivative which isfullyactive ininhibiting PP2A(Fig. 4).
DISCUSSION
Three mutations which alterinteraction of the SV40 small-t antigenwith PP2A in vitro and which failtotransformratcells
toanchorage-independent growth have been identified. These mutations fall inaregionof small-t which ispartiallyconserved in all papovaviruses. This region, amino acids 97 to 103 in SV40, is diagrammed below for SV40, the human BKvirus (BKV), mouse polyomavirus (PY), and hamster papovavirus (HaPV) (5, 6, 18, 25).ItcontainstwoCysresiduesseparated by five amino acidswhich containacentral proline residue:
SV40 Cys Lys Gln Trp Pro Glu Cys
BKV Cys Lys Glu Trp Pro Ile Cys
"Totalmultiplicityof infectionwas 100PFUper cell.
"Followinginfection of10"cells in suspension forI h, 6 x 104 cells were
platedperdish(eightdishes perinfection).Colonieswerecounted 6 weeks after
plating.
VOL. 68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.58.298.595.694.2]PY Cys Arg Met Pro Leu Thr Cys
HaPV Cys Arg Leu Pro Ile Thr Cys
These sequences lie within the unique region of the small-t antigen but upstream of the two CysXCysXXCys clusters whichappear to be involved in metal binding (7).
Whenpurifiedmutantproteinswereused to study inhibition of the enzymatic activity of the AC form of PP2A, it was clear that all three individual mutations reduced the affinity of small-t for PP2A. However, the mutant proteins did inhibit the enzyme when used at veryhigh concentrations, suggesting that this region may not beessential for PP2A interaction. All three mutantproteins were also defective in binding the A subunit synthesized in reticulocyte lysate, although the protein mutant atCys-103 consistently showed partial binding.
Studies with the peptide derived from the 97-103 region of small-t suggest that thisregionis a contact site between small-t and PP2A. Although the binding assay used radioactive A subunit, it is important to recognize that the 97-103 region of small-t may not be an A subunit-binding site. The assay is clearly dependent on stabilization by the C subunit, which is known to be present inreticulocyte lysates(4). This was shown byusing a mutant A subunit known to be unable to bind the C subunit. Our results based on use of this mutant indicate that interaction ofthe C subunit is necessary when small-t antigen is at low concentration. It is possible that the 97-103 region contactsthe C subunit, stabilizing the coprecipitation of the A subunit by small-t. Earlier studies (11) on the interaction of PP2A subunits showed that each of the subunits interacts directly with the other, although the stability of AB and BC complexes was less than that of the ABC enzyme. Stable complex formationrequired the presence of all three subunits, and the interaction ofBwith bothA and C was essential. In infected cells, small-tdisplaces the B subunit from complexes to form tAC (10). It seems likely that, as for the B subunit, there will be interactions between small-t and the C subunit. The 97-103 region may represent such a contact point.
Itis alsopossible that the 97-103 region represents one of several regions of small-t which contact the A subunit. A deletion analysis of the Asubunit has suggested that a rela-tively large region of the A subunit, possibly as long as 100 aminoacids, isrequired for interaction with small-t (20). This suggests that there could be more than one region of the small-t antigen participating in A subunit binding and that someof thecontactsbetweensmall-t and theAsubunit may be necessary not for initial binding but for high-affinity interac-tion.Recentevidence suggests thatatleastoneother region of small-t antigen is involved in A subunit binding. Sequences between amino acids 110 and 130, but not sequences from the 97-103 region, were required for binding to afusion protein which contains A subunit and glutathione-S-transferase se-quences (1Sa). Given the high concentrations of A subunit present in assays using the fusion protein, small-t sequences required for high-affinity interaction with the A subunit might notbe detected. The 97-103 region may be such a region.
Although the exact mechanism through which mutations within the 97-103 region reduce interaction of small-t and PP2Ais not completely understood, it is clear that the reduced affinity for PP2A in vitro correlates with reduced activity of small-t in enhancing transformation. It will be of interest to
study the effects of these mutations on intracellular proteins whosephosphorylation states change in the presence of small-t antigen during natural infection. Such substrates have not yet beenidentified, although recent studies suggest that a potential target of small-t following transfection is mitogen-activated
protein (MAP)kinase orits kinase(1Sa).
Transactivation of the adenovirus E2 promoter by small-t antigen may not depend on its ability to inhibit PP2A activity but may involve some mechanism other than controlling the phosphorylation state of specific transcription factors. The threemutations which reduced PP2Ainteraction had no effect on transactivation; in addition,mutation of residues 43 and 45 reduced small-t transactivation activity. Mutations of small-t in this region have also been shown to reducetransactivation of the human papillomavirus type 16 long control region in human cells (23). This region of small-t is not involved in inhibition of PP2A, because a 14-kDa derivative which com-pletely lacks the first 51 amino acids of small-t inhibits PP2A with the same efficiency as the WT protein. The mutations described in this study and several described earlier (9) also suggest that the transactivation activity of small-t is not suffi-cient for enhancement of transformation, as several viruses withmutant small-t antigens and normal transactivation capa-bility arecompletely unable to transform cells.
ACKNOWLEDGMENTS
This work was supported by Public Health Service grants CA-21327 (to K.R.) and CA-54726 (to M.C.M.). The support of the Lester Woods Foundation in providing equipment (to K.R.) is gratefully acknowledged.
REFERENCES
1. Bikel,I., H. Mamon, E. L. Brown, J. Boltax, M. Agha, and D. M. Livingston. 1986. The t-unique coding domain isimportant to the transformation maintenance function of the simian virus 40 small
tantigen. Mol. Cell. Biol. 6:1172-1178.
2. Bossert, A., P.Mulgaonkar, and K. Rundell. 1985. Interaction of simian virus 40 small-tantigen produced in bacteria with 56K and 32Kproteins of animal cells. J. Virol. 56:325-327.
3. Bouck, N., N. Bealer, T.Shenk, P. Berg, and G. di Mayorca. 1978. Newregion of simian virus 40 genome required for efficient viral transformation. Proc. Natl. Acad. Sci. USA 75:2473-2477. 4. Chen, S. C., G. Kramer, and B. Hardesty. 1989. Isolation and
partial characterization of an Mr 60,000 subunit of a type 2A phosphatase from rabbit reticulocytes. J. Biol. Chem. 264:7267-7275.
5. Delmas, V., C. Bastien, S. Scherneck, and J. Feunteun. 1985. A new member of thepolyomavirus family: the hamster papovavirus. Complete nucleotide sequence and transformation properties. EMBO J.4:1279-1286.
6. Friedman, T., R. Doolittle, and G. Walter. 1978. Amino acid sequencehomology between polyoma and SV 40 tumor antigens deduced from nucleotide sequence. Nature (London) 274:291-293.
7. Goswami, R. G., B. Turk, K. Enderle, A. Howe, and K. Rundell. 1992. Effect of zinc ions on thebiochemical behavior of simian virus 40small-t antigen expressed in bacteria. J. Virol. 66:1746-1751.
8. Harlow, E., L. V. Crawford, D. C. Pim, and N. M.Williamson. 1981. Monoclonal antibodies specific for simian virus 40 tumor antigens. J. Virol.39:861-869.
9. Jog, P., B.Joshi, V. Dhamankar, M. J. Imperiale, J. Rutila, and K. Rundell. 1990. Mutational analysis of simian virus 40 small-t antigen. J. Virol. 64:2895-2900.
10. Joshi, B., and K. Rundell. 1990. Association of simian virus 40 small-t antigen with the 61-kilodalton component of a cellular proteincomplex. J. Virol.64:5649-5651.
11. Kamibayashi, C., R. L. Lickteig, R. Estes, G. Walter, and M. C. Mumby. 1992. Expression of the A subunit of proteinphosphatase 2A andcharacterization of its interactions with the catalytic and regulatory subunits. J. Biol. Chem.267:21864-21872.
12. Loeken, M. R. 1992. Simian virus 40 small tantigen trans activates theadenovirus E2A promoter by using mechanisms distinct from those usedby adenovirusElA.J. Virol. 66:2551-2555.
13. Loeken, M., I. Bikel, D. M. Livingston, and J. Brady. 1988. Trans-activation of RNA polymerase II and III promoters by
on November 9, 2019 by guest
http://jvi.asm.org/
SMALL-t TRANSFORMATION AND PP2A INHIBITION 1681 SV40 small tantigen.Cell55:1171-1177.
14. Martin, R., V.Setlow, C. Edwards, and D. Vembu. 1979. Theroles ofthe simian virus 40tumor antigens intransformation of Chinese hamster lung cells. Cell 17:635-643.
15. Montano, X., R. C. Millikan, J. M. Milhaven, D. A.Newsome, J.W. Ludlow,A.K.Arthur, E. Fanning, I. Bikel, andD. M.Livingston. 1990. SV40 small t and anamino terminal domain of largeTshare
a common transforming function. Proc. Natl. Acad. Sci. USA 87:7448-7452.
15a.Mumby, M. Submitted for publication.
16. Pallas, D. C., L. K.Shahrik, B. L. Martin, S. Jaspers, T. B. Miller, D. L. Brautigan, and T. M. Roberts. 1990. Polyoma small and middle Tantigens andSV40 small t antigen form stable complexes withprotein phosphatase 2A. Cell 60:167-176.
17. Phillips, B., and K. Rundell. 1988.Failure of simian virus 40 small
tantigen to disorganize actin cables in nonpermissive cell lines. J. Virol.62:768-775.
18. Pipas, J. M. 1992. Common and unique features of Tantigens encoded bythepolyomavirus group. J. Virol. 66:3979-3985. 19. Rajan, P.,V.Dhamankar, K. Rundell, and B. Thimmapaya. 1991.
Simian virus 40 small-t does nottransactivateRNApolymerase II promoters in virusinfections. J. Virol. 65:6553-6561.
20. Ruediger, R., D. Roeckel, J. Fait, A.Bergqvist, G. Magnusson, and G. Walter.1992.Identificationof binding sites on the regulatoryA
subunit of protein phosphatase 2A for the catalytic C subunit and for tumorantigens of simian virus 40 andpolyomavirus. Mol. Cell. Biol. 12:4872-4882.
21. Rundell, K., E. 0. Major, and M. Lampert. 1981.Association of cellular 56,000- and 32,000-molecular-weight proteins with BK virus and polyoma virus t-antigens. J. Virol. 37:1090-1093. 22. Scheidtmann, K. H., M. C. Mumby, K. Rundell, and G. Walter.
1991. Dephosphorylation of simian virus 40 large-T antigen and p53 protein by protein phosphatase 2A: inhibition by small-t antigen. Mol. CellBiol.11:1996-2003.
23. Smits, P. H. M., H. L. Smits, R. P. Minnaar, B. A. Hemmings, R. E.
Mayer-Jaekel, R. Schuurman, J. van der Noordaa, and J. ter
Schegget. 1992. The 55 kDa regulatorysubunit ofprotein phos-phatase 2A plays a role in the activation of the HPV16 long control region in human cells withadeletion in the shortarmof chromosome 1 1.EMBO J. 11:4601-4606.
24. Thimmapaya, B., andT.Shenk. 1979.Nucleotide sequence anal-ysis of viable deletionmutantslacking segments of the simian virus 40genome coding for smalltantigen. J. Virol. 30:668-673. 25. Tooze, J.(ed.). 1981. Molecularbiology of the tumor viruses,2nd
ed. ColdSpring Harbor Laboratory, ColdSpring Harbor, N.Y. 26. Turk, B., A. Porras,M.C.Mumby, andK. Rundell. 1993.Simian
virus 40small-t antigen bindstwozinc ions. J. Virol.67:3671-3673. 27. Wadzinski,B.E.,W. H.Wheat,S.Jaspers,L.F.Peruski, Jr.,R. L.
Lickteig, G.L.Johnson,andD.J.Klemm. 1993.Nuclearprotein phosphatase 2A dephosphorylates protein kinase A-phosphory-lated CREB andregulates CREBtranscriptional stimulation.Mol. Cell. Biol. 13:2822-2834.
28. Walter, G., A. Carbone-Wiley, B. Joshi, and K. Rundell. 1988. Homologous cellular proteins associated with simian virus 40
small t antigen and polyomavirus medium T antigen. J. Virol. 62:4760-4762.
29. Walter, G., F. Ferre, 0. Espiritu, and A. Carbone-Wiley. 1989. Molecular cloning and sequencing of cDNA encoding polyoma medium tumor antigen-associated 61-kDa protein. Proc. Natl. Acad. Sci. USA 86:8669-8672.
30. Walter, G., R. Ruediger, C. Slaughter, and M. Mumby. 1990. Association ofprotein phosphatase 2A with polyoma virus
me-diumTantigen.Proc. Natl.Acad. Sci. USA87:2521-2525. 31. Yang, S.-I., R. L. Lickteig,R.Estes, K. Rundell, G.Walter,and
M.C. Mumby. 1991.Control of protein phosphatase 2A by simian virus 40 small-tantigen. Mol. Cell. Biol. 11:1988-1995.
32. Yang, Y.-C., P. Hearing, andK. Rundell. 1979.Cellularproteins associated with simian virus 40 early gene products in newly infectedcells. J. Virol. 32:147-154.
VOL. 68, 1994