Copyright © 2001, American Society for Microbiology. All Rights Reserved.
Characterization of the AdoMet-Dependent Guanylyltransferase
Activity That Is Associated with the N Terminus of Bamboo
Mosaic Virus Replicase
YI-IJA LI, YI-JUN CHEN, YAU-HEIU HSU,ANDMENGHSIAO MENG*
Graduate Institute of Agricultural Biotechnology, National Chung Hsing University, Taichung, Taiwan 40227, Republic of China
Received 7 February 2000/Accepted 15 October 2000
Bamboo mosaic virus (BaMV), a member of the potexvirus group, infects primarily members of the
Bambusoideae. Open reading frame 1 (ORF1) of BaMV encodes a 155-kDa polypeptide that has long been
postulated to be a replicase involved in the replication and formation of the cap structure at the 5ⴕend of the viral genome. To identify and characterize the enzymatic activities associated with the N-terminal domain of the BaMV ORF1 protein, the intact replicase and two C-terminally truncated proteins were expressed in
Saccharomyces cerevisiae. All three versions of BaMV ORF1 proteins could be radiolabeled by [␣-32P]GTP,
which is a characteristic of guanylyltransferase activity. The presence ofS-adenosylmethionine (AdoMet) was essential for this enzymatic activity. Thin-layer chromatography analysis suggests that the radiolabeled moiety linked to the N-terminal domain of the BaMV ORF1 protein is m7GMP. The N-terminal domain also exhibited
methyltransferase activity that catalyzes the transfer of the [3H]methyl group from AdoMet to GTP or
guanylylimidodiphosphate. Therefore, during cap structure formation in BaMV, methylation of GTP may occur prior to transguanylation as for alphaviruses and brome mosaic virus. This study establishes the association of RNA capping activity with the N-terminal domain of the replicase of potexviruses and further supports the idea that the reaction sequence of RNA capping is conserved throughout the alphavirus-like superfamily of RNA viruses.
Bamboo mosaic virus (BaMV), a member of potexvirus group, has a plus-strand RNA genome (⬃6.4 kb) with a 5⬘ m7G(5⬘)ppp(5⬘)G cap structure and a 3⬘poly(A) tail (18). The
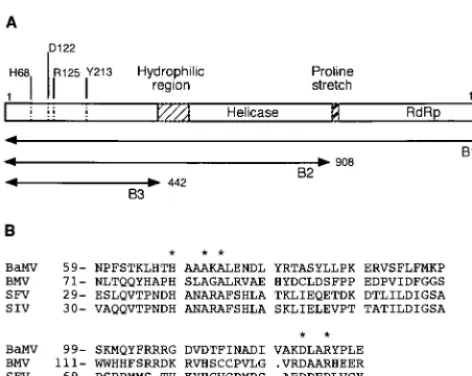
4.1-kb open reading frame 1 (ORF1) gene of BaMV encodes a 155-kDa polypeptide (17). The presence of conserved motifs such as GKS and GDD, signatures of helicase (6, 10) and polymerase (6, 13), respectively, led to the prediction that the 155-kDa protein may have an RNA helicase activity in the middle region and a polymerase activity in the C-terminal domain. The RNA-dependent RNA polymerase activity of the 155-kDa viral protein was recently corroborated by showing that the Escherichia coli-expressed viral protein was able to synthesize complementary RNA molecules using 3⬘end frag-ments of either the plus or minus strand of a BaMV RNA transcript as a template and that this polymerase activity was abolished when the GDD motif was deleted (16). The N-terminal region of the ORF1 product of potexviruses also shows distant similarity to the putative Sindbis virus-like meth-yltransferase (24), suggesting that the N terminus of the BaMV 155-kDa protein may be responsible for the cap formation at the 5⬘ends of genomic and subgenomic RNA transcripts. Fig-ure 1 shows the assumed domain organization of the 155-kDa BaMV replicase and the conserved His, Asp, and Arg residues found in the Sindbis virus-like methyltransferase.
The 5⬘-terminal cap structure m7G(5⬘)pppN is a
character-istic of eukaryotic mRNAs that is required for translation and stability. The formation of the cap requires three consecutive enzymatic activities according to the studies of capping reac-tions in mammalian and Saccharomyces cerevisiae cells and several viral systems. The proposed reaction scheme is sum-marized as the following (21, 28):
␥⬘p⬘p␣⬘pN(pN)n ORNA 5⬘-triphosphatase¡⬘p␣⬘pN(pN)n⫹pi
(1)
⬘p␣⬘pN(pN)n⫹␥pp␣pG Oguanylyltransferase¡
G␣p⬘p␣⬘pN(pN)n⫹ppi (2)
(The reaction at step 2 can be divided further into two half-reactions: enzyme⫹␥pp␣pG3enzyme-␣pG⫹ppi and
en-zyme⫺␣pG⫹⬘p␣⬘pN(pN)
n3G␣p⬘p␣⬘pN(pN)n⫹enzyme.)
G␣p⬘p␣⬘pN(pN)n⫹AdoMet Omethyltransferase¡
m7G␣p⬘p␣⬘pN(pN)n⫹AdoHcy (3) where AdoMet is S-adenosylmethionine and AdoHcy is S -adenosylhomocysteine. RNA 5⬘-triphosphatase removes the 5⬘ ␥phosphate of nascent mRNA (step 1), after which mRNA guanylyltransferase donates a GMP moiety, derived from GTP, to form a 5⬘–5⬘ triphosphate linkage, typical for a cap structure (step 2). In the step 2 reaction, the GMP moiety is linked first to the enzyme as a covalently bound intermediate (the first half-reaction) and then transferred to the 5⬘
diphos-* Corresponding author. Mailing address: Graduate Institute of Ag-ricultural Biotechnology, National Chung Hsing University, 250 Kuo-Kuang Rd., Taichung, Taiwan 40227, Republic of China. Phone: 886-4-2840328. Fax: 886-4-2853527. E-mail: [email protected] .tw.
782
on November 9, 2019 by guest
http://jvi.asm.org/
phate terminus of the RNA molecule (the second half-reac-tion). Thereafter, the cap is methylated by AdoMet at position 7 of the terminal guanosine, yielding a cap 0 structure and AdoHcy (step 3). A variety of RNA viruses have evolved di-verse pathways for cap formation (29). For example, GTP is methylated before the transguanylation reaction in alphavi-ruses (1) and brome mosaic virus (4, 12).
Because the capping of cellular mRNAs is a nuclear func-tion, it is not accessible to cytoplasmic viruses. Thus, BaMV, replicating in the cytoplasm of host cells, should be equipped with its own mRNA capping system. It was also interesting to know whether the capping activity of BaMV has characteristics similar to those of alphaviruses and brome mosaic virus since these viruses all belong to the large alphavirus-like superfamily of RNA viruses. To test the hypothesis that the N-terminal domain of the 155-kDa polypeptide harbors RNA capping activity, the ORF1-encoded polypeptide was expressed, in dif-ferent lengths, in Saccharomyces cerevisiaeand the activities associated with the viral proteins were characterized. The re-sults show that the N-terminal domain of the 155-kDa viral protein indeed harbors an AdoMet-dependent guanylyltrans-ferase activity. The viral protein forms a covalent complex with m7GMP as for alphaviruses and brome mosaic virus. This
discovery further supports the idea that the order of the cap-ping reaction is conserved among members of the alphavirus-like superfamily, although there are limited sequence similar-ities among them.
MATERIALS AND METHODS
Chemicals.Nucleotides and nucleotide analogs such as GTP, dGTP, GDP, GMP, m7GMP, and guanylylimidodiphosphate (GIDP) were purchased from
Sigma. AdoMet was from Boehringer Mannheim, and32P-labeled nucleotides
and Ado[methyl-3H]Met were from NEN.
Plasmid construction.Plasmid pYES2 (Invitrogen) was used to carry the BaMV cDNAs for viral protein expression inS. cerevisiae INVSc1 (MAT␣
his3-⌬1 leu2 trp1-289 ura3-52). Primer pair 5⬘-CCCAAGCTTATGGCACTCGT TTCTAAAGTCTTTGACand 5⬘-GCCAGATCTAGAGAGTAGGTCAGTTAT CCG was used to amplify the full-length cDNA of BaMV ORF1 (4,095 nucle-otides [nt]) in a 50-l PCR mixture that contained 1 ng of pBL (carrying the full-length cDNA of BaMV), 0.32M (each) primer, 0.2 mM (each) deoxyribo-nucleotide, and 2.5 U ofPfupolymerase. Sequences AAGCTT and TCTAGA represent the engineered cutting sites ofHindIII andXbaI, respectively, and the sequences in italics are within the coding region of BaMV ORF1. The PCR was carried out for 30 cycles (94°C for 45 s, 65°C for 45 s, 72°C for 8 min), followed by a 10-min extension at 72°C. Another set of primers, 5⬘-CCCAAGCTTATG GCACTCGTTTCTAAAGTCTTTGACand 5⬘-GCTCTAGAGCTCATTTGGGC TCCAAGGGTTCATC, was used to amplify a 3⬘-end-deleted BaMV ORF1 (2,724 nt) that encodes both the putative methyltransferase and helicase do-mains, whereas 5⬘-CCCAAGCTTATGGCACTCGTTTCTAAAGTCTTTGAC
and 5⬘-GCTCTAGAGCTCATTCGGTAGTTGCTGCGTCTGTwere used for the amplification of an even shorter 3⬘-end-deleted fragment of ORF1 (1,326 nt) that encodes only the putative methyltransferase domain under the PCR condi-tions described above. The PCR-amplified cDNAs were then digested with
HindIII andXbaI, ligated with pYES2, and transformed intoE. coliTop10F (Invitrogen) to obtain expression vector pYEB1 (containing a 4,095-nt cDNA), pYEB2 (containing a 2,724-nt cDNA), and pYEB3 (containing a 1,326-nt cDNA). The expressed viral proteins encoded by plasmids pYEB1, pYEB2, and pYEB3 are abbreviated B1 (1,365 amino acids), B2 (908 amino acids), and B3 (442 amino acids), respectively (Fig. 1).
Plasmid pET29 (Novagen) was used for the expression of the N-terminal domain of the BaMV ORF1 product inE. coli. Primer pair 5⬘-GTGCGGCAC ATATGGCACTCGTTTCTAAAGTCTTTGACand 5⬘-CTTGCGAAGCTTAAG CCGCTTTGCATTCTGGTwas used to amplify a 1,449-nt cDNA fragment con-taining the 5⬘end of BaMV ORF1 in a PCR. CATATG and AAGCTT represent the engineered cutting sites ofNdeI andHindIII, respectively. The sequences in italics are within the coding region of BaMV ORF1. The conditions of PCR were as described above. The endoribonuclease-digested cDNA fragment was inserted into pET29 and transformed into E. coli BL21(DE3). This construct (pEBM2984) allows the production of the N-terminal portion (the first 483 amino acids) of the 155-kDa protein inE. colicells.
Expression of recombinant viral proteins in yeast cells.To express the viral proteins in yeast cells, plasmids pYEB1, pYEB2, and pYEB3 were transferred separately intoS. cerevisiaecells by electroporation (pulse controller; Bio-Rad) under conditions of 1.5 kV, 25F, and 200⍀. The transformed yeast cells were selected by growing them in glucose-containing SC agar (2% glucose, 0.67% yeast nitrogen base without amino acids, 0.2% Kiwibrew solution, 2% agar). The absence of uracil in SC agar provides a selective pressure for the cells carrying plasmid derivatives of pYES2. The acquisition of the desired plasmid by yeast cells was confirmed by the amplification of the respective cDNA fragment by PCR.
To induce the BaMV proteins in yeast, recombinant cells grown in 10 ml of glucose-containing SC medium were pelleted down, washed, and resuspended in 10 ml of galactose-containing SC medium (2% galactose). The cultivation was continued for another 24 h at 30°C. The harvested pellets were suspended in 0.25 ml of ice-cold protein extract buffer (40 mM Tris [pH 8.0], 80 mM NaCl, 1.6 mM EDTA, 4 mM dithiothreitol [DTT], 4 mM EGTA, 4 mM phenylmethylsulfonyl fluoride), and disrupted at 4°C by vigorous mixing (10 times for 30 s each) with 0.45 g of acid-treated glass beads (0.3 to 0.45 mm) in a 1.5-ml Eppendorff tube. The cell extract was then subjected to centrifugation at 18,000⫻gfor 10 min at 4°C. The pellet fraction (P18) was further fractionated by sucrose discontinuous-gradient centrifugation. A 2-ml sample, which contained 60% (wt/wt) sucrose, was first overlaid on top of a 1.3-ml 67% sucrose solution in an SW41Ti (Beck-man) ultracentrifuge tube. Then 7.1- (50%) and 1.6-ml (10%) sucrose solutions were laid onto the tube. All sucrose solutions contained 100 mM NaCl and 50 mM Tris (pH 7.5). Centrifugation (35,000 rpm) was carried out at 4°C for 20 h. Preparation of antibodies against the N-terminal portion of the BaMV ORF1 product.The N-terminal domain of the 155-kDa viral protein was produced by
E. coliBL21(DE3) cells harboring pEBM2984 under conditions described pre-viously (16). The insolubleE. coli-expressed truncated protein was thereafter purified by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-FIG. 1. (A) Domain organization of the replicase (1,365 amino
acids) of BaMV. A highly hydrophilic region and a proline-rich stretch are thought to divide the 155-kDa replicase into methyltransferase, helicase, and RNA-dependent RNA polymerase (RdRp) domains. B1, B2 (908 amino acids), and B3 (442 amino acids) represent the intact BaMV replicase and two C-terminally truncated proteins expressed in yeast in this study. The conserved residues (His, Asp, Arg, and Tyr) of the Sindbis virus-like methyltransferase found in the N terminus of the BaMV replicase are indicated. (B) Alignment of partial sequences of the N termini of BaMV replicase, 1a protein of brome mosaic virus (BMV), and nsP1 proteins of Semliki Forest virus (SFV) and Sindbis virus (SIV). Asterisks, conserved residues including His, Asp, and Arg. The association of mRNA capping activities with nsP1 of Semliki Forest virus and Sindbis virus and 1a of brome mosaic virus has been established (2, 4, 12, 14, 19).
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.58.294.71.259.2]PAGE). The electroeluted protein from the corresponding protein band was then injected into rabbits to raise antibodies. The antibodies were purified by passing the rabbit serum through a DEAE Affi-Gel blue gel column (Bio-Rad). Immunoprecipitation and Western blotting.For immunoprecipitation, the SDS-denatured sample of the guanylyltransferase assay was diluted 20-fold with TET buffer (1% Triton X-100, 50 mM Tris [pH 7.5], 150 mM NaCl, 5 mM EDTA) and then mixed with antibodies conjugated to CNBr-activated Sephadex beads. After 2 h of gentle mixing, the beads were washed six times with TET buffer and subjected to SDS-PAGE analysis. The antibodies, which are specific for the N terminus of the ORF1 product of BaMV, were also used in Western blot analysis.
Activity assay.The guanylyltransferase activity was detected by the formation of32P-labeled proteins after incubation of proteins with [␣-32P]GTP (9, 27).
Unless otherwise stated, the standard reaction was carried out at 30°C for 1 h in a 20-l reaction buffer that contained 5l of enzyme preparation, 10Ci of [␣-32P]GTP (3,000 Ci/mmol), 50 mM Tris (pH 7.5), 5 mM DTT, 2 mM MgCl
2,
10 mM KCl, 1.2%n-octyl--D-glucopyranoside, and 100M AdoMet. The re-action was stopped by adding SDS (final concentration, 2%) and was followed by 3 min of boiling. The reaction products were analyzed by SDS-PAGE and visualized by autoradiography or phosphorimager.
The methyltransferase activity was detected by the transfer of [3H]methyl from
Ado[methyl-3H]Met to a methyl acceptor (14). The reaction was carried out at
30°C for 50 min in a 25-l solution that contained 50 mM Tris (pH 7.5), 2 mM MgCl2, 2 mM DTT, 1.2%n-octyl--D-glucopyranoside, 10M AdoMet, 0.75Ci
of Ado[methyl-3H]Met (80 Ci/mmol), 10l of enzyme solution, and 10 mM
methyl acceptor. At the end of the reaction, 1 ml of 10 mM ammonium acetate (pH 8.5) was added, and the reaction products were adsorbed to 1 ml DEAE-Sephadex in a Pasteur pipette, washed with the same buffer containing 100 mM NaCl, and then eluted with the same buffer containing 500 mM NaCl. The incorporated3H on the methyl acceptor was measured by liquid scintillation.
The covalent linkage of [3H]methyl to BaMV protein was performed at 30°C
for 20 min in a 30-l solution that contained 50 mM Tris (pH 7.5), 10 mM KCl, 2 mM MgCl2, 5 mM DTT, 1.2%n-octyl--D-glucopyranoside, 5Ci of Ado
[methyl-3H]Met (80 Ci/mmol), and 10l of enzyme solution. To determine the
effect of GTP on methylation of proteins, 100M GTP was included in the reaction mixture. The reaction was stopped by adding SDS to a final concentra-tion of 2% and was followed by boiling for 3 min. The sample was analyzed by SDS-PAGE in a 10% polyacrylamide gel, and products were visualized by fluo-rography.
Thin-layer chromatography (TLC) analysis of guanylate moiety.The reaction products of the guanylyltransferase assay were separated by SDS-PAGE, and the
32P-labeled proteins were eluted from the gel and concentrated. The protein
samples were then incubated at 65°C for 10 min with 0.5 N HCl or 0.5 N NaOH or incubated at 37°C for 20 min with 3.8 M acidic hydroxylamine (pH 4.3) as described by Jose et al. (9). The treated samples were then spotted on a poly-ethylenimine cellulose plate, developed with formic acid buffer (0.5 M formic acid, 0.5 M LiCl2), and visualized by autoradiography.
RESULTS
Viral protein expression in yeast cells.In order to study the inherent activities associated with the N-terminal region of the BaMV 155-kDa protein, we constructed three yeast-expression vectors in this study. Plasmid pYEB1 encodes the entire 155-kDa replicase (B1), while pYEB2 and pYEB3 encode C-ter-minally truncated proteins with deduced sizes of 100 (B2) and 50 kDa (B3), respectively. The positions of truncation were based on the assumption that the long hydrophilic stretch be-tween amino acids 406 and 520 and the proline-rich segment from amino acid 895 to 910 may represent the borders of functional domains. The expression of B1, B2, and B3 proteins in yeast was determined by Western blotting analysis (Fig. 2). After centrifugation of cell extracts at 18,000⫻g, B1 and B2 were found in both soluble (S18) and insoluble (P18) fractions, while B3, the putative methyltransferase domain, was present mostly in the P18 fraction.
Detection of guanylyltransferase activity.The formation of the covalently bound GMP-enzyme complex was used as an index for guanylyltransferase activity. In the initial trial we
incubated protein samples with [␣-32P]GTP with or without
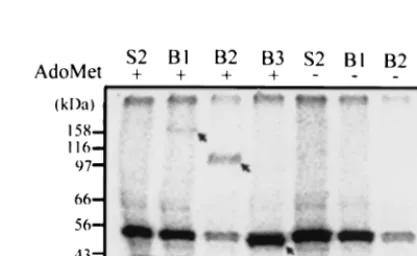
AdoMet and analyzed the reaction products by denaturing SDS-PAGE. The results indicate that B1, B2, and B3 proteins from the pellet fraction (P18) formed covalently bound com-plexes with [32P]GMP (Fig. 3). The proteins from the
super-natant fraction (S18), however, did not show the corresponding bands (data not shown). The complex formed only when AdoMet was included in the reaction buffer. The band that migrated slightly above the B3 protein and that appeared in every sample including S2 (the yeast background control) was suspected to be the cellular mRNA guanylyltransferase of yeast (53 kDa) according to its apparent molecular mass (26). The labeling reaction of the yeast cellular mRNA guanylyl-transferase does not require AdoMet. Therefore, the AdoMet dependence of activity distinguishes BaMV guanylyltrans-ferase from the yeast counterpart. In order to alleviate the interference of the yeast enzyme in the activity assay, we fur-ther purified the BaMV protein in the P18 fraction with a discontinuous sucrose gradient. Western blotting analysis of
[image:3.612.326.535.514.642.2]FIG. 2. Expression of the BaMV replicase (B1) and two C-termi-nally truncated proteins (B2 and B3) inS. cerevisiae. Yeast cells har-boring the desired plasmid were disrupted, and the supernatant (S18) and pellet (P18) after centrifugation at 18,000⫻gwere analyzed by SDS–10% PAGE. The BaMV proteins were detected by rabbit anti-bodies directed against the N-terminal portion of BaMV replicase. S2, extracted proteins from yeast cells harboring plasmid pYES2 (Invitro-gen).
FIG. 3. AdoMet-dependent guanylyltransferase activity. The yeast-expressed BaMV proteins from the pellet fraction (P18) were incu-bated with [␣-32P]GTP under the conditions described in Materials and Methods and analyzed by SDS-PAGE. The viral proteins (B1, B2, and B3) were radiolabeled only when AdoMet (100M) was included in the reaction buffer (arrows). The band immediately above B3 and shown in every lane probably represents the yeast cellular mRNA guanylyltransferase (53 kDa). S2, proteins from the P18 fraction of yeast cells harboring plasmid pYES2.
on November 9, 2019 by guest
http://jvi.asm.org/
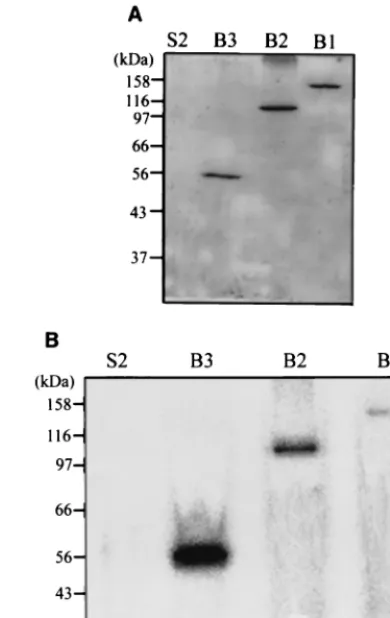
the fractionated samples (B3 in this case) showed that a por-tion of the viral protein floated to the interface between the 10 and 50% sucrose solution (Fig. 4), suggesting that this part of the viral protein is membrane associated. Similar to the B3 protein, the B1 and B2 proteins could also be found in the membrane fraction (Fig. 5A). All three membrane-associated viral proteins exhibited guanylyltransferase activity only in the presence of AdoMet, and obviously B3 had stronger activity than B2 and B1 (Fig. 5B). The result also shows that the background activity caused by the yeast protein was alleviated, so the membrane fraction was used in a subsequent guanylyl-transferase assay. To confirm the 32P-labeled protein as a
BaMV protein, the reaction products of the guanylyltrans-ferase assay were immunoprecipitated using antibodies against the N-terminal portion of the BaMV replicase and then the labeling of B2 and B3 by [␣-32P]GTP was examined by
SDS-PAGE analysis (Fig. 6). The requirement of AdoMet to form the covalent32P-labeled proteins was observed again. No yeast
cellular capping enzyme was found after immunoprecipitation. The aforementioned data show that the N-terminal portion of the BaMV replicase indeed harbors a guanylyltransferase tivity and that the presence of AdoMet is crucial for this ac-tivity.
Characteristics of BaMV guanylyltransferase activity.Other nucleoside triphosphates including [␣-32P]ATP, [␣-32P]UTP,
and [␣-32P]CTP were used to verify the nucleotide selection in
the guanylyltransferase assay. The results showed that both B2 and B3 were labeled by [␣-32P]GTP but not by either
[␣-32P]ATP, [␣-32P]UTP, or [␣-32P]CTP (Fig. 7). The time
course of the32P labeling of the viral proteins showed that the
extent of labeling increased with the incubation time and reached a plateau after 30 min (data not shown). The activity was also dependent on the concentration of AdoMet; the more AdoMet, the stronger was the activity (data not shown). EDTA
(5 mM) abolished the activity, suggesting that Mg2⫹is crucial.
AdoHcy (100M) or pyrophosphate (5 mM) was also included in the reaction buffer to determine the effects on the product. The result showed that AdoHcy was unable to stimulate the activity and, in fact, that it had a slightly inhibitory effect against the function of AdoMet. On the other hand, pyrophos-phate (5 mM) annihilated transguanylation completely (data not shown). This suggests strong binding of pyrophosphate to the enzyme, and the loss of activity may be caused by the competition between pyrophosphate and GTP binding.
TLC analysis of the guanylate moiety.The AdoMet depen-dence of guanylyltransferase activity could have different im-plications. AdoMet could act as an effector that induces pro-tein conformational changes and stimulates transguanylation. Alternatively, GTP could be methylated first by AdoMet and subsequently transferred as an m7GMP moiety onto the active
[image:4.612.103.246.71.255.2]site of the enzyme as for alphaviruses and brome mosaic virus. Two approaches were employed to clarify this uncertainty. In the first attempt, we looked into whether the methyl group
FIG. 4. Partition of B3 protein in a discontinuous sucrose gradient. The truncated protein (B3) from the P18 fraction was loaded onto a discontinuous sucrose gradient and subjected to ultracentrifugation as described in Materials and Methods. After centrifugation, the samples from the membrane fraction (interface between 10 and 50% sucrose), intermediate fraction (50% sucrose), sample loading layer (60% su-crose), and pellet fraction (bottom of tube) were collected and ana-lyzed by Western blotting.
FIG. 5. (A) Western blotting analysis of the viral proteins (B1, B2, and B3) in the membrane fraction. The yeast membrane fraction containing each viral protein was obtained from the discontinuous sucrose gradient and was analyzed with rabbit antiserum directed against the N-terminal portion of the BaMV replicase. (B) Guanylyl-transferase assay of the membrane-associated viral proteins. The assay buffer was as described in Materials and Methods and included 100 M AdoMet and 5l of enzyme solution. The intensity of each viral protein in the Western blotting analysis (A) suggests that approxi-mately equal amounts of viral protein were used in each assay. S2, proteins from the P18 fraction of yeast cells the harboring plasmid pYES2.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.333.528.75.384.2]from AdoMet would covalently link to the BaMV protein by incubating the B3 protein with Ado[methyl-3H]Met in the
pres-ence or abspres-ence of GTP. The results showed that the B3 protein was actually labeled by 3H and that this reaction
oc-curred only when GTP was present (Fig. 8). The second at-tempt was to identify the guanylate moiety released from the
32P-labeled BaMV protein. The32P-labeled B3 was isolated by
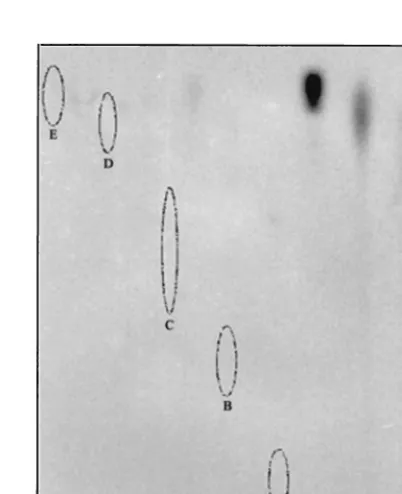
SDS-PAGE and concentrated by ultracentrifugation. The pro-tein was then treated with HCl, hydroxylamine, or NaOH, and the products were separated by TLC. The linkage of the32
P-labeled moiety to B3 was susceptible to acidic conditions but resistant to alkaline conditions, indicating a phosphoamide type of chemical bonding between the guanylate moiety and the protein (Fig. 9). The migration of the acid-released32
P-labeled moiety suggests that the guanylate moiety is identical to the standard m7GMP. It should be noted that the slightly
higher mobility of m7GMP in HCl than in acidic
hydroxyl-amine was also found in the standards. Taken together, the above data suggest that GTP is first methylated by the methyl donor, AdoMet, and then covalently linked to an active-site
residue of the BaMV capping enzyme through a phosphoam-ide bond. The active-site residue therefore most probably is lysine, histidine, or arginine. In the subsequent reaction, the m7GMP is transferred to the 5⬘-diphosphate terminus of the
RNA to complete the capping process.
Detection of methyltransferase activity.The methylation of B3 by Ado[methyl-3H]Met and the identification of m7GMP
[image:5.612.79.269.71.205.2]linked to B3 point to the existence of a methyltransferase
[image:5.612.330.531.404.651.2]FIG. 6. Immunoprecipitation of the 32P-labeled B2 and B3 pro-teins. The B2 or B3 protein from the membrane fraction was incubated with [␣-32P]GTP in the presence or absence of AdoMet (100M). The reaction mixtures were then incubated with antiserum against the N-terminal portion of the BaMV replicase, and the precipitated sam-ples were analyzed by SDS-PAGE and autoradiography. S2, proteins from the membrane fraction of yeast cells harboring plasmid pYES2.
FIG. 7. Nucleotide selection of the BaMV guanylyltransferase ac-tivity. The B2 or B3 protein from the membrane fraction was incubated with [␣-32P]GTP, [␣-32P]ATP, [␣-32P]UTP, or [␣-32P]CTP under stan-dard reaction conditions as described in Materials and Methods. The reaction products were analyzed by SDS-PAGE and autoradiography. S2, proteins from the membrane fraction of yeast cells harboring plasmid pYES2.
FIG. 8. Methylation of B3 protein by Ado[methyl-3H]Met. The B3 protein from the membrane fraction was incubated with Ado[methyl-3H]Met in the presence or absence of 100M GTP as described in Materials and Methods. The reaction products were then analyzed by SDS-PAGE and fluorography. S2, proteins from the membrane frac-tion of yeast cells harboring plasmid pYES2.
FIG. 9. TLC analysis of the guanylate moiety released from the 32P-labeled B3 protein. The32P-labeled B3 protein was treated with either HCl, NaOH, or acidic hydroxylamine, and the treated samples were then analyzed by TLC and detected by autoradiography as de-scribed in Materials and Methods. The standards GTP, GDP, GMP, m7GMP (in hydroxylamine), and m7GMP (in HCl) were run along with the analyzed products on a same TLC sheet and visualized by UV illumination (spots A to E, respectively).
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.73.273.543.662.2]activity on B3. To verify this possibility, B3 was incubated with Ado[methyl-3H]Met along with several potential methyl
accep-tors. The potential acceptors were then recovered with DEAE-Sephadex, and the labeling of3H on the acceptors was counted
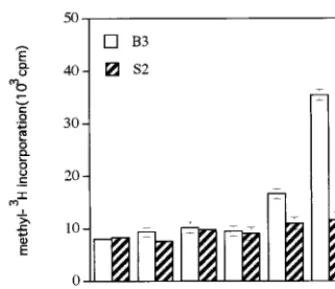
by liquid scintillation. The average results of three independent experiments are shown in Fig. 10. Although the incorporation of 3H was low, GTP could be methylated by Ado[methyl -3H]Met in a statistically significant way. GIDP, a
nonhydrolyz-able analog of GTP, was a better acceptor than GTP by a factor of approximately fourfold under the reaction conditions described in Materials and Methods. The methylation of GMP, m7GMP, and dGTP was insignificant.
DISCUSSION
ORF1 of potexviruses encodes a polypeptide whose size is larger than 150 kDa. This viral protein has long been proposed to be a replicase with polymerase activity residing at the C terminus and methyltransferase activity residing at the N ter-minus. The association of RNA-dependent RNA polymerase activity with the C-terminal domain has been demonstrated previously (16). In this study, the full-length 155-kDa protein of BaMV and two C-terminally truncated proteins were ex-pressed inS. cerevisiae. The activity assay and immunoprecipi-tation indicate that both the full-length and the truncated viral proteins have guanylyltransferase activity. The methyltrans-ferase assay also showed that the N-terminal portion has meth-yltransferase activity that catalyzes the transfer of a methyl group from AdoMet to GTP or GIDP. Therefore, our present study proves that the N terminus of the 155-kDa protein, from amino acid 1 to 442, harbors both guanylyltransferase and methyltransferase activities. Among the three recombinant proteins, B3 showed the strongest guanylyltransferase activity (Fig. 5). It is possible that the helicase domains in B2 and B1 hydrolyze GTP and, consequently, reduce the activity of trans-guanylation. The N-terminal portion of the BaMV replicase was also expressed inE. colicells with and without an S tag, a 15-amino-acid peptide specifically interacting with S protein derived from pancreatic RNase A, fused at the N terminus. No
activity was found in either of theE. coli-expressed proteins (data not shown). A problem of viral protein folding inE. coli or interference caused by the S tag might be the cause for enzyme inactivity.
Most replicase activities of the alphavirus-like superfamily, including alphaviruses and various plant viruses, were isolated from membrane fractions (11, 15, 23, 25). For Semliki Forest virus, membrane attachment has been demonstrated to be through the interaction of nsP1 (3). Moreover, the capping activity of nsP1 requires the association with anionic mem-brane phospholipids. The RNA-dependent RNA polymerase activity capable of synthesizing the BaMV genomic and sub-genomic RNA transcripts was also identified in the membrane fraction from the BaMV-infected plant cell extract (C. H. Tsai, unpublished data). The B3 protein expressed in yeast was found mainly in the P18 fraction, whereas B1 and B2 were found in both the P18 and S18 fractions. Fractionation of viral proteins by sucrose discontinuous-gradient centrifugation sup-ports the association of B3 protein with the membrane. Similar to what was found for nsP1 of Semliki Forest virus, membrane association could be crucial for BaMV capping activity since activities were found in the P18 fractions but not in the S18 fractions. BaMV capping activity may require a membrane environment to resume a correct protein conformation or ori-entation. The competition of yeast proteins in S18 for the utilization of GTP and AdoMet should offer another explana-tion. BaMV proteins need to be purified from the S18 fraction to further address the necessity of membrane association for transguanylation activity.
In general, the capping reaction at the 5⬘end of eukaryotic mRNA is accomplished by three consecutive enzymatic reac-tions. They involve RNA triphosphatase, mRNA guanylyl-transferase, and RNA (guanine-7-) methyltransferase. The or-der of these three catalytic reactions in alphaviruses and brome mosaic virus varies in that methylation of GTP occurs prior to transguanylation (1, 4, 12). It was therefore interesting to know whether potexviruses have similar characteristics with respect to the capping reactions. In the present study, we found that the guanylyltransferase activity of BaMV is AdoMet depen-dent and that m7GMP is the moiety linked to the enzyme. The
evidence provided in this study supports the conservation of the capping reaction throughout members within the alphavi-rus-like superfamily regardless of sequence dissimilarity among these capping enzymes. The N terminus of the BaMV replicase also exhibited methyltransferase activity. GIDP is a better methyl acceptor than GTP, and the preference was also found in nsP1 of Sindbis virus (20). For the 1a protein of brome mosaic virus, the formation of the adduct of m7GMP
[image:6.612.91.259.72.216.2]and 1a protein inhibited its own methyltransferase activity (4). It is possible that a similar phenomenon occurred in the BaMV capping enzyme when GTP was used as the methyl acceptor. GIDP is nonhydrolyzable; therefore it cannot link covalently to the B3 protein, and this may presumably render GIDP a better methyl acceptor. dGTP, a very good methyl acceptor in the case of the 1a protein, was a poor acceptor in this study. Despite the conservation of the main characteristics of the capping reaction, other subtle properties such as substrate specificity may differ between members of the alphavirus-like superfamily. Indeed, the overall sequence similarity between the first 450 amino acids of the BaMV replicase and the first FIG. 10. Methyltransferase activity of the B3 protein. Various
po-tential methyl acceptors (10 mM) were incubated with B3 protein and Ado[methyl-3H]Met. The methylated acceptor was recovered by a DEAE-Sephadex column and counted by liquid scintillation. CL, ac-ceptor was omitted from the reaction mixture; S2, proteins from the membrane fraction of yeast cells harboring plasmid pYES2. The re-sults are averages from three independent experiments.
on November 9, 2019 by guest
http://jvi.asm.org/
half of 1a of brome mosaic virus is no more than 10% based on the Clustal method.
Notwithstanding these distant relationships, several amino acid residues were found to be conservative among the capping enzymes of members within the alphavirus-like superfamily (24). The alignment of partial amino acid sequences of the capping enzymes from alphaviruses, brome mosaic virus, and BaMV is shown in Fig. 1. The importance of the conserved His residue for guanylyltransferase activity has been established by mutation analyses for Sindbis virus (30), Semliki Forest virus (2), and brome mosaic virus (4). In contrast, its role in meth-yltransferase activity is controversial in that the activity de-creased in Sindbis virus and brome mosaic virus, but actually increased in Semliki Forest virus, when the His residue was mutated. Mutation of the conserved Arg residue resulted in loss of methyltransferase activity and viral infectivity in Sindbis virus (30) and concomitant loss of guanylyltransferase and methyltransferase activities in Semliki Forest virus (2) and brome mosaic virus (4). Despite the above efforts, the in-depth function of these conserved residues remains obscure and re-quires extensive study. For the capping enzyme of yeast (5) and some well-studied viruses such as vaccinia virus (22) and Chlor-ellavirus (7, 8), the GMP moiety is attached to the enzyme via a lysine residue within a conserved active site motif (KXDG). No such conserved motif was found in the capping enzymes in the alphavirus-like superfamily. Based on a mutational result, Ahola and Ahlquist inferred that the conserved His residue links to m7GMP (4). Further physical evidence such as peptide
mapping and X-ray structure certainly would help to resolve these uncertainties.
ACKNOWLEDGMENTS
Y.-I.L. and Y.-J.C. contributed equally to this work.
This work was supported by grants NSC 87-2311-B-005-001-B11 and NSC 88-2311-B-005-001-B11 from the National Science Council, Re-public of China.
REFERENCES
1.Ahola, T., and L. Ka¨a¨ria¨inen.1995. Reaction in alphavirus mRNA capping: formation of a covalent complex of nonstructural protein nsP1 with 7-meth-yl-GMP. Proc. Natl. Acad. Sci. USA92:507–511.
2.Ahola, T., P. Laakkonen, H. Vihinen, and L. Ka¨a¨ria¨inen.1997. Critical residues of Semliki Forest virus RNA capping enzyme involved in methyl-transferase and guanylylmethyl-transferase-like activities. J. Virol.71:392–397. 3.Ahola, T., A. Lampio, P. Auvinen, and L. Ka¨a¨ria¨inen.1999. Semliki Forest
virus mRNA capping enzyme requires association with anionic membrane phospholipids for activity. EMBO J.18:3164–3172.
4.Ahola, T., and P. Ahlquist.1999. Putative RNA capping activities encoded by brome mosaic virus: methylation and covalent binding of guanylate by rep-licase protein 1a. J. Virol.73:10061–10069.
5.Fresco, L. D., and S. Buratowski.1994. Active site of the mRNA-capping enzyme guanylyltransferase fromSaccharomyces cerevisiae: similarity to the nucleotidyl attachment motif of DNA and RNA ligases. Proc. Natl. Acad. Sci. USA91:6624–6628.
6.Habili, N., and R. H. Symons. 1989. Evolutionary relationship between luteoviruses and other RNA plant viruses based on sequence motifs in their
putative RNA polymerase and nucleic acid helicase. Nucleic Acids Res. 17:9543–9555.
7.Håkansson, K., A. J. Doherty, S. Shuman, and D. B. Wigley.1997. X-ray crystallography reveals a large conformational change during guanyl transfer by mRNA capping enzymes. Cell89:545–553.
8.Ho, C. K., J. L. Van Etten, and S. Shuman.1996. Expression and charac-terization of an RNA capping enzyme encoded byChlorellavirus PBCV-1. J. Virol.70:6658–6664.
9.Jose, M.-C., G. Sutton, N. Ramadevi, and P. Roy.1998. Guanylyltransferase and RNA 5⬘-triphosphatase activities of the purified expressed VP4 protein of blue tongue virus. J. Mol. Biol.280:859–866.
10. Kadare´, G., and A.-L. Haenni.1997. Virus-encoded RNA helicases. J. Virol. 71:2583–2590.
11. Kao, C. C., and J.-H. Sun.1996. Initiation of minus-strand RNA synthesis by the brome mosaic virus RNA-dependent RNA polymerase: use of oligori-bonucleotide primers. J. Virol.70:6826–6830.
12. Kong, F., K. Sivakumaran, and C. Kao.1999. The N-terminal half of the brome mosaic virus 1a protein has RNA capping-associated activities: spec-ificity for GTP andS-adenosylmethionine. Virology259:200–210. 13. Koonin, E. V., and V. V. Dolja.1993. Evolution and taxonomy of
positive-strand RNA viruses: implication of comparative analysis of amino acid se-quence. Biochem. Mol. Biol.28:375–430.
14. Laakkonen, P., M. Hyvo¨nen, J. Pera¨nen, and L. Ka¨a¨ria¨inen.1994. Expres-sion of Semliki Forest virus nsP1-specific methyltransferase in insect cells and inEscherichia coli. J. Virol.68:7418–7425.
15. Laakkonen, P., T. Ahola, and L. Ka¨a¨ria¨inen.1996. The effects of palmitoyl-ation on membrane associpalmitoyl-ation of Semliki Forest virus RNA capping en-zyme. J. Biol. Chem.271:28567–28571.
16. Li, Y.-I., Y.-E. Cheng, Y.-L. Huang, C.-H. Tsai, Y.-H. Hsu, and M. Meng. 1998. Identification and characterization of theEscherichia coli-expressed RNA-dependent RNA polymerase of bamboo mosaic virus. J. Virol.72: 10093–10099.
17. Lin, N.-S., F.-Z. Lin, T.-Y. Huang, and Y.-H. Hsu.1992. Genome properties of bamboo mosaic virus. Phytopathology82:731–734.
18. Lin, N.-S., B.-Y. Lin, N.-W. Lo, C.-C. Hu, T.-Y. Chow, and Y.-H. Hsu.1994. Nucleotide sequence of the genomic RNA of bamboo mosaic potexvirus. J. Gen. Virol.75:2513–2518.
19. Mi, S., R. Durbin, H. V. Huang, C. M. Rice, and V. Stollar.1989. Association of the Sindbis virus RNA methyltransferase activity with the nonstructural protein nsP1. Virology170:385–391.
20. Mi, S., and V. Stollar.1991. Expression of Sindbis virus nsP1 and methyl-transferase activity inEscherichia coli. Virology184:423–427.
21. Mizumoto, K., and Y. Kaziro.1987. Messenger RNA capping enzymes from eukaryotic cells. Prog. Nucleic Acid Res. Mol. Biol.34:1–28.
22. Niles, E. G., and L. A. Christen.1993. Identification of the vaccinia virus mRNA guanylyltransferase active site lysine. J. Biol. Chem. 268: 24986–24989.
23. Osman, T. A. M., and K. W. Buck.1996. Complete replication in vitro of tobacco mosaic virus RNA by a template-dependent, membrane-bound RNA polymerase. J. Virol.70:6227–6234.
24. Rozanov, M. N., E. V. Koonin, and A. E. Gorbalenya.1992. Conservation of the putative methyltransferase domain: a hallmark of the ‘Sindbis-like’ su-pergroup of positive-strand RNA viruses. J. Gen. Virol.73:2129–2134. 25. Russo, P., P. Laakkonen, T. Ahola, and L. Ka¨a¨ria¨inen.1996. Synthesis of
Semliki Forest virus RNA polymerase components nsP1 through nsP4 in
Saccharomyces cerevisiaeby expression of cDNA encoding the nonstructural polyprotein. J. Virol.70:4086–4089.
26. Shibagaki, Y., N. Itoh, H. Yamada, S. Hagata, and K. Mizumoto.1992. mRNA capping enzyme: isolation and characterization of the gene encoding mRNA guanylyltransferase subunit fromSaccharomyces cerevisiae. J. Biol. Chem.267:9521–9528.
27. Shuman, S.1990. Catalytic activity of vaccinia mRNA capping enzyme sub-units coexpressed inEscherichia coli. J. Biol. Chem.265:11960–11966. 28. Shuman, S.1995. Capping enzyme in eukaryotic mRNA synthesis. Prog.
Nucleic Acid Res. Mol. Biol.50:101–129.
29. Shuman, S., and B. Schwer.1995. RNA capping enzyme and DNA ligase: a superfamily of covalent nucleotidyltransferase. Mol. Microbiol.17:405–410. 30. Wang, H.-L., J. O’Rear, and V. Stollar.1996. Mutagenesis of the Sindbis virus nsP1 protein: effects on methyltransferase activity and viral infectivity. Virology217:527–531.