Copyright ©1977 AmericanSociety for Microbiology PrintedinU.S.A.
Control
of
Protein
Synthesis by
a
Temperature-Sensitive
Mutant of
Reovirus 3
I.
Temperature-Sensitive Function of ts261-b Mutant
NOBUKO IKEGAMI
Division ofVirology, Memorial Sloan-Kettering Cancer Center, New York, New York 10021 Received for publication 25 May 1976
The ability of a temperature-sensitive (ts) mutant of reovirus, ts261-b, to synthesize virus-specificRNAsandproteinsduringinfectionatthe nonpermis-sive temperature (3700) was investigated. The relative amounts of the mutant virus-specificsingle-stranded (ss) RNAs anddouble-stranded(ds) RNAs synthe-sized in cells at 37C were 20 to 25% as much as those synthesynthe-sized in the wild-type virus-infected cells. The 10 segments of the mutant ds RNAs and the three size classes of the ss RNAs were synthesized in the usual proportions. The methyla-tion ofthe mutant viral mRNA's (ss RNAs) was not blocked at
3700
ininfected cells. A striking temperature-sensitive restricted function of the ts261-b mutant wasexpressed in the synthesis of the viral proteins. This study, which uses an in vitroprotein-synthesizing system reconstituted with an endogenous polysomal fraction andapostribosomal supernatant from reovirus-infected cells, has dem-onstrated that the endogenous polysomes obtained from ts261-b mutant-infected cellsat3700
are not active inthesynthesisof theviral polypeptides ofknown molecular weights, and the amounts ofthemutant viral polypeptides synthe-sized in vitro by these polysomes are 5 to 9% of those synthesized by the corresponding fraction from wild-type-infected cells. The impaired protein-synthesizing capacity ofthe mutant virus-specific polysomes can be restored during maintenance of the infected cellsat300C
after shift-down from370C.
The invitrosynthesis of viralpolypeptides ofknownsizeby theactiveendogenous polysomes derived from cells infectedatthepermissivetemperatureis acceler-ated by the addition of the postribosomal supernatant obtained from cells infected at the permissive temperature. The postribosomal supernatant from mutant-infected cells at370C
did nothave a stimulatory effect, but rather, it inhibited in vitro viral protein synthesis.The viralgenome of reovirus consists ofthe 10 segments ofdouble-stranded (ds) RNA, and eachsegmenttranscribesamonocistronic viral mRNA (5, 25). From this genomic structure, the existence of at least 10 genetic classes of temperature-sensitive (ts) conditional-lethal mutants ofreovirus can beanticipated. Thets mutants of reovirus type 3 (Dearing strain) have beenisolated in twolaboratories (12, 21). Those isolated by Fields andJoklikhavebeen classified into seven classes, A through G, by genetic recombination tests (12). Their investi-gations demonstrated that the ts mutants in
thedifferent classes expressed thedifferent re-stricted
functions
during the infective cycle at the nonpermissive temperature,390C.
The re-strictedfunctionsoftheir ts mutants were: (i) the virion-associated transcriptase (groupG; 7);(ii) the abilityto synthesize progenyds RNAs (groupsC, D, andE; 7, 23, 33); (iii)the assem-blyormaturation of progeny subviralparticles into the completed virus particles (groups B andG; 14, 34). All of thesetsmutants areable tosynthesize incells at 3900 the 10segments ofreovirus-specific
single-stranded
(ss) RNAs and all of themajorvirus-specific
polypeptides (7, 8, 13).The isolation and characterization of the ts
mutants that have been carried out in this laboratoryhave beenreported (21,22).The ge-netic class of thesets mutants isdifferent from each of the five classes A
through
E (unpub-lished data). Inthisreport, Idescribe thetem-perature-sensitive restricted function of a ts
mutant,ts261-b,isolatedinthislaboratoryand discusssomeaspectsrelatedtothe
regulation
of31
on November 10, 2019 by guest
http://jvi.asm.org/
32
reovirus proteinsynthesisthat have been
stud-iedinanin vitrosystemusing theendogenous virus-specific polysomes.
MATERIALS AND METHODS
Cells and media. L cells, strain 929, were used for all experiments. The media forgrowth of cellsand
the proceduresfor cultureofthe cellsweredescribed
previously (21, 22).
Viruses.Reovirus type 3, Dearing strain, cloned
(19), was used as thestock of wild-type virusafter
two further passages in L cells in suspension at
370C. The clonedts261-b mutants (22) were grown up
to astockin Lcellsonmonolayersafterone passage
at30'C. Fortheproductionof the mutant virus, cells
were infected at the low multiplicity of infection
(MOI) of 0.1 PFU/cell. When the complete
cyto-pathiceffect was observed, the infected cells were
frozen and thawed, and the lysate wascentrifuged
at alowspeedto removecelldebris. The virus
par-ticles in the clarified supernatant werecollected in
apellet by centrifugationat72,000 xg for3h,and
thepellet wasresuspendedinfresh mediumat1/20 of the original volume. This material was usedas the stockof ts mutant virus.
Isotopes and chemicals. [U-14C]uridine (447mCi/
mmol), L-[methyl-3H]methionine (11 Ci/mmol),
S-adenosyl-L-[methyl-3H]methionine (12.6 Ci/mmol),
L-[35S]methionine (150 to 380 Ci/mmol), 3H-labeled
L-aminoacids mixture,andGTPlabeledwith 32p in the a position werepurchasedfrom New England Nuclear Corp. (Boston, Mass.). Unlabeled ribonu-cleosidetriphosphateswereobtained from P-L Bio-chemicals (Milwaukee, Wis.). Goat antiserum to
rabbit 7S gammaglobulin was purchased from
Hy-land Laboratories(CostaMesa,Calif.).
Preparation ofcytoplasmicextracts forthe
ex-tractionofvirus-specificRNAs. The L cells in
sus-pensionculture were infected witheitherwild-type
virusorthets261-bmutant at an MOI of 20. After an
adsorption of2 h, the suspension ofinfected cells
wasmadeataconcentration of about106 cells/mlin
complete spinner medium containing 5% fetal
bo-vine serum. Whereindicated, the suspension of
in-fected cells was made in methionine-free spinner
medium containing 5% fetal bovine serum. Actino-mycin D (0.15 ,ug/ml) was present throughout the infective cycle. The cells were collected by
centrifu-gationand werewashedoncewithRSB-Na+ buffer
(0.01 M Tris-hydrochloride [pH7.4], 0.01 MNaCl, 0.0015 MMgCl2), andthen the infected cells were disrupted in RSB-Na+ buffer containing 2 mM dithi-othreitol, 0.1% Nonidet P-40, 20 ,ug of polyvinyl sulfate perml, and10% glycerol in a Dounce homog-enizer.The cytoplasmic extract, free ofnuclei, was obtained as described by Penman (37).
Extraction of RNA from cytoplasmic extract, separation ofreovirus-specificss anddsRNAs, and velocity sedimentation analysis. To the total
cyto-plasmicextract, sodium dodecyl sulfate (SDS) and
polyvinylsulfate wereaddedtofinal concentrations
of2% and 200j.g/ml,respectively;then the cytoplas-mic extract was diluted threefold in TNE buffer (0.01MTris-hydrochloride [pH7.4],0.001MEDTA,
0.1 M NaCl) containing 0.6% SDS. Extraction and purification ofcytoplasmic RNAs were carried out according totheproceduredescribedby Moyeretal. (35). The purified RNAs were precipitated by the addition of 2 volumes ofethanol, and the RNA pre-cipitate wasdissolved in TNE buffer. Anequal vol-ume of 4 M LiCl was added to the RNA solution to separate ss RNA from ds RNA. The mixture was kept overnight at 0°C and then centrifuged. The resulting precipitatecontained ss RNA. The ds RNA present in the supernatantsolution was precipitated once morebytheaddition of2volumesof ethanol. The RNA in each precipitate wasdissolvedinTNE
buffercontaining 0.5%SDS. TheRNAsamplewas
layeredonto alinear 15 to 30% sucrosegradientin
TNE buffer andcentrifugedat 30,000 rpm for 17 h in an SW41 rotor at 15°C. Fractions of 0.35 ml were
collectedfrombelow andassayedfor acid-insoluble
materiallabeled with theradioactivityasdescribed by Gomatos (18). The sedimentation values of the
radiolabeled RNAs weredeterminedrelative to the
positionsof 28S and 18SrRNA's,which sedimented ina companion sucrosegradient.
In vitro transcription. (i) RNA synthesis by
wild-typeviral cores. Theassayprocedures for in vitro
RNA synthesis by the wild-type virion-associated
transcriptase were the same as those described by
Shatkin (40). The complete reaction mixture,
con-taining the chymotrypsin-treated wild-type virion, was incubated for 60 min at 45°C. At the end of incubation, the reactionmixture was centrifuged at 30,000 xgfor 30 min. The supernatant, which con-tained the RNA products, was saved, and the pel-leted viral cores were resuspended in the fresh, com-plete reaction mixture. Incubation was carried out
for another 60 min at45°C. The reaction mixture
wascentrifuged as described above, and the super-natant obtained was combined with the previous
one. The RNA products in the combined
superna-tant werepurified by extracting three times with an
equal volume ofamixture (1:1) of phenol and
chloro-form containing isoamyl alcohol. The RNA
mole-cules in theaqueous phasewere chromatographed
through SephadexG-50. Fractions containing
radio-active RNA were pooled and precipitated with 2 volumes of ethanol in the presence of 0.3 M NaCl.
(ii) RNA synthesis by ts261-b mutant subviral
particlespresent inthe cytoplasmic pellet. The
cy-toplasmic extract wascentrifuged at4°Cfor 20min at 35,000 x g. The pelleted material was
resus-pended by homogenization in0.01 M Tris (pH 8.1)
and was used as a source of the mutant subviral
particleswithout furtherpurification.The
cytoplas-mic pellet containing 900 ,ug of protein was incu-bated at 37°C for 30 min in the presence of chymo-trypsin (450 ,ug). Atthe end of incubation, all of the components necessary for in vitro transcription wereadded, andthe final concentration ofeach com-ponent in the reaction mixture was the same as described by Shatkin (40). The incubation was con-tinued at 37°C for 90 min. The reaction mixture was centrifuged at 35,000 x g for 30 min. The RNA product in the supernatant waspurifiedby the pro-cedure described above.Theproductassociated with the pelleted material was extracted andpurified by
on November 10, 2019 by guest
http://jvi.asm.org/
the proceduresdescribed for purification ofssRNAs fromthe cytoplasmic extract.
Preparation ofsubcellular fraction for invitro
protein synthesis. Cells were infected with
wild-type virus at anMOI of20orwiththets261-b mu-tant at anMOI of 50.Afteradsorption of virus for 2 h, the cells were resuspended in complete spinner mediumcontaining 5%fetalbovine serum,and acti-nomycin D (0.15
Ag/ml)
wasadded to the infected cellsuspension(106/ml) andwaspresentthroughoutthe infective cycle. The infected cellswerecollected
atthe time indicated and were washedonce with
RSB-K+buffer(pH 7.8).Thewashedcellswere
dis-rupted in a Dounce homogenizer in the RSB-K+
buffer containing 2 mM dithiothreitol and 0.2%
NonidetP-40.Thecellhomogenatewascentrifuged
at250 xgfor8min. The postnuclear supernatant was collected and mixed with glycerol in a 9:1 (vol/ vol)proportion.The nuclear sedimentswere
resus-pendedinbufferA(RSB-K+ containing2mM
dithi-othreitol and10%glycerol); then the suspensionwas
centrifuged at 600 x g for 8 min. The resulting
supernatant fluidwascombinedwiththe
superna-tantcollectedpreviously. Thetreatmentofthe
nu-clear sediments wasrepeated once ortwice in the
same way,andthesupernatantfluidscollected after
each centrifugation were combined with the
pre-vious ones. The total postnuclear supernatant fluid was then centrifuged at 10,000 x g for 20 min. The resulting supernatant fluid was designated S-10.TheS-10 wasfurthercentrifugedat150,000x
gfor2h.The clearsupernatantfluid inthemiddle
part of the centrifuge tube was collected, and it wasdesignated S-150. The surface of the pellet was washed once with buffer A, and the material in the
pelletwasresuspended inbuffer Aby
homogeniza-tion, to give a protein concentration of 13 to 15 mg/ml. This suspension wasdesignated P-150.
Assay forin vitrosynthesis of viral polypeptides.
In a total volume of600 ,l, thefollowing
compo-nents were added to give the final concentrations
indicated:Tris-hydrochloride (pH 7.8),44mM;KCl,
40mM;NH4Cl,44mM;MgCl2,4.5mM;
2-mercapto-ethanol, 11 mM; ATP, 1.1 mM; GTP, 0.5 mM;
K-phosphoenolpyruvate, 5 mM; pyruvate kinase, 160
,g/ml; 19unlabeledamino acids(except
L-methio-nine), 2.2
AM;
L-[35S]methionine (150 to 380 Ci/mmol), 100 ,Ci; subcellular fraction, 420 ,l. The
reaction mixture wasincubatedat30°C.Toanalyze
kinetics of in vitro incorporation of -[35S]methio-nine, asampleof20,ul was removed and mixed with 2mlof5%trichloroaceticacidatthe timeindicated. After 30 min at 0°C, the samples were heated at 95°Cfor20minand then cooledat0°C.Theresulting
precipitates were collected on membrane filters
(Millipore Corp.), and the amount of radioactive
material on the filter was counted.
Immuneprecipitation of viralpolypeptides syn-thesized in vitro. Tworabbitserahyperimmune to reovirus werepreparedinthislaboratory.To
immu-nizerabbits,virusstockwaspreparedinthe
follow-ing way. Wild-typevirus wasgrowninmonkey
kid-ney cells, and the ts mutant virus was grown in
BHK-21 cells. Thelysateofcell infected with each viruswascentrifugedto removecelldebris,and the
clarified supernatant was used for immunization. One antiserum is hyperimmune to the wild-type virusand the otherishyperimmune to the ts mu-tant. The procedurefor immune precipitation was essentially as describedbyFields et al. (13). A rab-bit immune serum (20 ul) was added to the reaction mixtureof 480
tMl.
The mixture wasincubated at4°Covernight, andanadditional 20,ulof thesame
rab-bit immune serum was added to this mixture and incubated overnight at 4°C; then 0.2 ml of goat anti-rabbit serum was added. Incubationwas continued for an additional 24 h at 4°C.The immune precipi-tate wascollected by centrifugation andwaswashed three times with cold phosphate-buffered saline. Withthisimmune precipitationmethod,tworabbit immune sera were equally capable of precipitating over 90% ofradiolabeled virion structural proteins of aknown amount, as judged by the total radioactive counts precipitated and bythe polyacrylamide gel
electrophoresis (PAGE)patternof thelabeled
poly-peptides ofX-, ,u-, and a-size classes. When the
immune precipitation method was applied to the reaction mixture at the end of the in vitro assay, each ofthe reovirus antiseraprecipitated
approxi-mately15%ofthe total acid-precipitable
radioactiv-ity,whereas normal rabbit serumprecipitated less
than 1%oftheacid-precipitablecounts.
PAGE. Analysis of viral proteins. The im-mune precipitate was dissolved by heating at 100°C for 2 min in 100 ,ulof buffer containing 0.1 M
phos-phate(pH 7.4), 2%SDS,4%2-mercaptoethanol, and
0.5 Murea.Thetotaldissolvedsample was subjected
toPAGEanalysis.The proceduresforpreparationof
polyacrylamide gelcontainingSDS,urea,and
phos-phatebuffer and for electrophoresis of gel in a
phos-phate buffersystem were as described previously
(22). The gel was frozen and thawed once and then cut into 1-mm slices. Two slices were placed into
eachvial and solubilized by heating at 55°C
over-nightin0.5ml of hydrogen peroxide.Ten milliliters
ofatoluene-TritonX-100 (2:1)scintillationmixture
was added, and the amount ofradioactivity was
determined. Toserve as internal protein markers,
3H-aminoacid-labeledpurified reovirus, after
solu-bilization, was subjected to coelectrophoresis with
each immune precipitate.
Chemical analysis. Protein concentration was
determined by theLowryprocedure (30), using
crys-talline bovineserum albumin as the standard.
RESULTS
Synthesis of virus-specific RNAs by the
ts261-b mutant at the nonpermissive
tempera-ture, 37°C. Since the level of the yield of the infectiousts261-b mutantat37°Cis 0.01%of the maximalyieldproducedat30°C, 37°C has been chosen as the nonpermissive temperature for the ts261-b mutant. To compare the ability of the ts261-bmutant tosynthesizethe virus-spe-cific ds andssRNAsinvivo at37°Cwiththat of the wild-type virus, cells were infected with either ts261-b mutantor wild-typevirusat an
33
on November 10, 2019 by guest
http://jvi.asm.org/
MOI of20 and were labeled with ['4C]uridine
for the respective period, from2 to 6h(the early time) and from 6 to 11 h (the latetime) after infectionat370C. The RNAswereextracted and
purified from the cytoplasmic extract of each
set of the infected cells. The ss RNAs were
separated from the ds RNAs by2MLiCl precip-itation. After velocity gradient centrifugation of the RNAsamples, theamountsof the
reovi-rus ds RNAs that sedimented between 10.5S
and14S (1) and those of the reovirusssRNAsof
the three size classes1, m,and s (17, 25) were
determined. The resultsareshown inTables 1
and 2.
Theamountof themutantdsRNAs
synthe-sized andlabeled in vivoat370C relativetothat of thewild-type viralds RNAswas15%earlyin
the infective cycle and8%late inthe infective cycle (Table 1). In experiment 2, when cells
were infectedat 370Cwith the ts261-b mutant
at an MOI of 250, the increase in the total
synthesis of themutantviraldsRNAswasless
than twofold, whichwasnotproportionaltothe increase in MOI. These data indicate that the maximal level of ts261-bmutantds RNAs
syn-thesizedin vivoat370C is,atmost,20 to25%as
much as that of the wild-type viral ds RNAs
synthesizedat370C. PAGEanalysis of the
mu-tant ds RNAs synthesized in vivo at370C
re-vealed that the 10 segments ofthe mutantds RNAhad been synthesized (Fig. 1). The ts261-b
mutant, thus neither is adeletionmutant (43)
TABLE 1. Synthesis ofreovirus ds RNAs in vivoat 37°C
Period of 14CCpM
labeling inds Ratio
Expt Infectiona postinfec- RN
tionb X103
(h)
1 ts261-b 2-6 15 15.3
Wildtype 2-6 98 100
ts261-b 6-11 26 8.4
Wildtype 6-11 307 100
2 ts261-b 2-11 68
aAtotal of108cellswereinfected with each virus atanMOI of 20 inexperiment 1and 250 in experi-ment2. ActinomycinD (15jig)wasadded2h after theonsetofinfection andwaspresentuntil the end ofthelabeling period.
bInfected cellswerelabeled with['4C]uridine (50
,uCi)for theperiodindicated.
cRNAs were extracted and purified from the wholecytoplasmicextractof eachset,and ds RNAs
wereseparated fromssRNAsasdescribed in
Mate-rials and Methods. 14C cpm represents the total
counts incorporated intoreovirus-specific ds RNAs that sedimentedbetween 10.58 and14S(1)andwere
resistanttohydrolysis bypancreatic RNase(3 jug/
[image:4.508.264.456.74.184.2]ml).
TABLE 2. Comparison ofinvivotranscription at 370C
Period of '4Ccpm(x10-3) inssRNAs8 labeling
Infection
postin-fection 1 m 8 Total
(h)
ts261-b 2-6 17 20 18 55
Wild type 2-6 77 88 69 234
ts261-b 6-11 86 113 84 283
Wild type 6-11 438 499 388 1,325
aProceduresfor infection ofcells at an MOI of 20
and for labeling ofinfected cells are the same as
thosedescribedinTable1.
b RNAs were extracted and purified from the
whole cytoplasmicextractof each set,andssRNAs
wereprecipitatedwith2MLiCl. Threesizeclasses
ofvirus-specific ss RNAs (1, m, and s) were
sepa-rated by velocity gradient centrifugation as
de-scribed in Materials and Methods. The
sedimenta-tionvalues of the three sizeclasses,24S for1,19Sfor m, and 14S for s (17), weredeterminedrelative to the position of 28S and 18S rRNA's, which sedi-mented in a companion sucrose gradient. 14C cpm
represents the total counts incorporated into the respective ssRNAclass.
nordoes it seem to belong to the C, D, and E classes (23),butitcanbeplacedintothe cate-gory ofds RNA-positivemutantsbyusingthe criteriathathave been applied for the ts mu-tantsof classes A,B, F, and G (25).
The ability of the ts261-b mutant to tran-scribe the reovirus-specific ss RNAs invivo at
370C
is also20to 25%of the level of transcrip-tionexpressed by the wild-typevirusthrough-outthe infective cycle (Table2).Whenthe mu-tant subviral particles obtained from cells in-fectedat37 or300C and the wild-typesubviral particles obtained from the infected cells at 37°C were assayed invitrofor their transcrip-tase activities,theratesofthe invitro synthe-sis of the virus-specific ss RNAs were two- to threefoldgreater at
370C
astheassay tempera-turethan at300C
inall cases. It is not clearly understood at present what mechanism(s) is responsible for thereduced level of in vivo tran-scriptionintheearly time after infection with ts261-b mutant at370C.
Inhibition of protein synthesis in ts261-b mutant-infected cells at
370C.
Incells infected with the ts261-b mutant at the nonpermissive temperature,370C,
there occurs dissolution of pre-existing host cell polysomes, and the inhibi-tion ofsynthesis of host cell protein is evident by 7 h after infection. When such cells are pulse-labeled for 15 min with the radioactive aminoacidsatany time after7hpostinfectionat
370C,
the incorporation of the radiolabeled amino acids into the polysomal fraction ison November 10, 2019 by guest
http://jvi.asm.org/
[image:4.508.50.253.424.538.2]BY MUTANT
I IV
ts261-bdsRNAs
M S
i-_
WildtypedsRNAs
L S
4-
2-20 40 60 80
Mobility(mm)
FIG. 1. Polyacrylamide gel electropherogram of reovirus ds RNAssynthesizedin vivoat37°C. Virus-specific ds RNAs from infected cells labeled with [14C]uridine for 9 h (2 to11 h after infection) were
obtained and purifiedasdescribedinMaterialsand
Methods and in Table1. ThedsRNA sample was
analyzed in 7.5% gel. Theprocedure forpreparation ofgel and for electrophoresiswere asdescribedby Ito
andJoklik (23). Electrophoresiswascarriedout at4 mA/gel for 30 h. The gel wascutinto1-mmslices,
andtheradioactivityineachslicewasdeterminedas
described in Materials andMethods.
markedly inhibited. When the cytoplasmic
ex-tract of ts261-b mutant-infected cells at 370C
wasanalyzed byPAGE,itwasdifficult to
iden-tify any virus-specific polypeptides of known
molecular weights, because the amount of the
newly synthesizedand labeled proteins inthe
cytoplasmic extract ofts261-bmutant-infected
cellswas too little to resolve into
distinguish-able proteinbands. However, aswas reported previously(22), newly synthesizedmutant
sub-viral particles at a density of 1.43 g/ml were
detectable inCsCldensity gradient analysisof
a concentrated cytoplasmic pellet, but no
ma-ture mutant progeny particles at adensity of
1.36 g/ml were detectable. To determine
whether all ofthe viralproteinsaresynthesized
in a markedly reducedamount not detectable
in a conventional PAGE analysis or whether
the synthesis of a particular viral protein is
selectively inhibited in ts261-b mutant-infected cells at 3700, the present experiments have been carried outby analysis of in vitroproducts synthesized in a cell-free system reconstituted with a concentrated polysomal fraction and a
postribosomal supernatant from infected cells. Formationof polysomes in vitro byfraction S-10 obtained from reovirus-infected cells. Thepostmitochondrial supernatantfraction (S-10), obtained from a cytoplasmic extract of wild-type virus- or ts mutant-infected cells at the permissive temperature, containsaquantityof virus-specific ss RNA of the threesizeclasses, anamountequal to 50% of the total present in the original cytoplasmic extract. These virus-specific ss RNAs are already bound to ribo-somesandpolysomes. If all components neces-sary for protein synthesis are present and func-tionallyactive intheS-10fraction, these endog-enous viral mRNA's are translated during in vitro incubation of the S-10 fraction with the protein-synthesizing assay mixture, and the subsequent analysis of the reaction mixtureby velocity gradient centrifugation will demon-stratethe incorporation of35Slabelintothe80S monosomes and polysomes of various sizes. Three kinds of S-10 fractions were prepared to compare theirprotein-synthesizing abilities: (i) the wild-type S-10, which was obtained from cells infected with wild-type virus for 8 h at
370C
(the permissive temperature); (ii) the ts261-bS-10 (370C),fromcellsinfectedwiththe mutantvirusfor8.25h at370C (the nonpermis-sive temperature); (iii)the ts261-b S-10 (300C), fromcells infected with themutantvirusfor19 h at 300C (the permissive temperature). The length of infection atthe permissive tempera-ture chosen in this experiment wasoptimal to obtaintheS-10fractions, whose protein-synthe-sizing activitywasinthe range of themaximal level. Each of these S-10 fractions wasmixed with the the assay mixture and incubated in vitro at 30 or37°Cimmediatelyafter addition of[35S]methionine.
Atthe end ofincubation, eachreactionmixture wasanalyzedbysucrose gra-dientcentrifugation. Theresults are shownin Fig. 2. The incorporation of 35Slabelproceeded duringincubation at 30 or370C, but not at00C (Fig. 2a). The 35S-labeled 80S monosomes and polysomes were formed in the reaction mix-turesthatcontained theS-10fractionfromcells infected at the permissive temperature with either wild-type virus (Fig. 2b) or the ts261-b mutant (Fig. 2e and f). The increase in 35S
counts inthepelletedmaterials suggestedthat
the larger polysomes were formed and sedi-mentedduringvelocity gradientcentrifugation. In contrast tothese results, the datainFig. 2c and ddemonstrated that the ts261-b S-10
(3700)
VOL. 22, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.508.81.226.55.344.2]36
Wild-typeS-10 fromcells infected at37° for 8 hrs. a)0min.atO' 12Fb)370
ls261-b S-10 fromcells infected at370 for81/4hrs,
4 c)300 4 d)37°
E tP;12xi06 80S P06.106 80S
Qo
ts 261-bS-10 from cells infected at 30°for19 hrs. iz-e)30°1 l'' f) 370
80S 480S
8
8-4 4
tp; e8 loetp2 X0
0 10 20 30 0( 10 20 30
[image:6.508.67.244.65.326.2]Fraction number
FIG. 2. Formation of 80S monosomes and poly-somesduring in vitro incubation ofS-10 in theassay
mixture for protein synthesis. The reaction mixture (2.4ml) contained S-10 andL-[35S]methionine (250
,uCi); itwasincubatedfor 20 minat30°Corfor 30
min at37°Cand then treated with detergents (so-diumdeoxycholate,Brij-58,and EDTAatfinal
con-centrationsof0.7%, 0.5%, and 0.5 mM, respectively). Thedetergent-treated samplewaslayeredon alinear
15to30%sucrosegradient inRSB-Na+ buffer.
Cen-trifugation wasat24,000 rpmfor8 h in an SW27
rotor at4C. Fractions of1 ml were collected. The
absorbencyat260nmandacid-insoluble
radioactiv-ity in each fraction were determined as described
previously (22). Theamountofacid-insoluble radio-activity in the pelleted material( P)is shown in the
lowerleftcornerof each panel.
obtained from cells infected at the
nonpermis-sivetemperature(3700)hadessentiallyno
cata-lytic activity for incorporation of35Slabel into
polysomes atbothassaytemperatures, 30 and
370C.The data suggest that theremaybesome
defectsorrestrictioninfunctionofasubcellular
components) presentinthe ts261-bS-10 (370C).
Analysis of viral proteins synthesizedin vi-troby polysomal fraction fromcells infected at the permissive temperature. To examine
whetheradefect(s) responsible for inhibition of
protein synthesis in ts261-b mutant-infected
cellsat370Clies in thepolysomalfractionorin
a components) present in the postribosomal
supernatant, theS-10fractionwasfurther
cen-trifuged at 150,000 x g and separated into a
pellet, P-150, and a postribosomal supernatant, S-150. The P-150 contained free polysomes, monosomes, 60S and 40S ribosomal subunits, and viral mRNA's of the three size classes. The S-150, a clear portion of postribosomal superna-tant, did not contain ribosomes or their sub-units. The viral mRNA's of the three size classes were also undetectable in theS-150.
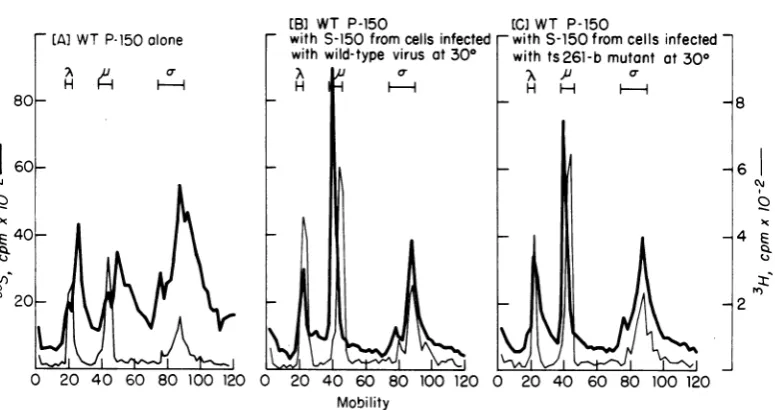
(i) In vitro products synthesized by the P-150(30CC). The P-150(30'C)wasobtained from cells infected with eitherwild-type virusorthe ts261-b mutant at300C for 21 h, and its protein-synthesizing ability was assayed in vitro in the absence of S-150. Virus-specific proteins syn-thesized by each P-150 (30'C)wereprecipitated with a reovirus-specific antiserum and
ana-lyzed by PAGE. Theelectrophoreticpatterns of in vitroproducts areshown in Fig. 3. Both P-150 (300C)preparationsof the wild type and the ts261-b mutant were capable of synthesizing reovirus polypeptides in vitro at 30and 370C. In the absence ofS-150, the P-150 (300C) synthe-sizedpredominantlythepolypeptidesof the size corresponding to the a- class (43,000 to 33,000 daltons). The 35S-labeled polypeptides, whose mobilities corresponded to the ,-size class (88,000 to 70,000daltons)and theA-size(155,000 to 140,000 daltons), were synthesized in one-third amountsof the35S-labeled polypeptidesof thea-size class. Thus, the relativeproportions of the three size classes (X, ,u, and c-) of 35S-labeled virus-specific polypeptides synthesized bythe P-150 (300C)did notcorrespond to those ofthe three size classes of 3H-labeled virion polypeptides. In addition, 35S-labeled polypep-tides oftwoother size ranges weredetectablein substantial amounts. They migrated faster than the polypeptides ofthe A-size class and those of the ,u-sizeclass, respectively. The ap-parentmolecular weightsofeach are approxi-mately140,000 to 120,000daltonsfor theformer and69,000 to 55,000daltonsfor the latter. Since these polypeptides were precipitated with the reovirus antiserum but did not correspond to thesizes ofthe reovirus structural proteins and to those oftwo nonstructural proteins (88,000 and 36,000daltons),itappears that they may be incomplete polypeptides of the K- and ,-size classes, respectively, that may have arisen from incomplete elongation or from premature termination underthe assay conditions in the absence of S-150. This assumption, however, remains to be proven by identification of the authentic tryptic peptides, which correspond totrypticpeptides from the virion polypeptides ofthe three size classes.
(ii) Effect of postribosomal supernatant (S-150[30°C1).When theS-150(300C)obtained from wild-type virus-infected cells was addedto the
19
IID
on November 10, 2019 by guest
http://jvi.asm.org/
Protein synthesis in vitro cells infected at 30°
product at 300
D 140,00072,000 34,000
Do H A
by Wild-type P-150 obtained from
product at 370
V740,2000 34,000
4 X
I H H-H
Mobility (mm)
Protein synthesis in vitro by ts 261-b P-150 obtained from cells infected at 30°
20 40 60 80 100 120 0 20 40 60 80 100 120 Mobility (mm)
FIG. 3. Reoviruspolypeptides synthesizedinvitrobywild-type P-150(300C)andbyts261-bP-150(300).
Each P-150(3090)wasprepared fromcellsinfected withwild-typevirusorthe ts261-bmutant at300C for20 h.The reaction mixture(600,d)contained 100
pi
ofeachP-150(1.3mgofprotein),320pi
ofbuffer A,and 60pi
of L-[35S]methionine andwas incubatedfor100 minat300Cor50 minat370C.During incubation,asample (20 p1) from each mixture was withdrawn sixtimes, at equal intervals, forkinetic analysis of
L-[35S]methionine incorporation (datanotshown). Afterterminationoftheassay,thereovirus-specificproteins intheremaining reactionmixture(480 d)wereprecipitatedwiththereovirus-specificantiserum. The whole immune precipitate wassolubilized andsubjectedtoPAGEanalysis. Theprocedureswere asdescribedin Materials and Methods. To serve asinternal markerproteins, 3H-labeled polypeptides fromthe wild-type
virionweresubjectedtocoelectrophoresiswitheach 35S labeledsample.Thepositions ofthe three size classes ofreovirus-specific polypeptidesareindicatedbyX(155,000to140,000 daltons), u(88,000to70,000 daltons),
and (43,000to33,000daltons) (46).
wild-type P-150 (300C) and the mixture was
assayed at3000 (see Fig. 4B),thepolypeptides
of the three size classes (X, g, and a-) were
synthesized and labeled with 35S in proportions
similarto thosesynthesized invitrobya
post-mitochondrial supernatant ofreovirus-infected
cells(31). The polypeptides of 140,000to120,000
daltons and those of69,000 to 55,000 daltons
were no longer detectable asthe major species
of theproduct.
Electrophoretic patterns similar to those of
the invitroproductswere seenwhen the
wild-typeP-150(3000)wasassayedinthepresenceof
the S-150 (3000) obtained from cells infected
withthe ts261-bmutant atthepermissive
tem-perature (seeFig. 4C)orinthepresenceof the
2
I
13
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.508.147.379.62.461.2]infected-at 300
80- 8
160
6~40-
[image:8.508.63.451.66.271.2]20-0 20 40 60 80 100 120 0 20 40 60 80 100 120 0 20 40 60 80 100 120 Mobility
FIG. 4. Viral polypeptides synthesized in vitro by wild-type P-150(309C)and effect ofS-150 (309C) on this protein synthesis. The wild-type P-150(300C)andthe S-150(300C) were preparedfromcells infected at300C
for20h,andthets261-bS-150(300C)waspreparedfrom mutant-infectedcells at 309Cfor 21 h. The reaction mixturecontained 100,4ofwild-type P-150(300C)(1.4 mgofprotein) and 320piofeitherS-150(300C).Ina
controlassay, theS-150(300C)wasreplaced bybuffer A. Incubation was at300Cfor 100min.Theprocedures
forassay andfor analysis ofthe invitroproducts were as described in thelegendtoFig.3and inMaterials
andMethods.(A)Product in the control assay. (B) Product in the presence of wild-typeS-150(300C).Protein
concentrationof the S-150(300C) added to thereaction mixture was 1.3 mg. (C) Product in the presence of ts261-b S-150(300C).Proteinconcentration of theS-150(300C)added was 1.4 mg. 3H label in each panel
denotes polypeptidesfromthewild-type virion subjected tocoelectrophoresis.
S-150 obtained from mock-infected cells (data notshown). When theS-150 (300C) from virus-infected cells was added to the P-150 from mock-infected cells and the mixture was as-sayed under thesameconditions,no35S-labeled polypeptides were detectable in the immuno-precipitates. ThemutantP-150 (30°C)wasalso analyzed under similar conditions. The re-sponseof themutantP-150
(3000)
tothese S-150 was similar to that seen in the assay of the wild-type P-150.Protein-synthesizing ability ofthe mutant P-150 (37°C) obtained from cells infected at thenonpermissive temperature
(370C).
To de-termine the level ofprotein-synthesizing abil-ity ofthe P-150 from mutant-infected cells at370C,
cells were infected at 37°C with either ts261-b mutant or wild-type virus for11h.Each cytoplasmic extract was made from an equal number of the viable infected cells. The P-150 (37°C) was obtained from it and analyzed invitro at
300C
for itsprotein-synthesizing activ-ity.Thekinetics of incorporation of 35Slabelinto hotacid-precipitable material are shown in Fig. 5(left panels). The level of the protein-synthe-sizing ability of the ts261-b P-150
(370C)
wasless than one-tenth that of thewild-typeP-150. PAGE analysis of the virus-specific
polypep-tides that were synthesized by each P-150 is showninFig.5Dand G (note 10-fold difference inscale for35S). Most of themutantviral poly-peptidessynthesizedbythe ts261-bP-150(370C) werepolypeptides of 140,000 to120,000daltons andthose of 43,000to33,000daltons(Fig. 5D). The electrophoreticpatternof theproductmade by the ts261-b P-150 (370C) was qualitatively similar to that of the wild-type product (Fig. 5G).However,theamountsofthemutant poly-peptides synthesized and labeledwith35Swere
only 5 to9%o of thosesynthesized by the
wild-typeP-150 (370C).
Effect ofpostribosomal supernatant(S-150 [370C0). To determine if
synthesis
of the mu-tant viral polypeptides ofthe ,u-size classin-creasesbythe addition ofS-150to these P-150
(370C),
the S-150 (370C)obtained from cellsin-fected at 370C with either wild-type virus or ts261-bmutant was addedtoeach oftheP-150 preparations, and virus-specific polypeptides synthesized byeachP-150(370C)wereanalyzed by PAGE. When the wild-type P-150 was
as-sayed in the presence of the wild-type S-150 (370C) (Fig. 5H), the synthesis of 35S-labeled wild-type polypeptides of the ,- and a-size classes increased two- to threefold above the levels observedinFig. 5G. It appears that the stimulatory effect of the added
wild-type
S-150J.
on November 10, 2019 by guest
http://jvi.asm.org/
- Invitro300
[D]-ts261-bP-150 alone
[El---with WT S- 150 [F]-- with 261-b S-150
D
F
20 40 60 80 100
6
1
&2
Invitro 30°
[G]-WTP-150 alone (HI---withWT S-150
[I)- -with261-b S-150
Time (minutes)
ts 261-b P-150 from cells infected for 11 hrsat37°
x t a, [DI Al a, (El 'A ) a, F]
20 40 60 80 100 120 0 20 40 60 80 100120 0 20 40 60 80 100 120
Wild-typep-150 from cells infected for 11 hrsot370
Mobility (mm)
FIG. 5. Viralpolypeptidessynthesizedin vitroby ts261-b P-150(37'C)and by wild-typeP-150 (37'C), and theeffect of S-150(37°)oninvitroprotein synthesis by P-150(379C).Anequal numberof cellswasinfected
witheither thets261-bmutantorwild-type virus at37'Cfor 11 h. The P-150(37'C) and the S-150 (37'C)were
prepared from these infected cells. Theassayprocedureswere asdescribed in Fig. 3 and 4. Incubation ofeach
reaction mixturewasat309C for 100 min. (Upper panels) Protein synthesis in vitro by ts261-b P-150 (37'C). Proteinconcentration ofts261-b P-150(379C) addedtoeach reaction mixturewas1.3mg.(Leftpanel)Kinetics
of incorporation ofL-[P5S]methionineintohot acid-insolublematerial. A sample of 20p1 fromeachreaction
mixturewaswithdrawnatthe times indicated. Theamountof hot acid-insoluble radioactivityateach time point isexpressedasthe totalamountpresent in 480 y1 ofeach reaction mixtureafter subtraction ofthe corresponding valueat0time:(D) without S-150(37'C);(E)withwild-typeS-150(37'C) (0.9mgofprotein); (F) with ts261-b S-150 (37'C) (0.8mgof protein). (D, E, and F) Electrophoreticanalysis ofmutant viral
polypeptides synthesizedin480piof each reactionmixture. 3Hlabel in eachpaneldenotespolypeptidesfrom
thewild-type virionsubjectedtocoelectrophoresis.(Lowerpanels) Protein synthesis in vitro by wild-type P-150
(37TC).Protein concentration of wild-type P-150 (379C) addedtoeach reaction mixture was1.4 mg. (Left
panel) Kinetics of incorporation of 35S label: (G) without S-150(370C);(H) with wild-typeS-150(37'C)(0.9
mgof protein); (I) withts-261-b S-150 (379C) (0.8mgof protein). (G, H, and I) Electrophoretic analysis of
wild-typeviralpolypeptidessynthesizedin480 ofeach reaction mixture.
results from the supplement ofsome necessary
soluble components for elongation of the
pep-tidechains,sothatsynthesis of thefulllength
ofthe viralpolypeptides has been accelerated.
The same wild-type S-150, however, was not
effective on thesynthesis of the mutant viral
polypeptides (Fig. 5E), suggestingafunctional
defect(s) of themutantP-150(37°C).
Figures5F and I show that themutantS-i150 (3700) obtained fromtsmutant-infected cellsat 3700 has essentially no stimulatory effect on
the synthesis of both wild-type and mutant
viralpolypeptidesof all size classes. This could
mean thatsomenecessarysoluble components are missing, or atype of inhibitors) may be
presentin the mutantS-150 (3700). When the
ts261-b P-150 (370C) was prepared from cells
infected for7.25 hat370Cand assayedat300C
inthesame mannerand under thesame
condi-tions,the results obtainedweresimilarto those
shown inFig. 5D, E, and F. These dataimply
that the mutant virus-specific polysomes
formed in vivo at 37C have alreadybeen
im-pairedasearlyas7 hafterinfection. The levels
15
°10
5
iilo5
0
6
4
2 :c7
E
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.508.67.451.66.375.2]40 IKEGAMI
ofinvivosynthesis at370C ofthe mutant virus-specific ss RNAs of the three size classeswere
20 to 25% of thatsynthesized by the wild-type virusthroughout the infective cycle (Table2), and it is assumed that about one-half of the amounts ofthe mutant ss RNAs synthesized would bethe mutant viral mRNA's,whichare
associated with polysomes and monosomes
present inthe P-150, sothat the observed low levelof theprotein-synthesizing capacity of the ts261-b P-150 (370C) would stem from the re-duced synthesis of themutantviral mRNA's. In addition, the finding that the mutant P-150 (370C) doesnotrespondtothe supplement of the wild-type S-150(3700)suggeststhat themutant viral mRNA's associated with polysomes or monosomes may be nicked or may have an abnormal secondary structure at
370C,
sothat synthesis of thefulllengthoftheviral polypep-tides ispresumably blocked.Methylationof mutant viral mRNA's. Most viral andcellularmRNA's have been foundto be blocked and methylated at the 5'-ternini (15, 16, 35, 38, 40),andanimportantrole of the blocked and methylated 5'-terminal structure
inefficienttranslationhas been reported(3, 4). Inregard totheinhibitionofviral protein syn-thesisinmutant-infected cellsat
370C,
one pos-sibility was that there might be a defect in methylation of the mutant viral mRNAs syn-thesized during the infective cycleat370C.
This wasexaminedinthe followingway.(i)Invivo methylation. The mutant-infected cells were labeled with L-[methyl-3H]methio-nineand [14C]uridineat
370C
for 9h(2to 11h after infection). The virus-specific ss RNAs were isolated from the cytoplasmic extract, purified, and then analyzed by velocity gra-dient centrifugation. Figure 6 (left) shows the sedimentation profile. The mutant virus-specificss RNAs of the three size classes1, m, and s werelabeled with both3H and'4C.
The complete hydrolysis of the labeled RNAs by pancreatic RNase(3 ug/ml) confirmed that the methyl-3H label had incorporatedintothemu-tantssRNAsofthethreesizeclasses.Asimilar sedimentation profile was also observedin an
analysis of the wild-type viral ss RNAs synthe-sizedinvivoat37°C underthe samecondition, except thatthelabelingof infected cellswasfor
5 h (6 to 11 h). Inboth analyses, the relative
amounts of the methyl-3H label incorporated
into the s-size RNA was slightly less than the expectedproportions onthe basis ofaninverse relationship to the molecular size of the respec-tive
1,
m, andsclasses.(ii) In vitro methylation. To determine whether the mutant subviral particles derived from infected cells at
370C
are capable ofsyn-J.
thesizing the methylated ss RNAs in vitro, a cytoplasmic pelletwas obtained from mutant-infected cells at370Cfor 9h,and it was used as a source ofthe mutantsubviralparticles. The material inthe pellet was treated with chymo-trypsinandincubatedat370C for 90 min in the presence of the optimal concentrations of all components describedbyShatkin (40). The
re-action mixture was thencentrifuged, and the sedimentable material wasseparatedfrom the supernatant. Under this assaycondition,most of themutant ssRNAs
synthesized
were associ-ated with the sedimentablematerial, and little was present in the supernatant fraction. The centerpanel shows thesedimentationprofileof the in vitro transcriptsisolated from the sedi-mentable material. The methyl-3H and 14C la-belswereincorporatedintothenascent mutantss RNAsof thethreesizeclasses (1, m, ands), and the relative proportionsof both labelswere
inversely related to the molecular size of the respective sizeclass. Thischaracteristic pattern of methyl-3H incorporation indicates that the mutant subviral particles present in cells in-fected at 370C are capable ofmethylatingthe mutant ssRNAsintheprocessofinvitro
tran-scription.
In vitro transcription by the chymotrypsin-treated wild-type virion was carried out for comparison. The reaction mixture was incu-bated for 60 min at
450C,
the optimal assay temperaturefor thewild-type viral cores. Un-der thiscondition, the wild-typessRNAstran-scribed were released from the core
particles
during the period of theassay.The sedimenta-tionprofile shownintheright panelwas simi-lartothatreported by Shatkin (40). These data inFig. 6 supportthe conclusion that methyla-tionof themutant ssRNAssynthesized in vivo
at370C is not restricted.
Effect of temperature shift-down of mu-tant-infected cells. The inhibition of produc-tionof the infectious mutant virus can be re-versed by shifting the temperature of mutant-infectedcells from 37 to300C,and the maximal yield of infectiousvirus can be obtained. It is of interest to ascertain whether the protein-syn-thesizing ability of the mutant virus-specific polysomes has been restored in these shift-down cells. Two sets of an equal number of mutant-infected cells were maintained at
370C
for 11 h. One set was harvested at 11 h. The
other set was shifted down to 30°C and was maintained for an additional4h. TheP-150 was prepared from each set of the mutant-infected cells,and theprotein-synthesizing ability ofthe P-1i0 was assayed in vitro at
300C
in theab-sence of the S-150. The results are shown in
Fig. 7. Theprotein-synthesizing ability of the
on November 10, 2019 by guest
http://jvi.asm.org/
REOVIRUS
ts 261-b mutant virusspecificssRNA's Invivo at370 In vitroat 370
Wild-typevirusspecificssRNA's invitro at450
25r- I m s
H |
20H
5'
4
1
'CQJ
E
1:0
100RNose
0 10 20 30
I m s
I-I MZ
rII
11
'C.
Ec
-
>t
w
t
Il
0 10 20 30
Fraction number
0 10 20 30
FIG. 6. Velocitysedimentation analysis ofts261-bmutantvirus-specificsingle-strandedRNAssynthesized invivo and in vitroat37`C. (Left) ts261-bmutantvirus-specificssRNAssynthesized andmethylated invivo
at 370C. The mutant-infected cells (108) were maintained at 370C in methionine-free spinner medium
supplemented with 20 mM sodium formate, 20 puMguanosine, 20 PM adenosine, and 5%fetal bovine
serum.Infectedcells were labeled with ['4C]uridine (50 PCi) andL-[methyl-3H]methionine (2.2 mCi) for
9 h (2to11 hafterinfection). ActinomycinD(15pg) waspresentfor2 hpriortoinfectionandthroughout the labelingperiod. Theprocedures for extraction andpurification ofssRNAsand foranalysis of velocity
gradientcentrifugation were as described in Materials and Methods and in Table 2. Hydrolysis ofRNAs
ineachfraction bypancreatic RNase (3 pglml) was carried outas described by Gomatos (18). (Center) ts261-b mutant viral ss RNAs synthesized and methylated in vitro at370C. The cytoplasmic pellet was
obtainedfrom cellsinfectedat370C for9 h with the ts261-b mutant in thepresenceofactinomycin D. The pelletwasresuspendedin0.01 MTris (pH 8.1)andwasusedasasourceofmutantsubviralparticles. The detailed procedures for in vitro synthesis of RNAsandforextraction andpurification ofssRNAsfrom the
assaymixturewere asdescribedinMaterials and Methods. In vitroassaywasat370C for90min,and the reaction mixturewascentrifugedat17,000rpmfor30min.ThessRNAswereextracted andpurified from
pelletedmaterials andanalyzed by velocity gradient centrifugation. (Right) Wild-typeviralssRNAs synthe-sizedinvitroat450C byviralcoresderivedfromthepurified wild-typevirus. Theproceduresusedforinvitro transcription and methylation by wild-typeviralcores werethesame asthosedescribedbyShatkin(40).The ssRNAs releasedasfree fromviralcoreswerepurifiedasdescribed inMaterialsand Methods.
ts261-b P-150 from the shift-down cells
in-creased almost 10-fold, and the ability to
syn-thesize thepolypeptides ofo- sizeand those of
140,000 to 120,000 daltons was markedly
re-stored (Fig. 7, center). These data imply that
the mutantvirus-specific polysomes have been
modifiedinafunctionally activestructure
dur-ing theperiod of shift-down incubation atthe
permissive temperature. When thesameP-150
fromtheshift-down cellswasassayedat300Cin
thepresenceofthets261-bS-150 (370C) (Fig. 7,
lowerpanel), theprotein-synthesizingability of
theP-150decreased by 70to80%suggesting the
presenceofaninhibitor(s) in themutantS-150
(370C).
Analysis ofpostribosomalsupernatant.The
S-150 from cells infected with the ts261-b
mu-tant at 370C did not manifest a stimulatory
effect on the viralprotein synthesis catalyzed
by the P-150 (Fig. 5, 7). Its effect was rather
inhibitory insomecases. Asfor the nature ofa
components)responsible for this inhibitory
ef-fect, thets261-b S-150 (370C)might contain an
increasedamountofanenzyme,likeanuclease
that hydrolyzed the viral mRNA's or cleaved
the virus-specific endogenous polysomes. For
elucidation of thisproblem,anexperimentwas
carriedout todetermine theextentof
hydroly-sisofthe free virus-specific ssRNAsduring in
vitroincubation withanS-150 sample. Three
S-I
1
15e 10
Q.
5
VOL. 22,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.508.68.454.68.328.2]42 IKEGAMI
[IiProteinsynthesisin vitro at30° byts 261-b P-150 obtained from cells infected for11 hrs at370
Q) 5_- 10 H H
a 6
u 0 20 40 60 80 100 0 20 40 60 80 100 120 Time (minutes) Mobility (mm)
[21 Protein synthesis in vitro at 30° by ts 261-b P-150 obtained from the infectedcells incubated for 4 hrs at300 after temperature shift down from 371 at11 hrs.
AdditionofS-150 fromncellsinfectedwith ts261-bat37°to the261-b P-150frominfected cells after shiftdown.
10 20 h4
5 010 2
0 20 40 60 80 100 0 20 40 60 80 100 120 Time(minutes) Mobility (mm)
FIG. 7. Restoration ofprotein-synthesizingability of ts261-b P-150 from mutant-infected cells after shifting down to 300C from 370C. Two sets ofan
equal number of cellswereinfected with ts261-b
mu-tantat370C. At 11 hafter infection, the ts261-b P-150 (370C) wasprepared from one setof infected cells. Anothersetof infected cellswas transferredto30'C and incubated for an additional 4 hat300C. The ts261-bP-150(370C/300C)wasprepared from it. The
assay and the subsequent analysis of the products
werecarriedoutinthesame manner asdescribed in
thelegendtoFig.5.(Top)Protein-synthesizing abil-ity invitroat30°C ofts261-b P-150 (37°C) (1.3mgof
protein) inthe absence of S-150 (370C) and electro-phoretic analysis of the in vitro product. (Center) Protein-synthesizing ability in vitroat30°C of ts261-bP-150 (37°C/30°C) in the absence of ts261-b S-150 (370C) andelectropherogram of thein vitroproduct. 3Hlabeldenotespolypeptides from the wild-type vir-ionsubjectedtocoelectrophoresis. (Bottom) Protein-synthesizing ability of ts261-b P-150 (370C/30°C) in thepresenceof ts261-b S-150(37°C) and electrophe-rogram of the in vitro product. The ts261-b S-150 (37°C) wasobtainedfromcells infected for 1I hat 37°C, and the protein concentration added to the reactionmixture was0.8mg.
150 samples (ts261-b 150 [370C], wild-type S-150 [370C], and mock-infected S-150 [370C]) were prepared from cells infected or mock-in-fected at 370C for 11 h. Two kinds of virus-specific ss RNAs (ts261-b ss RNAs and wild-type ss RNAs) were isolated from infected cells that had been exposed to [14C]uridine and
L-[methyl-3H]methioninefor9h at370C.Thesess
RNAs were used asthesubstratefordetection of nuclease activity. Each methylated viral ss RNA wasincubated in vitro at300C with each of theS-150 (370C)samples, and the radioactiv-ity in theacid-precipitable material was deter-mined at the timesindicated. Enzymatic activ-ity that hydrolyzed the virus-specific ss RNAs was present in all three S-150 (370C) samples (Fig. 8). Both ss RNAs were hydrolyzed to a similar extent. Judged by 14C counts (upper panels), 20 to 50% ofthe original radioactive countsremained in theacid-precipitable mole-cules. The observeddifferences in the extent of hydrolysis byeachS-150 appear to result from the differences in the protein concentration of each S-150 tested (see legend to Fig. 8). The methyl-3H-labeled portion of the RNA mole-cules was relatively insensitive to an enzyme presentin theS-150 (lower panels). About80% of themethylatedRNAs,either the intact mol-ecule or the oligonucleotides derived from it, appear tobeprotectedfrom the cleavageof the methyl-3H-containing 5'-terminal nucleotides. From these data, it is reasonable to conclude that the ts261-bS-150(370C)does not contain an increased amount of nuclease, and the inhibi-tory effect of this S-150 (370C) onprotein syn-thesis appears to be attributed to a compo-nent(s)otherthananuclease.Thenatureofthe inhibitory component(s) remains to be eluci-dated.
DISCUSSION
The ts261-b mutant isolated in this labora-torysynthesizesin vivoreduced amounts of the virus-specific ds RNAs and ss RNAs
through-out the infective cycle at the nonpermissive temperature,370C. Despitethe reductioninthe total amounts synthesized, the 10 segments of the mutant ds RNA andthreesizeclasses of the
ss RNAswere synthesized in the proper rela-tiveproportions.The methylation of the newly synthesized ss RNAs was not blocked in the mutant-infected cells at370C.Thecapabilityof the ts261-b mutant subviral particles to tran-scribe the virus-specific ss RNAs in vitro at 370C was comparable to that of the wild-type subviralparticles. As judged from the capabil-ity tosynthesize reovirus-specific ds RNAs and
ssRNAs,the ts261-bmutantcanbeplacedinto the category of dsRNA-positivemutants, like
on November 10, 2019 by guest
http://jvi.asm.org/
[image:12.508.61.246.48.440.2]ts261-b SS RNAs
% [iJ 14C-Uridinelabel
WT SS RNAs
ol [2] C-Uridinelabel
100
t 80
0 z
a 60
x, 40
!0 O
4-0 20
0.§
.q 100'
't 80
%
6040
A
I
~~~~~B
-A.:BufferB:261-bS-150(37 \ C lC: WT S-150 (37) D D:Mock-infectedS-150 (37 D
[3]
3H-methyl
labelh
KAA
BCD
'I ~~~~B
100
80 60
40
A
B
c
_ -uD
201 0
100
80
[4]
3H-methyl
labelA
B 60_
401-201_
I
0 30 60 90 0 30I 60 90
Minutes
FIG. 8. Detectionof nuclease activityinS-150thathydrolyzes free virus-specificssRNAs. Theanalysisat each timepoint for detection ofnucleaseactivityinaparticularS-150(37°C)wascarried out in the presence
ofprotein-synthesizingmixture inafinalvolumeoflOOg0 inindividual tubes that containedaknownamount
of labeledts261-bssRNA orwild-type ssRNA. ProteinconcentrationsoftheS-150tested were210 pg of ts261-bS-150 (37°C),300pgof wild-type S-150(37°C),and 370pgofthemock-infectedS-150(37C).Inthe controlassay, bufferA was included inplace oftheS-150. The in vitro incubation was at30°C. Samples takenat0, 30, and 90 min weremixed With 2 mlof5% trichloroaceticacid andkeptat0°Cfor30min. Theremainingacid-precipitable,labeled RNAswerecollectedonmembranefilters(Millipore Corp.).Thess
RNAs usedwereisolatedfrom cells infectedand labeled(2 to11 hpostinfection) at37°Candpurifiedas
describedinthelegendtoFig. 6 and in Materials and Methods.
those ofclasses A, B, F, and G, which have been isolated and characterized byFields and
Joklik andtheircollaborators (7, 12, 23, 25).
A major temperature-sensitive restricted
function of the ts261-bmutantwasexpressedat
the translational leveland resulted in the
inhi-bition of theviralproteinsynthesis.
The studies using thein vitro protein-synthe-sizing systemreconstituted withanendogenous
polysomalfraction, P-150, andapostribosomal
supernatant,S-150,from reovirus-infectedcells
demonstratedthat restriction of the viral
pro-tein synthesis in the mutant-infected cells at
3700 wasattributedtothe defectivefunctions of
bothmutantvirus-specific polysomesand
com-ponents present in thepostribosomal
superna-tant.Thesynthesisofthemutantviralproteins
wasrestrictedatsomestep(s)ofelongationand
completion of the peptide chains. Under the
experimental conditions in this study, it was
not clear whether the initiation of the viral
peptidechainswasblocked.
When P-150 and S-150 were obtained from
reovirus-infected cells at the permissive
tem-perature and in vitro proteinsynthesis by the
P-150was assayed intheabsence of the S-150,
the P-150 synthesized mostly polypeptides (43,000to33,000 daltons) of thea size and
poly-peptides of 140,000to120,000 daltons and those
of69,000to55,000daltons. The polypeptides of thelattertwosizeranges arevirusspecific, as
theycanbeprecipitated by thereovirus-specific
immune serum, but their apparentmolecular
weights, determined by theirmobilities in SDS-phosphate gel, donot correspondtoanyof the
virus-specific polypeptides reported previously (31, 32, 45,46). Wheninvitro protein synthesis bytheP-150wasassayedinthepresenceofthe
S-150, theelectrophoreticpatternofthe result-ing product showed polypeptides of88,000 to
70,000daltons and thedisappearance of poly-peptides of69,000to 55,000daltons. Ithas not yetbeen determined whether thesetwoevents
occurred by a direct conversion of the latter
)I--v_
20_
43
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:13.508.119.395.67.342.2]44 IKEGAMI
polypeptides (69,000 to 55,000 daltons) to the polypeptidesof the Asize (88,000to70,000 dal-tons),or whether they are unrelated. The effect of the added S-150 onthe in vitroprotein syn-thesis directed by the active P-150 probably results fromthe supplement ofthe active fac-tors, such as elongation factorsoramino acyl-tRNA synthetases, which are necessary for elongation and completionof thepeptidechain andsome of which mayberesponsible for the regulation oftranslational frequency.It is also conceivablethatapossible effect of the added S-150maybe partly relatedtophosphorylationor glycosylation. As aresult, the mobility ofthe modified viral polypeptides
(/i
size) maybe al-tered, since it has been found that the ,u-size polypeptides derived from thewild-type vision arephosphorylated (27) and glycosylated (26). Cross and Fields have reported that one or two species oftheak-size
polypeptides synthe-sizedinvivo atthe nonpermissive temperature(390C)
by thets mutantsofclassesA, D,F, andG manifestan aberrent electrophoretic mobil-ity in ahigh-resolution PAGE system. The mo-bilityisslightly fasterthan thatof the control, but the altered mobility remains within the range of the migration distances of the
4u-size
polypeptides (8). Thesetsmutants arecapable of synthesizing at 390C most of the reovirus polypeptides intheproperrelative amounts (8, 25). In contrast tothe ts mutants of classes A, F, and G, the ts261-b mutant was unable to synthesize the polypeptides of the sizes corre-sponding to the reovirus-specific capsid and noncapsid polypeptides in the mutant-infected cellsat370C.
Thiswasdue to the impairment of the mutant virus-specific polysomes, which wereunabletorespondtothe additionof active soluble factors that stimulate the synthesis of the completed lengthof the viralpolypeptides.The S-150obtained from the ts261-b mutant-infected cells at
370C
(see Fig. 5 and 7) had no stimulatory effectonthe synthesis of theviral polypeptides of allsizeclasses. The ineffective-nessof this S-150 appears to result from some defective functions of soluble proteins rather than an action of a nuclease, since no increased nuclease activity is found in thisS-150 obtained frommutant-infected cells at370C.
Alternatively, the ineffectiveness of the
mu-tantS-150
(370C)
may be due to a limited avail-ability of soluble components, which are re-quired for an efficient translation. Since high-speed supernatants from wild-typevirus-in-fected cells contain a quantity of virus-specific polypeptides (20;unpublishedobservation),
fur-ther work will be necessary to elucidate a
role(s)inprotein synthesis for reovirus-specific soluble proteins synthesized at
370C
by the ts261-b mutant.In the high-speed supernatantfrom frog vi-rus-infected cells, a newly synthesized frog vi-rus-specific protein kinase has been identified which isultimately assembledintovirion parti-cles. Ininfectedcells,certaints mutantsof this virus fail to synthesize the virus-specific pro-tein kinase at the nonpermissive temperature (41). This evidence from another system sug-gests a possibility of the presence of reovirus-specific protein kinase. A possible role of the proteinkinase inthe regulationof protein syn-thesis has been indirectly demonstratedinthe reticulocyte lysate (29). Underconditions such asthe incubationofthelysateabove30°Cinthe absenceof heminorthe addition of ds RNAs or oxidizedglutathione tolysate, the initiation of the peptide chainis inhibited(2, 6, 9, 10). This inhibition is due tothe formation ofa transla-tional inhibitor, which is formed in the high-speed supernatant (10, 29). The inhibition of peptide chain synthesis by the translational inhibitor can be reversed by the addition of cyclicAMP (10,29)ortheadditionof the active initiationfactors that areinvolvedinbinding of the initiator
Met-tRNAf
to the 40S ribosomal subunits (2, 6, 9), suggesting that the transla-tionalinhibitormayresembleaninactiveform of cyclic AMP-dependent protein kinase and that the phosphorylation of a particular pro-tein(s)mayplayanimportant role inthe regu-lation of proteinsynthesis.The ds RNAs are inducers ofproduction of interferon as well as inhibitors of protein syn-thesis. In reovirus-infected cells, the parental ds RNAs are conserved within the parental subviral particles (42), the progeny ds RNAs
arereplicated withinanewlysynthesized parti-cle structure (39, 44), and free ds RNAs have never been detected inanalysis of cellextract (17, 39). However, it is conceivable that an undetectable amount of free ds RNAs, if they
are present in mutant-infected cells at
370C,
may engage information of a translational in-hibitor similartothat foundinthereticulocyte lysates.
Another consideration is that the inhibition of the synthesis of the ts261-b mutant viral proteinsat
370C
may bedue toanactionof the induced interferon. That this would -not be thecaseissupported by(i) the evidencereported by Lai and Joklik (28) that during the infective cycleat
390C
withanyof thets mutantsof the sixclasses (A, B, C, D, E, andG), the level of interferon production is approximatelyone-sixth of thatproduced bythewild-typevirusat
thesametemperature, and that theamountof interferon producedcorrelates with the yields of the infectiousvirus;and(ii)myfindingsthat during the infective cycle at
370C
with the ts261-b mutant, the synthesis of the cellularon November 10, 2019 by guest
http://jvi.asm.org/
and viral proteins is inhibited and the inhibi-tion of the viralprotein synthesis is reversed by shifting the mutant-infected cells from 37 to 300C.
ACKNOWLEDGMENTS
I thankShirley Keelfor technical assistance. I am in-debtedtoPeterJ.Gomatos andthemembers of this labora-toryformanyhelpful discussions.
Thisinvestigation wassupported by Public Health Ser-vice grantCA-08748 fromthe National Cancer Institute.
LITERATURE CITED
1. Bellamy, A.R., L.Shapiro, J. T. August,and W. K. Joklik.1967.Studies onreovirusRNA.I. Characteri-zationofreovirus genome RNA. J.Mol. Biol.29:1-17. 2. Beuzard, Y., and I.M. London. 1974.The effects of hemin anddouble-strandedRNA on aand8globin synthesisinreticulocyteand Krebs IIascitescell-free systemsand the relationship of these effects to an initiationfactor preparation. Proc. Natl. Acad. Sci. U.S.A.71:2863-2866.
3. Both,G.W.,Y.Furuichi, S.Muthukrishnan, andA.J. Shatkin. 1975. Ribosomebindingtoreovirus mRNA inproteinsynthesisrequires5'-terminal 7-methylgu-anosine. Cell6:185-195.
4. Both,G.W.,Y. Furuichi,S.Muthukrishnan, andA.J. Shatkin. 1976. Effect of 5'-terminal structure and basecomposition onpolyribonucleotide bindingto ri-bosomes.J.Mol. Biol. 104:637-658.
5. Both,G.W.,S.Lavi, and A. J. Shatkin.1975.Synthesis of all thegeneproducts of the reovirusgenome in vivo andin vitro.Cell4:173-180.
6. Clemens, M.J.,E.C.Henshaw,H.Rahamimoff, andI. M. London.1974.Met-tRNAfmet,bindingto40S ribo-somal subunits:asitefor theregulation ofinitiation ofproteinsynthesisby hemin. Proc. Natl. Acad. Sci. U.S.A. 71:2946-2950.
7. Cross, R. K., andB. N. Fields. 1972. Temperature-sensitivemutantsofreovirus type 3:studies onthe synthesisof viral RNA.Virology50:799-809. 8. Cross, R. K., and B. N. Fields. 1976.
Temperature-sensitive mutants orreovirus type 3: evidence for aberrant ul and jA2 polypeptide species. J. Virol. 19:174-179.
9. Darnbrough, C., T.Hunt, and R. J.Jackson.1972. A complexbetween met-tRNAc andnative40Ssubunits inreticulocyte lysates anditsdisappearanceduring incubation withdouble-strandedRNA.Biochem. Bio-phys.Res.Commun.48:1556-1564.
10. Ernst, V. D., D. H. Levin, R. S. Ranu, and I. M. London. 1975.Controlof proteinsynthesisin reticu-locyte lysates: effects of3':5'-cyclic AMP, ATP, and GTP oninhibitions inducedbyheme-deficiency, dou-ble-stranded RNA, andareticulocyte translational inhibitor. Proc.Natl. Acad. Sci.U.S.A. 73:1112-1116. 11. Fields, B.N. 1971. Temperature-sensitive mutantsof
reovirustype 3: features ofgenetic recombination. Virology46:142-148.
12. Fields, B.N., and W. K. Joklik. 1969. Isolation and preliminarygeneticand biochemical characterization oftemperature-sensitivemutantsof reovirus. Virol-ogy 37:335-342.
13. Fields, B. N., R. Laskov, and M. D. Scharff. 1970. Temperature-sensitive mutants of reovirus type 3:
studies onthesynthesis of viral peptides. Virology 50:209-215.
14. Fields, B. N., C. S. Raine, and S. G. Baum. 1971. Temperature-sensitive mutants of reovirus type 3:
defects in viral maturation asstudiedby immunoflu-orescenceand electronmicroscopy.Virology 43:569-578.
15. Furuichi,Y., and K-I.Miura.1975.A blockedstructure at the 5' terminus of mRNA fromcytoplasmic polyhe-drosis virus. Nature (London) 253:374-375. 16. Furuichi, Y., M. Morgan, S. Muthukrishnan, and A. J.
Shatkin.1975.ReovirusmessengerRNA containsa methylated, blocked 5'-terminal structure: mTG(5') ppp(5') GmpCp. Proc. Natl. Acad. Sci. U.S.A. 72:363-366.
17. Gomatos, P. J. 1967. RNA synthesis in reovirus-in-fectedL929 mousefibroblasts. Proc.Natl.Acad. Sci. U.S.A. 58:1798-1805.
18. Gomatos,P. J. 1968.Reovirus-specific,single-stranded RNA'ssynthesizedin vitrowith enzymepurifiedfrom reovirus-infectedcells. J. Mol. Biol. 37:423-439. 19. Gomatos, P. J., and I. Tamm. 1963. The secondary
structure ofreovirus RNA. Proc. Natl. Acad. Sci. U.S.A.49:707-714.
20. Huismans,H., andW.K.Joklik.1976.Reovirus-coded polypeptidesininfected cells: isolation oftwonative monomeric polypeptides with affinity for single-stranded and double-single-strandedRNA, respectively. Vi-rology70:411-424.
21. Ikegami,N., andP.J. Gomatos. 1968. Temperature-sensitive conditional-lethal mutants ofreovirus 3. Virology 36:447-458.
22. Ikegami,N., andP.J.Gomatos.1972.Inhibitionof host and viralproteinsynthesis duringinfection at non-permissivetemperaturewithts mutantsof reovirus 3.Virology47:306-319.
23. Ito, Y., and W. K.Joklik. 1972.Temperature-sensitive mutantsofreovirus. I.Patternsofgene expression by mutantsof groups C, D,andE.Virology50:189-201. 24. Ito, Y.,andW. K.Joklik.1972.Temperature-sensitive mutants of reovirus. II. Anomalous electrophoretic migrationofcertainhybridRNA moleculescomposed ofmutantplus strand and wild-typeminusstrands. Virology59:202-208.
25. Joklik, W. K.1974. Reproductionof reoviridae,p. 231-334.In H. Frankel-Conrat andR. R. Wagner(ed.), Comprehensive virology, vol. 2. Plenum Press, New York.
26. Krystal, G., L. Perrault, and A. F. Graham. 1976. Evidence for a glycoprotein in reovirus. Virology 72:308-321.
27. Krystal, G., P. Winn, S. Millward, and S. Sakuma. 1975.Evidence forphosphoproteinsinreovirus. Virol-ogy64:505-512.
28. Lai, M-H. T., andW.K.Joklik.1973.The induction of interferonby temperature-sensitivemutantsof reovi-rus, uv-irradiated reovirus, and subviral reovirus particles.Virology51:191-204.
29. Levin, D. H., R. S.Ranu,V.Ernst, M. A.Fifer, andI. M. London. 1975. Associationofa cyclic AMP-de-pendent protein kinase withapurified translational inhibitor isolated from hemin-deficient rabbit reticu-locyte lysates. Proc. Natl. Acad. Sci. U.S.A. 72:4849-4853.
30. Lowry,0. H.,N.J.Rosebrough,A. L.Farr, and R.J. Randall. 1951.Proteinmeasurementwiththe Folin phenolreagent. J.Biol. Chem. 193:263-275. 31. McDowell,M. J.,andW. K. Joklik. 1971.An in vitro
proteinsynthesizingsystemfrommouse Lfibroblasts infected withreovirus.Virology 45:724-733. 32. McDowell, M. J., W. K.Joklik, L. Villa-Komaroff, and
H. F. Lodish. 1972.Translation of reovirus messen-ger RNAssynthesizedin vitro intoreovirus polypep-tidesbyseveralmammalian cell-freeextracts. Proc. Natl.Acad.Sci. U.S.A.69:2649-2653.
33. Matauhisa,T., and W. K.Joklik. 1974.
Temperature-sensitivemutantsof reovirus. V. Studies on the na-tureof thetemperature-sensitivelesion of the group C mutant ts 447.Virology 60:380-389.
34. Morgan, E. M., and H. J. Zweerink. 1974. Reovirus morphogenesis. Core-like particles in cells infected at
on November 10, 2019 by guest
http://jvi.asm.org/
46 IKEGAMI
390 with wild-type reovirusand temperature-sensi-tivemutantsofgroupsBand G. Virology 59:556-565.
35. Moyer,S. A., G. Abraham,R. Adler, and A.K. Baner-jee.1975.Methylated and blocked 5'termini in vesic-ularstomatitisvirus in vivo mRNAs.Cell5:59-67. 36. Nuss,D. L., Y.Furuichi, G. Koch, and A. J. Shatkin.
1975. Detection inHeLa cellextractsofa7-methyl guanosine specific enzyme activity that cleaves m7GpppN"'. Cell6:21-27.
37. Penman, S. 1966. RNA metabolism in the HeLacell nucleus.J.Mol. Biol.17:117-130.
38. Perry, R. P.,D. E.Kelley,K.Friderici, andF.
Rott-man. 1975. The methylated constituents of Lcell
messengerRNA:evidence foranunusual clusterat
the5' terminus. Cell 4:387-394.
39. Schonberg, M.,S. C.Silberstein, D.H. Levin, andG. Acs.1971.Asynchronous synthesisof the
complemen-tary strands of the reovirus genome. Proc. Natl. Acad.Sci. U.S.A. 68:505-508.
40. Shatkin, A. J. 1974. MethylatedmessengerRNA
syn-thesisin vitrobypurifiedreovirus.Proc. Natl. Acad.
Sci. U.S.A. 71:3204-3207.
41. Silberstein, H.,andJ. T.August. 1976. Characteriza-tion ofa virion proteinkinaseas avirus-specified enzyme.J.Biol.Chem. 251:3185-3190.
42. Silverstein,S. C.,M.Schonberg,D. H.Levin, and G. Acs. 1970. The reovirusreplicative cycle:
conserva-tionofparentalRNAandprotein.Proc. Natl.Acad. Sci.U.S.A.67:275-281.
43. Spandidos,D.A.,G.Krystal,and A. F. Graham. 1976. Regulated transcription of thegenomesofdefective
virionsandtemperature-sensitivemutantsof
reovi-rus.J.Virol. 18:7-19.
44. Zweerink,H.J.,Y. Ito, and T.Matsuhisa.1972. Syn-thesis of reovirus double-stranded RNA within vir-ion-likeparticles. Virology50:349-358.
45. Zweerink, H.J.,and W.K.Joklik.1970.Studiesonthe
intracellularsynthesis of reovirus-specifiedproteins. Virology 41:601-518.
46. Zweerink,H.J.,J.McDowell,andW.K.Joklik.1971. Essential and nonessential noncapsidreovirus
pro-teins.Virology 45:716-723.
on November 10, 2019 by guest
http://jvi.asm.org/

![FIG.1.and[14C]uridineanalyzedanddescribedofspecificobtainedMethodsmA/gelreovirusgel Polyacrylamide gel electropherogram of ds RNAs synthesized in vivo at 37°C](https://thumb-us.123doks.com/thumbv2/123dok_us/1546350.107167/5.508.81.226.55.344/fig-uridineanalyzedanddescribedofspecificobtainedmethodsma-gelreovirusgel-polyacrylamide-electropherogram-rnas-synthesized-vivo.webp)

![FIG. 3.Eachsample[35S]methionineh.andinMaterialsofvirionpiimmune The the reovirus-specific Reovirus polypeptides synthesized in vitro by wild-type P-150 (300C) and by ts261-b P-150 (300 P-150 (3090) prepared from cells infected with wild-type virus )](https://thumb-us.123doks.com/thumbv2/123dok_us/1546350.107167/7.508.147.379.62.461/eachsample-methionineh-andinmaterialsofvirionpiimmune-reovirus-reovirus-polypeptides-synthesized-infected.webp)