Vol. 62, No. 5 JOURNAL OF VIROLOGY, May1988, p. 1486-1494
0022-538X/88/051486-09$02.00/0
Copyright X) 1988, American Society for Microbiology
A
Herpes
Simplex Virus
Mutant
in
Which Glycoprotein D
Sequences Are
Replaced by
P-Galactosidase
Sequences
Binds
to
but Is Unable To Penetrate into Cells
MICHAEL W. LIGASANDDAVID C. JOHNSON*
MolecularVirology and Immunology Program, Department of Pathology, McMasterUniversity, Hamilton, Ontario, Canada L8N 3Z5
Received4November1987/Accepted11January1988
Herpes simplex virus(HSV) glycoprotein gDisamajorcomponentof the virion envelope and is thoughtto
play animportant role in the initial stagesof viral infection and stimulates the production of high titers of neutralizing antibodies. We assumed that gD playsanessential role in virusreplication, andsotocomplement viruses with mutations in the gDgene weconstructedacellline, denotedVD60, which is capable ofexpressing high levels of gD after infection with HSV. A recombinant virus, designated
F-gDoi,
in which sequencesencoding gD andanonessentialglycoprotein, gI,werereplaced by Escherichia coli ,I-galactosidasesequences, was selected on the basis that it produced blue plaques on VD60 cell monolayers underagarose overlays containing 5-bromo-4-chloro-3-indolyl-I-D-galactopyranoside (X-Gal). F-gDII wasable toreplicate normally oncomplementing VD60 cells. However, F-gD,I wasunabletoformplaquesonnoncomplementing Vero cells. Virionslacking gDwereproduced in normalamountsby Vero cells infected with F-gDfI, and the virusparticles
weredistributed throughout the cytoplasm andonthe cellsurface, suggesting that gD isnotessential forHSV envelopment andegress.Virionslacking gDwereabletobindtocells, butwereunabletoinitiatesynthesis of viral earlypolypeptides. Plaque production of F-gDfI particles lacking gDwasenhancedby polyethyleneglycol
treatment,suggesting that gD is essential for penetration of HSV into cells. Other HSV glycoproteins have been implicated in theentryof virus into cells, and thus thisprocessappearstoinvolvemultiple interactionsatthe cell surface.
Mostenvelopedanimal virusesincorporatealimited
num-ber of virus-encoded glycoproteins into host cell and viral
membranes. These polypeptides areutilized in the
envelop-ment of viral nucleocapsids and subsequentlyin the
recog-nition of and penetration into host cells. Viruses such as
influenza virus and vesicular stomatitis virus utilizeasingle glycoprotein in bindingtoandentering into cells (3, 20,38, 39).With these viruses, endocytosis precedes fusion ofthe virion envelope with the endosome membrane. Viral fusion
activity is triggered by the low pH of endosomes (reviewed in reference 20).
Other enveloped viruses, including Sendai virus, human
immunodeficiency virus, and herpes simplex virus type 1
(HSV-1), apparently enter cells by fusion of the virion
envelope directly withtheplasma membrane (9, 25, 34,38). However, incontrast tothe smallerRNAvirusesmentioned above, HSV-1
specifies
atleast seven membrane glycopro-teins (1, 4, 18, 27, 31). Fourofthesepolypeptides, gC, gE, gG, andgI,
have been found to be dispensable for virusreplication
in cultured cells (12, 18, 19, 23, 28, 37). Ofthe three remaining glycoproteins, gB, gD, and gH, there isevidence from studies involving virus mutants that gB and
gHareessentialforvirus entry intocellsorvirusspread (10, 17, 29; P. J.Desai,P. A.Schaffer, and A. C. Minson, Abstr. 12thInt.HerpesvirusWorkshop, p. 111, 1987). Experiments
involving monoclonal antibodies and liposomes suggested that gD might act as a virus attachment component or in virus entry into cells (8, 9, 12a, 16, 24).However, numerous attempts to isolate viruses with mutations in the gD gene
*Correspondingauthor.
have been unsuccessful, and therefore we have only weak
evidence for its role in viralreplication.
Inthis report we describe the construction ofa cell line which can express HSV-1 gDin an inducible fashion. The
cell linewasusedtoisolatea mutantHSV-1inwhichthegD structural sequences werereplaced by the Escherichia coli
,-galactosidase
gene. Virusparticles lacking gD
werepro-duced innoncomplementingVerocells, and these particles
were able to bind to cells but not to initiate an infection. Therefore, gD is essential for penetration of HSV-1 into cells,aprocesswhichappears tobemore
complicated
than that previously described for small RNA viruses such asvesicular stomatitis virus andinfluenza virus.
MATERIALS ANDMETHODS
Cells and viruses. Vero cells were grown in a minimal
essential medium (a-MEM) (GIBCOLaboratories,
Burling-ton, Ontario, Canada) supplemented with 7% fetal calf
serum (FCS). VD60 cells were maintained in Eagle MEM
lacking
histidine(MEM-his)
supplemented with 0.3 to 1.2 mMhistidinol(Sigma Chemical Co., St.Louis, Mo.)and7%FCS. Prior to infection, VD60cells were passaged at least onceina-MEMcontaining7% FCS. HSV-1 F and HSV-2 G wereobtained fromP. G. Spear, Northwestern University, Chicago, Ill., and were propagated and assayed by plaque
formation on Vero cells. The recombinant virusF-gD, was propagated and assayed on VD60 cells.
Plasmids. PlasmidpSS17 contains a copy of theBamHI J
fragment derived from HSV-1 KOS inserted into pUC19.
pMC1871 (5) was agiftfrom M. J. Casadaban. pTZ19R was
purchased from Pharmacia, Dorval, Quebec, Canada. pSV2HIS containing the Salmonella typhimurium histidinol
dehydrogenase gene under control of the simian virus 40
1486
on November 10, 2019 by guest
http://jvi.asm.org/
HSV-1 GLYCOPROTEIN D MUTANT 1487
earlypromoter and flanked by simian virus 40 polyadenyla-tion sequences was a generous gift of R. A. Weinberg,
Whitehead Institute, Cambridge, Mass., and will be
de-scribed in more detailelsewhere (S. C. Hartman and R. J.
Mulligan, manuscript in preparation). Plasmid pSV2HISgD contains the BamHI J fragment of HSV-1 derived from
pSS17 and inserted into theBamHI site ofpSV2HIS. Antibodies.The HSV-1 gD-specific monoclonal antibody, II-436, was a gift of P. G. Spear. Monoclonal antibodies 15IB32, which recognizes HSV-1 or HSV-2 gB, and 17aBl,
whichrecognizesHSV-1 and HSV-2 ICP6 (the large subunit
of ribonucleotide reductase) (2), were gifts from S.
Bac-chetti, McMasterUniversity. Polyclonalrabbitserum, made
in rabbits injected withcrystalline HSV-1 thymidine kinase
produced in E. coli, was a gift from W. Summers, Yale
University,New Haven, Conn.
Construction of VD60 cells. Subconfluent Vero cell mono-layers (100-mm dishes) were transfected with 20
jig
ofpSV2HISgD, using the CaPO4 technique (11), treated 4 h
laterfor60 to 70 swith mediumcontaining 15% glycerol and
7% FCS, andthenwashedtwice with medium. After 2 days theconfluentcellmonolayers weretrypsinizedand plated in 10 100-mm dishescontaining MEMminus hissupplemented
with 0.3 mM histidinol. Approximately 16 days later, indi-vidual colonies of cells were trypsinized by using steel
cloningcylinders and seeded in dishes containing selective medium. The transformants were screened by infecting
small dishes of cells with HSV-2, labeling the cells with
[35S]methionine,
and immunoprecipitating HSV-1 gD fromcell extracts by using HSV-1 gD-specific monoclonal
anti-body 11-436. A cell transformant, VD60, which expressed
high levels of HSV-1 gD when infected with HSV-2 was
chosen fromapool ofover 180transformants.
Transfection, marker rescue experiments, and selection of viruses expressing
3-galactosidase.
Infectious HSV-1 DNA was isolated from cytoplasmic nucleocapsids as describedpreviously (30) and cotransfected with appropriate plasmid
DNAbytheprocedure ofGraham and van der Eb (11). At 4 hafterthe DNA wasadded, cellmonolayersweretreated for 60 swith mediumcontaining 7% FCSand 15% glyceroland washedthree times with medium. Viralcytopathiceffect was
observed2to 4dayslater, and the cells wereharvested and
sonicated. Inmarker rescue experiments, virusyields were
assessedinplaqueassayswithVeroand VD60 cells.
Recom-binant viruses abletoexpress,B-galactosidase were isolated by infectingVD60cell monolayers andoverlayingthe
mono-layers36 to 48h laterwith medium containing 0.5% agarose, 5% FCS, and 300 ,ug of
5-bromo-4-chloro-3-indoyl-P-D-galactopyranoside (X-Gal; Boehringer Mannheim Canada Ltd., Dorval, Quebec, Canada) per ml. Blue plaques
ap-peared after18to 40 h.
[35S]methionine
labeling ofcells, immunoprecipitation,andgelelectrophoresis. Monolayers ofVero cells or cells
trans-formed with
pSV2HISgD
in35-mmdisheswereleftuninfec-ted or wereinfected with HSV-1 or HSV-2. After2to 3 h, the cells were labeled with
[35S]methionine
as described previously (13). Cell extracts were madeat 7to 8 h postin-fection,andextracts were sonicated, clarified bycentrifuga-tion,andmixedwith mouseascitesfluidor rabbit serum and
protein A-Sepharose asdescribed previously (13). Samples
ofprecipitated proteinswereelectrophoresed in8.5%
N,N'-diallytartardiamide cross-linked polyacrylamide gels,
in-fused with2,5-diphenyloxazole, dried, andexposed to XAR film (Eastman Kodak Co., Rochester, N.Y.) as described previously (13).
Purification of labeled virions and adsorption of virions to
cells. Virions were purified from Vero orVD60 cytoplasmic
extracts by usingdextran-T1O (Pharmacia) gradients essen-tially as previously described (16, 32), except that labeling took place from 3 to 24 hpostinfectionin medium 199lacking
methionine and supplemented with 2% FCS and 20 p.Ci of
[35S]methionine per ml and fewer cells per gradient were used (2 x
107
to 4x107
cells pergradient). To collect labeled virus, peak fractionscontaininglabeled virions were pooled, diluted 10-fold in 1 mMphosphatebuffer, and centrifuged at4°C for2 h at 25,000 rpm in aTi5Orotor(Beckman Instru-ments, Inc., Fullerton, Calif.). The pelleted virus was
sus-pended, Dounce homogenized briefly, diluted in medium, and storedat 4°C.Replicate culturesofVero cells(orhuman
R970-5 cells in other experiments) in 24-well dishes were exposed to radiolabeled virus diluted in medium containing
2%
FCS at 37°C, and at appropriate times the mediumcontaining unadsorbed virus was removed, the cells were washed twice, and cell extracts were made by using
Tris-saline (50 mM Tris hydrochloride (pH 7.5), 100 mM NaCI) containing0.5% sodium dodecyl sulfate. Fractions contain-ing unadsorbed virus, washes, and cell extracts were indi-vidually dried on glass fiber filters and counted by liquid
scintillation spectrophotometry.
Electronmicroscopy.At20 hpostinfection, Vero or VD60 cells infected with F or
F-gDP
were washed and fixed at roomtemperature with 0.1 Msodium cacodylatebuffer (pH 7.2) containing 2%gluteraldehyde. The cells were scrapedfromthe dishes, collected bycentrifugation, andprocessed
forelectron microscopyasdescribed previously (15).
Polyethyleneglycolenhancementof viruspenetration. Vero cells were infected with F or
F-gDp
for 2 h, washed three times with medium containing 1% pooled human immuno-globulin and once in medium without gamma globulin andharvested immediately or after 24 h. Samples of
virus-infected cells were sonicated, diluted appropriately, and
incubated with VD60 cell monolayers for 2 h at 37°C. The
cells were exposed briefly to polyethylene glycol, washed, and incubated as described previously (29). Plaques were
countedafter 48 h.
RESULTS
Construction of a cell line able to express HSV-1 gD and complement mutant viruses.Ontheassumptionthat gD is an
essential viral polypeptide, we constructed a cell line
con-taining endogenous gD gene copiesthat are expressed fol-lowing HSV infection. Vero cells were transfected with
plasmid pSV2HISgD, which containstheBamHI J fragment
ofHSV-1 (including the gD gene) inserted intothe BamHI
site ofpSV2HIS, avectorcontainingthehistidinol dehydro-genase genefromS.typhimurium under control of the simian virus 40earlypromoter (HartmanandMulligan,in
prepara-tion). Cells expressinghistidinol dehydrogenase areable to grow in medium containing histidinol and lacking histidine (an essential amino acid for most cells) by catalyzing the
oxidation ofhistidinol(toxicto mostcells)tohistidine. Cell
transformants able to grow in mediumcontaining histidinol
andlacking histidine were screened by infecting small dishes
ofthecells with HSV-2 and
immunoprecipitating
the HSV-1 gD expressed from endogenous HSV-1 gD gene copies byusingmonoclonal antibody II-436, whichspecifically recog-nizes HSV-1gD and not HSV-2 gD. One cell transformant,
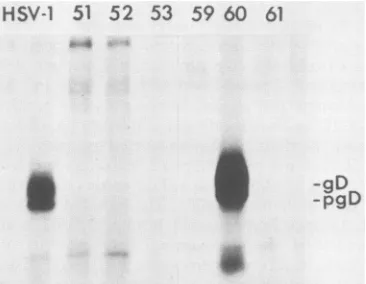
denoted VD60, was found toexpress high levelsofHSV-1 gD after infection with HSV-2(Fig. 1). All of the other 180 cell transformants screenedexpressedundetectable levelsor
muchlower levels ofgD than did VD60 cells and Vero cells
VOL. 62,1988
on November 10, 2019 by guest
http://jvi.asm.org/
1488 LIGAS AND JOHNSON
HSV-1
51
52 53
59
60 61
S
SF
-pgD
-gD
To confirm that the defect in replication of
F-gDP
results fromdisruption of the gD gene and not disruption of thegIgene, weperformed a series of marker rescue experiments. VD60cellsweretransfectedwithF-gD, DNA andplasmids containing an intactgD and gIgene (pSS17), an intactgD geneand adisruptedgIgene(pUS7kan),or an intactgI gene
anda disrupted gD gene (pUS6kan). The titer of the virus
from these transfections was determined using VD60 and Verocells. Plasmids containing an intactgDgene, coupled
toeitheran intact (pSS17) ordisrupted (pUS7kan) gIgene,
efficiently rescued the
F-gDP
mutation, whereas plasmids containing a disrupted gD gene (pUS6kan) or no gD geneFIG. 1. Expression of HSV-1 gD in Vero cell transformants infected with HSV-2. Cell lines VD51, VD52, VD53, VD59,VD60, and VD61 were derivedafter transfection of Vero cells withplasmid pSV2HISgD and selecting for cells abletogrowinmedium contain-ing histidinol andlacking histidine. Thetransformantswereinfected with HSV-2 and labeled with [35S]methionine. For comparison, Vero cells were infected with HSV-1 and labeled (lane HSV-1). HSV-1 gD was immunoprecipitated from extracts of the cells by using monoclonal antibody11-436,which doesnotrecognize HSV-2 gD.
infected with HSV-1. Insubsequentexperiments, wefound
that VD60 cells contain
approximately
110 copies of pSV2HISgD and donot express detectable amounts ofgD priortoinfection(data notshown),presumably becausethe gDpromoteris responsive toviral trans-activating factors.VD60cellsmaintained the propertyofinducibleexpression ofgD afterHSV-2 infection for atleast 20 passages in the
absence of selection. We also found that the inducible expressionofgDincreasesifthe cellsaregrownin medium lacking histidine and containing 1.2 mM histidinol for 12 passages (data not shown). The rate ofdoubling of VD60
cellsin selectiveornonselective mediumissimilartothatof
Vero cells in nonselective medium. We found previously thatthegrowthratesofneomycin-resistantor
methotrexate-resistant transformants were significantly reduced in
selec-tive media.
Construction ofavirusmutant,HSV-1
F-gDO,
in which the gD gene is replaced by theE. colijI-galactosidase
gene. Toisolate a mutant virus lacking the gD gene, we first
con-structed a plasmid, pDGAL11ZK, in which the sequences
encoding thegDpolypeptide andpart oftheneighboring gI
gene were replaced by E. coli
P-galactosidase
sequencesfused togD promoter elements (Fig. 2). VD60 cells were
transfected with pDGAL11ZK and intact HSV-1 F DNA,
andvirusesderivedfrom thetransfectionwere screened by
overlaying plaques formed on VD60 cell monolayers with
agarose containing X-Gal. A recombinant virus,
F-gD3,
whichproduced blue plaques under X-Gal overlay (Fig. 3a), was isolatedand plaque purified.
Southern
blot analysis ofF-gD,B DNA (data not shown) indicated that the virus
contained,-galactosidasesequences inserted inplace ofgD andgI structural sequences (Fig. 3b).
Phenotype of the mutant virus and marker rescue with
plasmids containing the gD gene.
F-gDP
formed plaques on VD60 cells which were of normal size, but was unable toformplaquesonVerocellsorhuman R970 cells, suggesting
that some stage of the replicative cycle of the virus was
disrupted. The substitution mutation disrupts both the gD and gI genes; however, the gI gene has been shown to be
dispensablefor HSV-1 replication in cultured cells (14, 18).
FIG. 2. Construction ofplasmid PDGAL11ZKusedto mutage-nize the HSV-1gD gene. Plasmid pSS17, which contains theBamHI
Jfragment of HSV-1 insertedintopUC19,wasdigestedwithNcoI,
treated with the Klenow fragment of DNA polymerase I, and digested withSmaI,andafragmentcontainingthegD promoterwas
purified and inserted into the SmaI site of pMC1871 directly upstreamof the
0-galactosidase
structural sequences.APstIfrag-mentcontaining the gD-p-galactosidase hybrid genewas excised frompDGAL11 and inserted into thePstIsite of pTZ19R. HSV-1 sequences 3' tothegD gene, extending fromauniqueBalI site in pSS17 toaBamHI site, were excisedfrom pSS17K (aplasmidin
which the BalI site in pSS17 was converted to a KpnI site) and inserted downstream of theP-galactosidasesequencesat aKpnIsite in pDGAL11Z to form pDGAL11ZK. Abbreviations: B, BamHI;
B1, BalI; K, KpnI; N, NcoI; Ps, PstI; S, SmaI; S1, SaII; Apr,
ampicillin resistance gene;Tcr,tetracycline resistance gene; ,-gal.,
P-galactosidasesequence.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.91.274.74.216.2] [image:3.612.318.552.213.574.2]HSV-1 GLYCOPROTEIN D MUTANT 1489
a
UL~~~ ~ ~ ~ ~ ~ ~ ~~
~~'-~~~-~'~
H N F Bal
_
...
HSV-1(F) USBIgo) Us7(gh Us81gE1
B H
HSV-1(F-gDL) 3-Galactosidase Us8(gE)
FIG. 3. Expression of ,-galactosidase by the recombinant virus
F-gD,B and schematic representations oftheDNA sequence arrange-mentsin wild-type HSV-1F and mutant strainF-gDf3.(a)VD60cells were infected with HSV strain F (right) or
F-gDP
(left), and after 36 h the monolayers were overlaid with medium containing agarose andX-Gal. Blue plaques appeared after 36 h. (b) The region of HSV-1 DNA containing the US6 (gD), US7 (gI), and US8 (gE) genes is
depicted for HSV-1 F and recombinant
F-gDP
(21). InF-gDP
P-galactosidase
sequencesreplacegDstructural sequences and partofthegIgenefromanNcolsiteneartheinitiationcodonofgD to a
BalI site in thegI gene. Abbreviations: B, BamHI; Bal, BalI; H,
HindIll; F, FspI; N, NcoI; UL, long unique segment; Us, short
uniquesegment.
(pUC19)wereunable to rescue
F-gDP
(Table 1). Therefore, the lossof the gD gene and not the gI gene leads to a defect in the replication of F-gD, on Vero cells. We find that viruses with disruptions in the gI gene replicate to titers equal to that of wild-type viruses (14; D. C. Johnson,un-published results).
Formation of noninfectious virus particles by
F-gDIO
in noncomplementing cells.AlthoughF-gDP
was unable to form plaques on Vero cells, Vero cells displayed typical HSV cytopathic effects when the cells were infected with a highmultiplicity (5 to 10 PFU per cell) of F-gD,3 grown on
complementing VD60 cells. This suggested that
F-gDP
vi-TABLE 1. Marker rescue of F-gD,Bby plasmidscontaining
intactordisruptedgD andgIgenes Titerb
Plasmid' (Vero/VD60)x 100%
VD60cells Verocells
pSS17 8.2 x 106 4 x 105 4.9
pUS6kan 8.0 x 106 <103 <0.013
pUS7kan 2.0 x 106 8.2 x 104 4.1
pUC19 2.4 x 107 <103 <0.004
"VD60 cells were transfected withF-gDj3 DNAand one of the plasmids indicated. pSS17 containstheBamHI Jfragment of HSV-1subcloned into pUC19; pUS6kanisidenticaltopSS17,exceptthat akanamycingene cassette isinserted intoaHindlIl siteinthe promoter of the gD gene; andpUS7kanis identicaltopSS17,except that akanamycingene cassetteis inserted into a BalI sitein thegIgene.
bTransfectionyieldsweremeasuredby plaqueformation on VD60 or Vero
cells.
rions grown on VD60 cells (and supplied with the gD polypeptide)wereabletoinitiate infection of Vero cells. To
testwhether
F-gDp
virionsgrown on VD60 cellswere able to complete the viral lytic cycle and produceprogenyvirions in Vero cells, weperformed two types of experiments. In the first set ofexperiments radiolabeled virus particles in cell extractswere quantitated by using dextran gradients. VD60 orVero cells were infected withF-gD,B or wild-type strain F and labeledwith[35S]methionine. Extracts from labeled cells were centrifuged on dextran T10 gradients (32), and labeled virions were quantitated. Peaks of labeled virions observedin extracts fromn
F-gDp-infected
Vero cells and F-gDp-infected VD60 cells were only slightly lower than those observed inextractsfromVero cells infected with wild-type strain F(Fig. 4). In the second set of experiments, electron micrographs of virus-infected cells revealed enveloped par-ticles inF-gDp-infected
Vero cells which were structurally and quantitatively similar to those observed in F-gDp-infected VD60 cells or F-F-gDp-infected Vero cells (Fig. 5). Inaddition, weobserved numerous envelopedparticles near to and attached to the surfaces of
F-gDp-infected
Vero cells. Since Vero cells infected with F-gD, do not synthesize detectablequantities ofgD (seebelow), we conclude that gD is notrequired forassembly, maturation, or egress of HSV-1 virions.Binding to cells of virus particles lacking gD. One hypoth-esis which might explain the inability of mutant viruses
bearing gD to form plaques on Vero cells is that gD is
required at a very early stage in the viral lytic cycle, i.e., virus adsorption to or penetration into cells. F-gD,B viruses
containing gD in their envelopes (grown on VD60 cells)
would be able to initiate a single round of infection on Verocells, but the progeny, lackinggD, would be unable to
initiate a second round of infection. To determine whether virions lacking gD could bind to Vero or VD60 cells, Vero cells were infected with F-gD, and labeled with
[35S]methionine,
and radiolabeled virus was prepared ondextran gradients. Vero cells were incubated with labeled
28
24
20
16
12
U
..t
5L_.,.".4r
10 15 20 25 30
Fraction number
FIG. 4. Virus particles produced in cells infected with F-gDf3.
Vero cells or VD60 cells were infected with HSV-1 F or HSV-1
F-gD,p,
labeled with[35S]methionine, and harvested after20to24h,and the virus particles were purified on dextran gradients. The trichloroacetic acid-precipitable label in gradient fractions was
quantitated,andonlyfractionscontainingradiolabeled virionswere
plotted. Symbols: 0, Vero cells infected with F; O, Vero cells infected with F-gD,;*, VD60 cells infected withF-gD,B.
VOL. 62, 1988
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.57.295.69.285.2] [image:4.612.318.554.470.649.2] [image:4.612.56.297.588.660.2]1490 LIGAS AND JOHNSON
'..-r
.,*.W
.4,;l:
I. ~s.
...
*4$.,j- 'j
4"..4Ka . ;iv
rk t ,;w.%w..;UP
vi
S;'ss~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~.If
^!'s'Xww
,
44,
-r~ ~ ~ ~ ~
J. VIROL.
.0
C .0
CUC
042
4)
40.
-o
-u
).-C)3 4)
c'4-'4-4 U
CU
--)
C).
Cd
>.2j
4).0
04), A
~
'i :1.4 z.'.
't. 'v ... s, .01
,z"l- &
-:-'"If." ..
r&,
:
.4w. v
..
v
.0t.
!F ..: ib
on November 10, 2019 by guest
http://jvi.asm.org/
HSV-1 GLYCOPROTEIN D MUTANT 1491
virus particles and washed and the label associated with the cell monolayer was quantitated. Virions purified from
F-gDp-infected
Vero cellswereabletobindtoVero cells and,in other experiments, to human R970-5 cells (results not
shown) asefficientlyaswerewild-type Fvirions and F-gD, virions purified from VD60 cells (Fig. 6). Binding
experi-ments performed at 4°C gave similar results, although the
overallbinding of all viruses waslower(resultsnot shown). Therefore, gD is notessential for attachment orbinding of HSV-1 to cells.
Penetration into cells of F-gD,B virions lacking gD. To determine whether F-gD, virions lacking gD could initiate
certain of the initial stages of virus replication, namely induction of early viral polypeptides, Vero cells were in-fected with F-gDpderived from VeroorVD60 cells and the cells were labeled with [35S]methionine. HSV-1 early
poly-peptides, ICP6 (the large subunit of HSV-1 ribonucleotide reductase), thymidine kinase, gB,andgDwere
immunopre-cipitatedfrom cellextractsby using monoclonal or
polyclo-nal antibodies.
F-gDp
viruses containing gD in the virion envelope were able to induce the synthesis of viral early polypeptides ICP6, thymidine kinase, andgB (Fig. 7, lanes3). However, these viruses cannot induce the synthesis of gD, as one would expect, because the gD gene has been deleted. In contrast,
F-gDP
viruses lacking gD (harvested fromVero cells)wereunabletoinduce the synthesis of viral earlypolypeptides (Fig. 7, lanes 2). Thisresultsuggeststhat virions lacking gDareunable topenetrate into cells.To further examine the possibility that virus particles lacking gDare unableto penetrate intocells and initiate an infection,
F-gDP
virions lacking gD were incubated withVD60 cells, and then thecells were treated with
polyethyl-ene glycol. Polyethylene glycol treatment has previously
been shown to facilitate the entry of gB mutants which
cannotpenetrateinto untreated cells (17,29). Inone
exper-iment, brief exposure ofVD60 cells infected with
F-gDP
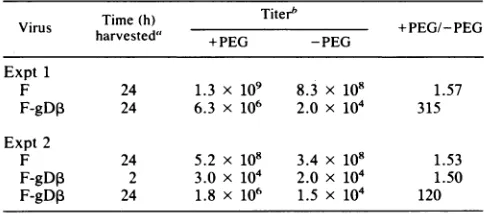
virionslacking gD increased byover300-fold thenumberof
plaques formed (Table 2). In a second experiment, the polyethylene glycol enhancement ofplaque formation was lower,andwefound that
F-gDP
harvested2 hafterinfection of Vero cellsproduced approximately2x 104PFUonVD600
u
0
-1
.0
[image:6.612.317.561.73.227.2]Time(minutes)
FIG. 6. Binding of purified viral particles lacking gD to cells. Virus particles labeled with [35S]methionine were purified on dex-tran gradients from Vero cells infected with F (0). Vero cells infected with
F-gDP
(OI),orVD60 cells infected withF-gD,3 (M)andincubatedwith Verocellsat37°Cin mediumcontaining2% FCS. At various times the cells were washed and radiolabel bound to the cells was quantitated. The results shown are the average oftwo
independent experiments.
ICP6 tk gB gD
1 2 3 1 2 3 1 2 3 1 2 3
97K-
68K-ICP6 gB
-pgB
* gD
a
-pgD-tk
46K-FIG. 7. Expression of early viral polypeptides in cells infected with HSV-1 F or HSV-1 F-gD,B. Vero cells were infected with HSV-1 F harvested from Vero cells (lanes 1), HSV-1
F-gDp
harvestedfromVerocells(lanes 2), orHSV-1F-gDpharvested from VD60 cells (lanes 3). The cells were labeled with [35S]methionine from2to 6hafterinfection, and then cell extracts were mixed with monoclonal antibody 17aBl specific for ICP6, rabbit antibody specific for thymidine kinase (tk), monoclonal antibody15PB5
specific for gD,orantibody 11-436 specific for gD. The immunopre-cipitated proteinswere eluted andelectrophoresed on sodium
do-decylsulfate-polyacrylamide gelsandsubjectedtoautoradiography. Molecularmassstandards areindicatedattheleft.
cellmonolayers.Thissuggeststhat therelatively high
back-grounds in these experiments result from residual F-gD, virionscontaining gD which were used to prepare stocks of virus in Vero cells.
DISCUSSION
HSV-1 gD is a majorcomponent ofthe virion envelope
andinfectedcellmembranes,andvirusneutralizing antibod-ies oftenrecognizegD (7, 12a, 22,26). Numerous attemptsto
isolate viruses with mutations in the gD gene have been unsuccessful. These results suggested that gD might be an
essential component of the HSV-1 envelope or carry out someessentialfunctionduringthereplication cycleof HSV-1. Assuming thatgD is essential for HSV-1 replication, we
beganourattemptsto mutagenizethegDgeneby
construct-ingacellline able tocomplementviruses withmutations in thegD gene. Cellswere transfected withaplasmid
[image:6.612.59.297.503.652.2]contain-ing the gD gene coupled to a novel selectable marker,
TABLE 2. Plaque production of F-gD, particleslackinggD afterpolyethylene glycolenhancement
Time(h)
Titerb
~
+E/EVirus harvested' EG G +PEG/-PEG
Expt1
F 24 1.3 x 109 8.3 x 108 1.57
F-gD, 24 6.3 x 106 2.0 x 104 315
Expt2
F 24 5.2 x 108 3.4 x 108 1.53
F-gD, 2 3.0 x 104 2.0 x 104 1.50
F-gD,3 24 1.8 x 106 1.5 x 104 120 "Vero cells were infected with F or F-gD,B, and infected cells were harvestedatthe indicatedtimes.
bVirus-infected Vero cellswere sonicated, and virus titers were deter-mined onVD60cellswithorwithoutpolyethylene glycol(PEG)treatment.
VOL. 62, 1988
40
It
A.qs,
40 a
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.314.556.585.692.2]1492 LIGAS AND JOHNSON
histidinol
dehydrogenase,
andtransformants abletogrowinmedium
containing
histidinol and lacking histidine wereisolated.
We
found that this selectionsystem
offeredsomeadvantages
overother available methodsinvolving
neomy-cin or
G418, hygromycin,
and methotrexateresistance,
whichareoftenmoretoxictotransformantsor more
expen-sive.
Although
many of our histidinol-resistanttransfor-mants
expressed
detectableamountsofgD, only
oneofthe 180 transformantsscreened, designated VD60,
was able toexpress
gD
at levelscomparable
to those observed in in-fected cells.To assess the effects of
deleting
thegD
gene on virusreplication
and,
for futurestudies,
to allow us torapidly
select forviruses in which the
gD
gene hasbeenmutagenized
in
vitro,
wereplaced
thegD
gene and aportion
of thedispensable
gI
gene with the E. coliP-galactosidase
gene.Recombinant viruses
expressing ,B-galactosidase
werereadily
selectedonthebasis of theirability
toproduce
blueplaques
underagarose-X-Gal overlays. Preliminary
experi-ments also indicate that we can select for viruses which
acquire, by
markertransfer, copies
of thegD
genecontain-ing
site-directed mutationsby isolating
viruses whichpro-duce colorless
plaques.
Inaddition,
we canreadily
screenDNA sequences essential for
gD
functionby using
markertransfer
experiments
suchasthose described inTable 1.The mutant virus
F-gD,
was able toreplicate
on VD60 cells, butwasunabletocomplete
afull round ofinfection inVero cells. Normal amounts of virus
particles
werepro-duced
by
themutantvirus in Verocells inwhichgD
is notexpressed.
These virionsappeared structurally
identical towild-type
virions, although
amore detailed structuralanal-ysis, perhaps involving negative staining
(33),
would berequired
toreveal smalldifferencesinenvelope
structure. Inaddition, F-gDP
virions weredistributed
throughout
thecytoplasm
and on the surfaces ofVero cells. These resultsshow that
gD
is not essential for HSV-1envelopment,
maturation,
oregressto thecellsurface.
Previous studies have
suggested
that gDmight
be anessential component in the attachment of virus to cell surfaces. Virosomes or
lipid
vesicles reconstituted with virionglycoproteins
were able to bind tocells,
and thebinding
was reduced whengD
orgB
was removedfromthelipid
vesicles(16).
Inaddition, polyclonal
and monoclonalantibodies
specific
forgD
inhibited
thebinding
of virionstocells
(9).
In the studiesreported here,
virusparticles
in whichgD
wascompletely
absent from thevirionenvelope
wereabletobindto
monkey
orhumanfibroblastsaswellasvirions
containing gD
were.Therefore,
itappears thatgD is alsonotessentialfor virusattachment,
atleasttothesecells.Itis
conceivable,
however,
thatgD plays
anessentialroleinHSV attachment to other cell types, for
example
neurons.We
plan
to testthispossibility using
virusmutants.Initially,
wefound it somewhatsurprising
thatgD
doesnotplay
an essential role in virusbinding
to culturedcells,
because there is now some evidence thata
majority
oftheother HSV-1
glycoproteins
are also not essential for virusattachment to cells or virus
replication.
Virus mutantsun-abletoexpress
gB (S. Person, personal communication),
gC(12, 28), gG, gE,
orgI
(14,18,
19,23,
37)areabletobind to and in some casesreplicate normally
incultured cells. Thesimplest
explanation
forthese
results isthattwoor moreofthese surface
glycoproteins
canindependently
mediate theinitialattachment of virus tothecell surface. Inadditionto
gD, gB
wouldseem tobealikely
candidate for membershipin this group ofcell attachment
proteins, because
notonlydid virosomes
depleted
ofgB
bind poorly to cells (16) butalso heparin binds to
g1B
(M. L. Parish and P. G. Spear, unpublished results)andblocks virusadsorptiontocells(35, 36). There is also evidence that gC and gEmay be involved in virusbinding tocells(8).Althoughable to bind to cells, virusparticles lacking
gD
wereunableto initiate the synthesis of early viral polypep-tides, suggesting that either entry of virions into cells or some subsequent stage in virus replication was blocked.
Polyethylene glycol treatment dramatically increased the
plaque production of
F-gDP
virionslacking gD on comple-menting VD60 cells.These resultssuggestthat HSV-1gDisessential for virus penetration into cells. However,
F-gDP
virions harvested from Vero cellsaredeficient ingI aswell
as gD. AlthoughgI has been shown to be dispensable for
virusreplication (14, 18)and aplasmid containing anintact
gD gene and a disrupted gI gene efficiently rescued the
replication
defect inF-gDp,
it ispossible that gI somehowaffects the phenotype of the
F-gDP
virions, perhaps by interacting with gD. We think it unlikely that gD and gIinteract,
because gI complexes with gE to form IgG Fc receptors,whicharedispensable forvirusreplication (14).Inaddition, we have recently characterized virus particles whichcontaingIbut notgD, andthesevirusesareunable to
infectVero cells (M.W. Ligas and D. C. Johnson, unpub-lishedresults).
There is evidence that gB, aswell as gD, is involved in
viruspenetration into cells (17, 29). Monoclonalantibodies to gH prevent virus-induced cell fusion (10), a process
thought
tobeanalogoustovirus entryinto cells,andviruseslacking
gH arenoninfectious (Desaietal., Abstr. 12th Int.Herpesvirus Workshop); thus,
gH mayalso be involved in viruspenetration.
One might suggest that gD, gB, and gHmolecules froma complex which acts as the virus
attach-ment
component
and subsequently induces membrane fu-sion.However,
there is evidence that gB and gD arespatially separated
in the virion envelope (33) and thatgB forms homodimers which are not associated with otherglycoproteins
(6). A more likely hypothesisis thatmultiple interactions occur between individual HSV glycoproteins andanumberof cell surfacecomponents, sothatthevirionenvelope
isbrought
into close proximity to the plasmamembrane and the twomembranes areinduced to fuse. In
this
model,
gD, gB,orgHmay notbe essential fortheinitialbinding
of virustocells,
butmaybe essential for interactions whichsubsequently lead toviruspenetration in additiontobeing directly
involved in the membrane fusion reaction. Fusion may involve conformational changes in the fusionpolypeptide,
aswith thepH-induced
conformational changeinthe influenza virushemaglutinin,and mayberegulated by other
polypeptides.
HSV-induced fusion of infected cells, most easily
ob-served with
syncytial
strains ofvirus,
is thought to be a processanalogoustofusion ofthevirionenvelope with cellmembranes during virus entry into cells (31). Antibodies
directedto gD, as well as to gH, block virus-induced cell fusion(10, 12a, 24). Wefound thatF-gD,Bcausedrapidcell
fusion ofVD60 cells, but did not fuse Vero cells (datanot
shown).
Thisresultimplies
thatgDplaysanessentialrole in cell-cellfusion inadditiontoviruspenetration.It is noteworthy that the gD gene, here shown to be essentialfor virusreplication, isembedded in a cluster of 12 geneswhich makeup the unique S componentofthe virus
genome (21). Eleven of these genes are dispensable for
replication
of HSV-1 in cultured cells (14, 18, 19, 23, 37). Theobservationthat the
majority
ofS-component
genescouldbedeleted ledLongneckerandRoizman(19) to suggest that the
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
HSV-1 GLYCOPROTEIN D MUTANT 1493
S component was acquired as a DNA fragment which subsequently evolved to allow the virus to survive in its environmental niche in the human host. If this hypothesis is
correct, the ancestral herpesvirus did notdependon gDto
enter cells. This scenario predicts alterations in the entry
pathway of HSV following acquisition of the Scomponent.
Alternatively, one might hypothesize that the genes sur-rounding gD have playedanimportant role in the replication of HSVthroughoutmostof the evolution of the virus andyet
thesegenes aredispensable in the limited setting of
labora-tory-cultured cells. There is evidence suggesting that
S-component genes confer a distinct selective advantage on the virus in the human host. We haverecently shown that S-component polypeptides gI and gE formacomplex which acts as an immunoglobulin G Fc receptor and may act to
shield virus-infected cells from the host immune response
(13, 14). Additionally, thegeneencoding gG, whichmapsto
the left ofgD (1, 27), has been implicated in virus replication inthe central nervous system(37). From this perspective it is not surprising that the gD gene is surrounded by genes
encoding functions which are advantageous and may be
essentialfor virus replication in humans.
ACKNOWLEDGMENTS
We are indebted to James Smiley for coaching, especially in matters havingtodo with genetics. We thankSteven Hartman for
suggestingtheuseofpSV2HISand PatriciaSpear,SilviaBacchetti,
and William Summers forgiftsofantibodies.JosephinaMaljarwas
invaluable inhelpingtoputthemanuscript together.
This work was supported by grants from the National Cancer Institute of Canada and the Medical Research Council of Canada. D.C.J. is a research scholar of the National Cancer Institute of Canada.
LITERATURE CITED
1. Ackermann, M., R. Longnecker, B. Roizman, and L. Pereira. 1986. Identification, properties, and gene location ofa novel
glycoprotein specified by herpes simplex virus 1. Virology
150:207-220.
2. Bacchetti, S.,M.J. Evelegh,and B.Muirhead. 1986. Identifica-tion andseparationoftwosubunits of theherpes simplexvirus ribonucleotide reductase. J. Virol. 57:1177-1181.
3. Bishop, D. H. L., P. Repik, J. F. Obijeski, N. F. Moore, and R. R. Wagner. 1975. Restitution of infectivity to spikeless
vesicular stomatitis virus by solubilized viral components. J. Virol. 16:74-84.
4. Buckmaster, E. A., U. Gompels, and A. C. Minson. 1984. Characterization and physical mappingofanHSV-1
glycopro-tein ofapproximately115 x103molecularweight. Virology139:
408-413.
5. Casadaban, M. J., A. Martinez-Arias, S. K. Shapira, and J.
Chou. 1983. 3-Galactosidase gene fusions foranalyzing gene expression in Escherichia coli and yeast. Methods Enzymol.
100:293-308.
6. Claesson-Welsh, L., and P. G.Spear. 1986.Oligomerizationof
herpes simplexvirusglycoprotein B.J. Virol. 60:803-806. 7. Cohen, G. H., B. Dietzschold, M. Ponce deLeon, D. Long,E.
Golub, A. Varrichio, L. Pereira, and R.J. Eisenberg. 1984. Localizationandsynthesisofanantigenicdeterminant ofherpes
simplex virus glycoprotein D that stimulates production of
neutralizing antibody. J. Virol. 49:102-108.
8. Fuller,A.O.,andP.G.Spear.1985.Specificitiesof monoclonal and polyclonal antibodies that inhibit adsorption of herpes simplexvirus tocells and lack ofinhibitionbypotent
neutral-izingantibodies. J. Virol. 55:475-482.
9. Fuller, A. O., and P. G. Spear. 1987. Anti-glycoprotein D antibodies thatpermit adsorptionbutblock infectionby herpes simplex virus 1 prevent virion-cell fusion atthe cell surface. Proc. Natl. Acad. Sci. USA 84:5454-5458.
10. Gompels, U.,andA.Minson. 1986. Thepropertiesandsequence
ofglycoprotein H of herpes simplex virustype1.Virology153:
230-247.
11. Graham, F. L., and A. J. van der Eb.1973. Anewtechnique for theassayof infectivity of adenovirus5DNA.Virology 52:456-467.
12. Heine, J. W., R. W. Honess, E. Cassai, and B. Roizman. 1974.
Proteins specified by herpes simplex virus. XII. The virion polypeptides oftype1 strains.J. Virol. 14:640-651.
12a.Highlander, S. L., S. L. Sutherland, P. J. Gage, D. C. Johnson, M. Levine, and J.C. Glorioso. 1987. Neutralizing monoclonal antibodies specific for herpes simplex virus glycoprotein D
inhibit viruspenetration. J.Virol. 61:3356-3364.
13. Johnson, D. C., and V. Feenstra.1987. Identification ofanovel herpes simplex virustype 1-induced glycoprotein which
com-plexes with gE and binds immunoglobulin. J. Virol. 61:2208-2216.
14. Johnson, D. C., M. C. Frame, M. W. Ligas, A. M. Cross, and N. D.Stow. 1988. Herpessimplex virus immunoglobulin G Fc receptoractivity dependson acomplex oftwoviral glycopro-teins, gE and gI. J. Virol. 62:1347-1354.
15. Johnson, D. C., and P. G. Spear. 1982. Monesin inhibits the processingof herpes simplex virus glycoproteins, their trans-porttothecellsurface, andthe egressof virions from infected cells. J. Virol. 43:1102-1112.
16. Johnson, D.C.,M. Wittels,and P. G.Spear. 1984. Bindingto
cells of virosomes containing herpes simplex virus type 1
glycoproteins and evidence for fusion.J. Virol. 52:238-247.
17. Little, S. P., J. T. Jofre, R. J. Courtney, and P. A. Schaffer.
1981. Avirion-associated glycoprotein essential for infectivity ofherpes simplex virustype 1. Virology 115:149-158. 18. Longnecker, R.,S.Chatterjee, R. J. Whitley, and B. Roizman.
1987. Identification ofa herpes simplex virus 1 glycoprotein
genewithinageneclusterdispensable for growth in cell culture. Proc. Natl. Acad. Sci. USA 84:4303-4307.
19. Longnecker, R., and B. Roizman. 1987. Clustering ofgenes
dispensable for growth in culture in the S componentof the HSV-1gene.Science236:573-576.
20. Marsh,M. 1984. Theentry ofenveloped viruses intocells by endocytosis. Biochem. J.218:1-10.
21. McGeoch, D.J., A. Dolan, S. Donald, and F. J. Rixon. 1985. Sequence determination and geneticcontentof the shortunique region in the genome of herpes simplex virustype 1. J. Mol. Biol. 181:1-13.
22. Minson,A.C.,T.C.Hodgman,P.Digard, D. C. Hancock, S. E.
Bell,and E. A. Buckmaster. 1986. Ananalysis of the biological properties of monoclonal antibodies against glycoproteinD of herpes simplex virus and identification of amino acid substitu-tions that confer resistance to neutralization. J. Gen. Virol. 67:1001-1013.
23. Neidhardt, H., C. H.Schroder,and H.C.Kaerner.1987.Herpes
simplexvirustype 1glycoproteinEisnotindispensable for viral infectivity.J. Virol.61:600-603.
24. Noble,A.G.,G. T.-Y.Lee, R.Spraque,M. L.Parish, and P. G.
Spear. 1983. Anti-gD monoclonalantibodies inhibit cellfusion inducedby herpessimplex virustype 1. Virology129:218-224. 25. Para, M. F., R. B. Baucke, and P. G.Spear.1980. Immunoglob-ulinG(Fc)-bindingreceptors onvirions ofherpes simplex virus
type 1 and transfer of these receptors to the cell surface by infection. J.Virol.34:512-520.
26. Para,M.F., M. L.Parish,A.G.Noble, and P. G. Spear. 1985.
Potentneutralizing activity associated with anti-glycoproteinD
specificityamongmonoclonal antibodies selected forbindingto
herpes simplex virions.J. Virol.55:483-488.
27. Richman, D. D.,A.Buckmaster, S. Bell, C. Hodgman, andA.C. Minson. 1986. Identification ofa new glycoprotein of herpes simplexvirus type 1andgenetic mappingof the gene that codes
for it. J. Virol.57:647-655.
28. Ruyechen,W. T.,L.S.Morse, D. M.Knipe,and B. Roizman. 1979.Moleculargeneticsofherpes simplex virus.II.Mappingof
themajorviralglycoproteinsandofthegeneticlocispecifying
thesocialbehavior of infected cells. J.Virol.29:677-687.
29. Sarmiento, M.,M. Haffey, and P. G. Spear. 1979. Membrane
proteins specified by herpes simplex viruses. III. Role of
VOL.62, 1988
on November 10, 2019 by guest
http://jvi.asm.org/
1494 LIGAS AND JOHNSON
glycoprotein VP7 (B2) in virion infectivity. J. Virol. 29:1149-1158.
30. Smiley,J., B. Fong, and W.-C.Leung. 1981. Construction ofa
doublejointed herpes simplex virus DNA molecule: inverted repeats are required for segment inversion and direct repeats promotedeletions. Virology 113:345-362.
31. Spear, P. G. 1985. Glycoproteins specified by herpes simplex viruses, p. 315-356. In B. Roizman (ed.), The herpesviruses, vol. 3. Plenum Publishing Corp., New York.
32. Spear, P. G., and B. Roizman. 1972. Proteins specified by herpes simplex virus. V. Purification and structural proteins of the herpes virion. J. Virol. 9:143-159.
33. Stannard, L. M., A. 0. Fuller, andP. G.Spear. 1987. Herpes
simplex virus glycoproteins associated with different morpho-logical entities projecting from the virion envelope. J. Gen. Virol. 68:715-725.
34. Stein, B. S., S. D. Gowda, J. D. Lifson, R. C. Penhallow, K. G.
Bensch, and E. G.Engleman. 1987. pH-independent HIVentry into CD4-positive T cells via virus envelope fusiontothe plasma membrane.Cell 49:659-668.
35. Takemoto, K. K., and P. Fabish.1964. Inhibition of herpes virus by natural and synthetic polysaccharide. Proc. Soc. Exp. Biol. Med. 116:140-144.
36. Vaheri, A., and K. Cantrell. 1963. The effect of heparin on
herpes simplex virus. Virology 21:661-662.
37. Weber, P. C., M. Levine, and J. C. Glorioso. 1987. Rapid identification of nonessentialgenesofherpes simplex virustype 1 by Tn5 mutagenesis. Science 236:576-579.
38. White, J., M. Kielian,andA.Helenius. 1983. Membrane fusion proteins of enveloped animal viruses. Quart. Rev. Biophys. 16: 151-195.
39. Wilson, I.A., J. J.Skehel, and D. C. Wiley. 1981. Structure of
the haemagglutininmembrane glycoprotein of influenza virusat 3 Aresolution. Nature(London) 289:366-373.
J. VIROL.