Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Replication and Pathogenicity of Primer Binding Site Mutants
of SL3-3 Murine Leukemia Viruses
ANDERS H. LUND,1JO¨ RG SCHMIDT,2ARNE LUZ,3ANNETTE BALLE SØRENSEN,1MOGENS DUCH,1 ANDFINN SKOU PEDERSEN1,4*
Department of Molecular and Structural Biology1and Department of Medical Microbiology and Immunology,4
University of Aarhus, DK-8000 Aarhus C, Denmark, and GSF Institute for Molecular Virology2and
GSF Institute for Pathology,3Neuherberg, D-85764, Germany
Received 24 November 1998/Accepted 5 April 1999
Retroviral reverse transcription is primed by a cellular tRNA molecule annealed to an 18-bp primer binding
site sequence. The sequence of the primer binding site coincides with that of a negatively actingciselement that
mediates transcriptional silencing of murine leukemia virus (MLV) in undifferentiated embryonic cells. In this
study we test whether SL3-3 MLV can replicate stably using tRNA primers other than the cognate tRNAProand
analyze the effect of altering the primer binding site sequence to match the 3*end of tRNA1Gln, tRNA3Lys, or
tRNA1,2Arg in a mouse pathogenicity model. Contrary to findings from cell culture studies of primer binding
site-modified human immunodeficiency virus type 1 and avian retroviruses, our findings were that SL3-3 MLV
may stably and efficiently replicate with tRNA primers other than tRNAPro. Although lymphoma induction of
the SL3-3 Lys3 mutant was significantly delayed relative to that of the wild-type virus, molecular tumor analysis indicated that all the primer binding site-modified viruses induce T-cell lymphomas similar to those induced by the wild-type virus in terms of frequencies of genomic rearrangements within the T-cell receptor
b-chain, the immunoglobulinklight chain, and the c-myclocus. Whereas none of the mutants were found to
revert to tRNAProprimer utilization, in two tumors resulting from the injection of the SL3-3 Lys3 mutant the
primer binding site was altered to match that of a new primer species, tRNA1,2Lys. In addition, recombination
with endogenous viruses resulting in the generation of recombinant viruses carrying a glutamine primer binding site was detected in the majority of the tumors induced by the SL3-3 Lys3 mutant as well as in two tumors induced by wild-type SL3-3 and the SL3-3 Arg1,2 mutant.
Reverse transcription of the retroviral RNA genome into proviral DNA is a hallmark of retroviruses. Upon infection, the process of reverse transcription is initiated near the 59end of the genomic RNA molecule where the 3918 nucleotides of a tRNA molecule derived from the former host cell are annealed to the primer binding site (PBS) sequence. The PBS sequence mediates two functions of vital importance during reverse tran-scription: it serves as a tRNA binding site and hence facilitates initiation of reverse transcription, and through base paring with a DNA copy of the 18 39nucleotides of the tRNA mole-cule generated during plus-strand DNA synthesis, it mediates the second template switch of reverse transcription (14). The tRNA primer molecule is derived from the host cell tRNA population, and different viruses utilize different tRNA mole-cules as primers for reverse transcription. Murine leukemia viruses (MLV) (36) and human T-cell leukemia virus (41) replicate via a tRNAPro, human immunodeficiency virus (HIV) (44) and mouse mammary tumor virus (27) replicate via a tRNA3Lys, while avian retroviruses (38) replicate via a tRNATrp primer molecule. Within a given virus species the sequence of the PBS, and thus the utilization of a specific tRNA isoaccep-tor, is highly conserved partly due to the inherent conservative mechanism of reverse transcription.
Molecular adaptation of a given retrovirus towards a single tRNA species is likely to be favorable by increasing the rate of correct initiations of reverse transcription and the fidelity of the second template switch. Genetic experiments with
PBS-modified HIV-1 and avian viruses in cell culture have con-firmed preferential tRNA usage in both viruses (11, 21, 45, 47). Likewise, a number of biochemical studies have demonstrated the presence of specific interactions between mature HIV-1 reverse transcriptase (RT) and tRNA3Lys, presumably through RT recognition of the anticodon loop (4, 5, 8, 30, 37). For the avian viruses, tRNATrprecognition by the avian RT dimer has been shown biochemically (3, 5, 15) and base pairings between tRNATrp and the viral genome upstream from the PBS se-quence have been demonstrated in genetic studies of mutant viruses (1, 9). In contrast to HIV and the avian retroviruses, MLV may be less stringent in its use of a tRNA replication primer. Accordingly, we have previously shown that retroviral vectors derived from Akv-MLV may be efficiently transduced with either tRNA1,2Gln or tRNA3Lys (24) or with a genetically engineered tRNA-like primer molecule (25). Previous bio-chemical investigations have not revealed evidence of tRNA primer preferences in MLV (5, 33). Furthermore, analysis of polmutants of Moloney MLV demonstrated that the presence in MLV virions of tRNAProcorrectly annealed to the PBS is independent of products of the polgene (13). Additional in-dications for a less stringent use of tRNA primers in MLV come from sequencing of MLV-related viruses endogenous to the murine genome carrying PBSs that match tRNAGln (10, 31).
Aside from having pivotal functions during reverse tran-scription, the PBS sequence that matches tRNAProhas been found to overlap with a negatively acting cis element that mediates provirus transcriptional silencing in the mouse germ line and in undifferentiated embryonal cell lines (6, 17, 46). Interestingly, it was recently demonstrated that the negatively actingciselement overlapping the proline PBS may affect the * Corresponding author. Mailing address: Department of Molecular
and Structural Biology, University of Aarhus, C. F. Møllers Alle´, Bldg. 130, DK-8000 Aarhus C, Denmark. Phone: 45 8942 3188. Fax: 45 8619 6500. E-mail: [email protected].
6117
on November 9, 2019 by guest
http://jvi.asm.org/
transcriptional activity of retroviral vectors in several lymphoid and myeloid cell types (7). Conceivably, these types of effects of PBS alterations on virus-host interactions might be revealed in mouse models of MLV pathogenesis.
As a first part of such studies, we chose to study the repli-cation and pathogenicity of PBS-modified mutants of the rap-idly disease-inducing virus SL3-3, altered to replicate via tRNA1Gln, tRNA3Lys, or tRNA1,2Arg. The aim of this study was dual: first, to analyze whether PBS-modified viruses can repli-cate stably and efficiently in vivo with tRNA primers other than tRNAPro and, second, to investigate whether the introduced PBS modifications overlapping the negatively acting cis ele-ment influence the pathogenicity of the viruses. SL3-3 MLV is a highly lymphomagenic, nonacutely transforming virus that induces T-cell lymphomas with 100% incidence (35). Depend-ing on the mouse strain, the latency period varies from 2 to 6 months (12, 18). The T-cell lymphomagenic potential of SL3-3 has been mapped to the U3 region of the long terminal repeat (LTR) (18, 20) and is likely mediated primarily through bind-ing of T-cell-specific transcription factors, resultbind-ing in a high rate of replication and rendering the virus a strong insertional mutagen in the T-cell compartment of the hematopoietic sys-tem.
The viruses were derived from a pBR328 subclone of lSL3-95 (19), T464, containing a functional SL3-3 provirus surrounded by sequences of genomic DNA. A deletion mutant, T464(DPvuI), lacking the 59LTR region and part of the leader region was linearized withSpeI and ligated to a 735-bpEco RI-SpeI fragment derived from either pPBS-gln1, pPBS-lys3, or pPBS-arg1,2 and encompassing the 59LTR and 157 bp of the leader region of Akv-MLV. Three SL3-3 replication-compe-tent viruses with mutations in their PBS sequences at Gln1, Lys3, Arg1,2, hereinafter referred to as Gln1, SL3-3-Lys3, and SL3-3-Arg1,2, were thereby created. pPBS-gln1 and lys3 have previously been described (24), and pPBS-arg1,2 is syngeneic to these vectors except in the PBS sequence. Within the leader region SL3-3 differs from Akv-MLV by hav-ing a cytosine at position 163 and adenosine residues at posi-tions 235, 247, and 249. SL3-3 also differs from Akv by having a 2-bp guanosine insertion after position 246. The effect of introducing these leader mutations was analyzed through con-struction of an additional mutant, SL3-3-Pro/Akv UTR, which contains the wild-type (wt) proline-specific PBS and the Akv-specific leader. The viruses were generated by direct transfec-tions of purified ligation reaction mixtures (total amount of DNA, approximately 5 mg) into NIH 3T3 cells by calcium phosphate precipitation, and the emergence of viruses was monitored by assaying the supernatant for RT activity (22). For the generation of wt SL3-3, T464 containing the proviral clone of SL3-3 was transfected into NIH 3T3 cells and is hereafter referred to as SL3-3-Pro. To verify the presence of the PBS mutations, RNA was purified from pelleted virus particles and subjected to RT-PCR followed by direct auto-mated sequencing of a 200-bp region encompassing the PBS. Randomly primed reverse transcription was performed with a First Strand cDNA synthesis kit (Pharmacia), and PCR ampli-fication was done with oligonucleotides matching regions within SL3-3 U3 (primer 1, 59-TCCGAATCGTGGTCTCGCT GATCCTTGG-39, matching SL3-3 nucleotide positions 69 to 96) andgag(primer 2, 59-TAGGGTCAGACTCAGAGGGG TGGT-39, specific for upstream MLV gag sequences and matching Akv positions 677 to 654). Sequencing was per-formed with primer 3 (59-CGCAGGCGCAAAAAGTAGAT GC-39), which is specific for Akv positions 268 to 289. All PCRs in this study were performed in 100ml of PCR buffer (Perkin-Elmer) containing 25 pmol of each primer, 0.2 mM
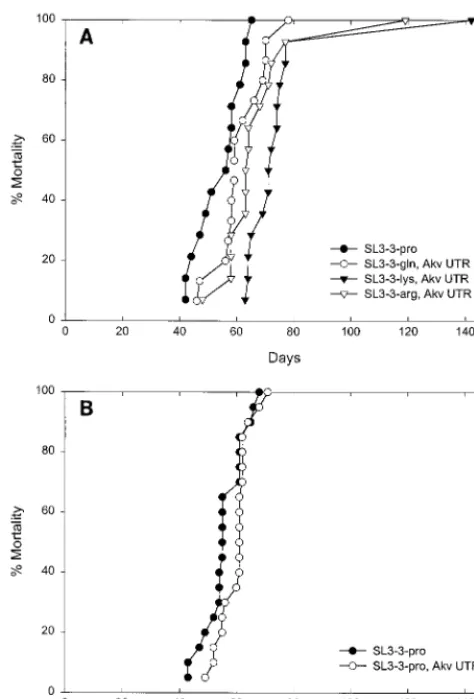
each deoxynucleoside triphosphate, and 3 U of AmpliTaq Gold polymerase (Perkin-Elmer). All viruses were found to replicate stably in cell culture regardless of their PBS sequence (data not shown). To evaluate the amount of infectious parti-cles produced, infectious-center assays were performed with a rabbit-anti-p30 antibody as described previously (40). The viral titers in the supernatants of the NIH 3T3 producer cell lines were determined in several independent assays to be 13106to 13 107, 0.4 3 106 to 0.5 3 106, and 1 3 106 to 83 106 infectious particles per ml of supernatant for 3-Gln1, SL3-3-Lys3, and SL3-3-Arg1,2, respectively, and thus to be within a 1.5-log-unit range of that of wild-type SL3-3-Pro, which was 0.83106to 13106infectious particles per ml of supernatant. To investigate the stability of the introduced PBS mutations in an in vivo model and to test their possible effects upon tumorigenesis, 100-ml aliquots of medium containing infec-tious particles were injected intraperitoneally into newborn inbred NMRI mice. The mice were checked for tumor devel-opment on 5 days of the week, and the mice were killed at the time of apparent illness or tumor development. Tumors were diagnosed on the basis of gross appearance of lymphoid organs as previously described (39) and according to cytologic and anatomic criteria (34). To monitor the development of viremia in the mice 2 weeks postinfection, blood samples (2.5 ml) drawn from the tail vein were analyzed for the presence of infectious particles by infectious-center assays (40). All mice injected with wt SL3-3-Pro and SL3-3-Gln1 carried infectious virus particles in peripheral blood. In contrast, 13 of 14 mice injected with SL3-3-Arg1,2 and 4 of 14 mice injected with SL3-3-Lys3 were found to be viremic at day 14 postinfection at the level of sensitivity of this assay. All viruses tested in this study were highly tumorigenic, giving rise to lymphomas in all of the injected mice (Fig. 1A and Table 1). All viruses induced tumors within a narrow time window of around 60 days, with the exception of one mouse injected with SL3-3-Lys3 and one mouse injected with SL3-3-Arg1,2, in which the tumors were detected at 142 and 119 days postinfection, respectively. Inter-estingly, lymphoma inductions by the PBS-modified viruses were significantly delayed compared to that of the wt virus (log rank test,P,0.05 for SL3-3-Gln1 and SL3-3-Arg1,2 andP, 0.001 for SL3-3-Lys3). Furthermore, tumor induction by the SL3-3-Lys3 mutant was significantly delayed compared to that of the SL3-3-Gln1 mutant (log rank test,P,0.05). To analyze whether the Akv-MLV-specific alterations introduced in the 59 leader region affected the time course of tumor induction, we performed a separate series of animal experiments in which the tumor induction of the SL3-3-Pro/Akv UTR mutant was compared to that of wt SL3-3 (Fig. 1B and Table 1). Since no significant differences in disease induction were found among these viruses, we conclude that the delays in tumor induction observed for the PBS-modified viruses are attributable mainly to the PBS alterations. However, a minor role for the Akv-MLV-specific alterations in the 59UTR cannot be excluded.
To analyze at the molecular level the disease tropism of the mutant viruses, Southern blotting was performed. Genomic DNA purified from tumors was digested withHindIII, blotted, and probed for the presence of genomic rearrangements within the T-cell receptor (TCR)b-chain with two probes specific for the TCR-joining regions (J1 and J2) (2). A representative autoradiogram is shown in Fig. 2. With a few exceptions where the blotting results were inconclusive, the tumors harbored genomic rearrangements within this region and hence repre-sented predominantly T-cell lymphomas (Table 1). We subse-quently tested for rearrangements within the immunoglobulin (Ig)klight chain (23) and found no genomic rearrangements in any of the analyzed tumors, consistent with the pathological
on November 9, 2019 by guest
http://jvi.asm.org/
finding of enlarged thymuses (Table 1). Hence, we conclude that the PBS-modified SL3-3 viruses, like wt SL3-3, induce T-cell lymphomas. The number of integrated ecotropic viruses per tumor was investigated with a probe specific for the eco-tropicenv gene (23). The provirus copy numbers varied be-tween 1 and 7, with no significant mutant-dependent differ-ences (data not shown). The c-mycgene has previously been reported to be frequently involved in T-cell lymphomas
in-duced by SL3-3, with 20 to 25% of tumors showing genomic rearrangements as a result of proviral integration into the c-mycpromoter region (16, 43). With a probe specific for the c-mycpromoter (43), clonal rearrangements within the c-myc promoter were detected in 25% (13 of 52) of the analyzed tumors, being found, for example, in 2 of the 12 tumors in-duced by 3-Gln1 and 4 of the 13 tumors inin-duced by SL3-3-Arg1,2 (Table 1). We therefore conclude that the introduced PBS mutations do not grossly affect tumorigenesis processes related to c-myc.
[image:3.612.55.292.72.421.2]To analyze if the introduced PBS mutations were genetically stable during in vivo replication and tumor induction, purified genomic DNAs from all tumors were subjected to PCR am-plification with primers specific for the ecotropic MLV U3 region (primer 4, 59-GATTCCCAGATGACCGGGGATC-39, which matches Akv positions 8179 to 8201) andgag(primer 2). The resulting ;750-bp amplicons were purified, and both strands of an ;250-bp region encompassing the PBS were directly sequenced with primers 1 and 3 (Table 2). While most of the sequence readouts were clean and allowed unambiguous assignments, direct sequencing of the PCR products from 14 of the tumors repeatedly resulted in poor-quality double se-quences, consistent with sequencing of two different proviral templates. In these cases the PCR products were cloned into pGEM-T (Promega) and 8 to 12 individual subclones were sequenced. Surprisingly, SL3-3-Gln1 was the only virus found to replicate stably in all tumors analyzed whereas alterations within the PBS were detected in individual tumors induced by SL3-3-Pro, SL3-3-Lys3, and SL3-3-Arg1,2 (Table 2). A double
[image:3.612.367.495.75.222.2]FIG. 1. Lymphoma development in inbred NMRI mice injected with wt and mutated SL3-3 MLVs. The cumulative mortality is plotted as a function of time after virus injection. (A) PBS-modified SL3-3 viruses; (B) 59 UTR-modified virus.
FIG. 2. Southern blot of tumor DNAs digested withHindIII and probed for rearrangements in their TCRbloci (J2 probe). The arrow indicates the position of the nonrearranged germ line band.
TABLE 1. Lymphoma induction in inbred NMRI mice by wt SL3-3 and mutant-PBS viruses
Virus
No. of mice with malignant lymphomas/no. of mice analyzed
Mean latency6
SD (days)
No. of tumors with genomic rearrangements within the TCRb-chain/no. of tumors
analyzed
No. of tumors with genomic rearrangements within the
Igklight chain/no. of tumors analyzed
No. of tumors with genomic rearrangements within the c-mycpromoter region/no.
of tumors analyzed
SL3-3-Pro 14/14 5468 10/11 0/12 4/14
SL3-3-Gln1 15/15 6169 10/11 0/8 2/12
SL3-3-Lys3 14/14 76620 12/12 0/11 3/13
SL3-3-Arg1,2 14/14 68616 9/9 0/10 4/13
SL3-3-Proa 20/20 5667 NDb ND ND
SL3-3-Pro/Akv UTRa 20/20 6065 ND ND ND
aSeparate experimental series. bND, not determined.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.54.551.602.712.2]T and C readout at position 8 for all of the analyzed Gln1 PBS sequences from tumor proviruses indicates that the SL3-3-Gln mutant may replicate with both tRNA1Glnand tRNA2Gln(Fig. 3), as was previously seen for Akv-MLV-derived vectors (24). wt SL3-3-Pro was ubiquitously found in tumors from mice originally injected with SL3-3-Pro, but in one tumor a PBS sequence matching tRNAGlnwas detected by subcloning the PCR product. The sequences flanking the Gln PBS contained a pattern of nucleotide substitutions, deletions, and insertions relative to the sequence of SL3-3-Pro virus originally injected and were highly homologous to previously identified proviral sequences endogenous to the murine genome (10, 29). A sim-ilar picture was seen with the lymphomas induced by the SL3-3-Arg1,2 mutant. SL3-SL3-3-Arg1,2 sequences were found again in all tumors, and one tumor contained additional proviruses harboring a glutamine PBS sequence (Table 2). Among the tumors taken from mice originally injected with SL3-3-Lys3, none contained only proviruses harboring the original PBS matching tRNA3Lys. In two of the tumors originally arising from infection with SL3-3-Lys3, the PBS had been altered to match the 39 18 nucleotides of tRNA1,2Lys. The Lys3 PBS and Lys1,2 PBS differ at five positions only (Fig. 3). Since the sequences flanking the Lys1,2 PBS were SL3-3 specific, these altered
viruses are likely to have originated from the binding of a tRNA1,2Lysdirectly to the Lys3 PBS sequence rather than from recombination with an endogenous virus harboring the Lys1,2 PBS. Interestingly, the tumor holding only a Lys1,2 PBS-con-taining SL3-3 mutant originated from the one SL3-3-Lys3-injected mouse in which disease induction was delayed to twice the average latency period. tRNA1,2Lys molecules are used as replication primers by visna (42) and spuma (28) viruses, and a PBS sequence matching tRNA1,2Lyshas previously been identi-fied in human endogenous retrovirus K (32). However, to our knowledge this represents the first report of an MLV replicat-ing via tRNA1,2Lys. Since direct sequencing of PCR products amplified from tumors originating from mice injected with SL3-3-Lys3 resulted in mixed sequences in 12 of 13 tumors analyzed, PCR products from these tumors were cloned and individual subclones were sequenced. In 11 of 12 tumors, anal-ysis of individual subclones revealed a mixture of proviral se-quences carrying either the Lys3 or Gln PBS (Table 2). Whereas the sequences flanking the Lys3 PBS were identical to those in the original SL3-3-Lys3 mutant, all the sequences harboring a Gln PBS also contained the endogenous proviral sequence-characteristic pattern of substitutions, insertions, and deletions. In 1 of 12 of the tumors analyzed by sequencing of subclones, three PBS sequences, the Lys3, Gln, and Lys1,2 PBSs, were detected (Table 2). The exact nature of the recom-binants is currently being investigated (26). To determine if second-site mutations were selectively induced as a function of replication with alternative tRNA primers, the sequences flanking the PBS (;250 bp in total) were inspected. Whereas single nucleotide substitutions were found in a number of proviruses, as were one 2-bp duplication and one 2-bp dele-tion, no evidence for a recurring tRNA-specific pattern of second-site mutations was found within the sequences flanking the PBS. Hence, we found no indications of molecular adap-tation towards replication with alternative tRNA primers within the region analyzed. However, we cannot from these analyses dismiss the possibility of second-site mutations within the coding regions resulting from replication with tRNAs other than tRNAPro.
Among the tumors originating from infection with
SL3-3-FIG. 3. (A) wt SL3-3 and mutant PBS-tRNA base pairings. Note that tRNA2Glnbinding to the Gln1 PBS does not disrupt base pairing. (B) Potential base pairings
between the Lys3 PBS and various tRNA species. Note that 14 base pairings may be formed between the Lys3 PBS and tRNA1,2Lys. Numbers to the right of the panels
[image:4.612.53.297.83.201.2]indicate numbers of bases paired.
TABLE 2. Analysis of proviral PBS sequences in tumor DNAs
Virus Frequency of tumors withconserved PBS sequences only (PBS type)
Frequency of tumors with altered PBS sequences
(PBS type[s])
SL3-3-Pro 13/14 (Pro) 1/14 (Pro and Gln1,2)a
SL3-3-Gln1 12/12 (Gln1,2) NDb
SL3-3-Lys3 ND 1/13 (Lys1,2)
11/13 (Lys3 and Gln1,2)a 1/13 (Lys3 and Gln1,2 or Lys1,2)a
SL3-3-Arg1,2 12/13 (Arg1,2) 1/13 (Arg1,2 and Gln1,2)a aSequences generated from cloned PCR products.
bND, not detected.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.66.541.503.700.2]Lys3, three were found to contain clonal rearrangements in the c-mycgene. Since recombinant proviruses containing Gln PBS sequences were detected in all three tumors, we wanted to analyze the PBS present in a c-myc-integrated provirus. With PCR primers specific for MLVgagand c-myc(43) an;1.5-kb PCR fragment was generated and directly sequenced. The sequencing verified the presence of an SL3-3-Lys3 provirus integrated in the opposite transcriptional orientation relative to c-myc 1,415 bp upstream from the first c-myc exon (se-quence information from GenBank accession no. M12345). SL3-3-Lys3 may therefore be involved in the tumorigenesis process in a manner similar to that of wt SL3-3, despite the observed delay in lymphoma induction (Fig. 1).
In conclusion, we find that the SL3-3 MLV may stably and efficiently replicate with tRNA primers other than tRNAPro, although in vivo replication with alternative tRNA primers exhibits lowered replication kinetics. The reason for this find-ing remains unclear. While the results may reflect that MLV tRNA primer specificities act on the level of packaging, an-nealing, or RT-tRNA recognition, they may also reflect non-viral phenomena such as tRNA availability. In addition, alter-ation of the PBS sequence to match tRNA3Lys renders the mutant highly prone to recombination with endogenous MLV-like sequences. The results obtained in this study are in con-trast to those of previous reports on PBS-modified retrovi-ruses. While in this study none of the PBS mutants were found to revert to tRNAProutilization, PBS-modified HIV-1 (11, 21, 45) and avian leukosis virus (47), analyzed by extended growth in cell culture, were found to eventually revert to the PBS sequence of the wt virus. Results from this study therefore sustain the notion of a less stringent usage of the MLV tRNA primer relative to those of other retroviruses and point to base pairings between the tRNA and the PBS sequence as the major determinant of replication primer selection. The PBS-modified viruses were found to induce malignant lymphomas indistin-guishable from tumors induced by wt SL3-3 in terms of gross appearance, frequencies of genomic rearrangements within the TCRb-chain and the c-myclocus, and an absence of genomic rearrangements within the Igklight chain. Hence, we find no evidence that disruption of the negatively actingciselement originally identified in embryonal cells and overlapping the Pro PBS sequence significantly affected the viral tumorigenesis process in this model system. However, the complex pattern of molecular interactions underlying the development of neo-plasms by MLV is still poorly understood, and it remains a formal possibility that negative effects of the Pro PBS sequence may reduce the appearance of revertants to Pro PBS. While several disease stages typical of MLV leukemogenesis, such as early viral replication within the bone marrow, rapid polyclonal expansion of preleukemic cells in the spleen, and proviral in-sertional mutagenesis and cisactivation of cellular proto-on-cogenes, may potentially be affected by the introduction of PBS mutations, the final effect of such PBS alterations cannot be predicted. We acknowledge, however, that the combination of highly pathogenic SL3-3 MLV isolates and a sensitive inbred mouse strain may have precluded the emergence of an altered pathogenicity pattern resulting from increased viral replication and transcriptional access in non-T cells.
The technical assistance of Jane Jensen, Angelika Appold, Anna Nickl, and Elenore Samson is gratefully acknowledged.
This work was supported by contracts CT 95-0100 (Biotechnology) and CT 95-0675 (Biomed 2) of the European Commission, the Karen Elise Jensen Foundation, the Danish Cancer Society, the Danish Bio-technology Program, and the Danish Natural Sciences and Medical Research Councils.
REFERENCES
1.Aiyar, A., D. Cobrinik, Z. Ge, H.-J. Kung, and J. Leis.1992. Interaction
between retroviral U5 RNA and the TcC loop of the tRNATrpprimer is
required for efficient initiation of reverse transcription. J. Virol.66:2464– 2472.
2.Amtoft, H. W., A. B. Sørensen, C. Bareil, J. Schmidt, A. Luz, and F. S.
Pedersen.1997. Stability of AML1 (core) site enhancer mutations in T
lymphomas induced by attenuated SL3-3 murine leukemia virus mutants. J. Virol.71:5080–5087.
3.Araya, A., L. Sarih, and S. Litvak.1979. Reverse transcriptase mediated
binding of primer tRNA to the viral genome. Nucleic Acids Res.6:3831– 3843.
4.Barat, C., S. F. Le Grice, and J. L. Darlix.1991. Interaction of HIV-1 reverse transcriptase with a synthetic form of its replication primer, tRNALys,3.
Nucleic Acids Res.19:751–757.
5.Barat, C., V. Lullien, O. Schatz, G. Keith, M. T. Nugeyre, F. Gruninger
Leitch, F. Barre-Sinoussi, S. F. LeGrice, and J. L. Darlix.1989. HIV-1
reverse transcriptase specifically interacts with the anticodon domain of its cognate primer tRNA. EMBO J.8:3279–3285.
6.Barklis, E., R. C. Mulligan, and R. Jaenisch.1986. Chromosomal position or virus mutation permits retrovirus expression in embryonal carcinoma cells. Cell47:391–399.
7.Baum, C., S. Hegewisch-Becker, H. G. Eckert, C. Stocking, and W. Ostertag.
1995. Novel retroviral vectors for efficient expression of the multidrug resis-tance (mdr-1) gene in early hematopoietic cells. J. Virol.69:7541–7547. 8.Bordier, B., L. Tarrago-Litvak, M. L. Sallafranque-Andreola, D. Robert, D.
Tharaud, M. Fournier, P. J. Barr, S. Litvak, and L. Sarih-Cottin.1990.
Inhibition of the p66/p51 form of human immunodeficiency virus reverse transcriptase by tRNALys. Nucleic Acids Res.18:429–436.
9.Cobrinik, D., A. Aiyar, Z. Ge, M. Katzman, H. Huang, and J. Leis.1991.
Overlapping retrovirus U5 sequence elements are required for efficient in-tegration and initiation of reverse transcription. J. Virol.65:3864–3872. 10. Colicelli, J., and S. P. Goff.1987. Isolation of a recombinant murine
leuke-mia virus utilizing a new primer tRNA. J. Virol.57:37–45.
11. Das, A. T., B. Klaver, and B. Berkhout.1995. Reduced replication of human immunodeficiency virus type 1 mutants that use reverse transcription primers other than the natural tRNA3Lys. J. Virol.69:3090–3097.
12. Ethelberg, S., J. Lovmand, J. Schmidt, A. Luz, and F. S. Pedersen.1997.
Increased lymphomagenicity and restored disease specificity of AML1 site (core) mutant SL3-3 murine leukemia virus by a second-site enhancer vari-ant evolved in vivo. J. Virol.71:7273–7280.
13. Fu, W., B. A. Ortiz-Conde, R. J. Gorelick, S. H. Hughes, and A. Rein.1997. Placement of tRNA primer on the primer-binding site requirespolgene expression in avian but not murine retroviruses. J. Virol.71:6940–6946. 14. Gilboa, E., S. W. Mitra, S. Goff, and D. Baltimore.1979. A detailed model
of reverse transcription and tests of crucial aspects. Cell18:93–100. 15. Haseltine, W. A., A. Panet, D. Smoler, D. Baltimore, G. Peters, F. Harada,
and J. E. Dahlberg.1977. Interaction of tryptophan tRNA and avian
my-eloblastosis virus reverse transcriptase: further characterization of the bind-ing reaction. Biochemistry16:3625–3632.
16. Hays, E. F., G. Bristol, and S. McDougall.1990. Mechanisms of thymic
lymphomagenesis by the retrovirus SL3-3. Cancer Res.50:5631S–5635S. 17. Ja¨hner, D., and R. Jaenisch.1985. Retrovirus-induced de novo methylation
of flanking host sequences correlates with gene inactivity. Nature315:594– 597.
18. Lenz, J., D. Celander, R. L. Crowther, R. Patarca, D. W. Perkins, and W. A.
Haseltine.1984. Determination of the leukaemogenicity of a murine
retro-virus by sequences within the long terminal repeat. Nature308:467–470.
19. Lenz, J., R. Crowther, S. Klimenko, and W. Haseltine.1982. Molecular
cloning of a highly leukemogenic, ecotropic retrovirus from an AKR mouse. J. Virol.43:943–951.
20. Lenz, J., and W. A. Haseltine.1983. Localization of the leukemogenic
de-terminants of SL3-3, an ecotropic, XC-positive murine leukemia virus of AKR mouse origin. J. Virol.47:317–328.
21. Li, X., J. Mak, E. J. Arts, Z. Gu, L. Kleiman, M. A. Wainberg, and M. A.
Parniak.1994. Effects of alterations of primer-binding site sequences on
human immunodeficiency virus type 1 replication. J. Virol.68:6198–6206. 22. Lovmand, J., A. H. Lund, and F. S. Pedersen.1997. Growth and purification
of murine leukemia virus, p. 528–533.InJ. E. Celis (ed.), Cell biology. A laboratory handbook. Academic Press, London, United Kingdom.
23. Lovmand, J., A. B. Sørensen, J. Schmidt, M. Østergaard, A. Luz, and F. S.
Pedersen.1998. B-cell lymphoma induction by Akv murine leukemia viruses
harboring one or both copies of the tandem repeat in the U3 enhancer. J. Virol.72:5745–5756.
24. Lund, A. H., M. Duch, J. Lovmand, P. Jørgensen, and F. S. Pedersen.1993.
Mutated primer binding sites interacting with different tRNAs allow efficient murine leukemia virus replication. J. Virol.67:7125–7130.
25. Lund, A. H., M. Duch, J. Lovmand, P. Jørgensen, and F. S. Pedersen.1997.
Complementation of a primer binding site-impaired murine leukemia virus-derived retroviral vector by a genetically engineered tRNA-like primer. J. Virol.71:1191–1195.
on November 9, 2019 by guest
http://jvi.asm.org/
26.Lund, A. H., J. G. Mikkelsen, M. Duch, J. Schmidt, A. Luz, and F. S.
Pedersen.Unpublished data.
27. Majors, J. E., and H. E. Varmus.1983. Nucleotide sequencing of an
appar-ent proviral copy ofenvmRNA defines determinants of expression of the mouse mammary tumor virusenvgene. J. Virol.47:495–504.
28. Maurer, B., H. Bannert, G. Darai, and R. M. Flu¨gel.1988. Analysis of the primary structure of the long terminal repeat and thegagandpolgenes of the human spumaretrovirus. J. Virol.62:1590–1597.
29.Mikkelsen, J. G., A. H. Lund, K. D. Kristensen, M. Duch, M. S. Sørensen, P. Jørgensen, and F. S. Pedersen.1996. A preferred region for recombinational patch repair in the 59untranslated region of primer binding site-impaired murine leukemia virus vectors. J. Virol.70:1439–1447.
30.Mishima, Y., and J. A. Steitz.1995. Site-specific crosslinking of 4-thiouri-dine-modified human tRNA3Lysto reverse transcriptase from human
immu-nodeficiency virus type I. EMBO J.14:2679–2687.
31. Nikbakht, K. N., C. Y. Ou, L. R. Boone, P. L. Glover, and W. K. Yang.1985. Nucleotide sequence analysis of endogenous murine leukemia virus-related proviral clones reveals primer-binding sites for glutamine tRNA. J. Virol.
54:889–893.
32. Ono, M.1986. Molecular cloning and long terminal repeat sequences of
human endogenous retrovirus genes related to types A and B retrovirus genes. J. Virol.58:937–944.
33. Panet, A., and H. Berliner.1978. Binding of tRNA to reverse transcriptase of RNA tumor viruses. J. Virol.26:214–220.
34. Pattengale, P. K.1994. Tumours of the lymphohaematopoietic system, p.
651–670.InV. S. Turusov and U. Mohr (ed.), Pathology of tumours in laboratory animals, vol. II. Tumours of the mouse. IARC Scientific Publica-tion no. 111. InternaPublica-tional Agency for Research on Cancer, Lyon, France.
35. Pedersen, F. S., R. L. Crowther, D. Y. Tenney, A. M. Reimold, and W. A.
Haseltine.1981. Novel leukaemogenic retroviruses isolated from cell line
derived from spontaneous AKR tumour. Nature292:167–170.
36. Peters, G., F. Harada, J. E. Dahlberg, A. Panet, W. A. Haseltine, and D.
Baltimore.1977. Low-molecular-weight RNAs of Moloney murine leukemia
virus: identification of the primer for RNA-directed DNA synthesis. J. Virol.
21:1031–1041.
37. Sarih-Cottin, L., B. Bordier, K. Musier-Forsyth, M. L. Andreola, P. J. Barr,
and S. Litvak.1992. Preferential interaction of human immunodeficiency
virus reverse transcriptase with two regions of primer tRNALysas evidenced
by footprinting studies and inhibition with synthetic oligoribonucleotides. J. Mol. Biol.226:1–6.
38.Sawyer, R. C., and J. E. Dahlberg.1973. Small RNAs of Rous sarcoma virus: characterization by two-dimensional polyacrylamide gel electrophoresis and fingerprint analysis. J. Virol.12:1226–1237.
39.Schmidt, J., V. Erfle, F. S. Pedersen, H. Rohmer, H. Schetters, K. H.
Mar-quart, and A. Luz.1984. Oncogenic retrovirus from spontaneous murine
osteomas. I. Isolation and biological characterization. J. Gen. Virol.65:2237– 2248.
40.Schmidt, J., A. Luz, and V. Erfle.1988. Endogenous murine leukemia
vi-ruses: frequency of radiation-activation and novel pathogenic effects of viral isolates. Leuk. Res.12:393–403.
41.Seiki, M., S. Hattori, Y. Hirayama, and M. Yoshida.1983. Human adult
T-cell leukemia virus: complete nucleotide sequence of the provirus genome integrated in leukemia cell DNA. Proc. Natl. Acad. Sci. USA80:3618–3622.
42.Sonigo, P., M. Alizon, K. Staskus, D. Klatzmann, S. Cole, O. Danos, E.
Retzel, P. Tiollais, A. Haase, and S. Wain-Hobson.1985. Nucleotide
se-quence of the visna lentivirus: relationship to the AIDS virus. Cell42:369– 382.
43.Sørensen, A. B., M. Duch, H. W. Amtoft, P. Jørgensen, and F. S. Pedersen.
1996. Sequence tags of provirus integration sites in DNAs of tumors induced by the murine retrovirus SL3-3. J. Virol.70:4063–4070.
44.Wain-Hobson, S., P. Sonigo, O. Danos, S. Cole, and M. Alizon.1985.
Nu-cleotide sequence of the AIDS virus, LAV. Cell40:9–17.
45.Wakefield, J. K., A. G. Wolf, and C. D. Morrow.1995. Human
immunode-ficiency virus type 1 can use different tRNAs as primers for reverse tran-scription but selectively maintains a primer binding site complementary to tRNA3Lys. J. Virol.69:6021–6029.
46. Weiher, H., E. Barklis, W. Ostertag, and R. Jaenisch.1987. Two distinct
sequence elements mediate retroviral gene expression in embryonal carci-noma cells. J. Virol.61:2742–2746.
47. Whitcomb, J. M., B. A. Ortiz-Conde, and S. H. Hughes.1995. Replication of avian leukosis viruses with mutations at the primer binding site: use of alternative tRNAs as primers. J. Virol.69:6228–6238.