0022-538X/81/030860-11$02.00/0
Comparative
Analysis: Intracellular
Precursor
Polyproteins
of
Baboon
Endogenous
Retroviruses and Human Viral Isolate
HL23V
S. A. WHITELEYANDR. B. NASO*
Departmentof Tumor Virology,The University ofTexasSystemCancerCenter M. D. AndersonHospital
andTumorInstitute,Houston, Texas 77030
Intracellular precursor polyproteins of three baboon endogenous retrovirus (BaEV) isolates, m7, 455K, and BILN, were compared with the intracellular proteins of the type C human isolate HL23V by radioimmunoprecipitation, followedby sodiumdodecylsulfate-polyacrylamidegelelectrophoresisandtryptic peptide analysis. Human and canine cells infected with m7-BaEV and canine thymus cells infected withBILN-BaEVwerecharacterizedbyidenticalprecursor polyproteinsPr85V",
Pr70-719"9,
Pr659"9, and gPr85env. Caninecellsinfectedwith 455K-BaEVconsistentlyshowedaslightly different patternof precursor polypro-teins. These included Pr85V", Pr7O9'9, Pr67gag, andgPr85env. Bytryptic digest mapping of peptides containing [3H]leucine, m7-BaEV and 455K-BaEV wereshowntobehighlyrelated.Bycomparison, mappingstudies showed that BILN-BaEVwaslesshighlyrelatedtom7-BaEV thanwas455K-BaEV.Differences in these related BaEV isolatespresumablyreflectedvirus-specific differential cleav-ageofcoreproteinprecursorsoralterations inpolyproteinprimarystructure or
both. Chase-incubated cells infected with BaEV also contained a stable, p28-relatedpolyprotein termed P72r". This polyprotein migrateduponsodium do-decyl sulfate-polyacrylamide gel electrophoresis slightly slowerthan the major
core protein precursor Pr70-719'9 and appeared to arise by posttranslational modification of Pr7O-719'9. Immunoprecipitation ofextracts ofHL23V-infected cellswith antiseratosimiansarcoma-simian-associated virusproteinsand BaEV proteins confirmedthat thesecells containedtwounrelated viralcomponents,one
thatwassimilar tom7-BaEVor BILN-BaEV anda secondthatwasrelatedto
simian sarcoma-simian-associated virus. Tryptic digest mapping of BaEV and HL23V precursor polyproteins suggested that the BaEV-like component of HL23V was more closely related to m7-BaEV than to 455K-BaEV or BILN-BaEV.
Baboon endogenous type C retroviruses (BaEV) have been isolated fromseveralspecies of the baboon (Papio) genus, including Papio cynocephalus (m7-BaEV),Papioanubis (455K-BaEV), Papio hamadryas (BILN-BaEV), and Papiopapio (PP-1-BaEV) (4, 13, 28, 29). The
m7 and 455K isolates are indistinguishable by nucleic acidhybridization, but partial nucleotide sequence data suggest thatthey maydiffer by
at least one nucleotide at the 5' end of the genome (14, 28). The other isolates have been shown to be more readily distinguishable by nucleic acid hybridization (28) or by type-spe-cific immunological differences (23, 25, 26). These studies have also demonstrated that, al-though the various BaEV isolates are closely related, theyare distinct from the retroviruses isolated from otherprimates, includingthe sim-ian sarcoma virus-gibbon ape leukemia virus
group(SiSV-GaLV) (12-14). A type C retrovirus (HL23V) isolated fromculturedleukocytesof a patient with acute myelogenous leukemia has been showntocontain components immunolog-ically relatedtobothSiSV-GaLV and BaEV (9, 11, 18, 19). We report that, analogous to the well-characterizedmurine and avian type C ret-rovirus systems(1-3, 7), BaEV viral proteins are made by synthesis and cleavage of precursor polyproteins. In this paper we characterize the virus-specific proteins and intracellular
precur-sor polyproteins of three BaEV isolates by ra-dioimmunoprecipitation assay, followed by so-diumdodecylsulfate-polyacrylamidegel electro-phoresis(SDS-PAGE)and ion-exchange column chromatography of trypsin-digested proteins. Thecharacteristicsofthese primate retroviruses
are compared with the BaEV-like viral gene products expressed in HL23V-infected cells.
860
on November 10, 2019 by guest
http://jvi.asm.org/
BaEV PRECURSOR POLYPROTEINS 861
MATERLALS AND METHODS
Cell cultures. Thevariousprimatevirus isolates
wereobtainedfrom cultureskindly supplied byRobert C.Galo,National Institutes ofHealth,Bethesda, Md.,
and HEM Research Inc., Rockville, Md.
CT/455K-BaEVwasoriginallyobtainedbycocultivation of
ca-ninethymuscells withkidneytissue of P.anubis (29). A204/m7-BaEVandCT/m7-BaEVwereoriginally
ob-tainedbycocultivation of either human
rhabdomyo-sarcomacellsorcaninethymuscells, respectively,with
placentaltissue from P.cynocephalus(4). CT/BILN-BaEVwasoriginally obtainedbyinfectionofcanine thymuscellswithavirusisolated fromthetissuesof
alymphomatousP.hamadryas (13).Simian
sarcoma-simian-associatedvirus [SiSV(SiAV)], originally
iso-lated from a fibrosarcoma ofawoollymonkey, was
propagated in marmoset cells, 71AP1 (27, 31). The human virusisolate HL23V (9) wasgrown in either A204cells or7573 caninethymuscelLs.TheSiSV-like
component of HL23V (9) was grown in normal rat
kidney (NRK) cells,whicharerefractiletoinfection
bythe BaEV-like component ofHL23V. This
SiSV-like virus, termed 23SiSV(SiAV), was then used to infectcanine fibroblast CF2thcells.
Allcellsweregrowntoconfluenceinminimal Eagle
medium containing 10% fetal calfserum and
genta-micinat50,ug/mlinT-75 flasks. Eachinfectedculture
wasshownto be capable ofproducing aninfectious
virus,asdeterminedby the abilityofafiltered growth
mediumtoproductively infectcaninethymuscellsin
culture.
Isolation ofradioactively labeled intracellular
viralproteins.Labelingofcytoplasmic proteinswas
performedbyincubation ofculturesfor 20minat37°C
in10 ml ofEarlebalanced salt solutionsupplemented
with 2.5 mCi of[3H]leucine (45 Ci/mmol; New
Eng-land Nuclear Corp., Boston, Mass.). Cultures were
eitherlysedimmediatelyorchaseincubated for60to
90 min in a complete growth medium without the
isotope before lysis. Forsome experiments, cultures werepulsedfor 6 h with[3H]glucosamine(38Ci/mol;
Amersham Corp., Arlington Heights, Ill.) in a
com-pletegrowth medium and then lysed.Thelysis
pro-cedure andimmunoprecipitation ofcytoplasmicviral
proteinswereperformedasdescribedby Arcementet
al. (1). MonospecificantiseratoBaEVp28,Rauscher
murine leukemia virus (RLV) p30,and RLVgp70or
polyspecificantisera toBaEV, SiSV,andGaLVwere
obtained through the Office ofProgram Resources
and Logistics, ViralOncology Program, National
In-stitutes of Health.Immunoprecipitationwasenhanced
bythe addition of theFormalin-fixed Cowan I strain
ofStaphylococcusaureus(21).Theprecipitateswere
collected by centrifugation, washedthree times, and
dissolvedinsamplebufferbeforeelectrophoresis.
Im-munoprecipitated cytoplasmicviralproteinswere
an-alyzed bySDS-PAGEon6 to12%gradientgels, using
the buffer system of Laemmli (16). The gels were
subjectedtofluorography (5),and theX-rayfilmwas preflashedtoobtaina linearresponse(17).
Peptide mapping. Tryptic digestionof viral
pro-teins andseparationofthe resultantpolypeptide
frag-mentsbyion-exchange chromatography on Chromo-beadtype Presin(TechniconCorp., Inc., Tarrytown,
N.Y.)wereperformed as previously described (1). The two-dimensional analysis of peptides has also been described previously(15). Briefly, dry slab gel bands were digested with trypsin, and the soluble peptides wereseparated on thin-layercellulose plates (20 by 20 cm) by electrophoresis in 28% formic acid in the first dimension and by ascending chromatography in the second dimension, using a butanol-pyridine-acetic acid-water(6.2:1:3.3:2.8) buffer system. The spots rep-resenting radioactive peptides were detected by fluo-rography (15).
RESULTS
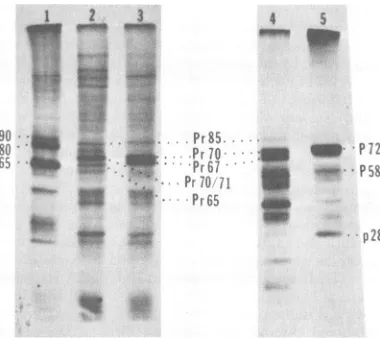
Immunoprecipitation of BaEV-specific intracellular polyproteins from infected cells. Various cell lines infected with BaEV isolates were compared for the presence of BaEV-related precursor-like polyproteins. An-tiserum to m7-BaEV p28 recognized several high-molecular-weight proteins from the m7-BaEV-infectedcells (Fig. 1,lanes1and 5). These included proteins of approximnately 85,000
(Pr5ag),71,000 (Pr7lgag), 70,000 (Pr70V¶), and
65,000
(Pr659'¶)
Mr and some smallerproteinsin the 40,000- to 55,000-Mrrange. Inpulse-labeled cells infected withm7-BaEV, Pr70Vg andPr719'9 always appearedas aclosely migratingdoublet in gels. Identical profiles ofprecursor-like pro-teinswere obtained from either canine (Fig. 1, lane 1) or human (Fig. 1, lane 5) cells infected with m7-BaEVorfrom cells infected with BILN-BaEV (data not shown). Chase incubation of pulse-labeled canine or human cells infected with m7-BaEV (Fig. 1, lanes 2 and 6,respec-tively) allowed the removal of labeled p28-re-lated precursorpolyproteins,theappearance of radioactive p28, and an intensification in the levels of an intermediate-sized protein of
ap-proximately 58,000 Mr (P58W). In addition, a newradiolabeled polyprotein of approximately 72,000 Mr (P72¶) appeared inthe cells.
Analy-sesofHL23V-infected,pulse-labeled canine and human cells with anti-BaEV p28 serum (Fig. 1, lanes 3 and 7,respectively) revealed polyprotein profiles that were nearly identical to those of m7-BaEV-infected cells (Fig. 1, lanes 1 and 5). Pulse-labeledHL23V-infected cellsconsistently contained lowerlevels of the 40,000- to 50,000-Mr proteins than did cells infected with m7-BaEV or BILN-BaEV. Aspreviously noted for the other BaEV isolates, chase incubation of HL23V-infected cellsresulted in thecleavageof thelarge-Mr precursor-like proteinsand the ap-pearance ofp28and twolargerproteins, p58gag and P72-G` (Fig. 1, lanes 4 and 8). The P729KG band from BaEV- or HL23V-infected cells al-ways migrated slightly slower upon SDS-PAGE than did the 71,000Mr band in the Pr70-71gag doublet (Fig. 1, lanes7and8). TheP72" band VOL. 37, 1981
on November 10, 2019 by guest
http://jvi.asm.org/
FIG. 1. Synthesis andcleavageofBaEV and HL23Vprecursorpolyproteins. Cultures werepulse-labeled
for15min with[3HJleucine (lanes 1, 3, 5, 7,and9) and chase incubatedfor1 h (lanes2, 4, 6, 8, and10).
Cultures labeled includedCT/m7-BaEV(lanesI and2),A204/HL23V(lanes3 and4),A204/m7-BaEV (lanes 5and 6), and 7573/HL23V(lanes7to10). Equal portionsofcytoplasmicextracts wereimmunoprecipitated
withanti-BaEVp28(lanes1to8),followedbyreimmunoprecipitationwith anti-BaEV(lanes9 and10).Lane 11 representsamarkerof RLVproteinsandprecursorpolyproteins.
appearedtobelabeledonlyinchase-incubated cells(Fig.1,lanes 2,4, 6, and8).Noneof thecell linesinfected with the BaEV isolatesorHL23V exhibited agag-pol-likeprecursorof180,000to
200,000Mrprecipitable withanti-BaEVp28 se-rum (Fig. 1,lanes 1,3, 5, and7). If the extracts
were reimmunoprecipitated with a polyspecific
antiserum to detergent-disrupted m7-BaEV aftertheywerecleared withanti-BaEVp28
se-rum, the BaEV envelope protein precursors could be visualized free fromcontamination by
coreprotein-related precursors. When pulse-la-beled HL23V-infected cells were thus treated, the precipitates contained only one protein of approximately 85,000(gPr85env)Mr (Fig. 1, lane 9). The same-sizedprotein couldbeobservedin any of the cell lines infected with the BaEV isolates (data notshown). Chase incubations of upto90minresulted inonlyaslightdecrease in size of the envelope protein precursor (Fig. 1,
lane 10). The 18,000-M, protein (p18)
precipi-tatedfrom thesepulse-labeled,chase-incubated cells byanti-BaEV serum (Fig. 1, lane 10) is a nonglycosylated BaEV-like envelope protein that shares peptides with gPr85env (data not shown).
Canine cells infected with 455K-BaEV con-tainedaslightlydifferent patternof p28-related proteins than didcellsinfected with m7-BaEV, BILN-BaEV, orHL23V (Fig. 2, lanes 2 and 3). Pulse-labeled cells infected with 455K-BaEV
- '-P 7.-'0' i :.' 4 ' _ P7
128
FIG. 2. Intracellular precursor proteins of CT!
455K-BaE versus7573/HL23V. Cultures
werepulse-labeledfor15mnwith[3H].eucine(lanes1to4)and
chase incubatedfor 1 h (lane 5). Extracts of7573/
HL23V(lane 2) and CT/455K-BaEV(lanes3 to 5)
cellswere immunoprecipitatedwith anti-BaEVp28.
Lane 1representsamarkerofRLV-specific proteins. contained severalprecursor-likeproteins
precip-itable withanti-p28serum(Fig.2,lane3).These included proteins of (PrPr1),85,000 70,000
(Pr7OgaR1),-and 67,000 (Pr679a") Mr and variable
amounts of
lower-M,
proteins. Upon chasein-cubation, these radioactiveproteinsdiminished
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.488.253.443.354.523.2]VOL. 37, 1981
in amounts, whereas amounts of labeled p28 increased (Fig. 2, lane 5). Chase-incubated cells infected with 455K-BaEV were also character-izedbythe presence ofP72gwandP58gw(Fig.2, lane5).
BaEV- and SiSV-like components in HL23V-infected celis.
Electrophoretic
analy-sesofimmunoprecipitatesofextractsfrom cells
infected withHL23Vconfirmedthatthese cells contained bothaBaEV-likecomponentandan SiSV-like component (Fig. 3). The precursor polyproteins of the two components could be separatedreadilyforanaysisbydifferential
im-munoprecipitation with theappropriate
antise-rum. Aspreviouslynoted (Fig. 1,lane3),
treat-ment of extracts of pulse-labeled HL23V-in-fected caninefibroblast cellswithantiserumto
BaEV p28 precipitated the typical BaEV
p28-related polyproteins Pr851'1,
Pr70-719"9,
and Pr65V9(Fig.3,lane1). Likewise,reimmunopre-cipitation of extracts with a polyspecific anti-BaEV serum, after clearing them with anti-BaEVp28 serum,revealed the BaEV-like enve-lopeprotein precursor
gPr85e"v
(Fig. 3, lane2).Obviously,allBaEVprecursorsrelatedtoBaEV
p28 wereremovedbythe firstprecipitationwith anti-BaEV p28.
Reimmunoprecipitation
of the anti-BaEV p28-cleared cellextract withan an-tiserum to SiSV p28, however, revealed a newPr8O
~Pr6O*
BaEV PRECURSOR POLYPROTEINS 863
set of precursor polyproteins that included a
minor protein, Pr8O'9, and a major
protein,
Pr6fg (Fig. 3, lane 3). Only trace amounts of these SiSV-like core protein precursors wereprecipitated by anti-BaEV p28 serum (Fig. 3,
lane 1).Immunoprecipitationofextracts of
sim-ilarly pulse-labeledcells withantiserumtoSiSV
p28identifiedthe same twoSiSV-like core pro-teinprecursorpolyproteins,Pr809`9andPr6OY",
but not the BaEV-like precursors (Fig. 3, lane
4).Reimmunoprecipitation of thesupernatants
ofthe cellextractswithapolyspecific antiserum
toSiSV,afterpriorclearing with anti-SiSV p28,
revealed the presence of a SiSV-like envelope
proteinprecursor,gPr8Oenv (Fig. 3, lane 5). The
SiSV-like envelope protein precursor was not
precipitable byanantiserum to BaEV proteins
(Fig. 3, lane 2).TheSiSV-like envelope protein precursor gPr8Oenv migrated significantly faster uponSDS-PAGEthandidthe analogous BaEV-likeprecursorgPr85e (Fig. 3,compare lanes 2 and4). The SiSV-like componentofHL23V is infectious forNRK cellstothe exclusion of the BaEV-like component. This SiSV-like virus, termed 23SiSV(SiAV), derived from infected NRK cellswas usedto infect canine fibroblast cells.Thesecellswerepulse-labeledasdescribed above, and cell extracts were treated with an antiserum to SiSV p28 (Fig. 3, lane6).
Super-Pr200
-gPr8O.
Pr8O0
Pr6O
* *FIG. 3. Intracellular precursorproteins ofHL23V. Cultures of canine thymus cells infected with HL23V
(lanes 1 to5), orinfectedwith onlytheSiSV-likecomponentof HL23V (lanes6and7), ormarmoset cells
infectedwithSiSV(SiAV) (lanes8and9) werepulse-labeled for 15 min with
[3H]leucine.
Equalportions of thecytoplasmicextractwereimmunoprecipitatedwithanti-BaEVp28 (lane 1), followed by reimmunoprecip-itation with eitheranti-BaEV(lane2)oranti-SiSV(lanes3),orwithanti-SiSVp28 (lanes 4,6, and 8), followedbyreimmunoprecipitationwith anti-SiSV(lanes5, 7,and9).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.488.44.443.372.598.2]864 WHITELEY AND NASO
natantsfrom this cleared cellextractwere then treated with an antiserum to detergent-dis-ruptedSiSV (Fig.3, lane7). Immunoprecipita-tion resulted in the usual SiSV-like precursors
Pr2OOgagpol,
Pr8O"',
andPr6Og (Fig.3, lane 6)andgPr8Oenv (Fig. 3, lane 7). Treatment of
ex-tracts from these 23SiSV(SiAV)-infected cells withantiserum toBaEVproteins didnotreveal anyBaEV-likeprecursorpolyproteins (datanot shown).Figure3,lanes8and9, showsthe results ofsimilarimmunoprecipitations ofextracts de-rivedfrompulse-labeled marmosetcells(71AP1) infected withSiSV(SiAV).
Labeling of HL23V precursor proteins with
['H]glucosamine.
Figure 4 shows the SDS-PAGE profile ofHL23V-infected cells that were long term labeled with [3H]glucosamine. Nocore-relatedproteins werelabeled with glu-cosamine (Fig.4,lanes2and3). BothBaEV-likegPr85env (Fig. 4,lane 5) and SiSV-likegPr8Oenv
(Fig. 4, lane 6) contained glucosamine. Antise-rum to GaLV (Fig. 4,lane 7) also precipitated anSiSV-likeglycoprotein, indicativeofthe high degree of relatedness between GaLV and SiSV. Inmmunoprecipitationwithanti-RLVgp7O (Fig. 4,lane4)precipitatedsmallamounts of only the SiSV-like envelopeproteingPr8Oenv.
Tryptic peptide maps of BaEV and HL23V precursor proteins. BaEV- or HL23V-infected cells were pulse-labeled with [3H]leucine and analyzed by SDS-PAGE and
FIG. 4. Glucosamine labeling of 7573/HL23V.
Cells were labeled for 6 h with [3H]glucosamine.
Equal portions of cytoplasmicextractwere
immuno-precipitated with anti-RLVp3O (lane2),anti-BaEV
p28 (lane 3), anti-RLVgp7O (lane 4), anti-BaEV(lane 5), anti-SiSV (lane 6), and anti-GaLV (lane 7). Lane Irepresentsamarkerof RL V-specific polyproteins.
autoradiofluorography. Bands exhibiting radio-activity as detected by fluorography were cut fromgelsand subjectedto tryptic digestion as previously described (1). The resultant polypep-tidefragmentswereseparated by ion-exchange chromatography (2, 7).
A comparison of Pr70-719'9 from BaEV-in-fected canine (CT/m7-BaEV) orhuman(A204/ m7-BaEV) cells with Pr70-71956 from HL23V-infectedcanine cells (7573/HL23V) isshownin Fig.5.Thepeptidemapofm7-BaEVPr70-715'9,
isolated from either infected canine or human
cells,appearedtobe identicaltothemapofthe
BaEV-like core-related Pr70-715'9 seen in HL23V-infected cells. All 21 to 23 leucine-con-taining peptide peaks seen in m7-BaEV Pr7O-719'9werealso represented inHL23V
Pr70-715'19.
Other experiments indicated thatPr70-715'9
fromcells infected with BILN-BaEVhas a pep-tide map identical to that of m7-BaEV Pr7O-71gag (datanotshown).The p28 content of the Pr70-719`9 proteins isolatedfromcells infectedwith any of the three BaEV isolates or with the HL23V isolate was
confinned bytryptic digest mapping (data not
shown). Mappeaks numbered5 to7, 9, 10, 12 to 15, 17, 18,and20werepresent in both p28 and Pr70-719`9 (Fig. 5).
As notedin the SDS-PAGE analysisof pre-cursorproteinsinCT/455K-BaEVcells
(Fig.
2,lanes1 and2), 455K-BaEV exhibitedan
appar-entlyuniqueprecursor
protein cleavage
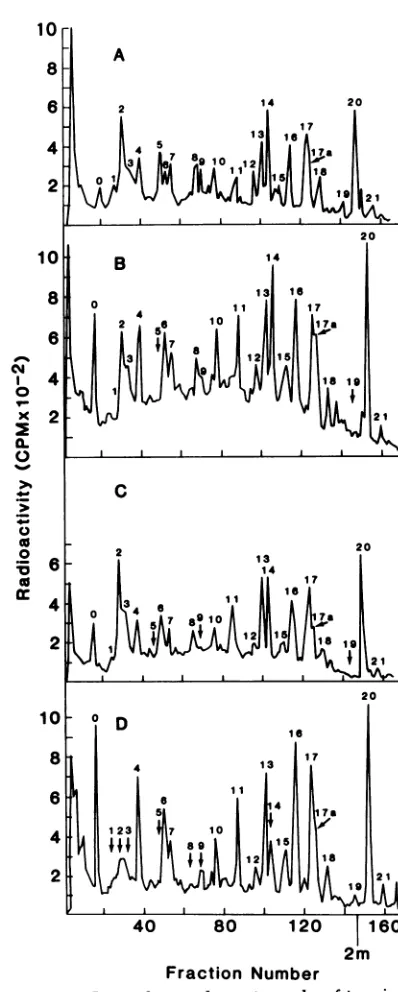
pattern.Figure6shows themapsof the
leucine-contain-ing peptides characteristic of the three
major
coreproteinprecursorsderivedfrom
CT/455K-BaEVcells.Astructuralrelationshipof
Pr85V9,
Pr709'9, and Pr679'9 fromCT/455KBaEVcells
was evident from these mapping studies. Also evident in Fig. 6D was the relatedness of the chase protein P72rg to thegagprecursor pro-teins. In this analysis no new
peptides
werepresent inP72w'9when itwascomparedwith the
pulse-labeled precursors
Pr85Y9,
Pr709'9, andPr67gag.Infact,P72gagwasmissingatleastone
peptide(peak 8) foundintheothercore
protein-relatedpolyproteins.
Avery similarrelationshipexisted among the coreproteinprecursors fromcells infectedwith the other BaEV isolates or with HL23V (data not shown). It was apparent from pulse-chase studies(Fig. 1and2) that thelevels of
P72w'9
in chase-incubated cells correlated well with the levels ofPr709'9 andPr70-719'9inpulse-labeledcells.Thisresultsuggestedthat
P72w'9
may havebeen derived from posttranslational modifica-tionof
Pr709"9
in455K-BaEV-infectedcellsorofPr70-719'9incells infected with the otherBaEV
isolates or with HL23V. It was not apparent from thesingle-dimensional analysis of
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.488.48.239.392.581.2]BaEV PRECURSOR POLYPROTEINS 865
that Pr70-71gwcontained onepeptide (Fig.7A, spot5a) whichwasmissing from P72wg (Fig. 7B, arrow). Likewise, P729ga contained onepeptide (Fig.7B, spot5b)which wasmissing from
Pr7O-7159(Fig. 7B, arrow). Wepresume that spot 5a
'_ NI
-
BBoI1
14 20012
L
7-2102
0IG. 7Inecagecrmtgapyo asn
0
8 ~~~~5
4 I10 13
co
BaEV(A),AX04/m7-BaEV(B and7573/H213519 21
1 2
C 2 14
10 7
1p0 20
8 4 p 13
178
6 0 351
4 ~~~~~~1112
2 2 1
40 80 120 160
2m Fraction Number
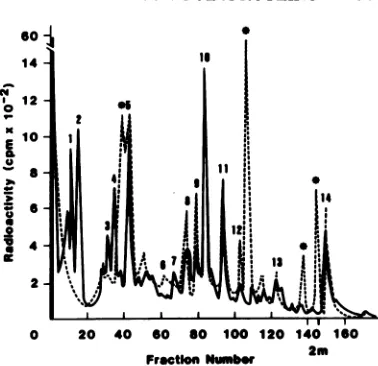
FIG. 5. Ion-exchange chromatography of
trypsin-digested[3HJleucine-labeled Pr7O-71""'from CT/m7-BaEV(A),A204/1m7-EaEV(B), and7573HL23eV(C).
Thepeptides are numbered forreference, andaheavy
bar indicates the elution ofthe column with 2 M
pyridineacetate(11).
cine-containing tryptic peptides howP72b'a
mighthave differed fromPr7O""gor Pr727w
Tomore
fully
consider therelationship betweenP72""9 and
Pr7o-719"",
we analyzed the trypticpeptides derived from these polypeptides by a
technique which allows the separation of
pep-tides intwodimensions. The resultsof this
anal-ysis (Fig.7) confirmedourpreviousobservation
that thepeptidemapsof Pr7O-719'w andPr72"'`
werenearly identical. It wasobvious, however,
40 80 120 1160
2m Fraction Number
FIG. 6. Ion-exchange chromatographyof trypsin-digested[3H]leucine-labeled polyproteinsfrom CT/
455K-BaEV.ComparedarePr85"" (A),Pr7Og"g(B),
Pr67"'' (C),andP72"- (D).
VOL. 37, 1981
CMJ 0
v-x
2 a.
0
o 0
06
co 0c
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.488.46.246.45.525.2] [image:6.488.244.443.128.624.2]866
(Fig. 7A) was the same peptide as that labeled peak 8 (Fig. 6B). The peptide in spot 5b (Fig. 7B) was apparently notseparated by the one-dimensional analysis (Fig. 6D) and may have been elutingin the void volume. Tryptic maps of P729'9 from cells infected with any of the BaEVisolatesorwithHL23V werenearly iden-tical (datanotshown).Likewise,tryptic maps of Pr85V9 from m7-BaEV and BILN-BaEV were identical to those ofPr85'9455K-BaEV (data not shown). The map of 455K-BaEV Pr7099 peptides was similar to the map ofm7-BaEV Pr70-719'9 (compare Fig. 6B with5A). The ab-sence of peak5 in Pr70O` derived from 455K-BaEV-infected celLsmay have been related to the differences in precursor profiles noted for this isolate in comparison with those for m7-BaEV(Fig. 2).
Figure8showsacomparison of theBaEV and HL23V BaEV-like glycoprotein precursor
gPr85env. Apparent immediately was the high
degree of relatedness togPr85env from cells in-fected with the various isolates of BaEV (Fig. 8A to C) and gPr85env from cells infectedwith the HL23V BaEV-like isolate (Fig. 8D). The
gPr85envproteins of m7-BaEVand455K-BaEV
exhibited identical tryptic peptide maps(Fig. 8A andC).The CT/m7-BaEVglycoprotein precur-sorapparently containedoneadditionalpeptide (Fig. 8B, peak 6) notseen in the other BaEV-infected cells. Like A204/m7-BaEV and CT/
455K-BaEV, HL23V gPr859'9 lacked peptide 6 (Fig. 8D) but contained one additional peak (peak 15). A comparison of the maps of the leucine-containing peptides of gPr85env from cells infected with m7-BaEV orBILN-BaEVis
shown inFig.9.Theenvelopeproteinprecursors of thesetwoBaEVisolates wereclearly
distin-guishable from one another by this mapping
procedure. As previously stated, however, the major core protein precursor,
Pr?Oga,
of these twoisolateswas indistinguishable bythis tech-nique(datanotshown).DISCUSSION
Several significant facts
pertaining
to BaEVreplication are apparent from the results pre-sentedinthisreport. First,BaEV coreand
en-velopeproteinsaresynthesizedininfected cells byprecursorpolyproteins,whichare
apparently
analogousto the precursors ofmurine retrovi-ruses. The immunoprecipitation-SDS-PAGEanalysesdescribedarecapableofdifferentiating
between the precursorpolyproteinsof m7-BaEV and455K-BaEV. We have also used 455K-BaEV obtained from CT/455K-BaEV to infect other canine thymus cells (CF2th). The SDS-PAGE profile ofpolyproteinsobtained from these cells wasidenticaltothat obtained from the original
CT/455K-BaEV cell culture (datanot shown).
This result suggests that the minor precursor
profiledifferences between m7-BaEV and
455K-BaEV are reproducible and virus specific. As
previously mentioned,peptidemappingstudies indicated an extensive structural relationship
among 455K-BaEV core protein precursors
Pr85Ya,
Pr709'9,
andPr67""
(Fig. 6). Asequen-tialprocessing from
Pr85Y'"
(Fig. 6A) toPr7Og"
(Fig. 6B) and finally toPr67Iag
(Fig.60)
was alsosuggestedbythe peptidemaps. ProcessingofPr85Y,if it
actually
occurs,apparently resultsinthe loss ofpeptides5and19 toyield
Pr7Ofgf,
FIG. 7. Two-dimensionalpeptide analysis
oftrypsin-digested[3H]leucine-labeledPr70-71'"
(A)andP72"w(B)from A204/m7-BaEVcells.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.488.49.446.443.631.2]BaEV PRECURSOR POLYPROTEINS 867
c 0
x
2
0.
1
0 0
10
FIG. 8. Ion-ex digested[3Hfleu
BaEV (A), CT/i
and A204/HL23
which subsequ Pr679'9.Altern is not a direct
10 60j
14 is
5 3 12
10
~~~~11~~~
9~~~~~FIG 9.Inecag~ 1hoatgah1ftysn
13. < befrom2di8tinctsynthesized but related
14~ ~
Typi dies aalye
lucn-cnaiinI precursors
suggest~~~
a
hihdge f eaens121 4
6 I yse 2f th neopeepodcsidiaena
10
0 20 40 60 60 100 120 140 1.0 FractionNumbar 2m
*
vME1E RFIG. 9. Ion-exchange chromatography of
trypsin-4 digested[3H]leucine-IlabeledgPr85nv fr-omA04/m
7-I WA J 14 BaEV(-)versusCT/BILN-BaEV(a b
12
Pr675,.
These relatedpolyproteins
actuallymaybe synthesized from distinct but related
mRNA's.
Tryptic
digest analysesofleucine-containing
10 peptidesof
SDS-PAGE-purified
BaEV-specificprecursors suggesta
high
degree of relatednesssamong
the threeBaEVisolates,especiallyinthe coreprotein-related gene products.
Similaranal-34^
eyses
of theenvelope
geneproducts
indicatenearg7t111 16
possidentity
for m7-BaEV and 455K-BaEV, but4 6 2 14 clearly
distinguish
these isolates fromBILN-BaEV. These observations are in
agreement
3 nwithpreviously published
estimatesof therela-Fato
Nutionship
amongvariousBaEV isolates based onimmunological and nucleic acid hybridization
data (9, 18, 19, 22, 24). Second, it isinteresting
that none of the cel lines, when they were
examined with anti-BaEVp28oranti-BaEV
se-rum, expressedanobvious
gagtpol
commonpre-I
(. lcursor
analogous
toPr2Oretlee observedinmu-rine retrovirus-infected cells (1, 2, 15). Several
14
possibilities
exist toexplain the
apparentab-34ntly
loses peptide 9 tosenceofthisexpectedprecursor.itis,ofcourse,12 16 possible that BaEV areunique among retrovi-ruses anddonotrequire synthesisandcleavage
ofa coreprotein
polymerase
read-throughtrans-_______________________lationproductforsynthesisof theirpolymerase.
40 80 120 1160 The absence of this common precursor may 2m indicate theexistence ofauniquesubsetmnRNA Fraction Number in BaEV-infected cells that codes solely for cchange chromatogaphy oftrpsn BaEV polymerase. It seems more likely,
how-tcine-1abeledgPr85enfromA2O4lm7_
ever, that the BaEV core protein polymerasemn7-BaEV (B), CT/4,55K-BaEV (C), precursor is synthesized in these cells but at
V(D). levels below thecurrentlevel of detection ofour
techniques. Thiswouldrequireasignificant
re-.iently loses peptide 9 to yield duction in polymerase synthesis in comparison
atively,it ispossiblethat Pr859a with that needed for other retrovirus systems
tprecursor to either Pr7OIag or (e.g., murine or avian retroviruses) in which
VOL. 37, 1981
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.488.255.444.61.244.2]868
Pr20O0""6P' is easily identified by these proce-dures. A third possibility is that antisera to
BaEVcore proteins do notimmunoprecipitate the BaEV core protein polymerase precursor polyprotein because of the alteration of core
proteinantigenicsites in thecommonprecursor. Further studies with antisera to BaEV
polym-erase will be required to explain the apparent absenceofPr2009`"-1°in BaEV-infected cells.
Athird interesting observation pertainingto BaEVreplicationis the presence ofa72,000-Mr, p28-related polyprotein
(P729'¶)
in extracts of pulse-labeled, chase-incubated BaEV-infected cells.P72w'9migratesuponSDS-PAGEslightly,but consistently, more slowly than does
pulse-labeledprecursorPr70-719'9.Thisrelatively
sta-blepolyprotein isnotlabeledduringshort pulse-labelingandis,therefore, apparentlynot a pri-maryproduct of BaEV mRNA translation. Syn-thesis ofP72rg presumably involves posttrans-lational modification of anothercoreprotein pre-cursor. As we have already mentioned, the amount ofP72rg in chase-incubated cells cor-relates relatively well with theamountofPr70ga
orPr70-719cI labeledduringpulses. This
corre-lation suggests that it is Pr709'9 or Pr70-719`9 thatis aprecursor toP72w'9.Thisinterpretation is favored by the results of peptide mapping experiments (Fig. 6 and7). The mostplausible interpretation of these results is thatpeptide 5a (Fig. 7A) in Pr70-719`9is altered by posttrans-lational modification to become peptide 5b in Pr729`9 (Fig. 7B). We could postulate that the alteration of this peptide is by glycosylation. Glycosylation isknown toresultinthe retarda-tionofprotein migrationuponSDS-PAGE,and glycosylated peptides often migratein the void volume upon column chromatography or near theorigin upontwo-dimensional analysis (L. J. Arcement, R. B. Arlinghaus, and R. B. Naso, unpublished data). Also, glycosylatedcore pro-teinprecursors have been observedin the mu-rine retroviral systems (6, 8, 30). Unlike the glycosylatedmurine retrovirus core protein pre-cursor gPr809'a, P72""9is an apparently stable intracellularprotein.Murine
gPr80O"'
is synthe-sized in pulse-labeled cells and is chased intogPr93-95Vg on the cell surface (30) and to a
lower-Mr precursor,
Pr45""9,
containing p15,ppl2, and theso-calledleader sequence (R. B.
Naso andR. B.Arlinghaus,manuscript in prep-aration).Furthermore, if
P72"ag
isglycosylated,it does notcontain enough glucosamine tolabel
with [3H]glucosamine (Fig. 4, lane 3). Similar
labeling experiments with [3H]mannose are in progress. The relatively large intracellular amountsof such astable core protein precursor undoubtedly relate to the rather poor
produc-tion of BaEV particles by most infected cells studiedin laboratories. Assuch,thepresence of
P72"''may representthefunctioning ofa
post-translationalcontrol mechanism in BaEV repli-cation.
Finally, this study has provided some inter-esting facts concerning the BaEV-like compo-nentof thehumanisolate retrovirus HL23Vand itsrelationship to trueBaEV isolates. It is ob-vious from the results described that HL23V containstwocomponents, one related to BaEV and theotherrelatedtoSiSV(SiAV).This find-ingiscertainlyin agreementwith the results of other analytical procedures (9, 19). It is also apparentthat antiseratoBaEV andSiSV(SiAV)
proteins, suchasthoseobtained fromthe Office of ProgramResourcesandLogisticsof the Na-tional Institutes of Health, are capable, when used in radioimmunoprecipitation-SDS-PAGE analyses, ofdistinguishingbetween the precur-sorproteins of thesetwocomponents in HL23V-infected cells. Thisfactis substantiatedbythe trypticdigest results presented here. Ithas
re-cently beenreportedthat thegenomeof viruses isolated fromHL23V-infected caninefibroblasts (A7573)wassimilarbyoligonucleotidemapping tothe genome ofSiAV (24). This report, how-ever, was unable to detect a species of RNA fromavirusreleased from these cells thatwas
similartoBaEV,suggestingthat the BaEV-like component of HL23V released from infected A7573cellscomprises less than 10% ofthetotal virus (24). The intracellular gene products re-sultingfromtranslation of theBaEV component RNA, however, canbeobserved readilyby
ra-dioimmunoprecipitation ofcell extracts, as
re-ported here. It is interesting to postulate that
thelowextracellular leveloftheBaEV-like com-ponent of HL23V relates to the synthesis of stable P72"'' in these cells. It has also been reported that HL23V derived from early pas-sages of the human peripheral leukocytes and obtained by cocultivation withhuman (NC37) cells orinfection of bat (CCL88) cells (10) con-tainedavirus componentthatwasidenticalto the San Francisco isolate ofGaLV(24). We have notobserved this component in A7573cells in-fected with virus derived from alaterpassage of the humanleukocytes.Itappears that a GaLV-like virus is the predominant species of SiSV-GaLV-like virus isolated from early passage HL23leukocytes,whereas anSiSV-likevirus is the predominant species of virus isolated from laterpassageleukocytes(24).
We have demonstrated that the intracellular proteinsof theBaEV-likecomponent of HL23V areidentical in size to theproteins of m7-BaEV. By tryptic peptide mapping, Pr709'9 from
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 37, 1981
HL23V is identical to Pr70O49 from human or canine cells infected withm7-BaEV. Compari-sonsof precursor polyproteinantigenic similari-ties,molecular weights, and peptide content il-lustratethat the BaEV-like viralcomponentof HL23V is more related to m7-BaEV than to 455K-BaEV. Similarly, antibodiesraised against m7-BaEVreversetranscriptase cross-reactmore significantlywithHL23V than with455K-BaEV (23). The envelope precursor of HL23V,
gPr85env, contains glucosamine and is nearly
identicalto boththe m7-BaEV and the 455K-BaEVenvelope precursors. Althoughthehuman isolate HL23V is showntocontain acomponent that is substantially related to BaEV, conven-tionalliquid hybridization studiesindicate that HL23V is not ahuman endogenousvirus (22). TheseresultssuggestthattheBaEV-like com-ponentofHL23Vrepresentsahorizontal cross-speciesinfectionof human cellswithm7-BaEV. The SiSV-like intracellular viral component of HL23Vwasisolated byimmunoprecipitation withseradirected against SiSV(SiAV) proteins. The SiSV-like proteins of HL23V are
distin-guishable from the BaEV-like component of
HL23V by analysis of intracellular precursor profiles (Fig. 3) and by tryptic digest analyses
(datanotshown). The HL23V SiSV-related viral
componentisreplicated by synthesis and
cleav-age ofamajor core protein precursor, Pr60`,
minor core protein precursors,
Pr20(Y"9°"
andPr80(c9,
and an envelope protein precursor,gPr8Oe"n.
A more complete characterization of the precursor polyproteins of the SiSV(SiAV)-like component of HL23V and their relation-ships to true SiSV(SiAV), GaLV, and another humanisolate, HEL-12V(20),will bepresented later (S.A.Whiteley,C.A.Edbauer, andR. B. Naso,manuscriptinpreparation).ACKNOWLEDGMENTS
Wethank Robert C. Galo for his gift of variouscelllines and antiserausedinthisstudy.We thank Maria Elena Leroux forexperttechnical assistance and Rebecca Bazer fortyping
themanuscript.
Thiswork wassupportedin partby grants MV-86 and NCS-245 from the American Cancer Society and Public Health Service Biomedical ResearchSupportgrant RR5511-18andCancer Center Core grant CA-16672 from the National Institutes of Health. R.B.N. isaLeukemiaSociety of America
Scholar, and S.A.W. is. the recipientofa Rosalie B. Hite
Fellowship.
LITERATURE CITED
1. Arcement,LJ.,W. LKarshin, R. B. Naso, andR. B.
Arlinghaus. 1977. 'gag' polyprotein precursors of
Rauscher murineleukemiavirus.Virology81:284-297. 2. Arcement, L, J., W. L. Karshin,R. B.Naso, G. A.
Jamjoom,and R. B.Arlinghaus.1976.Biosynthesis
ofRauscher leukemia virusproteins: presence of p30 andenvelopep15sequencesinprecursorpolyproteins. Virology69:763-774.
BaEV PRECURSOR POLYPROTEINS 869
3. Arlinghaus,R.B., R. B. Naso, G. A. Jamjoom, and L.
J.Arcement.1976.Biosynthesis of Rauscher leukemia
viral proteins, p.293-309. In D.Baltimore, A. Huang, andC. F. Fox(ed.),Animalvirology, vol. 4. ICN-UCLA Symposium on Molecular and Cellular Biology. Aca-demic Press,Inc., New York.
4. Benveniste, R.E., M. M. Lieber, D. M.Livingston,C. J.Sherr,andG. J. Todaro.1974.Infectious C-type virusfrom a baboonplacenta. Nature (London) 248: 17-19.
5. Bonner, W.M.,and R. A.Laskey.1974.Afilm detection
methodfor tritiumlabeledproteinsand nucleic acids in
polyacrylamide gels. Eur. J. Biochem.46:834-8.
6. Edwards, S.A., and H. Fan. 1979. gag-related polypro-teinsof Moloney murine leukemia virus: evidence for independent synthesis of glycosylated and unglycosy-lated forms. J. Virol. 30:551-563.
7. Eisenman,R., and V.Vogt.1978.Thebiosynthesisof oncornavirus proteins. Biochim. Biophys. Acta 473: 187-239.
8. Evans, LH.,S.Dresler,and D. Kabat. 1977. Synthesis
andglycosylationofpolyprotein precursorstothe
in-ternal core proteins ofFriendmurine leukemia virus. J. Virol.24:865-874.
9. Gallagher, R., and R. C.Gallo. 1975. Type-C RNA
tumorvirusisolated from cultured human myelogenous leukemiacelLs.Science187:350-353.
10. Gallagher,R.E.,S. Z.Salahuddin,W. T.Hall,K. B.
McCredie,and R.C.Gallo. 1975. Growth and differ-entiation inculture of leukemicleukocytes froma pa-tientwithacutemyelogenousleukemia and re-identifi-cation oftype-Cvirus. Proc.Natl.Acad. Sci. U.S.A. 72: 4137-4141.
11. Gabo,R.C.,and M.Gurof.1979.Type-specific binding
antibodytobaboonendogenousvirus(m7)reverse
tran-scriptase.J.Gen. Virol. 43:241-246.
12. Gilden,R.V., R.Toni,M. Hanson,D.Bora, H. P.
Charmen, andS.Oroszlan. 1974.Immunochemical studies of the major internal polypeptide ofwoolly monkey and gibbonape type-Cviruses. J. Immunol. 112:1250-1257.
13. Goldberg,R.J.,E.M.Scolnick,W. P.Parks,LA.
Yakovleva, and B. A. Lapin. 1974. Isolation of a
primatetype C virus fromalymphomatousbaboon.Int. J. Cancer14:722-730.
14. Haseltine,W.A.,and D. G. Kleid.1978.Amethod for
classificationof 5'-termini of retroviruses. Nature
(Lon-don)273:358-364.
15.Kopchick,J.J.,W.L. Karshin,and R. B.Arlinghaus.
1979.Trypticpeptide analysisof gagandgag-polgene
productsof Rauscher murine leukemia virus. J. Virol.
30:610-623.
16. Laenmmli, U. K. 1970. Cleavage of structural proteins duringtheassemblyof thehead ofbacteriophageT4. Nature(London)227:680-685.
17.Laskey,RIA.,and A. D.Mills.1975.Quantitativefilm detection of3Hand"4Cinpolyacrylamidegels by
fluo-rography.Eur.J. Biochem.56:-335-341.
18. Mishra,L,D.Hong,andMLA.Baluda.1979.
Homol-ogy betweenHL-23V andprimateviruses and search
forproviralDNA sequencesof simiansarcoma
associ-ated virus andbaboonendogenousvirusin DNA from humanleukemiccells.Leuk.Res.3:285-296.
19. Okabe,H.,R. V.Gilden,M.Hatanaka,R. J.
Stephen-son, R. E. Gallagher, R.C. Gallo,S. R.Tronick, and S.A.Aaronson. 1976. Immunologicaland bio-chemical characterization of type C virusesisolated fromhumanAML cells. Nature(London)260:264-266. 20. Panem,S.,E. V.Prochownik,F. R.Reale,and W.H. Kirsten.1975.Isolationoftype Cvirions fromanormal humanfibroblast strain. Science187:350-353.
21. Prekumar-Reddy,E.,S.G.Devare,R.Vasudev,and
on November 10, 2019 by guest
http://jvi.asm.org/
P. S. Sarma. 1977. Simplified radioimmunoassayfor viralantigens:useofStaphylococGusaureus as an
ab-sorbentforantigen-antibody complexes:brief
commu-nication.J. Natl. Cancer Inst. 58:1859-1861. 22. Reitz, M. S., N. R. Miller, F. Wong-Staal,R. E.
Gal-lagher, R. C. Gallo, and D. H. Gillespie.1976. Pri-matetypeCvirus nucleic acidsequences(woolly
mon-key and baboontypes)in tissues fromapatient with
acute myelogenous leukemia and in viruses isolated from cultured cellsof the same patient. Proc. Natl. Acad.Sci. U.S.A.73:2113-2117.
23. Robert-Gurof, M., and R. C. Gallo.1979.Type specific binding antibody to baboon endogenous virus (m7)
reversetranscriptase.J.Gen.Virol. 43:241-246. 24. Sahagan,B.G.,and W. A. Haseltine. 1980.
Relation-shipofretroviruses isolated from humanleukemia
tis-suestothewoolly monkey-gibbonapeleukemia viruses. J. Virol. 34:390-401.
25. Stephenson,J.R., and S. A. Aaronson.1977.
Endog-enousC-type viral expression inprimates. Nature (Lon-don) 266:469-472.
26. Stephenson, J. R., R. K. Reynolds, and S. A.
Aaron-son. 1976.Comparisons of the immunological
proper-ties oftwo structural polypeptides of type C RNA virusesendogenoustoOld Worldmonkeys. J. Virol.17: 374-384.
27. Theillen,G.H.,D.Gould,M.Fowler,andD. L. Dung-worth. 1971. C-type virus intumortissue ofa woolly
monkey (Lagothrix spp.) withfibrosarcoma. J. Natl. Cancer Inst. 47:881-889.
28. Todaro,G.J.,C. J.Sherr,and R.E.Benveniste. 1976. Baboons and theirclose relativesareunusualas pri-matesin theirabilitytoreleasenon-defective
endoge-noustypeC viruses. Virology72:278-282.
29. Todaro, G. J., C.J.Sherr,R. E.Benveniste, M. M. Lieber, and J. L.Melnick. 1974.Type C viruses of baboons: isolationfrom normalcellcultures. Cell 2:55-61.
30. Tung, J.-S.,T.Yoshiki,and E. Fleissner. 1976.Acore
polyprotein of murineleukemia virusonthe surfaceof
mouseleukemia cells. Cell 9:573-578.
31. Wolfe,L.G.,F.Deinhardt, G. H.Theilen,H.Rabin, T.Kawakami,andL. K. Bustad. 1971. Induction of tumorsin marmosetmonkeys bysimiansarcomavirus, type1(Lagothrix): preliminaryreport. J.Natl.Cancer Inst.47:1115-1120.
on November 10, 2019 by guest
http://jvi.asm.org/


![FIG. 7.(B) Two-dimensionalpeptide analysis oftrypsin-digested[3H]leucine-labeledPr70-71'" (A) andP72"w from A204/m7-BaEV cells.](https://thumb-us.123doks.com/thumbv2/123dok_us/1482770.100914/7.488.49.446.443.631/dimensionalpeptide-analysis-oftrypsin-digested-leucine-labeledpr-baev-cells.webp)