JOURNAL OFVIROLOGY, May1978, P. 389-410 Vol. 26,NO.2
0022-538X/78/0026-0389$02.00/0
Copyrighti 1978 AmericanSocietyforMicrobiology Printed in U.S.A.
Anatomy
of
Herpes Simplex Virus (HSV) DNA
X.
Mapping of Viral Genes by Analysis
of
Polypeptides
and
Functions
Specified
by
HSV-1 x
HSV-2 Recombinants
LAWRENCE S.
MORSE,'
LENORE PEREIRA,' BERNARDROIZMAN,I*
AND PRISCILLA A.SCHAFFER2
MarjorieB. Kovler ViralOncologyLaboratories, University of Chicago, Chicago, Illinois60637,'and
Sidney
Farber CancerInstitute,Harvard MedicalSchool,Boston,Massachusetts 021152 Received for publication 13 October 1977In an
earlier
paper(Morse et al., J. Virol. 24:231-248, 1977) we reported on theprovenance of theDNA sequencesin26herpessimplex virustype 1 (HSV-1) x
HSV-2 recombinants as determined from analyses of their DNAs with atleast
five restrictionendonucleases. Thisreport deals with the polypeptidesspecified
by the recombinants and
by
their HSV-1 and HSV-2parents.Wehave identified(i) the corresponding HSV-1 and HSV-2 polypeptides with molecular weights
ranging from 20,000tomorethan200,000,(ii)the polypeptides that undergorapid
post-translational processing,
and(iii) polypeptides
that vary intratypically inapparentmolecular
weight. By comparing
thesegregation
patterns of thepoly-peptides with those of the DNAsequenceof therecombinants,wehave mapped
the
templates
specifying
26polypeptides
and several viral functions on thephysicalmapof HSV DNA. The data show thefollowing: (i)apolypeptidesmap
atthe terminiof theLandScomponentsofthe HSV DNA.AlthoughaICP27
mapsentirely within the reiteratedregion ofthe L component, the template for
aICP4maylie
only
inpartwithinthe reiteratedsequencesof theScomponent.Ofnoteisthe
finding
thatcellsinfected witharecombinantthat contains bothHSV-1 and HSV-2 DNAsequencesin theS componentproduced a ICP4of both
HSV-1and HSV-2.
(ii)
Templatesspecifying,8
andypolypeptidesmapinthe Lcomponentand appearto be
randomly
distributed. (iii) Thymidine kinase andresistance to
phosphonoacetic
acidmapped in theL component.Inaddition,wehave takenadvantage of the rapid inhibition of host protein synthesis
character-istic of HSV-2 infections and
syncytial plaque morphology
to also map thetemplate(s) responsible
forthesefunctions in theL component.Theimplications
of thetemplate arrangement in HSV DNAarediscussed.
In this paper we report the
location,
in the arethefollowing
data:(i)
Honess andRoizmanDNAs of herpes
simplex
virus types 1 and 2 (13)reported
that HSV-1specifies
in infected(humanherpesviruses1and 2;HSV-1 and HSV- cellsatleast48
polypeptides
ranging
from 20,002), of the
templates
specifying
26 polypeptides to greater than 200,000 in molecularweight.
and several viral functions. The
experimental
Several additionalpolypeptides
less than20,000design
used in these studieswassimilartothat inmolecularweight
werereported
by
Marsdenused in
mapping
adenovirustemplates (9, 25)
et al.(24).
Asimilarlist
ofHSV-2polypeptides
andconsisted of
comparing
thesegregation
pat- wasreported by
Powell andCourtney
(30).
Stud-terns of
polypeptides specified by
HSV-1 x iesby
Courtney
and Powell(3),
Pereira etal.HSV-2intertypic recombinants with their DNA (29), Gibson and Roizman
(8),
and Cassai etal.sequence arrangements.The DNAsequencear- (2) showed thatmany HSV-1 and HSV-2 virion
rangements of the intertypic recombinants used polypeptides and infected-cell polypeptides
in these studies were reported in a previous (ICP)differ in
electrophoretic
mobility,
butonly
paper in this series (26). We nowreportonthe a few of these
polypeptides
were identified aspolypeptides producedincellsinfected with the functionally identical.
intertypicrecombinants andonthephysicallo- (ii) The
synthesis
of bothHSV-1 and HSV-2cation in HSV DNA of thetemplates specified polypeptides is regulated. On the basis of the
bythem. temporal
pattern
andrequirementsfor theirsyn-Pertinent totheresultspresented in this paper thesis,HSV-1ICPswereshownto
form
atleast389
on November 10, 2019 by guest
http://jvi.asm.org/
390 MORSE ET AL. J. VIROL.
threegroups,designateda,
,8,
andy,whosesyn- MATERIALS AND METHODSthesis is coordinately regulated andsequentially
ordered inacascadefashion (14, 15,32-34). Radiochemicals. L-[U-14C]isoleucine,
L-[U-_4C]-(iii) HSV-1 and HSV-2 DNAs are linear dou- leucine, andL-[U-_4C]valine (allwith specificactivities
ble-stranded molecules approximately 97 x 106 of approximately 300 mCi/mmol) were purchased
to 99 X 106 in molecularweight (21, 39). HSV-1 fromNewEngland Nuclear Corp.,Cambridge,Mass.
DNA consists oftwo covalently linked compo- Cels. HEp-2
ceUs
were grown in Eagle minimalnents, L and S, containing 82 and 18% of DNA, essentialmediumsupplementedwith 10% calf serum,
respectively.
The terminal reiterated sequence 0.001% ferric nitrate, and 1% sodium pyruvate.bracketing.thne
Lcomponent,designated
asequence
Viruses. The virus strains used in thisstudywerebracketing the L component, designated as ab, (i) HSV-1 (KOS tsE6), a DNA' ts mutant, and
HSV-and its inverted repeatb'a'each contain 6.0% of 2 (186
tsB5),
a DNA- mutant described elsewhere(35,the DNA, whereas the sequences bracketing the 36); (ii) HSV-1 (17 tsJ), a DNA- ts mutant (24, 38)
S component, designated as a'c' and ca, each kindly provided by J. Subak-Sharpe; (iii) HSV-1
contain4.3% of the viral DNA(32,33, 39). DNA (HFEMtsN102),aDNA- mutantkindly providedby
extracted from virions or from infected cells A. Buchan; (iv) HSV-1 (B2006 ts-), kindly provided
consists of fourequimolar populations differing by S. Kit (20); (v) HSV-2(186), theparent strainof
in the orientation of L and S components
(10,
HSV-2 (186 tsB5); (vi) HSV-2 (GP6), asyn-
mutant33
39).
StudiesofHSV-2DNA withrestriction (1)of HSV-2 (G); (vii) HSV-2(GP6PAAN),
amutant39).
Studies
ofdHSV-2
DNa
with res
trsictio
ofGP6resistant
tophosphonoacetic acid (PAA),se-endonucleases
iiaycate
that ithas
a similar lectedbyprocedures described in reference 26; (viii)structure (G. S. Hayward, T. G. Buchman, and HSV-1 (F) and HSV-2 (G)
(32-34);
and(ix)
24inter-B. Roizman, manuscript in preparation). Anal- typic recombinants derived by crossing HSV-1x
HSV-yses of HSV-1 x HSV-2 intertypic recombinants 2 parental strains (26). Virus stocks were prepared
reported in the preceding paper (26) suggested fromplaque-purified seedaspreviously described (26).
that only a limited number of HSV DNA ar- Labeling ofproteins synthesized by infected
rangementsparticipatedin theformation of re- celis. Confluent HEp-2 cell monolayers in 25-cm2
combinants and by extension were capable of tissueculture flasks wereexposed to 20 PFU ofvirus
replication. The HSV DNAarrangement shown per
cell
in 1.0ml of maintenancemedium. After1hoftorbeP11at10n.e
topaicipateinteforaementinown
o incubation of33.5°C
with constantagitation,
thein-to be able in-toparticipate in the formation ofall oculum was replaced with 5.0 ml of maintenance
me-recombinants analyzed in the preceding study dium, and incubation was continued at 33.5°C. For
wasdesignated as prototype (P) (26). Thethree. labeling polypeptides, the cultures were replenished
otherarrangements were designated as Is, inver- with labeling medium containingone-tenththenormal
sion of S component; IL, inversion ofL compo- amountofleucine, isoleucine, and valine, but
supple-nent;andISL,inversion ofboth L and S compo- mented with
["4C]leucine,
-isoleucine, and -valine (2.0nents. For the sake of simplicity, only the P uCi of each amino acid per ml of medium). For labeling
arrangements of HSV-1 andHSV-2 are shown early viralpolypeptides, HEp-2cellswereexposedto
inthispaper. virusat370C for 1 h and then labeled for 2 h at39°C.
mi
this
paper. y At the end of the labeling period, the cellswere(iV) The location of templates specifying rinsed with ice-coldphosphate-bufferedsaline (3 x 5.0
knownviralfunctionsin HSV DNAarelargely
ml/flask)
toterminate incorporation, either harvestedunknown. Previous studies have shown that immediately (pulse) or washed, and then reincubated
viralRNAsequences accumulating in thecyto- in the absence oflabeledaminoacids(chase).
plasm in thepresence ofcycloheximide (aRNA) Preparation of samples for
electrophoresis.
hybridize
predominantly with terminal frag- The labeledcells werestripped from the dish,dena-ments ofL and S components and to a lesser tured, and
solubilized
byheatingfor 2 to 3min
at80°Cextentwith internalfragments intheL compo- in the presence of 2%sodiumdodecyl sulfate (SDS),
nent (19). 5',B-mercaptoethanol, and 0.05 M Tris-hydrochloride
(v)Rcentlwerportdon he usfulnes of (pH 7.0).
(v) Recently wereported on the usefulness of
Polyacrylamide
gel electrophoresis. Theelec-intertypic (HSV-1 x HSV-2) recombinantsfor trophoretic, staining, andautoradiographictechniques
mapping viral markers on the DNA (26). The were as described previously (11, 37). The
polyacryl-recombinants used in these studies were pro- amide gelelectrophoresis was done in a discontinuous
duced by crossing ts HSV-1 x tsHSV-2 or by buffersystem containing0.1% SDS. Thestacker and
crossing tsPAAr HSV-1 x HSV-2. The prove- separation gel contained 3 and 9%acrylamide,
respec-nance of the DNA sequences in the cloned re- tively, cross-linked with N,N-diallyltartardiamide
combinants were determined by mapping with (Aldrich Chemical Co., Milwaukee, Wis.) in an amount
Hpa I,
Bgl
restr,BglII,XbaI,Ection ThuI,and,ioe
II,XbaI, EcoRI,HsuI,and,insome Thecorrespondingseparation gel
to 2.6% of the weight of acrylamide.was20cmin
length.
Theproteins
instances,K~pn Irestr±ctl endonucleases. The used for molecular-weight calibration were
f,',
,B,andnomenclatureofthe recombinants, themapping asubunits of Escherichia coli RNApolymerase,
bo-procedure,and detailedrestriction endonuclease vine serum albumin, and soybean trypsin inhibitor
maps werereportedinthepreceding paper (26). (T,) (Boehringer Mannheim, Indianapolis, Ind.) with
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 26, 1978 ANATOMY OF HSV DNA 391
molecularweights of165,000, 155,000, 39,000, 69,000, vary intratypically among HSV-1 and HSV-2
and21,000,respectively. strains, (iv) provisionally matches the
corre-sponding HSV-1 and HSV-2 polypeptides, and
RESULTS (v)describes thekinetic classto whichthe
poly-Comparison of HSV-1 and HSV-2 ICP. peptidesbelong.
Table 1 summarizes the data derived in the The experimental data relevant to the
con-courseof these studiesontheapparent molecu- structionofthistableare asfollows:
larweights and otherproperties of HSV-1 and (i) The criteria for
identifying
virus-specificHSV-2polypeptides. The polypeptideswere des- ICP was that of Honess and Roizman (13) as
ignated accordingto Honess andRoizman (13) applied by them foridentifying HSV-1
polypep-as extended by Pereira et al. (29). Thus, the tides and byPowelland Courtney (30) for
iden-polypeptidesarenumberednumericallyin order
tifying
those of HSV-2. We have largely retainedof decreasing molecular weight (13). Polypep- the numerical designation ofHoness and
Roiz-tides undergoing rapid post-translational proc- man (13) for HSV-1 ICPs and, for simplicity,
essingareidentifiedby number and lettersa, b, assignedtoHSV-2ICPnumbers that correspond
c, etc.,
denoting
theprecursor(a) andproducts tothoseof HSV-1 ICP.(b, c, etc.) (29). Table 1 (i) enumerates HSV-1 (ii) Identification of polypeptides whose
mo-and HSV-2 specific polypeptides in infected-cell lecular weightswerealteredas aconsequenceof
lysates, (ii) identifies the polypeptides thatun- rapidpost-translational processingwasbasedon
dergorapid post-translational processing, result- experiments involving pulse-labeling followed by
ing in an
appreciable change
in the apparent anappropriate
chase in the absence ofradioac-molecular weight, (iii) lists the polypeptides tive precursors. An example of the results of
[image:3.509.52.451.330.637.2]whoseapparentmolecularweightwasfoundto such anexperiment is shownin Fig. 1, where we
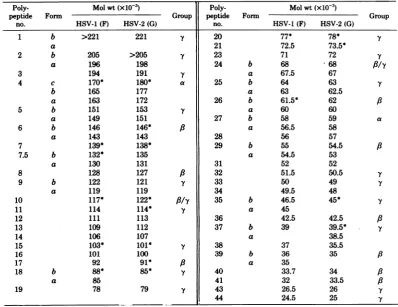
TABLE 1. ElectrophoreticpropertiesofHSVpolypeptidesa
Poly- Molwt(x10-3) Poly- Molwt(xlO-3)
peptide Form Group peptide Form Group
no. HSV-1(F) HSV-2(G) no. HSV-1 (F) HSV-2 (G)
1 b >221 221 Y 20 77* 78* Y
a 21 72.5 73.5*
2 b 205 >205 Y 23 71 72 Y
a 196 198 24 b 68 68 /Y
3 194 191 Y a 67.5 67
4 c 170* 180* a 25 b 64 63 y
b 165 177 a 63 62.5
a 163 172 26 b 61.5* 62 /3
5 b 151 153 .Y a 60 60
a 149 151 27 b 58 59 a
6 b 146 146* a a 56.5 58
a 143 143 28 56 57
7 139* 138* 29 b 55 54.5 a
7.5 b 132* 135 a 54.5 53
a 130 131 31 52 52
8 128 127 /3 32 51.5 50.5
-Y
9 b 122 121 Y 33 50 49
a 119 119 34 49.5 48
10 117* 122* //y 35 b 46.5 45* Y
11 114 114* Y a 45
12 111 113 36 42.5 42.5 /3
13 109 112 37 b 39 39.5* Y
14 106 107 a 38.5
15 103* 101* Y 38 37 35.5
16 101 100 39 b 36 35 /3
17 92 91* /3 a 35
18 b 88* 85* Y 40 33.7 34 A
a 85 41 32 33.5 A
19 78 79 y 43 26.5 26 Y
44 24.5 25
-Y
aThe polypeptides are numbered numerically in order ofdecreasing molecular weight. The table also
identifies thepolypeptidesthatundergo changesin theirelectrophoretic mobility,whicharedesignatedasa,b, or cforms.Polypeptides showingintratypic variabilityaremarked withanasterisk.
on November 10, 2019 by guest
http://jvi.asm.org/
392 MORSE ET AL.
J.
VIROL.
HSV-1
CYCLO
H SV- 2
3
4
C
~~~~~~~~~~4
a)4 - 4 4
5
5we
m
5O~54
S
v~~
S6_I6
a
US 61S
j
nn
0bl10
9afb
9
11 ab I19'1 '- b1
15
;8 17
24 C 24
25
-26
nU.AJW7
~
427--
- 7 _-4
-,
22UU7
JUl
32 32
34
344
4W
153
,,W,,U_'151
S
j
35-o-
e
ae
- me
-37 a
38
39
ekf
cut0.
5m41
P
C
P
C
P
C
P
P
C
PC
P
C
3
hr
4
hr
5hr
3
hr
4hr
5Shr
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 26,1978 ANATOMY OF HSV DNA 393
show the polypeptides undergoing rapid post- peakrates ofsynthesis between 5 and 7 h
post-translational cleavages relativelyearlyin infec- infection and were identified on that basis.
Iden-tion. Some of these (e.g., ICP4and6) have been tification of y polypeptides was based on the
previously showntobeprocessed after synthesis. observation that they are made at increasing
Table 1lists 16polypeptides thatwerefound to ratesuntil12 to 15hpostinfection.
besignificantly alteredinelectrophoreticmobil- (v) Identification of corresponding HSV-1 and
ity after synthesis. This is a minimal estimate HSV-2 ICPswasbasedonthe following
ration-becausepost-translational modifications that do ale. Comparison of the twosetsof ICPs showed
notresult inaperceptible change in electropho- that they fall into two classes. The first
com-retic mobility wouldnotbe detected. Two points prised HSV-1 and HSV-2 ICPs that fell into
are noteworthy concerning the processing of identical kinetic classes and didnotdiffer with
polypeptides. First, with onlyoneexception,all respect to electrophoretic mobility. Although
processed forms of the polypeptides showed a they are not central to the objectives of this
decrease in
electrophoretic
mobility. Secondly, study, for the purpose of identification weas-weresolved in these experimentstwobands for sumed that they have identical functions and
the major capsid protein, ICP 5. Pulse-chase numbered them accordingly. The second set
experiments of the kind described in the legend comprises HSV-1 and HSV-2 ICPs that could
to Fig. 1 failed to show a product-precursor notbematched withrespect toelectrophoretic
relationship. One possible interpretation of our mobility. Examples of such are ICP 4 and 27,
observations is that of a fraction of ICP 5 is whichfallintothea group,andICP5,member
altered during assembly of the HSV capsids. of the y group(14). These polypeptidesare
cen-Thedesignations5aand5b are arbitrary. tral to theobjectivesof thisstudy because
iden-(iii) Intratypic variability of structural poly- tification of the HSV-1 andHSV-2 polypeptides
peptides among HSV-1 strains has been re- specified by therecombinantswasbased inpart
portedpreviously (28). Comparisonofthe paren- ontheirelectrophoretic mobilities. Thepairing
tal strains usedto construct the recombinants of thecorrespondingpolypeptideswasbasedon
revealed numerous ICPs that varied among analysesofICPs specifiedby therecombinants
HSV-1 andamongHSV-2 strains. The dataare andrestedon twocriteria. First,withone
excep-basedonelectrophoreticmobility of the labeled tion identified later in thetext, thepresence in
ICPs on the same gels. Some of the data are thecell lysate ofapolypeptide specified byone
shown in
Fig.
2through 10 andsummarized in serotype excluded the presence of the other.Table 1. Aninteresting example ofanonstruc- Second,eachICP specifiedbyHSV-1orHSV-2
tural
polypeptide
thatappeared to be variable could be mappedindependently as eitherpres-wasICP 4;asshowninFig. 2, the fully processed ent or absent, and in each instance the paired
form (ICP 4c) of HSV-2 (G) migrated more ICPsmappedinthesamelocation.
slowly
than the corresponding polypeptide of Mapping of HSV polypeptides. Inprinci-HSV-2 (186). ple, by correlating the segregation patterns of
(iv) The identificationof anICP as a,
f,,
or y the polypetides specified by the recombinantwasbasedonthefollowing operational definition viruses with the DNAsequence arrangement of
andonthebasis of the kinetics oftheirsynthesis the recombinant virus, it should be possible to
(14, 15).Bydefinition, apolypeptidesaremade map the physicallocation of the templates
spec-immediately
after withdrawal ofcycloheximide ifying viral polypeptides on the HSV genome.presentin the mediumduring infection andfor The experimental procedure was as follows.
several hours thereafter (14, 15). An example of Cells infected with parental strains or with
re-a cycloheximide withdrawal experiment is combinants were pulse-labeled at three time
in-shown in
Fig.
1.Inthatexperiment,
only ICP4 tervals after infection as described in Materialsand27 weremade inHSV-1-infectedcells after andMethods and in the legends to Fig. 2 through
withdrawal of the drug.a Polypeptidesreached 10. Table 2 shows thesegregationpattemofthe
peakratesofsynthesis between2and4hpost- paired HSV-1-HSV-2polypeptides whose
tem-infection. In contrast, ,B polypeptides reached plateswere mapped. Figure 11 shows the DNA
FIG. 1. Autoradiographic images of electrophoretically separatedpolypeptides from HSV-1- and
HSV-2-infectedcells.HEp-2cellswereinfectedwitheitherHSV-1(F)orHSV-2(G)strains and thenlabeledfor15
minwith
"4C-amino
acidsat the times indicated. At the endof the pulse-labeling period, the inoculated cultureswereeither harvestedimmediately (P)orchasedby incubation inunlabeled maintenancemediumfor anadditionalhour(C). Thecycloheximidewithdrawalexperimentwasperformedby incubating HEp-2cellsinfectedwithHSV-I(F)in thepresenceof50 pgof cycloheximidepermlofmedium.After 7h,thecycloheximide
waswashedoutandreplaced with"C-aminoacidlabeling medium. The cellswereharvestedafter 15 minof incubation.
on November 10, 2019 by guest
http://jvi.asm.org/
394
MORSE ET AL. J. VIROL.Series A and B
4 44 4
4W
4 4 4 4 4 4 165155
cre5_
WP5 5;_17 7 74117 7 7J7 7 7 7 7 7
7e7
t 8 8S8 8 84_8 8 8 8 8 8 8S_8
10 tO
~~~~~~~~~~~~~~~101
10ITse -w
1S 15 15 15
_ow7aA 17
-
18si8
.\4A. .S A 20 20
P't.
24~~~~~~~~~~~~~~2
: 25
26
__ _
534
oiLL-i
-_
_SIO
O
eO
e
4
39
[image:6.509.70.456.69.586.2]G tsJ
A1EJ
GP6 A7DA1G3
GP6 A5C A2DA4D tsJ A6D Bl E 186 mockFIG. 2. Autoradiographic images of electrophoretically separated polypeptideslabeledwith
"4'C-amino
acidfr-om1to3hpostinfection.ParentalstrainsforseriesAintertypicrecombinantswereHSV-1(tsJ)and
HSV-2(0P6). TheparentalstrainsforseriesBwereHSV-1(tsej andHSV-2(186).
sequence arrangementofthe recombinants de-
fying
IOP
5 illustrates themapping procedure.
termined
previously
(26).Figure
12 shows theThus,
examination of theIOP
5specified by
physical
location of thetemplates
mapped
in recombinants C4D andA6D (Table
2;Fig.
4, 7, thisstudy.
Themapping
of thetemplate speci-
and10) indicatesthat thetemplate (Fig.
11) ison November 10, 2019 by guest
http://jvi.asm.org/
VOL. 26, 1978 ANATOMY OF HSV DNA
395
Series A and B
4 4 4 4 4 4 4 4
6a,4
_6_s z4hi- -.q.5
S F5emw<m5-
--S
JWJ*4
~~aB._nri~~
5n&
5_1
le IAD 7DulO.0aw-Of7
1St' 41 ~ 4 ...4.s.4n141Wj1
GP6AlEl A1G3 tsJA7
A2D
TIA4DOG6SIADAE
with~~~~zseriesjrA in*teryi reobiat frm . to 6 h potnecin
~~~~~~~~~~~~~~~~~~~~~~~~~~~-18
4 4Do-.4 "OW0-rM ~, do*-_ 4
41 4 - 41 41
-.4
443
43Vp,
43 4.4
4Q
3'43|
--,e,3 A44 44 44. 44 44 44 44 4-4 44 44 4
[image:7.509.106.389.66.621.2]GP6
AlElA1G3
tsJ A7D A2DA4D
GP6A5C
A6D A1E2FIG. 3. Autoradiographic images of electrophoretically separated polypeptides labeled in cells infected
with series Aintertypicrecombinants
firom
4.5to6hpostinfection.on November 10, 2019 by guest
http://jvi.asm.org/
Series
A
and
B4 4
w7
7mr=r
=
18
f8
*w.
4;
8
*b..8
8
8
7}t?@i8*#$-8
'
8a
8 8.st
"8
999999 9 9 9
10 0 ?.1
4MW _
fl
4S#
PSI
_1M1
1
8_
_ 18 -;
18*P'84+
'18
18~
- ss 1 8 '18*Lr18
1 -eA 18w I
___.'
I_
'33_:23;
72223:.0
23
2
4t2wmW*..24~lY E:=4 wQ4
_24,
...~~~~~~~~~~~~~~2 . w.WaoQ 27 - 2727
35
-~~"40~ 40'
-t394~39_
39
df9l39
*4
GP6
AlE)AlG3tsJ1
41
41
4
7
4D
n404Au11
As
.4_t40 44 44.0E18A
E16
441
4 [image:8.509.82.453.45.645.2]41~
~~~~~~
41a
'4~
~~~~~~~~~F
416IMW-TFIG. 4.
AutoradiograpI
iaeo et
s
infected
wIth seriesAuinter
gryphicrcmbnans
ofro9etro1hposticy
e p infectl e in clwithseries A
intertypi
recomb tsfm
9to po 40on November 10, 2019 by guest
http://jvi.asm.org/
Series C
UF"
fl4IIIS44_
*t44
= S4fl4
a_ 5i _5a 5
SE7w-7aiOSz
.7inZ
~7
_
7'
77fl
7
.., ..10
eI fi aa4Taf_ 8_ *~~1 8w am4
-15
:-
151
5,-15-1 -
-.S-15auL 18a-15-Ia
-@-20;)2D
a2O
20
20 20-~
~~~~~~~~~~
4-4*tU C.
__*-r4U
'
PM-~a
_ _ 334 34
_O O4ine 0ta
G
N102 C2D
186
C3DI
C4D
N102
C5D C6D C7D 186
mock
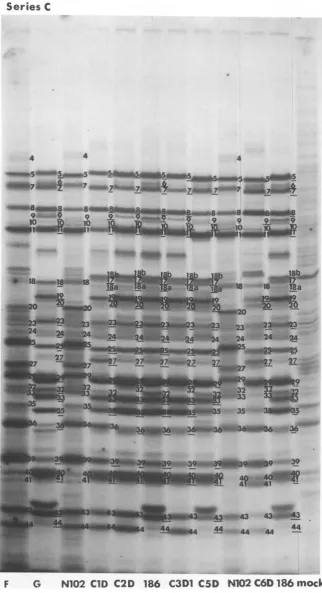
FIG. 5. Autoradiographic images of electrophoretically separated polypeptides labeled in cellsinfected
with series Cintertypic recombinantsfr-om 1 to3hpostinfection as described inMaterials andMethods. Parental strainsforseriesCintertypic recombinantswereHSV-1asNlo2 andHSV-2(186).
.b_i~~ ~~~~~9
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.509.78.422.45.647.2]398 MORSE ET AL. J. VIROL.
Series C
4
4 4 44 4
4 -,a4 4
5-5Amm5
5-ma. 5A 5-. 5__5_5 50oo5-7esLUIf eW b f7~7u7fl.0#Fa
8 cm
N3
eu> 8s aa m a n.. .XW5th_ e
18
4000
a
-4 2 0 2Q
~~~~~~20
20a~*}.
->s.ti
_
1
jje_~
?
__*~n3~
3__
4_07
7 WI__fS
35
|l35
5
|
5
SX
35
3535
4 * e 4 35 -G40
;F41
40 40,;00A|-m41 M41_
00-743 43 -4 3 4- .& 43_
44 44 44 4 44nA444 44 44
F
GN102
ClDC2D
186
C3D10NI2
C5D C6D
186FIG. 6. Autoradiographic
images of
electrophoretically separated
polypeptides
labeledin cellsinfected
with seriesCintertypicrecombinantsfrom
4.5to6hpostinfection.
on November 10, 2019 by guest
http://jvi.asm.org/
Series
C
4 4 4
-~~~-
*
cIinsSSin5-amIgI7aa~glom
7004WZM7&4aas.tmo7flO
7007 ILh4_9
^~~~00
99
9j"n_49
a"1
ORa
_
__.v
24*24
..
i36-~~~~~~~~~~~~~~3
F
0
N102
CID
C2D
186
C31D1
C5D
N1O2
C6Dl86
mock
FIG. 7. Autoradiographic images of electrophoretically separated polypeptides labeled in cells infected
with series Cintertypicrecombinantsfrom9to10hpostinfection.
o399
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.509.92.415.46.640.2]Series
D
4 4!;4r44 4 4
S-W~~~
4~~~~~~~~~~~~~~~~
4Sqw
4~~~~~~~~~
=4 4 4
v 5 5
.ub7? 7uIUht_ _ 7 7a1t *7* .7 7 7 7 7
CP8- faS^Uh8iB _ usu8e.e a8 8 8 8 8
10
15 .15
-17t-- *< <
18B
58a
20
2~~~~~~4
20* teU 4ibP~'W 'f
- 4j -D --,¢t.
_.
a
a aaa
a
SS
.40~~ ~ ~~~~0
-_9F~~~~~~~~~~~~~~~~~~~~~~~~~~~~~v
gR.7
G sE
DEIAE
4E DE D E tE D4ED5E
D5Emock:re
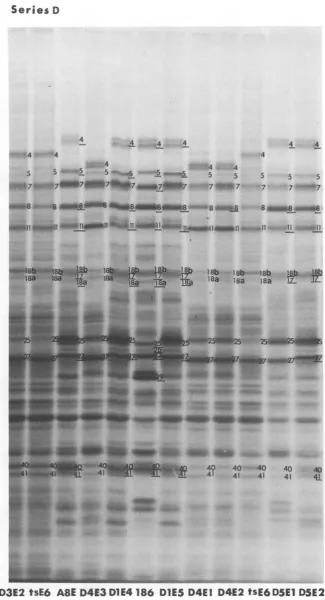
lPW}; FI. Auordogapi image of_ elcrohrtial seaaedoyepie labeled_ in celsifetewitseie D-inerypi reobiat fro t3hpoinein. Paena stais fo eisDitryi
recombinantaweeHV1(O tWsE6 an HS-16tB
on November 10, 2019 by guest
http://jvi.asm.org/
Series
D
4 4 4 4 4 4
4 4 4
5 5 5s-,.6 5 5 5 5 5
Mpw
7
0*wA7 IM7Ul7IU77
717
77
7W?
7
7
n-7
_ w8-.-4 j E sm8 88 3 8e8_naB
ugah
_nw9
nub
Zll _11tUns_
l W*A6.RJl 119'el
18 8-- IL 18b
l8b
- 18b yjj s*-w^1*}Y-1856 *118b8a*riWu) l18a Ila 185 ]L.Z.
40
40 40
40
40
41 41 41 41
41-D3E2
tsE6
A8E D4E3
D1E4
186
DIES D4E1
D4E2
tSE6
D5E1
DSE2
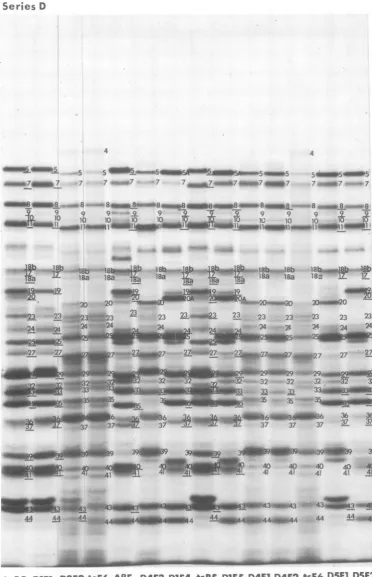
FIG. 9. Autoradiographic images ofelectrophoretically separated polypeptides labeled in cells infected
with series Dintertypicrecombinantsfrom4.5 to 6hpostinfection.
-;t ~~~~~40
on November 10, 2019 by guest
http://jvi.asm.org/
[image:13.509.87.413.44.645.2]Series
D4 4
_t_555 4_5e.e5eI 5R u5e5 5
54n5
a7ain7 7 7 7 7 7
d-777
7.7
7080__-8 8 8ee..--8--8
ewwin8
ea e e8m 8 a A 899 9 9!.1992-9
10 lO 10O T10 10 10IL 10 10 o 10
IO.b~~~
1 18b 1 #9
b_1ib
18b 18ba j I_;iga
wb 18a 18a 18a iTala
18a ISa188'
1720 20 20m2O--20 20- 20
-.23
~~~~~24
23 ---23 ;-23 23 23_1 -0-23 23 23 23 23 23 2324_3
24. 24 244224 21 24 24 24
25 2 Oa~
.27
27,iv
?.2Mp7
27--- 2 v 27 2729*fl629 29 e _2g
32_32 32 32 _ 32 32 32 32 32 32
_
'333m34_
_ ; s 3;3 ;033
3-_
t~~~~~~~~~~~~~~~~~~35
_,35zsE % ~~~~~--n660U 3 6*# 3 36 36
@
_~7
iulwZ 37 43 37 37 37"_
9 339133139Si3939301P4W39
'39_'U39
39_I 40 40 40 40 40
_I_ L 4
~~~~~41-RW,41
41 +_ 41 41 41 41 41;2Sa4S3
~~Ua43W31m_
_ 43m43 43winj3
-v
44 44 4-
444
4a4E4
44e44_ 44e 4 44 44 4tsB5
DlEl
D3E2 tsE6A8E
D4E3
D1E4tsB5
D1E5
D4E1 D4E2
tsE6D5E1
D5E2FIG. 10. Autoradiographic imagesofelectrophoretically separatedpolypeptides labeled in cellsinfected
with series Dintertypic recombinantsfrom9to10hpostinfection.
402
on November 10, 2019 by guest
http://jvi.asm.org/
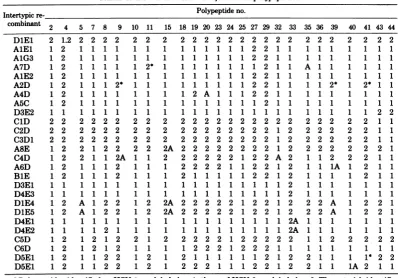
[image:14.509.81.454.51.629.2]VOL. 26, 1978 ANATOMY OF HSV DNA 403 TABLE 2. HSV-1- and HSV-2-infectedcellpolypeptidesa
Intertypicre- Polypeptideno.
combinant 2 4 5 7 8 9 10 11 15 18 19 20 23 24 25 27 29 32 33 35 36 39 40 41 43 44 DlEl 2 1.2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2
AlEl 1 2 1 1 1 1 11 1 1 1 1 1 1 1 2 2 1 1 1 11 1 1 1 1
A1G3 1 2 1 1 1 1 1 1 1 11 1 11 2 2 111 1 11 1 1 1 1
A7D 1 2 1 1 1 1 1 2* 1 1 1 1 1 1 1 1 2 1 1 A 1 1 1 1 1 1
A1E2 1 2 1 1 1 1 11 1 1 1 1 1 1 1 2 2 1 1 1 11 1 1 1 1
A2D 1 2 1 1 1 2* 111 1 1 1 1 1 1 2 2 1 1 1 1 2* 1 2* 1 1
A4D 1 2 1 1 1 1 11 1 1 2 A 1 1 1 2 2 1 1 111 1 1 1 1
A5C 1 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 2 1 1 111 1 1 1 1
D3E2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 2 2
ClD 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 1 1
C2D 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 1 2 2 2 2 2 2 1 1
C3D1 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 1 2 2 2 2 2 2 1 1
A8E 1 2 2 1 2 2 2 2 2A 2 2 2 2 2 2 2 2 1 2 2 2 2 2 2 2 1
C4D 1 2 2 1 1 2A 1 1 2 2 2 2 2 2 1 2 2 A 2 1 1 2 2 2 1 1
A6D 1 2 1 1 1 2 1 1 1 222211 2 1 2 1 1 1A 1 2 1 1
BlE 1 2 1112 1 1 1 2 1 1 1 1 1 2 2 1 2 1 1 1 1 2 1 1
D3E1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 2 1 1 1 1 1 1 1
D4E3 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 2 1 1 1 1 1 1 1
DlE4 1 2 A 1 2 2 1 2 2A 2 2 2 2 2 1 2 2 1 2 2 2 A 1 2 2 1
DiE5 1 2 A 1 2 2 1 2 2A 2 2 2 2 2 1 2 2 1 2 2 2 A 1 2 2 1
D4E1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 2A 1 1 1 1 1 1 1
D4E2 1 1 1 1 2 1 1 1 1 1 1 1 1 1 1 1 1 1 2A 1 1 1 1 1 1 1
C5D 1 2 1 2 1 2 2 1 2 2 2 2 2 1 2 2 2 2 2 1 1 2 2 2 2 2
C6D 1 2 12 1 2 1 1 1 1 2 2 2 1 2 2 2 1 1 1 11 1 1 1 1
D5E1 1 2 1 1 2 2 1 2 1 2 1 1 1 1 1 1 2 1 2 2 1 1 1 1* 2 2
D5E1 1 2 1 1 2 2 1 2 1 2 2 2 1 1 1 2 2 1 2 2 1 1 1A 2 1 1
aPolypeptides identifiedasHSV-1 arelabeledas1;those of HSV-2arelabeled as 2. The asterisk identifies
the few instances in which the polypeptides produced by thecells infected with the recombinants did not
cosegregatewith theDNA sequencespredicted byotherrecombinants.TheletterAdenotespolypeptideswith
anaberrantelectrophoreticmobility.
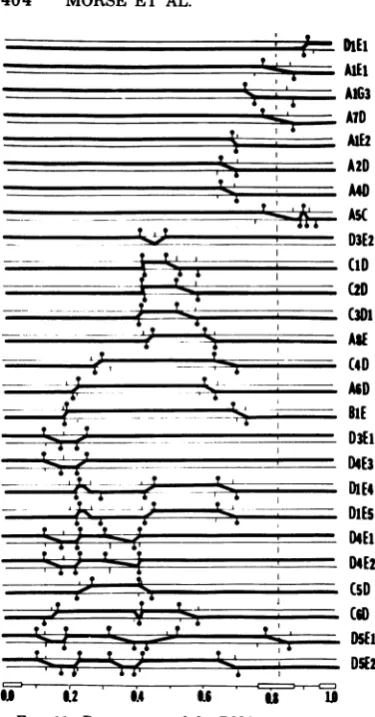
tothe leftof theleftcrossoversiteofC4D(0.30 a few instances inwhichthe
polypeptides
pro-mapunit) buttothe right of theleft crossover duced by the cells infected with the
recombi-site ofA6D (0.22 mapunit). The map position nants did not cosegregate with the DNA
se-oftheICP5template ismoreprecisely defined quences
predicted by
other recombinants.Pos-by the crucial recombinants D1E4 and
DiE5,
sibleexplanations
forthese inconsistencies are:each of which
specifies
anICP5withanelectro- (i) intragenic recombinationevents resultinginphoreticmobility intermediatebetween those of a recombinant gene
specifying
a polypeptideHSV-1 andHSV-2. Thissuggeststhepossibility with an electrophoretic
mobility
characteristicofan
intragenic
recombinationevent. Compari- ofoneof theparental
types, as illustrateddia-son with the ICP 5
specified
by D4E1, D4E3,grammatically
inFig. 13A; (ii)
DNAheterodu-andC5D indicates that the left
boundary
ofthe plex mismatchrepair;
and(iii)
undetecteddou-template
for ICP 5does notextend tothe left ble-crossovereventsinregions
of the DNAlack-beyond
0.23mapunit,
which indicates thatit ising
restrictionenzymesites. We notedpreviously
theright
crossoversite ofD1E4 and DlE5 that(26)
that double-crossovereventsencompassing
ismost
likely
responsible
fortheaberrant elec- smallregions
of the DNA would be detectedtrophoretic properties
of ICP 5 produced byonly
ifthey
bracketed a restriction enzymethese recombinants. The map
position
of thecleavage
site. The secondset, identified with thetemplate forICP 5 was thus determined to be letterA,denotespolypeptideswithan aberrant
between 0.23 and 0.30 map units. The DNA
electrophoretic
mobility.
Insomeinstances(e.g.,
sequencearrangements andtheICP5specified ICP 5 of D1E4 and
DlE5),
theelectrophoretic
by all other recombinants are consistent with
mobility
was intermediate between that of thethisconclusion. parental types. In other
instances,
thepoly-The segregation patterns of thepolypeptides peptidemigratedfaster than thecorresponding
mappedin thisstudy aresummarizedin Table parental
polypeptides.
Possibleexplanationsfor2, which identifies two sets of inconsistencies. the emergence of
polypeptides
with aberrantThefirst, identified withanasterisk, comprises
electrophoretic mobility
are:(i)
arecombinationon November 10, 2019 by guest
http://jvi.asm.org/
404 MORSE ET AL. J. VIROL.
:E.
DiE
matically
inFig.
13A and C maynothave been_ AlEl processedproperly.
__________________________ A From
Fig.
11and12,
itbecomesapparentthata A somepolypeptides are located in cluster groups
_______AID
A7D withoverlapping
boundaries. Thesepolypep-A=E2
tides can,however,
be orderedby analysis
of- A
^2D
theirsegregation pattern
among therecombi-- D~ nants. For
example,
ICP 10 must lie to the left____________, ,,,of the ICP 7 and 25, since recombinant C6D
--
St specifies HSV-1 ICP 10 but HSV-2 ICP 7 and- _____________ D3E2 25.Accordingly, a correlation of this segregation
(D1
pattern with the DNA map of C6DplacesICPC2D
10totheleft ofICP 7 and 25. The linear order_____,*L-< C3DI ofthe polypeptides deduced in this fashion
is,
301
**;from leftto
right,
9, 41, 18, 33(15, 39),40, 5, 24,'3
Al[
36(35, 11),8, 43, 44, 32, 2, 10(7, 25),23, 20,27,C(4
29,
4.-LP--
9ZASD
The map positions of severalpolypeptidesare__ ____ of
special
interest andaredealt with below.ICP4.Thesegregation
patterns
ofICP4(Fig._
-~D3EI
2,3, 5, 6,8,9, 11, and 12)indicate
that thegene__ _____________ D4E3 forthat polypeptide is located to the right of
=~
D1E4
0.787 map unitasdefinedby
recombinantsAIE1,
DIES
AI63,
A7D,
andDIEL.
Several observationsU64 made in this and otherstudies define the
loca-4D4E
tion of thistemplate
moreprecisely.
Specificaly:- f D4E2 (i) Recombinant DIE1 specifies in infected
(sD
cells(Fig.
8 and9)
thefully processed
fonn of,t_____________t_____.
_________ HSV-2ICP
4(ICP
4e)
andtheunderprocessed
7
7t(tD
forms
ofHSV-1 ICP 4(ICP 4a).
Theonly
de-==
OSEl
tectedHSV-1sequencesinDIE1arelocatedatc
==:~
_0~ :--n%D5E2
map positions 0.93 to 1.00, indicating that thetemplate
forHSV-1ICP4is in theScomponent
0 12 0 is and,
by
extension,
that thetemplate
for HSV-2FIG. 11. Provenance of the DNA sequences in the ICP4mustalso be in the S component. It would
intertypic recombinantsused in thisstudy.Theupper follow, therefore, that in DIE1 HSV-2 ICP 4
andlowerlinesof each doubletrepresent HSV-1 and maps between 0.83 and 0.93, whereas HSV-1
HSV-2DNA, respectively. Theheavy line identifies
ICP
4mapsbetween 0.93 and 1.0. In Fig.12,
the the DNA sequences presentintherecombinant virus. * * H *Thediagonallinespanstheboundariesofthecross- map
position
ofHSV-1 LOP4isshown
asdeter-over sites asdefined bythefive restrictionendonucle-
mined
from analyses of recombinants DIEl1 ases.Hatch marks at or near thediagonallineiden- There is evidence,however, that HSV-1 DNA is tifycriticalendonuclease cleavage sites thatdefinediploid
for theICP4gene and thatDIE1isonethe boundaries of the crossover event. The hatch
example
ofanICP4heterodiploid
recombinant;marks with aknob identify the restriction sitesre- this conclusion emerges from the observation
tainedinrecombinantDNA,whereasthose without thatrecombinants formed between intact
HSV-knobsrepresent parentalcleavagesites thatareab- 1 DNA and Xba-1 fragment I of HSV-2 DNA sent in therecombinant DNA. The boxes on themap also specify both HSV-1 and HSV-2
ICP
4. Inunitlinerepresent thereiteratedsequence ofLand theserecombinants the HSV-2 sequencesarein
Scomponents of HSV
DNA.teercmmnsteHV2sqecsaei
*
componentsofHSVDNA. mappositions0.945 to1.000;i.e., theyare
recip-rocal to those of the DIE1 recombinant (D.
eventmayhaveoccurred within a gene, resulting Knipe,W. Ruyechen, B.Roizman,and I.
Halli-in premature termination of transcription or burton,submitted forpublication).
translation(Fig.13B);(ii)arecombinationevent (ii) A possiblecomplicationthat might affect
mayhave occurred within ageneonly partially the mappositions of the genesspecifyingICP4
aligned,
resulting
in a recombinant gene speci- stemsfrom the observation thatalthoughviralfying
apolypeptide
withanapparentmolecular RNA extracted frompolyribosomes primedforweight
intermediate
betweenthat of the paren- synthesis ofapolypeptideshybridizestorestric-tal types (Fig. 13C); or (iii) the product of a tion endonuclease fragments of the S
compo-recombinant gene such as
illustrated
diagram- nent, the amount of the reiterated sequenceson November 10, 2019 by guest
http://jvi.asm.org/
[image:16.509.70.258.56.415.2]VOL. 26,1978 ANATOMY OF HSV DNA 405
Component . -l
s-
s->
Sequenceurnumgmuns.t US UL US C
Hybridintion
ofaRNA TOrigin
P~pelde
ofdefetive DNA
4Il I|mber
I
a: . I
II
41
JJ3I
2MFrIG.n funLcations of tepae spe TKnHS makes fucin,adplppidsdtrierma
analysisofHSV-I44V-2
royepresentedbin a'i RN wa to smll by a
I20
factr1
221
~
~
~
7Fr(tionowever,Fratioal
itoesumofthesequ
estnce0
0.1 0.2 s0.30.4
i 44e0.5 0.6 19I0.7 0.8 0.9 1.0FIG. 12. Locationof templatesspecifyingHSVmarkers,futnctions, and
polypeptides
determinedfr-omananalysis ofHSV-1 xHSV-2 recombinants.
represented
in this RNA was toosmall,
by
a Afactor of
2,
to contain the entireICP4template
Iv-1
(19).
However,
thesum of thesequencesinthe Nreiterated and unique sequences that are
pre-sented inthe a RNA could contain the ICP 4 flV-2
template. Additional evidence derivesfrom
stud-ies (5, 6)on defective virus generatedby serial B
passage of HSV-1 (Justin). Cells infected with l
certain populations containing defective
parti-cles
overproduce
ICP4.The defectiveDNA wasshowntoconsist ofareiteratedsegmentarising
from the ac and
Us
sequences of S located Cbetween map positions 0.939 and 1.000 in the
arrangement ofHSV DNA shown in Fig. 11(6).
Analyses of thesequences of the defective DNA L
that hybridize witha RNA (H. Locker andN.
Frenkel, in Proceedings ofthe Symposium on FIG. 13.
Hypothetical
models of recombinationalHerpesvirus and Oncogenesis, in press) also eventsgivingrise to inconsistent or aberrant
polypep-show that the amounts of reiterated ac se- tidesspecifiedbysomeofthe HSV-1 xHSV-2
recom-quences of the S component represented in a binants. (A) Intragenic recombinational event
be-RNA are not sufficient to specifyICP 4.
Thus,
tweengenes in whichonly the5'
endsare aligned.(B)although the evidence clearly indicates that
Intragenic
recombinational eventresulting
in apre-least partially diploid for ..OP.4 ge, mature termination oftranscription ortranslation.
HSaV
iSatleastpartially
diploid
for ICP4gene,HSV s at .
(C)
Intragenic recombinational
eventbetween geneswe cannot at this time determine whzether the only partially aligned resulting in a recombinant
putative
remainder ofICP 4gene extends into genespecifyingpolypeptideswith an apparentmolec-theuniquesequencesof Sorinto the Lcompo- ularweightintermediatebetweenthose specifiedby
nent. HSV-1 andHSV-2parents. The heavy line represents
ICP
27.BecauseA7D, A5C,
D5E1, D4E1,and thecorresponding
polypeptide template. The dashedD4E2allspecifiedICP27ofHSV-1 and because linerepresents the DNA sequences in the
recombi-the right crossover site in A7D and A5C near nant.
theL-Sjunctionhad to be to theleftof 0.84 map
unitasdeterminedbyKpn Idigestion(26), the the
right-hand
end of the Lcomponent betweentemplate specifyingthispolypeptide mustmap 0.787 and 0.83 map unit. In support of this
within the Lcomponent of the DNA. By similar location for the
template
specifying IOP
27 isreasoning, analyses of recombinants AlEl, theobservationthatthe
parental
strainsusedtoA1G3,and A2Dsuggest that ICP 27 maps within construct the recombinants in series Awere a
on November 10, 2019 by guest
http://jvi.asm.org/
[image:17.509.56.444.57.262.2] [image:17.509.276.423.297.455.2]406 MORSE ET AL. J. VIROL.
PAA' mutant of 17 tsJ (HSV-1) and wild-type pcU
HSV-2 (GP6) and that the selection was for Recombinant Boundaries 27 33.5
PAArts+progeny. From thoseexperiments, we L S
previously concluded that tsJ maps to the right A HSV-1 HoI EcoRI
ofmapposition0.78 (26). Marker rescue exper- 3 1.32
iments(Knipe, Ruyechan, and Roizman, unpub- SV;2.
lished data) showed that the
temperature-sen-sitive lesion of tsJ is located between mapposi- L2
tions0.70and0.83and therefore that the selec- B . 1.0
tion is consistent with a requirement for replace- -_
_________2
1.0ment of HSV-1 sequences with HSV-2
se-quences.
Threecommentsconcerning ICP27shouldbe C 12
made. First, ICP 27 is an a polypeptide. It is.1 (0
thereforeof interest to note that the DNA se- - -s --- - .
quencesarising from the region containing the
template forICP 27 are represented in a RNA 0oy"7 o08
of
(Fig. 12). Second,ourdataplace ICP27within
theregions of the reiteratedsequenceab ofthe
map
unitsLcomponent. However, recombinants contain- FIG. 14. Possible location of crossover sites near
ingheterologous ab sequences (e.g., A4D,
A1G3,
thejunctionsof L and Scomponentsin theDNAsofA2D, A1E2, and D5E2) do not produce both recombinants
AlEl,
A5C, and A7D. (A) Thebound-HSV-1 and HSV-2 ICP 27.Third,ICP 27 can be aries of the crossover sitesbased on restriction
en-used to better define the position ofcrossover donucleasemapsoftherecombinant
DNAs.
Asindi-catedin the text, andpreviously reported, the
recom-events in the DNA
ot
severalrecombinants.
binational event occurredbetween theHpa
I cleavageThus, because of the absence of known restric- site inHSV-1 and the Kpn I site in HSV-2. (B) The
tion endonuclease cleavage sites between map probable location ofthe crossover site inrecombinant
positions0.78and 0.83 for HSV-1 and 0.76 and AlEl defined by the electrophoretic mobility of ICP
0.83 for HSV-2, the crossover events near the 27 andefficiency ofplating. (C) Theprobablelocation
junctionof the L andScomponents inthe DNA ofthe crossoversite inrecombinants A5C andA7D
of recombinants D5E1, A5C, A1G3, AlEl, and based onthe electrophoretic mobility of ICP27and
A7D are shown as occurring within a broad partialtemperature sensitivity of the recombinants.
regionsanningthejunction.Comp s of In(A)and(B),the
heavy
linerepresentsthetemplateregion
spannmng
thejunction. Comparisons
orfo IC 7 h ahdlnerpeet h Nthe DNA sequence arrangements, polypeptides, for ICP 27.
The dashed line
represents the DNAandbiological propertiesof recombinants shown sequencesin the recombinant.
in Fig. 14 suggest that in AlEl the crossover
event is nearer the map position 0.78 than in Mapping of specific viral functions. (i)
A5C and in A7D. The conclusionis based on the HSV-1
(HFEM)
yn- locus. The biologicalobservation that AlEl
specifies
the ICP 27 of properties of the intertypic recombinants wereHSV-2,whereasthe othersspecify the ICP27of
reported previously (26).
In that report it wasHSV-1 andare
partially
temperature sensitive. observed thatintertypic
recombinants (C3D2,ICP6. Wesuspectthat the broad band des-
C4D, C5D, C6D,
andC7D) expressed thesyn-ignatedasICP6andICP7containsatleasttwo
plaque morphology phenotype
characterizedbypolypeptides. In earlier studies (29), it was fusion of cells intopolykaryocytes. Inasmuch as
shown thatasingle bandappears inthatregion series Crecombinants werederivedfromacross
during a
pulse
andthat onlya portion of that ofHSV-1(HFEM
tsN102 syn-) x HSV-2 (186band is then chased to a higher molecular syn+), those recombinants expressing the
syn-weight.Current data indicate that early ininfec-
phenotype
mostlikely
contain HFEM DNAse-tion, thepulse-labeled bandsimplybroadens to quences
specifying
thisfunction. Analysisofre-a higher apparent molecular weight during the combinant DNAs shows that the template
re-chase (Fig. 1). WehavetentativelydefinedICP
sponsible
for thesyn-phenotype
mustmap to6 as that
polypeptide
which shifts to a higher theright
oftheleftcrossoversite ofC4D (0.30)apparent molecular weight and ICP 7 as that andtothe left of therightcrossoversite ofC5D
polypeptidewhich does not becomeappreciably (0.42).
alteredduringthe chase. Wehave not been able
PAAr.
PAA has been shown toselectively
to map ICP 6 because theelectrophoretic mo- inhibit the HSV DNA
polymerase (23, 27).
Re-bility ofHSV-1 ICP 6 is not different enough sistancetoPAAwasused for selection of
recom-fromthat of HSV-2 ICP 6 to obtain a reliable binantprogeny in seriesA,B,and C.
Previously
segregationpattern. we
reported
that the PAA resistance markeron November 10, 2019 by guest
http://jvi.asm.org/
[image:18.509.275.461.58.258.2]VOL. 26, 1978 ANATOMY OF HSV DNA 407
mapsbetween0.43and0.52mapunit basedon be 44,000andbyCourtneyetal. (4) to be 43,000.
DNAsequence arrangementsofClD, C2D, and ICP 35 (molecular weight 42,000) mapped
be-C3D1. tween 0.27and0.35 map unit, whereas ICP36,
Shutoff of host protein synthesis. HSV-1 another candidate with a molecular weight of
and HSV-2shut off hostprotein synthesis within 47,000, mappedbetween 0.31 and 0.38 map unit.
a few hours after infection. Pereira et al. (29) Theseresults are also consistent with a recent
and Powell andCourtney(30) noted that HSV- reportthat theHpa I DNA fragment contained
2 shuts off host protein synthesis much more the geneticinformation necessary to
biochemi-rapidly and efficiently than HSV-1. We have cally transform tk- cells to the tk+ phenotype
used this differentialproperty to mapthe func- (41;S. Silverstein, personal communication).
tionresponsible for the accelerated inhibition of
hostprotein synthesis. DISCUSSION
Analysis of HSV-infected HEp-2 cells labeled In this paper we have (i) enumerated and
from 1 to 3 h postinfection (Fig. 2, 5, and 8) provisionally matched HSV-1 and HSV-2
poly-clearly demonstrated that five recombinants
peptides
with molecular weights of 20,000 to(C2D,C3D1, DlEl, C5D, and C6D) rapidly and more than 200,000, (ii) identified polypeptides
efficiently shut off host protein synthesis in a that vary in apparent molecular weight from
mannercharacteristic of HSV-2. Correlation of strain to strain and those that undergo rapid
thisphenotype with the
physical
DNA mapof post-translationalprocessing,resulting
in anap-the recombinants suggests that function in- preciable change in the apparent molecular
volvedinthe accelerated inhibitionofhostpro- weight, and (iii) mapped the approximate
loca-tein
synthesis
mustmapbetween0.52 and 0.59 tionof thetemplates
specifying
26polypeptidesmapunit. Itmaybe
significant
that thispheno-
and several viral functions. Several aspects oftypecorrelated with thepresenceof HSV-2ICP thepresentdata meritdiscussion.
7and27.Contrarytoexpectations, ClDdid not limitations of the
mapping
procedures.exhibit this phenotype,
possibly
because the The HSV-1 and HSV-2 genes that wereame-gene functionwas inactivated
by
the recombi- nabletomapping by the procedure used in thisnationeventin thatregion. Sincewe weremap- paper arethose thatdiffer in theapparent
mo-ping accelerated inhibition of host protein syn- lecularweight oftheirproduct. Thegenes whose
thesis, these observations should not be con- products do not differ in apparent molecular
struedtoindicate that the number and locations
weight
orinimmunologicalspecificity
will haveof all genes responsible for inhibition of host tobemapped by other
procedures.
functionsareknown. The
precision
of themapping procedure
usedThymidine kinase. Several linesofevidence in this study is
governed
by two factors: thesuggestthatthe gene
specifying
the thymidine number and distribution ofcrossoversites andkinasemapsbetween0.27and0.35mapunit. the numberofrestrictionenzyme cleavage sites
(i) HSV-1 x HSV-2 intertypic recombinants available topinpoint the exact location of
cross-were constructed from acrossbetween HSV-1 over sites. There were no crossover sites between
(B2006
tk-)
xHSV-2(GP6PAAr). Progeny were map positions 0 and 10, and only onerecombi-selected that were PAAr tk- by plating in the nant contained a crossover site in the S
compo-presenceof100 ,ugof PAA(17, 23) and 100
,ug
of nent suitable for mappingpurposes. In addition,thymidine arabinoside (ara-T) (7) permlof me- the exact position of crossover sites was
deter-dium. The only recombinant obtained in this mined (26) by a total of 56 restriction enzyme
crosscontained HSV-2sequencesbetweenmap cleavage sites in HSV-1 DNA and 40 sites in
positions 0.38 and 0.44, suggesting that the tk HSV-2 DNA. In consequence, no polypeptides
gene maps totheleftof 0.38 map unit or tothe were mapped in mappositions 0 to 10, only one
right of0.44mapunit. polypeptide wasunambiguously shown to arise
(ii) Theparental HSV-1 mutant (tsE6) used from a template in the S component, and, as
in the construction of the Dseries of recombi- shown inFig. 11, the boundaries for some ofthe
nantsistk- (4). Wehaveanalyzed the abilityof polypeptide locationsare quite broad and may
recombinantsD1E4,
DiE5,
D5E1,and D5E2 to have to beredefined byothertechniques.form plaques in the presence of 20
,ug
of ara-T Considering thecomplexity
of the genome,per ml of medium. Only D5E1 and D5E2 were the
correspondence
between thesegregation
abletoplaqueinthepresence of ara-T and were patternsof the DNA sequences and that of the
therefore tk-. This suggests that the tk- gene
polypeptides specified
by
the recombinantswasmaps between 0.27 and 0.35 map unit. quite
good.
The number ofinconsistencies was(iii) The molecular weight of the
HSV-1-in-
small(Table
2) and could be ascribedtoone orducedthymidinekinasepolypeptide in SDS-gels moreofseveral
possibilities
asdescribed earlierwas
determined
by Honess and Watson (16) to inthetext.on November 10, 2019 by guest
http://jvi.asm.org/
408 MORSE ET AL. J. VIROL.
Apparent gene arrangement in HSV double-strandedRNA. Wehaveinfactreported
DNA. Several aspects of the arrangement of that nuclear transcripts are many times larger
HSVgenetemplatesareofinterest.First, there than
polyribosomal
viral RNA(18,
40) and thatappears to be a clearsegregation of templates avery
large
fraction of the DNA is transcribedspecifying a polypeptides. Thus both a tem-
symmetrically (17, 22).
platesmap atthe terminiof L andScomponents Presence ofgene
templates
inreiteratedatleastpartly within the reiterated regions. The sequencesof viral DNA. As indicated in
Re-position ofa templates coincides with themap sults, ICP27maps
entirely
within the abregion
position ofsome aRNAsequences asreported of theDNA, whereas ICP4mapsatleast inpart
previously. Theproximity ofatemplatestothe in the reiteratedacsequencesof the S
compo-termini of theLandScomponents suggeststhat nent
(Fig. 12).
These observationswould
indi-the promoterfortranscriptionofthesefunctions catethatHSV isdiploid for ICP27andatleast
maybeatthe termini.Second, althoughno
fl
or partially diploid for ICP 4. However, whereasy polypeptides were mapped in the S compo- ICP4heterodiploidywasreadily demonstrated
nent,thoseintheLcomponent appeared to be inrecombinants withheterotypic acsequences
intermixed. It is also interesting to note that (e.g.,DIE1), itwas notdemonstrable for ICP27
viral functionsmostlikely involving
,8
polypep- inrecombinants withheterotypic
absequencestides (e.g.,tk, PAAr, and accelerated shutoffof (AIG3, AIE2,A2D, A4D, and
D5E2).
Twopos-hostmacromolecularsynthesis)wereintermixed sible
explanations
may accountfor thisobser-withyfunctions(syncytial plaque morphology). vation. First, it is conceivable that an ICP 27
These observations have several implications. template is located in the left end of the L
The apparent random distribution of tem- component,butthat it lacksapromotersite and
plates of
/8
andypolypeptides
is ofspecial inter- is nottranscribed.Itshould be noted that if theestin view ofthe evidence
indicating
that the promoterfortranscription
ofICP 27wereinS,
sequential switch-on of the
coordinately
regu- asconsideredinthepreceding
section,
thenbothlated ,B and y
polypeptide
syntheses is deter- theright and lefttemplates
ofICP27would beminedatthe
transcriptional
andpost-transcrip- transcribed off the P and ILarrangements
oftional levels (14, 15,32,34). Onepossible expla- HSV DNA, respectively, and both HSV-1 and
nation for the data is that
,B
and ytemplatesare HSV-2 ICP 27wouldbe made.The secondhy-segregatedonopposite strands.Toillustrate this pothesisis thatatleast
portions
of the terminalhypothesis further, it could be that (i) thepro- inverted sequences must be identical but
in-moter for a template transcription is in the verted and that the ICP 27 gene lies entirely
reiterated sequences of the S component and within suchsequences, whereastheICP4gene
allows
transcription
fromright
toleftacrossthe does not. Thishypothesis arises from the notion,function, (ii)
the promoter for,8 polypeptides
in tobedetailed elsewhere(manuscript
inprepa-the L component istothe left ofmapposition ration), that the inversion of L and S
compo-0.78andallowstranscriptiontotheleft, and (iii) nents relative to each other arises as a
conse-atleastone promoter fory
templates
isto the quenceofanobligatory
post-synthesis eventinleftofmapposition0.12andpromotestranscrip- which the terminal ab and ac sequences are
tion tothe
right.
In support ofthis hypothesis repaired or regenerated, using the reiterated are thefollowing
data.(i)
Trace amounts ofa baacjunction as a template. To evaluate this RNAhybridize
tosequencesoccupied by,8
and hypothesis, wereexamined the data indicating ytemplates
(19). Consistentwiththisfinding,
in thattherecombinantsA1G3,A1E2,
A2D, A4D,manyexperimentsinwhichattempts weremade and D5E2 have heterotypic ab regions. The
torestrict viral
protein
synthesistoapolypep-
conclusionsarebasedonanalyses ofthe DNAstides,
traceamountsofasubsetof/8
polypeptides
ofrecombinantviruses withthe restrictionen-designated as /3' (ICP 6, 8, 17, 29, 39, and 41) donuclease Hpa I which cleaves HSV-1 DNA
were also made (29; Fig. 2, 5, and 8).
Normal
within the ab sequences at mappositions0.037synthesisof
/31
polypeptides, however, required and 0.787. The ICP 27 template was mappedtranscription in the presence of functional /3 betweenmap positions 0.787 and 0.83. Since we
polypeptides (29). One possible explanation is have norestriction enzymecleavagesites in the
that the trace amounts of/3' polypeptides and ab sequences between 0.000 and 0.037 and
be-corresponding RNA represent read-through tween 0.787 and 0.83, we cannot exclude the
from right to left throughtheputative
/3,
pro- possibility that these regions are infact
homo-moter.(ii)Thehypothesispredictsnuclear tran- typic, andconsequently onlyoneICP 27 would
scripts considerably greater inlength than the be expected.
polyribosomal mRNA as
well
asthe accumula- Contribution of the recombinants to thetion oflargeamountsof RNA transcribed from study of the evolutionary divergence of
both strands andcapable ofannealingtoform HSV-1 and HSV-2. Numerous studies have
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 26, 1978 ANATOMY OF HSV DNA 409
shown the close genetic relationship between Proceedingsof the Symposium on Herpesvirusesand
HSV-1 and HSV-2, and it has been suggested Oncogenesis.I.A.R.C., Lyon.
that their current evolutionary divergence re- 4. Courtney,R. J., P. A. Schaffer, and K.Powell. 1976.
flectsacquisitionofaffinityfor differentecolog- Synthesisof virusspecific polypeptides by temperature-sensitivemutantsofherpes simplex type 1.
Virology
ical nichesas aconsequence ofdifferent modes 75:306-318.
oftransmission (31).Implicitinthis hypothesis 5. Frenkel,N., R. J.Jacob, R. W. Honess, G. S.
Hay-is the heurHay-isticconceptthat the viruscarriestwo ward, H. Locker,and B. Roizman. 1975.Anatomyof
offunctions: those that pertaintothe inter- herpessimplexvirus DNA.III.Characterizationof
de-setsottunctlons: t lose t zat pertam to t le mter- fective DNA molecules and biological properties of
action of viral macromolecules among them- virus populations containingthem. J. Virol. 16:153-167.
selves and those that involve interaction of viral 6. Frenkel, N., H.Locker,W.Batterson, G. Hayward,
and hostmacromolecules (31). Analyses of the and B. Roizman. 1976. Anatomy of herpes simplex
DNAsof therecombinantvirusesreportedpre- DNA. VI. Defective DNAoriginatesfrom the S
com-ponent. J. Virol.20:527-531.
viously (26) led to the conclusion that HSV-1 7. Gentry, G. A., and J. A. Aswell. 1975. Inhibition of
and HSV-2 DNAsare atleastgrossly colinear, herpes simplex virusreplication by Ara-T. Virology
i.e., that the arrangement of the genes was not 65:294-296.
alere
drat
the courseof the.evolution
8. Gibson, W.,and B. Roizman. 1972.Proteins
specifiedaltered
drastically
inthecourseoftheevolution- by herpessimplex
virus. VIII. Characterization andarydivergence. This study contributedanaddi- compositionof multiple capsid forms ofsubtypes1 and
tional conclusion based on the finding that at 2. J. Virol.10:1044-1052.
leastsomeof therecombinantsspecifyanearly 9. Grodzicker, T., J.Williams, P.Sharp,.and J.
Sam-equal
number of HSV-1 and HSV-2 polypep- brook. 1974. Physical mapping oftemperature-sensi-tive mutations ofadenoviruses. Cold Spring HArbor
tides,implyingthatHSV-1andHSV-2 polypep- Symp. Quant. Biol. 39:430-446.
tides arecapable of interacting. Althoughnot all 10.Hayward,G.S.,R.J.Jacob, S. C. Wadsworth, and
permutations have been observed, and therefore B. Roizman. 1975. Anatomy of herpes simplexvirus
wedo notknowwhetherprohibited, i.e.,lethal, DNA: evidence for fourpopulationsofmolecules that
wedonotknoweterproibte,.differin therelative orientationsoftheirlongandshort
genecombinations exist,the dataare adequate segments. Proc. Natl. Acad.Sci.U.S.A.72:4243-4247.
toconclude that the sitesonviralpolypeptides 11. Heine, J. W., R. W.Honess, E. Cassai,and B.
Roiz-specifically
definingtheinteractionof viralmac- man.1974. Proteins specified by herpessimplex virus.romoleculesromoleculesamongamongthemselves have beencon- XII. The virionpolypeptides of type1strains.J.Virol.
~~~~~~~~~14:640-651.
served. The conclusions are consistent with 12.
Hilliker,g;.,
and D. Botstein. 1976. Specificity of geneticthose ofHillikerand Botstein (12) fromstudies elements controlling regulation ofearly functions in
on bacteriophage lambda that evolution pro- temperate bacteriophages. J. Mol. Biol. 106:537-566.
ceededsuch that sequence
divergence
occurred13-
Honess,R. W., and B.Roizman.1973.Proteinsspecifiedby herpes simplexvirus.XI.Identificationandrelative
between recombinable segments. molar rates of synthesis of structural andnon-structural
herpesvirus polypeptides in infected cells. J. Virol.
ACKNOWLEDIGMENTS 12:1346-1365.
WegratefulyacknowledgethehelpofIanHalliburtonin 14. Honess, R.W.,and B. Roizman. 1974.Regulation of
settingupthenewconditions forpolyacrylamide gelelectro- herpesvirusmacromolecularsynthesis.I.Cascade
reg-shorettingsup.the
new conditionsfor polyacrylamide geleectro ulationofthesynthesis
of threegroupsofviralproteins.phoresis. J. Virol. 14:8-19. I
The studiesconductedattheUniversity of Chicago were 15. Honess,R.
W.,
and B.Roizman. 1975. Regulation ofaidedbygrantsfromthe National Cancer Institute, Public hersu macromolecula
tion-Health Service (CA 08494 and CA 19264), the American
herpesvirus
macromolecularsynthesis: sequential tran-CancerSociety (VC103L), and theNationalScience Foun- sition ofpolypeptide synthesis requires functional vi-dation(PCM 76-06254). The studiesatSidney FarberCancer ral polypeptides. Proc. Natl. Acad. Sci. U.S.A. Center,HarvArdMedicalSchool,wereaided by grants from 16.72:276128.
theatinalCanernsttut, Pbli HelthSerice(CA
16-Honess,
R. W., and D. H. Watson. 1974. Absence of a the National Cancer Institute, Public Health Service (CA requirement forhost polypeptidesin the herpesvirus10893and CA 20260). L.S.M. is apredoctoral trainee sup-
requinement
J.Gen.poly
22th7
er185
uported by Public Health Service training grant 5-T32 17. tymidine kinase. J. Gen. Virol. 22:171-185.
GM07183 from the National Institute of General Medical 1-Jacquemont,B., and B. Roizman. 1975.RNAsynthesis
Sciences.L.P.isapostdoctoral trainee supportedbyPublic in cells infected with herpes simplex virus. X. Properties
Sciences.Servis a grantrAInee frte bytional of viral symmetric transcripts and ofdouble-stranded
Health Service training grant AI-00184 from the National RNApreparedfromthem.J. Virol.15:707-713. Institute ofAllergyand Infectious Diseases. 18.Jacquemont, B., and B. Roizman. 1975. Ribonucleic
LITERATURE CITED acid synthesis in cells infected with herpes simplex virus: characterization of viralhigh molecular weight 1. Cassai,E., R. Manservigi, A. Corallin,and M. Terni. nuclear RNA. J.Gen. Virol. 29:155-165. 1
1976. Plaque dissociation ofherpes simplex viruses: 19. Jones, P. C., G. S. Hayward, and B. Roizman. 1977. biochemical andbiological charactersof the viral var- Anatomy of herpes simplex virus DNA.VII. a RNA is
iants.Intervirology6:212-223. homologous tononcontinuous sites in both the L and S 2. Cassai,E.N., M.Sarmiento,and P. G. Spear. 1975. components of viral DNA. J. Virol.21:268-276.
Comparison of the virion proteins specified by HSV 20. Kit, S., and D. Dubbs. 1963. A non-functional thymidine types1and2.J.Virol.16:1327-1331. kinase cistron inbromodeoxyuridine resistant strains of 3. Courtney,R.J.,and K. L.Powell.1974.Immunological herpes simplex virus. Biochem. Biophys. Res. Commun.
and biochemical characterization ofpolypeptides in- 13:500-504.
ducedbyherpes simplexvirus types 1 and 2, p. 63-73. 21. Kieff, E. D.,S. L. Bachenheimer, and B.Roizman. InG. deThe,M. A.Epstein, and H. Zur Hausen (ed.), 1971.Size,composition, and structure of the DNA of