0022-538X/80/11-0627/05$02.00/0
Viruses Isolated from
Cells
Persistently Infected with
Vesicular Stomatitis Virus Show Altered Interactions with
Defective Interfering Particles
FRANK
M.HORODYSKIAND JOHN J. HOLLAND*Department of Biology, University ofCalifornia,San Diego, La Jolla,California92093
Virusmutantsisolated
from
persistent infections ofvesicular stomatitis virusin BHK-21 cells were much less susceptible to interference mediated by the
defective interfering particle used to establish the persistent infection. This
mutational change
occurred asearly
as 34days
in the persistent infection and continued forover 5 years.Theearliest variants showednooligonucleotide
mapchanges and no difference in the temperature-sensitive phenotype from the
original
virus, but the later variants exhibited extensive map changes. Theseresultssuggest apossiblerole for defectiveinterfering particles in the selection of
themutants.
A number of animal viruses which are
nor-mally cytocidal are capable of establishing a
long-term persistentinfection in culturedcells.
Although the precise mechanisms responsible
for the establishment and maintenance of
per-sistent infections of RNA viruses havenotbeen
clearly defined,
anumber offactors have been implicated: (i)temperature-sensitive
mutants(18, 27),
(ii)
interferonproduction (14,
15,19),
and
(iii)
defectiveinterfering (DI)particles (1,5, 10, 17, 20, 22,26).Each of these factorsmayplayagreater orlesser role
depending
onthe virus-cellsystemstudied.Huang and Baltimore
(7)
firstsuggested
that DIparticles might play
animportant
role in viralpersistence.
Thebest-characterized
persist-entinfection in which DI particles playamajor role is that of vesicular stomatitis virus
(VSV)
inBHK-21 cells (5, 6). DI
particles
were neces-sary for both the establishment andmainte-nanceof the
persistent
infection(5).
TheBHK-21CAR4
persistently
infectedcellline has been maintained for 6.5 yearsandcontinues toshed low levels of standard virus and aconstantly
changing population
of DIparticles.
Standard virus isolated from thepersistent
infection at various timeswasanalyzed by
T1 oligonucleotide
mapping
ofthe viralRNA,
and it was shown that thevirusshedby
thisculture isundergoing
extensiveand continuous mutational
change (4).
Mutationsareexhibitedinall five viralproteins andinthe endsoftheviralRNA
(21,
23).
The recovered virues exhibitasmall-plaque
pheno-type; they have become
increasingly
lesscyto-pathic
and canestablish apersistent
infection after69months withoutadded DIparticles (6,
21). Little is known about the selective
mecha-nisms involved inpromoting sucha rapid
mu-tationrate,and it isdifficulttodetermine which mutations havebiological importance since the mutational evolution of the virus is both
exten-sive and continuous.
Many investigators have obtained mutations inthe virusisolated from
persistent
infections in othersystems(1,12, 13, 18, 24,25).These viruses often exhibitasmall-plaqueortemperature-sen-sitive
phenotype.
Ofparticular
interestwasthefinding by Kawai and Matsumoto (9) that a
small-plaque
variant ofrabies virus isolated froma
persistent
infection was lesssusceptible
to interferenceby
theoriginal
DIparticle
used to establish this persistent infection. Theypro-posed
that thissmall-plaque
variant could beeffectively
selected since it is much lesssuscep-tibleto interference
by
thepreexisting
DIpar-ticles.
Wereporthereasimilar
finding
in thecaseofpersistent
infections ofVSV in BHK-21 cells andpresent a quantitative preliminary characteri-zation of these viruses.
The persistently infected
line
studied here(designatedCAR4)was
originally
establishedbycoinfection of BHK-21 cells with ahigh
multi-plicity
of the cloned tsG31 mutant of the VSV Indianaserotypeand itshomologousDIparticle (5). After establishment of thepersistentinfec-tion at
330C,
thecellsweremaintained at370C.
Virus was isolated at various
times
from thepersistently infected culture by cocultivationof carriercells and nornalBHK-21cells.Virus was
plaque purified,andhigh-titerclonalpoolswere
prepared forusein thesubsequent
experiments.
Wethenexaminedthe
susceptibility
ofthese virusestointerferencemediatedbythe original627
on November 10, 2019 by guest
http://jvi.asm.org/
628 NOTES
DI particle used toestablish this persistent in-fection (tsG31 DI). We also quantitated the abil-ity of the recovered virusestoreplicate this DI particle. Thiswasdonebycoinfecting identical
monolayers of BHK-21cellswith theinfectious
virus at a multiplicity of5 PFU/cell together
with increasing amounts of the purified tsG31 DI particle. Interference was quantitated by
plaqueassayof the yield. Replication of the DI
particle was examined after velocity gradient
centrifugation ofprogenyvirus onsucrose
gra-dientstoseparate standardvirus and DI parti-cles. Wequantitated theDIparticleyield
spec-trophotometricallyasdescribed in the legendto
Fig.1.Atypicalsucrosegradient profile is shown
in Fig. la. The resultsareshown inFig. lb and
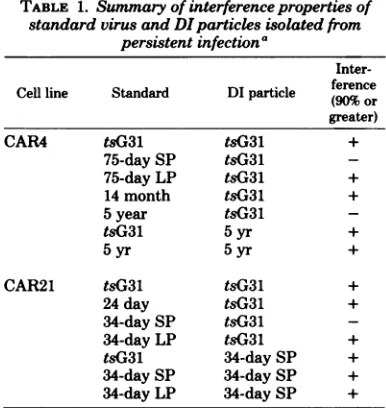
candsummarized in Table1.Themixed
popu-lation of virusisolated from the persistent infec-tionat 75days contained 70%smallplaques (1
to2mm). The remainder of the plaqueswereof
asize comparable to the tsG3M standard virus (4 mm). The small-plaque phenotypebredtrue,
and no detectable interference or DI particle replicationwasobservedevenatalevelofinput
DIparticles which givesover1 log interference with the homologous standard virus. The large-plaque virus at 75 days postinfection (75-day LP)wasstill subjecttotsG31 DI particle-medi-ated interference and still replicated that DI particle, althoughsomequantitative differences wereobserved. At 14months, the virus isolated
from thepersistent infectionwasoflarge-plaque phenotype andwasonceagain sensitivetotsG31 DI particle-mediated interference. In contrast, the small-plaque virus isolated at 5 years was
insensitive tothese levels of the tsG31 DI
par-ticle. For each virusexamined, the amount of interferenceobservedwasdirectlyrelatedtothe level of DIparticlereplication (compare Fig. lb andc). This correlationshowsthat eitherassay canbequantitatively usedtostudyDI
particle-mediatedinterference.
To determine how quickly this mutational changecan occur in the standard virus during
persistent infection,aseparatecarrier line (des-ignated CAR21)wasestablished in BHK-21cells
by the method by which the CAR4 persistent
infection had been established, and with the exception that these carrier cells were
main-tainedat33°Catalltimes (5). Viruswasisolated
by direct plaquepurification from the
superna-tantof thepersistentinfection. Figure ld ande
shows thatthis mutationwhichconfersa
rela-tive resistance to the original DI particle can
take place by 34 days,atwhichtime asimilar
mixed population of small-plaque (34-day SP)
andlarge-plaque (34-day LP) virus was found.
The34-daySP viruswasnotsensitiveto
inter-ference mediated by the tsG31 DI particles at
the levels of input DI particles employedin this
experiment. However, the virus isolated at 24
days in this persistent infection was entirely
largeplaqueandshowed normal levels of
inter-ferencebythe tsG31 DIparticle (Table 1). Wenextdeterminedwhetherextremelyhigh multiplicitiesofthe tsG31 DIparticlecould in-terfere well with the 34-day SP and 75-day SP viruses. Detectable levels of DI particle
replica-tion and interference did occur with these
vi-ruses at aninputof 10 DI units per culture, and greater than 90% interference was obtained
when 100 DI units of the tsG3M particle were
used. However, weobservedthat thetsG31 DI
particle always depressed theyield of homolo-gous tsG31 virusat least 10- to1,000-foldgreater
than it depressedtheyieldof34-daySP and
75-day SPviruses,evenunder the extreme interfer-enceconditions ofhigh inputDIparticles.
Sim-ilar results were seen if a low multiplicity of
infection (0.01 PFU/cell) of standard virus was
employed with a high inputoftsG31DI parti-cles,and the virusyieldwasassayed duringthe first cycle of infection (at 9 h postinfection). Clearly the mutant viruses are much more
re-sistant tothe
original
DIparticle,
but thisre-sistanceis
quantitative
andnotabsolute. Wenextcarriedoutreciprocal experimentstodetermine whether the originalstandard virus
(tsG31)
issubject
tointerferenceby
DIparticles
generated
by
the virus variants isolated after various periods of persistent infection. BHK-21 cellswerecoinfected withahigh multiplicity (5PFU/cell)
of standardvirusand5to 10DIunits of each DIparticlestudied. Table 1showsthatgreater than 90% interference occurred with the
original
tsG31 standard virusproduction
when5 to 10 DI units of either the 34-day SP DI
particle
orthe 5-year DIparticle
wasused. It is evident fromthese results that this mutational change is unidirectional. Standard virusunder-went amutationalchange which rendered it less sensitivetothe
original
DIparticle,
whereas DIparticles
generated
by these viruses remainedpotent interfering agents against the original
standardvirus.
Itwasofinteresttodetermine whether these mutations whichconferresistance to the tsG31 DIparticle alwayscause a detectableT1 oligo-nucleotide map change in viral RNA. Cloned viruswaslabeled with32Pandpurified. The viral RNAwasisolatedand digested with
T,
nuclease,and theresulting oligonucleotideswere mapped ontwo-dimensionalgels as previously described
(4). The oligonucleotide mapsof the large- and
small-plaque viruses isolated at 75 days from
CAR4 and34daysfromCAR21areidenticalto
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
2 DI STD a
0
TOP BOTTOM
b
O.OI Li2348 55
4
:8' ~~~~~~~~~~~~3
DI UNITS/CULTURE DI UNITS/CULTURE
>
d
ine
>0.1 cm
2
0.01'~~~~~~~~~~~~~~~~~~~
I 2 3 4 3 2 3 4 5
DI UNITS/CULTURE DIUNITS/CULTURE
FIG. 1. Interference and replication properties of thetsG31 DI particle when coinfected with standard viruses isolated from CAR4 (b and c) andCAR21(d and e). Matched monolayers of 4 x 107BHK-21 cells were coinfected with cloned standard virus at a multiplicity of infection of 5PFU/cell andtsG31 DI particles which were purified by two successive centrifugations in a 5 to 40%(wt/vol) sucrosegradient. Since these
virusesare temperature sensitive, allinfections and assayswere done at33°C. This DI particle purification procedure reduced the level of contaminating standardvirusto less than 1 particle per 108(3).Culture fluids wereharvested at 48 h postinfection and assayed for infectivity by plaque assay. Progeny virus was pelleted and centrifuged on a 5 to 40%(wt/vol) sucrose gradient (3). The gradients were fractionatedcontinuously
fromthe top through an ISCO model UA-5 absorbance monitor which measures absorbance at 254 nm, and the DI particle yields werequantitated by weighing the peaks. Atypicalsucrose gradient is shown in (a). V, PFU per milliliter after additionofvarious doses of DIparticles; VoPFUper milliliter without addition of DI particles. (b andc)@*,tsG31;E475-daySP;E475-day LP;*,14 month;0,55year.(d and
e)@*,
tsG31;4l
34-daySP;EN, 34-day LP.NOTES 629
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.499.98.390.67.536.2]630 NOTES
TABLE 1. Summary of interference properties of standard virus andDIparticles isolatedfrom
persistentinfectiona
Inter-Cell line Standard DIparticle (90%or
greater)
CAR4 tsG31 tsG31 +
75-day SP tsG31
75-day LP tsG31 +
14 month tsG31 +
5year tsG31
tsG31 5yr +
5yr 5yr +
CAR21 tsG31 tsG31 +
24day tsG31 +
34-day SP tsG31
34-day LP tsG31 +
tsG31 34-day SP +
34-day SP 34-day SP +
34-day LP 34-daySP +
aMonolayers of BHK-21
cells
werecoinfected withstandard virus at amultiplicity of infection of 5 PFU/ cell and 5 to 10 DI units ofpurified DI particles. One DI unit isdefined as the number of DI particles in the inoculum which gave 37% of theyield of DI particle-free inocula (i.e., which gave one hit biologically [2]).
the
original
tsG31virus(data
notshown).
It is not surprising that these mutations are unde-tectable byT,
oligonucleotide mapping
of the viral RNA, sinceonly about 10% ofallmutations result in alterations in the unique oligonucleo-tides.We next determined whether the tempera-ture-sensitive
phenotype
of the recoveredvi-ruseshad beenalteredduring thecourseof the persistent infection. The
yield
of these virusesgrown at33,37,and
390C
wasdetermined(Table
2). Allof the viruses recovered from the
persist-entinfectionwerestilltemperaturesensitiveas evidencedbyatleast a
2-log
reductioninyield
when the viruses were grown at 390C as com-pared with the yield when the viruses were grown at
330C.
There are undoubtedly many selective pres-sures exerted on the virus during a persistent
infection,and DIparticlesappear toprovideone type ofselective pressure. The virus, once
se-lected for greater resistance tothe original DI
particlepopulation can then generate DI parti-cles which can interfere withthereplication of thenewly selected virus as well as theoriginal
viruses
still
present inthe population. Thepop-ulation ofDI particlespresent has been shown to change during the course of the persistent infection(5).We do not yetknowwhethernewly arising DIparticlesinpersistent infections can
select for multiple rounds of mutations in a
similar manner or whether this selection only
occurs once, soonafter the establishment of the
persistent infection. Wearecarryingoutfurther
isolation of viruses fromCAR4andCAR21 and testing their susceptibilitytointerference
medi-ated by these DI particlestoresolve this
ques-tion.
We haverecently obtainedsomeevidence that
DIparticles might be involved in the selection ofmutations. We repeatedly passaged standard virus in BHK-21cellswithout dilution(toensure
thathigh levels of DI particlesarepresentduring
each serial passage). These serial undiluted
pas-sages appeartobegenerating mutations detect-able by
T,
oligonucleotide fingerprinting at amuch faster rate than passage either at high dilutionoralternating high and low dilutions (4,
21; K.Spindler, unpublished data). DI particles
may be partly involved directly by selection
effects and perhaps indirectly by slowing virus replication rates and prolonging cell survival times.However,manyother factors in
high-mul-tiplicity passages (such as complementation
amongmultiplemutantvirusesinthesamecell)
mayplayarole.J. S.Youngner, E.V.Jones,M.
Kelley, and D. W. Frielle (submittedfor publi-cation) have observedastriking increase in the
percentageoftemperature-sensitive mutantsof VSVpresentin thevirus population after 4 to
10 high-multiplicity passages in L cells.
How-ever,they didnotobserve this in BHK-21 cells
unlesstheypretreated the cells with actinomy-cinD, and they couldnotcorrelate the increase intemperature-sensitive mutantswith DI
par-ticle production. They postulate the
involve-mentof hostcellfidelity factors. Further work will be needed to assess the influence of DI
particlesonaccumulation ofmutations in virus
populations.
The location of the sites of mutationonthe
viral genomewhich conferarelative resistance
TABLE 2. Growth characteristicsof viruses isolated from persistent infectiona
Yieldat Yieldat
390C/
Virus 370C/yield at yieldat3300330C elat3C
ts31 0.24 0.00025
34-day SP 1.0 0.0093
34-dayLP 0.04 0.00025
75-day SP 0.50 0.0014
75-day LP 0.84 0.010
MSB(wild type) 0.69 0.11
aMonolayers of BHK-21
cells
were infected at amultiplicity of infection of 5 PFU/cell. Infections were carriedout at33, 37, and 39°C.Virusyield was deter-minedby plaque assay at 33°C.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
toDI particles shouldshedsomelight onthose
regions of the viral RNA which play a role in
autointerference. It has been suggested that the
complementary terminal sequences present in
the DI particle RNA may contribute to the
replicativeadvantage of the DI RNAduringan
infection (8, 16). Since the 5' end of the viral
RNAis conserved in theRNAof the DIparticle
(11, 16), and itscomplement is substitutedatthe 3' end, both of these sequences would be
re-flected in the 5' end of the viral RNA. It has
been shown that the 5' end of the viral RNA
mutatesmuchmorerapidly than does the 3' end
during persistence (21, 23). The virus variants described here may result from a coordinated
coselection of mutations in the 5' endof theviral RNAand in the gene(s) coding for thereplicase
(andpossiblyothergeneproducts).Sequencing
andhybridization studiesto testtheimportance of theseregions of the genome arebeing
initi-ated.
We thank EstelleBussey forexcellenttechnicalassistance. We also thank Julius Youngnerforcommunicating hisresults
tousbeforepublication.
This workwassupported byPublicHealthServicegrant
Al14627fromthe National Institutes ofHealth.F.M.H.was
supported byPublic Health ServicePredoctoral Traineeship GM07313.
LITERATURE CITED
1.Ahmed, R., and A. F.Graham.1977.Persistentinfection
in Lcellswithtemperature-sensitivemutantsof
reovi-rus.J. Virol. 23:250-262.
2. Bellet,A. J. D.,andP. D.Cooper.1959.Some properties
of thetransmissible interferingcomponentofvesicular
stomatitis virus preparations. J. Gen. Microbiol. 21:
498-509.
3. Doyle, M., andJ. J. Holland. 1973.Prophylaxis and
immunization inmice byuseof virus-freedefectiveT
particlestoprotectagainstintracerebral infectionby
vesicularstomatitisvirus.Proc.Natl.Acad.Sci.U.S.A.
70:2105-2108.
4. Holland, J. J., E. A.Grabau,C. L.Jones,and B.L
Semler.1979.Evolutionofmultiplegenomemutations
duringlong-tennpersistentinfectionbyvesicular
sto-matitis virus.Cell16:495-504.
5. Holland, J.J., andLP.Villarreal. 1974. Persistent
noncytocidalvesicularstomatitisvirusinfections
me-diated by defective T particles that suppressvirion
transcriptase. Proc.Natl. Acad.Sci. U.S.A.
71:2956-2960.
6. Holland,J. J.,LP.Villarreal,R.M.Welsh, M. B. A.,
Oldstone, D. Kohne,R Iazzarini, and E. Scolnick.
1976. Long-term persistent vesicular stomatitis virus
andrabiesvirusinfectionofcellsinvitro. J.Gen.Virol.
233:193-211.
7. Huang,A. S.,and D.Baltimore.1970.Defectiveviral
particlesandviraldiseaseprocesses.Nature(London)
226:325-327.
8. Huang,A.S.,S. P.Little,M.B. A.Oldstone, and D.
Rao.1978.Defective interferingparticles: theireffect
ongeneexpressionandreplicationofvesicular
stoma-titis virus,p.399-408.InJ. G.Stevens, G.J.Todaro,
and C.F.Fox(ed.),Persistentviruses.AcademicPress,
Inc.,New York.
9. Kawai, A., and S. Mat8umoto.1977. Interferingand noninterferingdefectiveparticlesgeneratedbyarabies smallplaquevariantvirus.Virology76:60-71.
10. Kawai,A.,S.Mataumoto,andKTanabe.1975. Char-acterizationof rabiesvimsesrecovered from
persist-entlyinfectedBHKcells.Virology67:520-533.
11. Leppert,M.,LKort,and D.Kolakofsky.1977.Further characterizationofSendai virus DI-RNA's:amodel for theirgeneration.Cell 12:539-552.
12. Meinkoth, J., and S. L.T. Kennedy. 1980. Semliki Forestviruspersistence inmouseL929cells.Virology 100:141-155.
13. Mudd,J.A.,R.W.Leavitt,D.T.Kingsbury,and J. J. Holland. 1973. Natural selection ofmutantsof
ve-sicular stomatitis virusbyculturedcellsofDrosophila
Melanogaster.J. Gen. Virol. 20:341-351.
14. Nishiyama,Y.1977. Studies of L cellspersistently in-fected with VSV: factors involved in theregulationof persistentinfection. J.Gen. Virol. 36:265-279. 15. Nishiyama,Y.,J.Ito,andK.Shimokata 1978.
Prop-erties of viruses selectedduring persistentinfection of Lcells with VSV. J. Gen. Virol. 40:481-484.
16. Perrault, J.,B.L Semler, R.W.Leavitt, and J. J. Holland. 1978. Inverted complementaryterminal
se-quences indefectiveinterfering particleRNAs of vesic-ular stomatitis virus and theirpossiblerole in autoin-terference, p. 527-538. In B. W. J. Mahy andR. D. Barry(ed.), Negativestrand viruses and the host cell. AcademicPress,Inc.,New York.
17. Popescu,M.,andF.Lehmann-Grube.1977.Defective interferingparticlesinmiceinfectedwithlymphocytic choriomeningitisvirus.Virology77:78-83.
18. Preble,0.T.,and J. S.Youngner.1973.
Temperature-sensitive defect ofmutantsisolated from L cells
persist-entlyinfected with Newcastle disease virus. J. Virol.12: 472-480.
19. Ramseur,J.IL,and R. M. Friedman.1977.Prolonged
infectionofinterferon-treatedcellsbyvesicular
stoma-titisvirus:possible role oftemperature sensitive
mu-tantsandinterferon.J.Gen.Virol.37:523-533.
20. Rima,B.K.,B.Davidson, and S. J.Martin.1977. The role of defectiveinterferingparticlesinpersistent infec-tionof Vero cellsbymeasles virus. J. Gen.Virol.35: 89-97.
21. Rowlands, D.,E.Grabau,K.Spindler,C.Jones,B. Semler,and J.Holland.1980.Virusprotein changes and RNAterminal alterationsevolvingduring
persist-entinfection.Cell. 19:871-880.
22. Schmaljohn,C.,and C. D. Blair.1977.Persistent infec-tion ofcultured mammali cellsbyJapanese
enceph-alitis virus. J.Virol.24:580-589.
23. Semler,B.L,and J. J.Holland.1979.Persistent vesic-ular stomatitis virus infection mediates base substitu-tions in viral RNAtermini. J.Virol.32:420-428. 24. Wagner,R.R.,A.Levy,R.Snyder,G. Ratcliff,and
D.Hyatt.1963.Biologicpropertiesoftwoplaque
var-iants of vesicularstomatitisvirus(Indianaserotype).J. Immunol.91:112-122.
25. Weiss, B.,R. Rosenthal, and S. Schlesinger. 1980.
Establishment and maintenance ofpersistentinfection bySindbis virus in BHK cells. J. Virol. 33:463-474.
26. Welsh,R. M.,P.A.Burner,J. J.Holland,M. B. A.
Oldstone,H. A.Thompson,andL. P.Villarreal.
1975. Acomparisonofbiochemical andbiological
prop-erties ofstandard and defective lymphocytic chorio-meningitisvirus.Bull.W.H.0.52:403-408.
27. Youngner, J. S., J. Dubovi, D. 0. Quagliana, M.
Kelley, and0.T. Preble. 1976.Role oftemperature sensitivemutantsinpersistentinfections initiatedwith
vesicular stomatitisvirus. J.Virol.19:90-101.