0022-538X/83/110396-09$02.00/0
Copyright ©1983, American SocietyforMicrobiology
Insertion Mutants
of Herpes
Simplex Virus
Have a
Duplication
of
the
Glycoprotein
D
Gene and Express Two
Different
Forms
of
Glycoprotein
D
MARYLOU G. GIBSON ANDPATRICIAG. SPEAR*
DepartmenitofMicrobiology, The University ofChicago, Chicago, Illinois 60637
Received 10 May 1983/Accepted 9 August 1983
Weproduced insertion mutants ofherpessimplex virus (HSV) that contain two functional copies of genesencoding different forms of glycoprotein D (gD). These viruses have the gene for HSV type 2(HSV-2) gD at the normal locus and thegene forHSV-1gD insertedinto thethymidine kinase locus. Results of immunoprecipi-tationexperiments done with monoclonalantibodies revealed thatboth gDgenes wereexpressed by these viruses, regardless oforientation of the inserted HSV-1 gD gene, and that maximal synthesis of both glycoproteins depended on viral
DNA replication. This apparently normal expression of the inserted HSV-1 gD
gene was from a DNA fragment (SauI fragment, 0.906 to 0.924 map units)
containing nucleotide sequences extending from approximately 400 base pairs
upstreamof the5'end of the gDmRNAtoabout 200 basepairs upstream of the 3'
end. The glycoproteins expressed from both genes were incorporated into the
surfaces of infected cells. Electrophoretic analyses of purified virions and neutralization studies suggest that bothglycoproteinswerealsoincorporated into virions. This nonpreferential utilization of both gene products makes these viruses ideal strainsforthe generationand characterization ofa variety ofmutations.
Glycoprotein D (gD) is one of several
enve-lope glycoproteins specified by herpes simplex
virus (HSV) (39). The two serotypes of HSV (HSV-1 and HSV-2) produce related, but
anti-genically and structurally differentiable, forms
ofgD (1, 4, 6, 13, 29, 30, 35, 36, 42). Little is knownabout thephysiological roleof this
glyco-protein except that itcanelicit theproduction of
neutralizing antibodies(4,30, 35,42),aproperty
shared with other HSVglycoproteins, and that
anti-gD antibodies can block virus-induced cell
fusion(A. G. Noble, G. T.-Y. Lee, R. Sprague,
M. L. Parish, and P. G. Spear, Virology, in
press).
The gene forgDwasmapped tothe S compo-nent of HSV DNA by analyses of HSV-1 x
HSV-2 recombinant viruses (23, 34) and, later, to a 2.9-kilobase (kb) Sacl fragment of HSV-1 DNA (Fig. 1) by selection and translation in vitro of gDmRNA(16,44). Nucleotide sequence
determinations, coupled with construction ofa
fused gene that expresses a X cro-gD hybrid
protein inbacteria, havepermittedprecise local-ization of the coding sequence forthe gD
poly-peptide of HSV-1 (44). The requirements for
adjacent nucleotide sequences upstream and downstreamofthecoding sequence for gDtobe expressed inproductivelyinfectedcells have not previously been explored, although it has been shown that gD can be expressed from the
BamHIJDNAfragment (0.89 to 0.93 map units) injected into oocytes (43).
Insertion of appropriate HSV-1 DNA frag-ments into thethymidine kinase (TK) gene of a gC- HSV-1 mutant permitted us to monitor expression of gC from the inserted DNA se-quencesof selectedthymidine arabinoside-resis-tant (AraTr) progeny and thereby to define the boundaries of the region necessary for expres-sion of the gC polypeptide in productively in-fected cells (17). In this report we describe a similar approach foridentification of sequences necessary forexpression of the gD polypeptide inproductively infected cells. Inthe processwe havealsogenerated viruses that contain duplica-tions ofthe gD gene. They express HSV-1 gD (gD-1) from the inserted DNA in the TK gene and HSV-2 gD(gD-2) from the normal genomic locus. We demonstrate expression and appar-ently normalregulation of both genesaswell as utilization of both products in virion morphogen-esis and transport to the cellsurface.
MATERIALS ANDMETHODS
Cells and viruses. Vero(African greenmonkey
kid-ney) cells, HEp-2 (human epidermoid carcinoma) cells, 143 (human TK--) cells, and rabbit skin cells
wereused in this study. The143cells(2),producedby C. Croce and K. Huebner, and the rabbit skin cells, isolated by J. McLaren, were obtained from B.
Roiz-396
on November 10, 2019 by guest
http://jvi.asm.org/
HSV MUTANTS EXPRESSING TWO FORMS OF gD 397
L b
a''c
S ca{ZJ~
mapunits 0 0 0 0 0 0
0.0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0 tk
~-4I
Ba SaNr Hn Sa Ec Ba
_gD-t
_[
_
§--s~~~~~~~~
I L
)
N
Eo
NrSo Ec Be
FIG. 1. ViralDNAsequencearrangements ofHSVstrain lAl, ofplasmidsused, and of insertion mutants
resulting fromrecombination betweenlAl and theplasmids. StrainlAl, the derivation of which is described in thetext,isaTK+ AraT5, HSV-1 x HSV-2 recombinant virus;theregionsof thegenomederived from HSV-1 andHSV-2areindicatedasreported by Tognonetal.(40).Location of the BamHIfragmentQof lAl is indicated bythe solid barnear0.3mapunits. TheTKgene mapswithin BamHIfragment Q (24, 41),and thegD-2gene
mapswithin the shortsegmentof the recombinantgenome(34)asshown. PlasmidpRB309 (25)containsaSacl
fragment of HSV-1(F) DNA(striped bar;mapcoordinates 0.906to0.924)inserted intothesingle Saclsite of BamHIfragment Q of HSV-1(F)DNA(solid bar), interruptingthecodingsequenceof the TKgene(24, 41).We reversed the orientationof theSacl fragmentwithrespecttothe TKgenetoobtain thenewplasmidpMG903. The gD-1gene hasbeenmappedtothisSacI fragment (16, 44).On the basis of nucleotidesequenceanalysesand otherresults,Watsonetal.(43, 44)located thegD-1 codingsequencetotheregionenclosedwithinbracketsand
reported that thesequencehomologoustogD-1mRNAspansfromneartheHindIIIsite(5' end)totheSaclsite
at0.925mapunits;the3'end of the mRNA liesapproximately200bp beyondthisSaclsite. Of thetwoNruI sites reportedtobein thisSacI fragmentofHSV-1(Patton)DNA(44), onlytheoneshown ispresentin HSV-1(F) DNA. Sequences homologous to uninterrupted TK mRNA are also indicated along with the direction of transcription (24, 41). Recombinationbetween either oftheplasmidsshown and strain lAl DNAresulted in the generation of insertion mutantsthat were AraTr. Abbreviations used for restriction endonucleases are: Ba, BamHI; Ec, EcoRI; Hn,Hindlll; Nr, NruI; Sa, Sacl.
man. An HSV-1 x HSV-2 recombinant virus,
desig-natedR5OBG13(alsoobtained from B. Roizman)and isolatedby Tognonetal.(40),wasmodified foruse as
theparentalvirus in thesestudies. Thisvirus, original-ly TK- and resistant to AraT, was made TK+ and
AraT-sensitive (AraTs) by rescue with the intact cloned TK gene (pRB103). Selection for the TK+ AraTs viruswasperformedin143 cells in thepresence of HAT medium(32).Theresultant virus, designated R5OBG13-1A1,willhereafter be called lAl.HSV-1(F) (8)andHSV-2(G) (8)werealso used in these studies.
Plasmid and viralDNAs. Therecombinantplasmids pRB103 [BamHI fragment QofHSV-1(F)DNA cloned intopBR322] (31)andpRB309 (25) (described inFig. 1)wereprovided byB.Roizman;pMG903was gener-atedasdescribed in thelegendtoFig.1, andpSKS309 containsaSaclfragment (0.906to0.924mapunits)of HSV-1(F) DNA cloned into the single Sacl site of pSKS106 (S.K.Shapira,J. Chou,F. V. Richard,and M. J. Casadaban, Gene, in press). Preparation and
determination ofauthenticity of each plasmid DNA
wereperformedasdescribed (16).
BamHI digests of genomic HSV DNAs prepared from infected Vero cells were separated on 0.6% agarose gels byelectrophoresis. Southern transfer of
DNA and hybridization with appropriate 32P-labeled
probeswereperformed(31).
Construction of recombinant viruses. Rabbit skin cells were cotransfected with mixtures of lAl viral
DNA andpRB309orpMG903DNAatvarious
concen-trations of each. Calcium phosphate precipitates of DNA were prepared, containing 50 to 200 ng of BamHI-digested plasmid DNA,8or12,ugof cytoplas-mic DNA from lAl-infected Verocells,and 12 ,ugof carrier DNAper0.6 ml. Rabbit skin cell cultureswere
treated with this DNAasdescribed(46). Progenyfrom
the transfected cellswereplatedonVero cells in the absence or presence ofAraT (Raylo Chemical,
Ed-monton,Alberta, Canada)at100,Lg/ml (33),and AraTr isolateswerescreened forgD-1 production by
immun-oprecipitationwithamonoclonalantibody specificfor gD-1 (anti-gD-1 [11436]). Isolates of interest were
plaquepurified twice.
Isotopic labelingof cellsand virions. (i) Continuous
labeling with [3.S] methionine. HEp-2 cells infected wtih HSV at a multiplicity of 10 PFU percell were
labeled from 4to 22 h afterinfection with [35S]methi-onine (10 p.Ci/ml, 1,084 Ci/mmol; New England
Nu-clearCorp.)forthepreparationofextractsfor
immuno-precipitation. Purified virionswerepreparedas previ-ously described (3) from HEp-2 cells infected at a
multiplicityof 3 PFUpercell and labeledat34WCfrom
5h after infection with[35S]methionine (5 ,uCi/ml)until
cytopathiceffect wascomplete. Purified virionswere
a b
[: D
lAI
pRB309
pMG903
gD-2
A.-
I/ HSV-I- 1% SV-2
-a S;a Hn
Bo Sa Hn
VOL.48,1983
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.491.68.413.70.246.2]pelleted, and proteins were solubilized in extraction
buffer forimmunoprecipitation.
(ii) Pulse-chase with [35S]methionine. Pulse-chase experimentswereperformedat4.5 hafterinfectionby labeling cells for 7 min with [35S]methionine (50
V.Ci/ml)
in methionine-deficient medium. Somecul-tures were harvested immediately after the pulse,
whereasotherswereincubated foranadditional 3 h in
nonradioactivemediumcontainingfive timestheusual
levels of methionine.
(iii)Cell surface labeling with Na'251. Intact HEp-2 cell monolayers infected at a multiplicity of 10 PFU
per cell were iodinated at room temperature at 24 h
after infectionaccordingtotheprocedureofSmith and
Brown(38)withNa'251 (250 p.Ciper4 x 106cells,17.4
Ci/mg; New England Nuclear Corp.). Post-iodination
washes of the labeled monolayerwereperformedwith
phosphate-buffered saline plus 10 mM Nal. Soluble proteins wereextractedas described below.
Antibodiesandimmunoassays. Monoclonal
antibod-ies directed againstHSVglycoproteinswereproduced
by M. F. Para inthislaboratory (Nobleetal.,inpress;
M. F.Para, R. Sprague,A. G. Noble, K. M.Zezulak, M.L. Parish, andP. G.Spear,manuscriptin
prepara-tion). The antibodies selected for use in this study
were 11436, specific for gD-1 (anti-gD-1); 111255 and
111114, both of whichreactwithgD-1 andgD-2 (anti-gD-1/2); and 1173, specific forgC-1.
Sequential immunoprecipitations were performed
on extracts preparedfrom infected HEp-2cells.
La-beled infected-cell monolayers were washed once in
phosphate-bufferedsaline andlysedin extraction
buff-erconsistingof 140 mMTris-hydrochloridebuffer(pH 7.4), 20 mM NaCl, 1% Nonidet P-40, 0.5% sodium
deoxycholate, bovine serum albumin (1 mg/ml), and
aprotinin (100 U/ml; Mobay Chemical Corp., New York, N.Y.). Cellextracts werecleared of nonsolubi-lized material by centrifugation at 178,000 x g in a
Beckman airfuge. Cell extracts were mixed with an
appropriate amountofantibody; after 30 minon ice,
Formalin-fixedStaphylococcusauireuis(14)wasadded for 10min, and immunecomplexes werecollectedby centrifugation. Antibody-treated supernatants from
onereactionwereusuallymixedwithasecondaliquot
ofantibody, orwith adifferentantibody, for second and subsequent rounds of immunoprecipitation. Pre-cipitated proteins were eluted from S. auirelis after washingasdescribed (16) and subjectedto
electropho-resis on 8.5% polyacrylamide gels linked with
N,N'-diallyltartardiamide (10). Fluorographywasperformed
asdescribed previously (15).
Antibody-mediatedneutralizationof viralinfectivity in thepresenceofcomplementwastestedbyaplaque
reductionassay. Mixtures containing 500PFU ofthe virus, the ascites form of the monoclonalantibodyata
final concentration of either 1/50or1/500, and
comple-ment at afinal concentration of 1/45 in 1 ml oftotal
volumewereincubatedfor1 hat37°Cand thenadded
to Vero cell monolayers as described (27). Percent
neutralized virus was calculated. Total transfection yields were assayed for the percentage of progeny expressinggD-1 byaslightlymodified version(17)ofa
plaque immunoassay developed byHolland et al.(11). RESULTS
Selection and characterization of insertion mu-tantsexpressing gD-1 and gD-2. The procedure for construction of the insertion mutants is de-tailed above andin thelegendtoFig.1.The 2.9-kb Sacl fragment containing the gD-1 coding
sequence was inserted in both orientations into
the TK gene of lAl virus by recombination
occurring aftercotransfection ofcells with lAl viral DNA and pRB309 or pMG903. The
per-centageofprogenyresistanttoAraTwas
signifi-cantly higher from cells transfected with lAI DNA plus plasmids than with lAl viral DNA alone(Table 1).Dataarealsopresentedshowing
that, of 63 AraTr plaque isolates obtained from transfections with plasmid and viral DNA, 62 (98%) were found to express gD-1 by immuno-precipitation analysis. Four isolates obtained from transfection with viral DNAalonewereall gD-1-. A total of 87% of 24 additional AraTr isolates from other transfections (data not
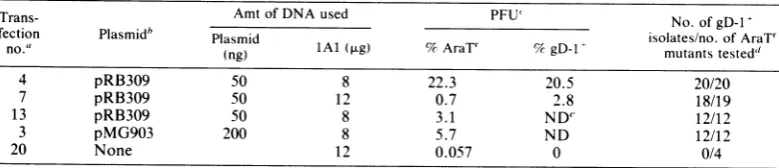
TABLE 1. Analyses of progeny virus obtained from cells cotransfected withlAl viral DNA and plasmid DNAs
Trans- Amt of DNA used PFU' No. of
gD-1i
fection Plasmidb Plasmid isolates/no.ofAraT'
no.' (ng) lAl (rig) % AraTY %gD-1l mutantstested'
4 pRB309 50 8 22.3 20.5 20/20
7 pRB309 50 12 0.7 2.8 18/19
13 pRB309 50 8 3.1 ND" 12/12
3 pMG903 200 8 5.7 ND 12/12
20 None 12 0.057 0 0/4
aPlaque isolates mentioned in the text were obtained from these transfections. The first number in the designationof each AraTr isolate denotes the transfection from which it was obtained.
bSeeFig. 1 and the text for derivation of plasmids.
CVirus titrations were done on Vero cells in the absence and presence of AraT (100
pLg/ml).
Titers in the absenceof AraT ranged from 20 x10'to 8.8 x 107 PFU/ml. Yields of some transfections were screened forgD-1 expressionby the in situ plaque immunoassay mentioned in the text and described elsewhere (11, 17).dVirus isolated from plaques formed in the presence of AraT was used to infect Vero cells in a 24-well dish. Eachculturewaslabeled with[35S]methioninebefore immunoprecipitation with anti-gD-1.
e ND, Not done.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.491.57.447.507.591.2]HSV MUTANTS EXPRESSING TWO FORMS OF gD 399
shown) also expressed gD-1 consistent with the expecta quences were inserted intoti ing the virus AraTr, and th;
expressed from the inserted I of two transfection yields in 20.5 and 2.8%,
respectively
expressed gD-1. Inthefirstc was thesame asthepercent ny; in the second case, 10-expressed gD-1 than were indicate that marker rescue insertion occurred to a var transfection, possibly due toconcentrations or ratios.
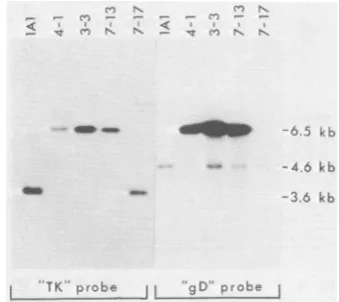
Genomic analyses of Ai performed to determine wh quence containing the gD-1
theBamHI Qsequence cont
Immobilized BamHI restri<
DNAs of representative Ara
were hybridized with either
(TK probe) or pSKS309 (g from allthree AraTrgD-1- is 7-13, have a novel 6.5-kb fre both to the TK probe and These isolates did not have t fragment characteristic of
strain. All isolates containe
N.n
Co -
-TK rob gC
[image:4.491.59.229.393.544.2]FIG. 2. Analysisof the genor progenyobtained fromcells cot
lAl DNA and either pRB309
restrictionfragmentsofgenomic
virus(lA1),threeAraTrgD-1+is
andoneAraTrgD-1-isolate
(7-a0.6%agarosegel by electropho
tonitrocellulose.Isolates4-1an
different transfections with pR
isolate 3-3 was from a transf DNA.Fragments containingseq
pRB103 (TK probe) or pSKS:
identifiedby hybridization with
probes.
These findings are kb which is derived from the HSV-2 DNA ttion that gD-1 se- sequence in the S component oflAl and deriva-he TK gene, render- tives and which hybridizes to the gD-1 probe. at the gD-1 gene is These results indicate that most AraTr isolates gene. Immunoassay have an insertion of the gD-1 gene into the TK
i situ indicated that gene. One AraTr isolate (7-17) identified as
gD-, of progeny virus 1- did not have an insertion in the TK gene, ase,this percentage indicating that it is a spontaneous TK mutant. age of AraTr proge- Thissamegenomicanalysiswas
performed
on -fold more progeny 35AraTrgD-1 + isolates before the second cycleAraTr. This would of plaque purification. All of these isolates had as well as marker an identical insertion of the Saclfragment into
*iable extent during the BamHI Qfragment (data not shown). Of four variations in DNA AraTrgD-1- isolates obtained from cells trans-fected with pRB309 and lAl DNA, none had raTr progeny were insertions, nor did four other AraTr gD-1- iso-iether the Sacl se- lates obtained from transfection with lAl DNA
gene wasjoined to alone (Table 1).
aining the TK gene. Sequential immunoprecipitation experiments
ction fragments of (Fig. 3) of infected cell extracts showed that
LTrprogeny (Fig. 2) mutant viruses 4-1, 7-13, and 3-3 expressed the
32P-labeled
pRB103 insertedgD-1 gene as well as retained the ability,D-1 probe). DNAs to express gD-2. Data are also presented in Fig. ,olates, 4-1, 3-3, and 3showing that the parental viruslAl, the AraTr agment homologous gD-1- isolate 7-17, and HSV-2(G) expressed to the gD-1 probe. only gD-2, whereas HSV-1(F), the strain from :he 3.6-kb BamHI Q which the cloned gD-1 gene was obtained, ex-the parental lAl pressed only gD-1. Apparent differences in the d a fragment of 4.6 amounts ofgD-1 and gD-2 precipitated may be due at least in part to differences in immunopre-cipitating efficiencies of the antibodies used. When both antibodies were tested against
sam-ples
of thesameantigen
preparation
fromHSV-M N s
1(F)-infected
cells,
anti-gD-1/2
precipitated
lessgD-1 than did anti-gD-1.
Theinserted gD-1 gene was expressed regard-less ofits orientation within the TK gene, indi-cating that transcription did not require the viral _
-6.5
kb TK promoter. The genes forgD-1
and TK aretranscribed in the same direction in isolates 4-1
--4 6 k b and 7-13, whereas transcription is in opposite -3.6 kb directions in isolate 3-3 (Fig. 1).
All of the insertion mutants analyzed in this study replicated in HEp-2cellsto titers compa-rable to those obtained with the lAl strain,
) probe HSV-1(F), orHSV-2(G). There were no
indica--P-Jitions that replication ofthe mutants wasin any nestructures ofAraTr way impaired by the insertion.
:ransfected with strain Processing of
gD-1
and gD-2 by the insertionor pMG903. BamHI mutants. Pulse-chase experiments were per-DNAsof theparental formed to compare the immature and mature solates(4-1, 3-3, 7-13), forms ofgD-1 and gD-2 made by an insertion 17) wereseparated on mutant with those made by other HSV strains ioresis andtransferred (Fig. 4). Infected-cell extracts (pulseand pulse-d7-13 camefromtwo chase) were sequentially and exhaustively
pre-B309 DNA, whereas
cipitated,
first
with
anti-gD-i
and
then
with
anti-e'ction with
pMG9O3
uenceshomologousto
gD-1/2.
The precursor and product forms of309 (gD probe) were gD-1produced by theinsertionmutant 7-13 were the indicatedlabeled indistinguishable by electrophoresis from those made byHSV-1(F). Similarly, the precursor and
VOL.48, 1983
on November 10, 2019 by guest
http://jvi.asm.org/
A
B
Uu.
FIG. 3. Production of gD-1 and gD-2 by AraTr insertion mutants. HEp-2 cellsinfected with plaque-purified individual isolates were labeled with [35S]methioninefrom4 to22hafterinfection, andthe extractsobtained were sequentially and exhaustively immunoprecipitated,first with anti-gD-1 and thenwith anti-gD-1/2 (111255). The resulting precipitates were analyzed by polyacrylamidegel electrophoresis. This autoradiogram of the gel shows the first precipitates obtained withanti-gD-1 (lanes designated 1), the sec-ondprecipitates obtained with anti-gD-1 (lanes desig-nated1*), and the third precipitates obtained with anti-gD-1/2 (lanes designated 2). Results obtained withthe parental strain lAl, HSV-1(F), and HSV-2(G) are shown for comparison with the results obtainedwith the AraTrisolates 4-1, 3-3, 7-13, and 7-17(previously describedin thelegendtoFig. 2). (A) 1AI, 4-1, 4-13, HSV-1(F); (B) HSV-2(G),7-17, 3-3.
product forms of gD-2 expressedby strain 7-13 could-notbedistinguishedfrom thegD-2of lAl and HSV-2(G). As previously noted (3, 6, 23,
34), both forms of gD-2 have slightly faster
electrophoretic mobilities than thoseofgD-1.
Effects of a DNA synthesis inhibitor on gD-1
and gD-2 accumulation. To determine whether the amountsofgD-1 and gD-2 made or accumu-latedbytheinsertionmutantsdependedonviral
DNAreplication,cellswereinfected and labeled
inthepresence ofphosphonoaceticacid (PAA), aspecific inhibitor of herpesvirus DNA
replica-tion (22, 26). Treatment of insertion
mutant-infected cells (7-13, 3-3) withPAA significantly
reduced the amounts of gD-1 and gD-2 that
accumulatedas comparedto untreated infected
cells (Fig. 5). The productionofgD-1 by
HSV-1(F) virus was also significantly inhibited by
PAA, as was gD-2 production by HSV-2(G)
virus. Similar findingswere previously reported
by Peake etal. (28).
The lateglycoprotein gC, immunoprecipitable
by ascites fluid I173, was undetectable in
PAA-treated cells infected with insertion mutant 3-3.
Extractsofuntreated and PAA-treated infected
cellsarepresentedinFig.5 toshow the
differen-tial effects of PAA on accumulation ofvarious
viral proteins.
Transport ofgD-1 to the infected-cell surface.
Lacto-peroxidase-catalyzed
iodination ofinfect-ed cells was performed to determine whether
both gDs made by the insertion mutants were
transportedtothe cellsurface.Extractsof
iodin-atedcellsweresequentially immunoprecipitated
as described. Analyses of these precipitates are
presented in Fig.6. Insertionmutants 4-1and
7-13 displayed both gD-1 and gD-2 on the cell
surface, whereas 7-17, an AraTr gD-1- mutant
A A3
...
FIG. 4. Processing ofantigenicallydistinctgDsby
an insertion mutant virus producing both glycopro-teins. HEp-2 cells infected with the mutant or with otherviruses as indicatedwerelabeled at4.5 h after infectionfor7min with[35S]methionine;extractswere
preparedfrom the cellseither immediately(pulse) or
after3hof additionalincubationinmediumcontaining
a fivefold excess of methionine (pulse-chase). For AraTr insertion mutant 7-13, theseextracts were
se-quentially precipitated with anti-gD-1 and then with anti-gD-1/2(111114)asdescribed inthelegendtoFig.
3.Onlythefirstandthirdprecipitatesareshown. For virusesexpressing onlyoneantigenic form of gD,the extractswereprecipitatedwithanti-gD-1in thecaseof HSV-1(F), orwithanti-gD-1/2 (111114)in the casesof lAl andHSV-2(G).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.491.52.242.72.381.2] [image:5.491.271.440.377.528.2]HSV MUTANTS EXPRESSING TWO FORMS OF gD 401
ZW
am^
*"
-a
so
an
; "_ _'- "I .L
FIG. 5. Effects of PAA treatment on production of gD-1 and gD-2 bytheinsertionmutants7-13and 3-3 and by strains lAl and HSV-1(F). HEp-2 cell monolayerswere pretreated with PAA (300 ,ug/ml) for4 hbefore infection; this level of PAA was maintained throughout the infection and labelingperiodwith[35S]methionine(5 to 22 h postinfection) in treated cultures (+). Control cultures (-) were not exposed to PAA. Sequential immunoprecipitations asdescribedin the legend to Fig. 3 wereperformedon extractsobtained from thecells, using first anti-gD-1 (lanes designated 1) and thenanti-gD-1/2(111114)(lanes designated 2). Extracts of control and treated culturesof insertion mutant 3-3 are also shown (E) along with immunoprecipitates obtained with anti-gC-1 (lanes designated 3).
with no insertion, displayed only gD-2. The viruses lAl, HSV-1(F), and HSV-2(G) ex-pressed either gD-1 orgD-2 only oninfected-cell
surfaces,asexpected. Anti-gD-1 precipitatedan
additional iodinated protein of125,000 daltons
from virusesexpressing gD-1.This
high-molecu-lar-weightmaterialwas notprecipitable by
anti-gB or anti-gC monoclonal antibodies (data not
shown). It could be a dimeric form of gD-1,
previously detected by Eisenberg et al. (7).
Alternatively, the iodination procedure could
have induced cross-linkingofgD-1 toothercell
surface components.
Incorporation of gD into the virion.
[35S]methi-onine-labeled virions of the insertion mutant 7-13 were purified on Dextran T10 gradients to
determinewhether bothgDs were incorporated
intotheviralenvelope. Electrophoretic analysis
of immunoprecipitates (Fig. 7) demonstrated
that both gD-1 and gD-2 were present in the
purified virion preparations oftheinsertion
mu-tant. The parental virus lAl incorporated only
gD-2 intothe virion, asexpected.
Neutralization tests were done with selected
insertion mutants to verify that gD-1 is on the virion surface and can serve as a target for immune recognition (Table2). Viruses
express-ing both types of gD were neutralized by anti-gD-1 but not as efficiently as was HSV-1(F)
virus, which expresses only gD-1. Although the extentof neutralizationofinsertion mutantsand
HSV-1(F) increased with increasing
concentra-tions of anti-gD-1, it is unclear whether total
neutralization might be achieved with this
anti-bodyunder theseconditions.Virusesexpressing
gD-1 alone, gD-2 alone, or both glycoproteins could be totally neutralized by anti-gD-1/2 at a dilution of 1/50.
DISCUSSION
We have isolated insertion mutants of HSV
which containtwodifferent forms of the gene for
gDandexpressandutilize theproducts of both
genes in a codominant fashion. These
experi--200 K
-130K
- 94 K
- 68 K
- 43 K
Al F 4-1 7-13 7-7 A' F 4- 7-13 7-17
W^S a ^ t| - g - 2
FIG. 6. Exposure of gD on the surfaces of cells infectedwith theparental viruslAl,HSV-1(F),orthe AraTr isolates 4-1, 7-13,or 7-17.lodinationofinfected cells was done as described in the text at 24 h after addition of virus. Extracts prepared from the cells were sequentially precipitated as described in the legend to Fig. 3. In this autoradiogram of the poly-acrylamide gel only the first immunoprecipitates ob-tained with each monoclonalantibodyareshown.
VOL.48,1983
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.491.114.377.76.224.2] [image:6.491.265.432.443.589.2]FIG. 7. IncorporationofgD-1andgD-2into virions
produced by an insertion mutant. Extracts (E)
pre-paredfrom[35S]methionine-labeledparental(1A1)and recombinant(7-13)virions are shown. Theseextracts
weresequentially immunoprecipitated, first with
anti-gD-1(lanesdesignated1)and thenagainwithanti-gD-i
(lanes designated 1*), followed by two successive
precipitations withanti-gD-1/2(111114, lanes
designat-ed 2 and2*).
mentsindicate thatgenetic informationrequired
in the vicinity of the gD-i coding sequence for theexpressionandproperregulationof thegene
is located within the Sacl fragment at map
coordinates 0.906through0.924onHSV-1DNA and that this information is functional when inserted into the TK gene.
In previous studies from this laboratory, in-sertionmutants were produced inpart todefine the coding sequence for HSV-1 gC (17). It was
pertinent in those studies as well as in these to prove that the additionalglycoprotein produced by the insertion mutants was being expressed
from sequences inserted into the TK gene and
not from the normal locus which had been convertedbymarkerrescue. ExpressionofgD-i bythemutantsproduced herealwayscorrelated withan insertion ofgD-i sequences into the TK
gene. Each isolate studied consisted of a pure
population of virus. Only one kind of genome
organization was detected when hybridization
studies were performed on insertion mutants,
making it unlikely that mixtures of virus could be responsible for the simultaneous production
of bothgD-i andgD-2. The salient evidence for the expression ofgD-i from inserted sequences
is that each clonally derived insertion mutant
expressed two antigenically distinguishable
forms ofgD. After exhaustive
immnoprecipita-tion and removal ofgD-i fromlysatesof infected
cells,gD-2 couldsubsequentlybe immunopreci-pitated. Consequently, although marker rescue was observed in some of ourexperiments
(ac-countingfor theAraT' gD-1 progeny;Table 1),
none of the insertion mutants analyzed was
alteredatthe normalgDlocusbytheacquisition
ofgD-1 sequences necessary for expression of
the gD-1 specific epitope recognized by the
monoclonal antibody used.
Differences in theamounts of gD-1 and gD-2 precipitable from recombinant virus-infected
cells could be due to lesser avidity of the
anti-gD-1/2 antibodies for solubilized antigen or for
S. aureus, rather than to differences in the
amountofgD-1andgD-2 produced. Alternative-ly, gD-1 may accumulate in larger amounts,
possibly due to the fact that, in large part, the
genetic background of the insertion mutants is HSV-1, and type 1 proteins may be selectively
produced by these viruses or may selectively
accumulate. Experiments different in design from thosereportedhere mustbedoneto
quan-titate the relativeratesof synthesisand
accumu-lation of gD-1 and gD-2 by the mutants.
ExpressionofagD-1-relatedprotein in
bacte-rialcellswasachievedbyfusingpartof the gD-1 coding sequence to the cro coding sequence of
bacteriophage X (44, 45). Expression of gD-1
proteinuponinjection of Xenopus oocyteswith the BamHI J fragment of HSV-1 DNA (map coordinates 0.89to0.93) has also been reported (43). Those results, coupled with nucleotide
se-quence analyses (44), have defined the coding sequencefor gD-1 aswell asdetermined thatno
additional viral functions coding outside of the BamHI Jfragmentare necessaryforexpression
of the genein oocytes.
Our results extend these findings and set the boundaries for any cis-acting sequences adja-centto thegD-1 coding sequence thatmight be necessary for the appropriate expression and
regulation of gD-1 in theinfected cell.
Apparent-TABLE 2. Neutralization ofinfectivity of insertion mutantsby monoclonal antibodies
PFU(% ofcontrol)W
Anti-gD-1at Anti-Virus gD expressed concn: gD-1/2
1/50 1/500 (1/50)
7-13 gD-1 + gD-2 23.2 35.6 <0.002
3-3 gD-1 + gD-2 25.3 32.3 <0.002
7-17 gD-2 86.4 106.0 <0.002
lAl gD-2 81.0 89.8 <0.002
HSV-2(G) gD-2 89.6 102.4 <0.003 HSV-1(F) gD-1 6.8 11.3 <0.002
aNeutralization assays were performed as de-scribed in the text. The valuesgivenarederived from the numbers of PFU detected after treatment with antibody plus complement, expressedas apercentage of the PFU obtained inthe presence ofcomplement alone. Numbers of PFU in control cultures ranged from368 to 556.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.491.60.235.62.228.2] [image:7.491.259.448.502.607.2]HSV MUTANTS EXPRESSING TWO FORMS OF gD 403
ly the SacI fragment (coordinates 0.906-0.924), already known to contain the gD-1 coding se-quence (16, 44), also contains any necessary promoter and regulatorysignals forgD-1
expres-sion. Several pieces of evidence support this
conclusion. First, gD-1 is expressed by the
in-sertion mutants in equivalent amounts
regard-less of orientation of the Sacl fragment within
theTK gene. Second,theinsertedgene
(regard-less of its orientation) is apparently expressed
under the same kind of control as is the gD-2 gene at the normal locus. Experiments with
PAA showed a significant decrease in the amounts of both gD-1 and gD-2 produced by
insertion mutants as well as by conventional
viruses in the presence of the drug, whereas expression of TK, an early or ,B product, is
enhanced under these conditions (9, 12, 18).
Therefore it seemslikely that the400-base-pair
(bp) sequence upstream of the gD-1 mRNA
initiationsite (bounded on the left by a Sacl site;
seeFig. 1) contains allpromoter andregulatory
signals necessary for expression ofgD-1 under
normal control intheinfectedcell. Mackem and
Roizman(19-21) showed that regulatory signals
influencingthe expression of HSVa geneslieat
least 110 to 140 bp upstream of thetranscription
initiation site and may even extend more than
330 bpupstream in some instances.
The 3' terminus of gD-1 mRNAlies200bp 3'
totheSaclsite oftheclonedgene(43, 44)and is
thereforenotcontained inthe sequences
insert-ed into the TK gene. Transcription termination
signals outside the Sacl fragment are
presum-ably used for
production
of functional gD-1mRNAsbytheinsertionmutants, as must be the
casefor theproduction of gC-1 mRNAby other
insertionmutants(17). Thusthereappears to be
no strict requirement with respect to sequence
of nucleotides in the 3' noncoding region, at
least forgD-1 and gC-1 mRNAs.
Several
investigators
have classified gDasanearlyor,Bpolypeptide, based on its appearance
by2h postinfection(1,5)oritsexpression after
acycloheximide block orwhen DNA synthesis
is inhibited (28),or basedon quantitation of gD
mRNAaccumulated in thepresenceofcytosine
arabinoside,aninhibitor ofDNA synthesis(44).
Our studies show thatsynthesisoraccumulation
of the gD polypeptide is significantly inhibited
by PAA,amoreeffectiveinhibitorofviral DNA
synthesis. EithergD is a late or y polypeptide
according to the original definitions proposed
(synthesis inhibited by the inhibition of viral
DNA replication; see reference 12), or
transla-tion of the gD mRNA (if actually made in
significant quantities when viral DNA
replica-tion is completely blocked) to yield a stable
product requiresafactormade under -y control.
The findings that both copies of thegD gene
are expressed in the insertion mutants under
apparently normal controls for this gene, and
that both glycoprotein products appear in the
cellsurfaceandinvirions, makethe
experimen-tal system described here especially attractive
forcertainkinds of studies of HSV gene
expres-sion. Mutatedforms ofaglycoproteingene can
beinserted into theTKgene to assess effects of
themutations onexpression of the gene and on
synthesis, posttranslational processing,
intracel-lular transport, andincorporation intocell mem-branes and virions. The effects of lethal
muta-tions can be studied because the product
expressed from the normal genetic locus can
supply all essential functions. The results
pre-sented here and earlier (17,25, 32, 33, 37) show
thatHSVcan be acloningvectorusefulforthe
isolationandstudy ofparticular HSVregulatory
sequences, genes, and their products in the
environment oftheHSV-infected cell. Thevirus
may also be a suitable cloning vector for the
study ofcertainnon-HSV genes.
ACKNOWLEDGMENTS
Variousaspectsof this workweresupported bygrantsfrom the NationalCancer Institute(CA 21776 and CA 19264) and theAmerican CancerSociety. M.G.G. was atrainee under PublicHealthServicegrant 5 T32CA09241from the National Institutes ofHealth and is now a fellow ofthe Leukemia Society of America.
LITERATURE CITED
1. Balachandran, N., D. Harnish, W. E. Rawls, and S. Bacchetti. 1982. Glycoproteins of herpes simplex virus type 2 as defined by monoclonal antibodies. J. Virol. 44:344-355.
2. Campione-Piccardo, J., W. E. Rawls, andS. Bacchetti. 1979.Selectiveassayforherpessimplex viruses express-ing thymidine kinase.J.Virol. 31:281-287.
3. Cassai, E. N., M. Sarmiento, and P. G. Spear. 1975.
Comparison of the virion proteins specified by herpes simplex virustypes 1and2.J. Virol. 16:1327-1331. 4. Cohen, G. H., M. Katze, C. Hydrean-Stern, and R. J.
Eisenberg. 1978. Type-common CP-1 antigen of herpes simplex virus is associated with a 59,000-molecular-weightenvelopeglycoprotein. J.Virol.27:172-181. 5. Cohen, G. H., D. Long, and R. J. Eisenberg. 1980.
SynthesisandprocessingofglycoproteinsgD and gC of herpes simplex virustype 1.J. Virol. 36:429-439. 6. Eisenberg, R.J., M. Ponce deLeon, andG. H.Cohen.
1980.Comparative structural analysis of glycoprotein gD ofherpes simplexvirustypes 1 and 2. J.Virol. 35:428-435.
7. Eisenberg,R.J.,M.Ponce deLeon,L.Pereira,D.Long, andG.H. Cohen.1982.Purification ofglycoprotein gD of herpessimplexvirus types 1 and 2byuseof monoclonal antibody.J. Virol. 41:1099-1104.
8. Ejercito, P. M., E. D. Kieff, and B. Roizman. 1968. Characterization ofherpes simplex virus strainsdiffering
in their effect on social behavior of infected cells. J. Gen. Virol. 3:357-364.
9. Garfinkle, B., and B. McAuslan. 1974. Regulation of herpessimplexvirusinducedthymidine kinase. Biochem. Biophys.Res. Commun. 58:822-829.
10. Heine, J. W.,R. W.Honess,E.Cassai, and B. Roizman. 1974.Proteinsspecified by herpes simplex virus.XII. The virionpolypeptides oftype 1strains.J.Virol.14:640-651. 11. Holland, T. C., R. M. Sandri-Goldin, L. E. Holland, S. D.
VOL. 48,1983
on November 10, 2019 by guest
http://jvi.asm.org/
Marlin, M. Levine, and J. C. Glorioso. 1983. Physical mapping of the mutation inanantigenic variant of herpes
simplex virustype1byuseofanimmunoreactiveplaque assay.J.Virol. 46:649-652.
12. Honess, R. W., and B. Roizman. 1974. Regulation of herpesvirus macromolecular synthesis.I.Cascade
regula-tion of the synthesisof threegroupsof viral proteins. J.
Virol. 14:8-19.
13. Honess, R. W., and D. H. Watson. 1974. Herpessimplex virus-specific polypeptides studiedby polyacrylamide gel
electrophoresis of immune precipitates. J. Gen. Virol. 22:171-185.
14. Kessler, S. 1975. Rapid isolation of antigens from cells
witha staphylococcal proteinA-antibody absorbent:
pa-rametersof the interaction ofantibody-antigen complexes
with protein A.J.Immunol. 115:1617-1624.
15. Laskey, R. A., and A. D. Mills. 1975. Quantitative film detection of 3H and14Cin polyacrylamide gels by
fluorog-raphy. Eur.J. Biochem. 56:335-341.
16. Lee, G. T.-Y., M. F. Para, and P. G. Spear. 1982. Locationof the structuralgenesf6rglycoproteins gD and
gE and for other polypeptides in the S component of
herpessimplex virustype1 DNA.J.Virol. 43:41-49. 17. Lee,G. T.-Y., K. L.Pogue-Geile, L.Pereira, and P. G.
Spear.1982.Expression ofherpessimplex virus
glycopro-tein C from aDNAfragmentinserted intothethymidine
kinasegene of this virus. Proc. Natl. Acad. Sci. U.S.A.
79:6612-6616.
18. Leiden,J.M., R. Buttyan, andP.G.Spear. 1976.Herpes
simplex virus gene expression in transformed cells. I.
Regulation of the viral thymidine kinase gene in
trans-formed L cells by products of superinfecting virus. J.
Virol. 20:413-424.
19. Mackem, S., and B. Roizman. 1982. Differentiation be-tweenpromoterand regulatorregions of herpes simplex
virus1: thefunctionaldomains ofamovablea regulator.
Proc.NatI. Acad. Sci.U.S.A. 79:4917-4721.
20. Mackem,S.,andB.Roizman.1982.Regulation ofagenes
of herpes simplexvirus: thea27genepromoter-thymidine
kinase chimera is positively regulated in converted L
cells. J. Virol. 43:1015-1023.
21. Mackem,S., andB.Roizman.1982.Structural featuresof
the herpes simplex virusa gene 4, 0, and 27
promoter-regulatorysequenceswhich conferaregulationon
chime-ricthymidinekinasegenes.J. Virol. 44:939-949.
22. Mao,J. C.-H., E. E. Robishaw, andL. R. Overby. 1975.
Inhibition of DNApolymeraseactivity from herpes
sim-plexvirus-infected Wi-38cells byphosphonoacetic acid.
J.Virol. 15:1281-1283.
23. Marsden, H. S., N. D. Stow, V. G. Preston, M. C.
Timbury, and N. M. Wilke. 1978. Physical mapping of
herpes simplex virus-induced polypeptides. J. Virol.
28:624-642.
24. McKnight, S. L. 1980. The nucleotide sequence and transcript mapof the herpessimplex virustype 1 thymi-dinekinasegene.Nucleic Acids Res.8:5949-5964. 25. Mocarski, E. S., L. E. Post, and B. Roizman. 1980.
Molecular engineering of the herpes simplex virus
genome: insertion of a second L-S junction into the
genome causesadditionalgenomeinversions. Cell
22:243-255.
26. Overby, L. R., E. E. Robishaw, J. B. Schleicher, A.
Reuter,N. L.Shipkowitz,andJ.C.-H. Mao.1974.
Inhibi-tion of herpessimplexvirusreplication by
phosphonoace-tic acid.Antimicrob. AgentsChemother. 6:360-365.
27. Para, M. F., R. B. Baucke, and P. G. Spear. 1982. GlycoproteingEof herpes simplexvirustype1:effects of
anti-gE on virion infectivity and on virus-induced
Fc-binding receptors.J. Virol. 41:129-136.
28. Peake, M. L., P. Nystrom, andL. I.Pizer.1982.
Herpesvi-rusglycoproteinsynthesis andinsertion into plasma mem-branes. J.Virol. 42:678-690.
29. Pereira, L., D. V.Dondero, D.Gallo, V.Devlin, andJ.D.
Woodie. 1982. Serological analysis ofherpessimplex virus types 1 and 2 with monoclonal antibodies. Infect. Immun. 35:363-367.
30. Pereira, L., T.Klassen, and J. R.Baringer. 1980. Type-common andtype-specific monoclonalantibodytoherpes simplexvirus type 1. Infect. Immun. 29:724-732. 31. Post, L. E., A. J. Conley, E. S. Mocarski,and B. Roizman.
1980.Cloningof the reiteratedandnon-reiteratedherpes simplex virus 1 sequences as BamHI fragments. Proc. Natl. Acad.Sci. U.S.A. 77:4201-4205.
32. Post, L. E., S.Mackem, and B. Roizman. 1981. Regulation of agenes ofherpes simplex virus: expression of chimeric genes produced by fusion of thymidine kinase withagene promoters.Cell 24:555-565.
33. Post, L. E.,and B. Roizman. 1981. A generalized tech-nique for deletion of specific genes in large genomes: a gene 22 ofherpes simplex virus 1 is not essential for growth. Cell25:227-232.
34. Ruyechan, W. T., L. S. Morse, D. M. Knipe, and B. Roizman. 1979. Molecular genetics of herpes simplex
virus. II. Mapping of the major viral glycoproteins and of thegenetic locispecifying the social behavior of infected cells.J. Virol. 29:667-697.
35. Showalter, J. D., M. Zweig, andB.Hampar. 1981. Mono-clonalantibodiestoherpessimplexvirus type1proteins,
including the immediate-early protein ICP4. Infect. Im-mun. 34:684-692.
36. Sim, C., and D. Watson. 1973. Therole of typespecific
andcrossreacting structuralantigens in the neutralization of herpes simplex virus types 1 and 2. J. Gen. Virol. 19:217-233.
37. Smiley, J. R.,B.S. Fong,and W.-C. Leung. 1981. Con-struction ofa double-jointed herpes simplexvirus DNA molecule: inverted repeatsarerequired for segment inver-sion and direct repeats promote deletions. Virology
11:345-362.
38. Smith, J. F., and D. T. Brown. 1977. Envelopment of Sindbis virus: synthesis and organization ofproteins in cells infected with wild type and maturation-defective mutants. J. Virol. 22:662-678.
39. Spear, P. G. 1980. Herpesviruses, p. 709-750. In H. A. Blough and J. M.Tiffany(ed.),Cell membranes and viral envelopes. AcademicPress, London.
40. Tognon, M.,D. Furlong, A. J. Conley,and B. Roizman. 1981. Molecular genetics of herpes simplex virus. V. Characterization ofa mutantdefective inabilitytoform plaques at low temperaturesandinaviralfunction which prevents accumulation of coreless capsids at nuclear pores late in infection. J. Virol. 40:870-880.
41. Wagner, M. J., J. A. Sharp, and W. C. Summers. 1981. Nucleotide sequence of the thymidine kinase gene of herpes simplex virus type 1. Proc. NatI. Acad. Sci. U.S.A. 78:1441-1445.
42. Watson, D. H., and P. Wildy. 1969. The preparation of "monoprecipitin" antisera to herpes virus specific anti-gens. J. Gen. Virol. 4:163-168.
43. Watson, R. J.,A.M.Colberg-Poley,C.J. Markus-Sekura, B.J. Carter, andL. W.Enquist.1983.Characterization of theherpessimplex virus type1glycoproteinDmRNA and expression of this protein in Xenopus oocytes. Nucleic Acids Res. 11:1507-1522.
44. Watson, R. J., J. H. Weis, J. S. Salstrom, and L. W. Enquist.1982.Herpessimplex virus type1glycoproteinD gene:nucleotide sequence and expression inEscherichia coli. Science218:381-384.
45. Weis, J. H., L. W. Enquist, J. S. Salstrom, and R. J. Watson. 1983. Animmunologically active chimaeric pro-teincontaining herpes simplex virustype 1glycoprotein
D. Nature (London)302:72-74.
46. Wigler, M.,A.Pellicer,S.Silverstein,R.Axel,G.Urlaub,
and L.Chasin. 1979. DNA-mediated transfer ofthe ade-nine phosphoribosyltransferase locus into mammalian cells. Proc. Natl. Acad. Sci. U.S.A. 76:1373-1376.



![FIG.7.gD-1ed(lanesprecipitationsproducedrecombinantwerepared 2Incorporation of gD-1 and gD-2 into virions byaninsertionmutant.Extracts(E) [35S]methionine-labeledpre- from parental (1A1) and (7-13) virionsare shown](https://thumb-us.123doks.com/thumbv2/123dok_us/1441147.96512/7.491.259.448.502.607/lanesprecipitationsproducedrecombinantwerepared-incorporation-byaninsertionmutant-extracts-methionine-labeledpre-parental-virionsare.webp)
