JOURNAL OFVIROLOGY, Apr. 1972, p.652-658 Copyright©1972 AmericanSocietyforMicrobiology
Vol. 9, No. 4 Printed in U.S.A.
A
Bacteriophage of Bacillus subtilis
Which
Forms
Plaques Only
at
Temperatures
Above
50
C
II.
Reduction
of
TSP-i-Specific Receptor
Sites
onCells Grown
at37
C
or
45
C,
and the
Temperature-Dependent
Inactivation
of
Replicating Phage1
J. R. LAMONTAGNE2 AND W. C. McDONALD
Departmenit ofMicrobiologyanidImmunlology, Tulante Uniiversity School of Medicinie, anid Biology Department,
Tullanie
Uniiversity,
NewOrleans,
Louisiania
70118Received forpublication 26 October 1971
The adsorption ofsubtilisphage TSP-1 to Bacillussubtilis WI 68wastested with
cells grownat 37, 45, and 53 C. Kineticdata, electron micrographs, and
quantita-tive measurement of the per cent phage adsorbed all indicated that irreversible
adsorption occurs only with cells grown at 53 C. IfTSP-1 was allowed to infect
cells at 53 C, subsequent shifts to 37 C inhibited phage replication and resulted in the inactivation ofplaque-forming units.
Thephysical andchemicalcharacteristics of the
subtilisphage,TSP-1, haverecentlybeendescribed (8). It was isolated from soil, contains
double-stranded deoxyribonucleicacid (DNA)withouta
thymineanalogue, and ismorphologically similar
toothersubtilisphages (3,5,10).Itdiffers drama-tically, however, from other B. subtilis
bacterio-phages in that it will formplaques only at
tem-peratures above 50 C. Clear plaques are formed
on any of the B.subtilis strains that grow above 50C, andonB. licheniformis9945A above 50C. This rigorous restriction to temperatures above 50 C (toanupperlimitof55C) maybea reflec-tion of the interaction of one or more factors, actingsinglyorinconcerttoinhibitplaque forma-tion below 50 C. These factors include: (i)
a temperature-sensitive restriction mechanism
which is inactivated above 50 C (6); (ii) the
ab-senceofphagereceptor sites below 50C, dueto
changes in cell wall structure(4);and(iii) TSP-1 may haveintermediatereplicativeenzymeswhich
require the higher temperature for optimal
activity.
As aninitial step inidentifying which of these factors mayaccountfor thisstringenttemperature
requirement, we have asked the following ques-tions: (i) will TSP-1 adsorb to B. subtilis W168 growing attemperatures below 50 C, and (ii) if
1This paper wastaken in partfrom adissertaition submitted by J. R. LaMontagne inpartial fulfillmenit ofthereqluiremiients for thePh.D.degreeatTulanieUniversity,NewOrleans,La.70118.
2Present address: Depairtment of Microbiology, School of Medicine,Universit) ofPittsbturgh,Pittsburgh,Pa.15213.
adsorption is a limiting factor, can TSP-1
replica-tion continue below 50C once phageadsorption and DNA injection have occurred at 53 C? We
approached the first question by analyzing the kinetics of phage adsorption with experiments similar tothe classicexperiment of Hershey and Chase (7), by measuring the amount of phage which would adsorb to B. subtilis cells growing at
various temperatures and by examining negative stains of TSP-1-infected B. subtilis with the elec-tron microscope. To answer the secondquestion,
the initial steps in phage replication, phage
ad-sorption, and DNA injection were allowed to occur at 53 C. The infected cultures were then transferred to37 Cto determine if phage replica-tion could be completed at this temperature.
MATERIALS AND METHODS
Bacterial strains and bacteriophage used. All of theseexperiments were donewith TSP-1 infecting B.
suibtilis
W168, a prototrophic transformant of B.suibtilis
168, unless otherwise noted.Growth conditions. All bacterial cultures were
growninDifco BrainHeartInfusion (BHI) broth
me-dium.Brothcultureswereincubatedat53,45,or37C
in reciprocating shaker-water baths (New Brunswick Scientific Co.).Plaquecounts weredoneonBHI agar
(1.5',) by the agaroverlay technique (1) with0.1 ml ofa B.
suibtilis
W168sporesuspension (108spores perml)astheinoculum forthelawn. Serialdilutionswere
made in BHIbroth,and theoverlaywasBHIsoftagar
(0.65,).
Preparationof TSP-1 antisera.Anti-TSP-1 antisera
wasprepared in rabbits withTSP-1 suspensions
puri-652
on November 10, 2019 by guest
http://jvi.asm.org/
fied as previously described (8). Primary inoculations consisted of 0.25 ml of a TSP-1 suspension (1012 plaque-forming units) emulsified in an equal volume of complete Freund's adjuvant (Difco) and injected into the footpad. This was followed by two 0.5-ml subcutaneous injections ofTSP-1 at 10-day intervals. Rabbits were bled by cardiac puncture 6 to 8 weeks after the primary inoculation. The sera were collected andheated for 30 min at 36 C to inactivate comple-ment. Antisera obtained by this procedure had K values of at least 400.
Temperature shift experiments. Bacteria were incu-bated in 300-ml Nephelo flasks (side arm = 14 by 130mm;Bellco, Vineland, N.J.) containing 40 ml of BHI in areciprocating shaker-water bath at 53 C. The bacterial growthwasmonitored witha Klett-Summer-soncolorimeter with a red filter (640 to 700 nm) and incubated until the culture reached a turbidity of 75 Klettunits (equivalent to 5 X 107 bacteria per ml). At thistime, TSP-1 wasaddedtothedesired cultures at a multiplicity of infection (MOI) of 1. The shift to a lower temperature was made by transferring the desired flask to a second reciprocating shaker-water bath at the appropriate temperature Under these
conditions,aculturetransferred from53 to 37C,for
example, reached 37 C within 1.5 to 2.0 min after
transfer. In those experiments which involved the transferfrom53 to37C,followed byareturn to53C, 0.1 mlof undiluted phage antiserawas addedshortly after the shift to 37 C.
Radioactive labeling of TSP-1 DNA. TSP-1 was
labeled with 32P-orthophosphate (285 Ci/mg; Inter-national Chemical and Nuclear Corp.). The radio-active phosphatewasadded to aculture ofB.subtilis W168 growing in BHI to afinal concentration of 3
,gCi/ml
2 hr before addition of TSP-1. The labeled phage were purified by the polyethylene glycol-sodiumchloride procedurepreviouslydescribed (8).Kinetics of TSP-1 adsorption and DNA injection. Thekinetics of TSP-1 adsorption and DNAinjection
werestudied byaproceduresimilartothat ofHershey
andChase (7). Phagelabeled with 32pwere addedto
late log-phase culture of B. subtilis W168
(approxi-mately 108 bacteria perml) growingat53 C at aMOI of 10.Samples (3ml)weretakenat5-minintervals and divided into twofractions as follows: (i) 0.5 ml was
filteredimmediatelyontonitrocellulose filters(25mm
diameter; 0.45 ,um pore size; Millipore Corp.) and washed with 15 ml ofminimal salts (7); (ii) the
re-maining 2.5 ml washomogenized in aSorvall
Omni-mixer (IvanSorvall, Inc., Norwalk, Conn.),equipped
witha5.0-mlmicroattachment, for 60 secat asetting
of5. A0.5-ml sample of this homogenate was then
filtered and washed. This homogenization procedure
wassufficientto removeany adsorbed phageparticles
asjudged by removal ofTSP-1 particles labeledwith
'4C-isoleucine. Thefilterswere placed in scintillation vials, dried, and counted with Spectrafluor (Amer-sham/Searle)-toluenein aBeckman Instruments 200B
liquid scintillation counter.
Electron microscopy. TSP-1 was addedto cultures ofB.subtilis W168(MOI = 50) growingat53or37C inBHI. After 10 minof furtherincubation, 10ml of
the culture was centrifuged in a clinical centrifuge
(International Equipment Co., Needham, Mass.) at maximum speed (ca. 3,000 X g) for 10 min. The pelletwas suspended in 5ml of 10-2 M tris(hydroxy-methyl)aminomethane (Tris)-10-3 M MgSO4 buffer,
pH7.4. Adropof the suspendedpelletwasmixed with
an equal amount of
2%/o
phosphotungstic acid-2%0 normalrabbit serum, pH 7.0,andexaminedonForm-var-coated grids in aPhillips 300E electronmicroscope operating at 80 kv.
RESULTS
Adsorption of TSP-1 to B. subtilis W168.
TSP-1waslabeledwith
32p,
purified, and added toB.subtilis W168 growing under various tempera-ture conditions. If adsorption was not
accom-panied with DNA injection, the cell-associated radioactivitywould be removable by homogeniza-tion. In contrast, ifadsorption was followed by DNA injection, the radioactivity would be
re-tainedafterhomogenization.
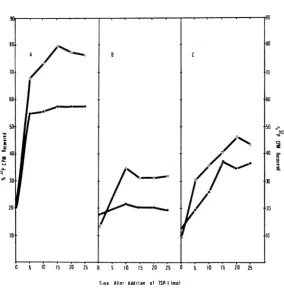
It is apparentfromtheresults of these experi-ments that TSP-1 will adsorb efficiently to 53 C growncells and thatDNAinjectionaccompanies
this adsorption (Fig. IA). Adsorption at 53 C occurred rapidly, and a plateau wasreached by 5
min after infection. A comparable situation does
not prevail at 37 C, however (Fig. 1B). In this case,TSP-1 adsorbed to 37 C cells, but not to the degree observed with 53 C grown cells, andonly
20%of the labelremained afterhomogenization. These results suggest two alternative explana-tions: (i)B. subtilisgrown at 37 C does nothave TSP-1 receptors on itssurface, and (ii) B. subtilis
grown at37 Chas the proper receptors,butDNA injection will not occur because it is a thermophilic
process.Toanalyze thispossibility,B.sutbtiliswas grown at 53 C, shifted to 37 C 10 min before addition ofTSP-1 (MOI = 10),and the kinetics of phage adsorption and DNA injection were
determined. Theresultsshowthatadsorptionand DNAinjection take place under these conditions, indicating that TSP-1 DNA injection is not a
thermophilic process (Fig. IC). Collectively, these results can be interpreted to indicate that
TSP-1adsorption to 53 C cells isspecific,whereas
adsorption to 37 C cells is not. In this context,
specific adsorption is defined as that adsorption
whichleadstoDNAinjection.Theseexperiments suggest that specific phage receptors are not present on cells grown at 37 C and that any
ad-sorptionto37Ccells isprobably nonspecific.Itis alsoclearthatphageDNAinjectioncan occurat
37Ciftheproperphage receptorsarepresent.
To confirm this suggestion, B. subtilis W168
grown at 37 and 53 C were infected withTSP-1 (MOIca.50),negatively stained,andexaminedby electron microscopy. These micrographs clearly
indicated thatTSP-1 does not irreversibly attach
on November 10, 2019 by guest
http://jvi.asm.org/
LAMONTAGNE AND McDONALD
5 10 5 20 25 5 tO 15 20 25 5 10 15 20 i5
Time After Addition of TSP-1(min)
FIG. 1. KitneticsofTSP-IadsorptioniandDNAinjection. Bacillussubtilis W168 wasgrowti tolate logphase
(108 cells/ml) antd 32P-labeled TSP-I wasadded(MOI = 10) tocellsgrownz at53 C (A), 37 C (B),and 53 C
cells shiftedto37 C 10mitibeforeTSP-Iaddition (C).Cell-associated radioactivity after (0) antdbefore (0) ho-mogenization.
toB.subtilisW168cellsgrownat37 C, but does
attachto53 Cgrowncells.
An additional experiment was done to verify the absenceofphagereceptorsitesat37C. In this instance, TSP-1 was added to B. subtilis W168
growingat37 C(MOI = 1.0)andincubatedat37
Cforanadditional 5or10min before theculture wastransferredto53 Candmonitored turbidime-tricallyfor thedevelopmentoflysis. IfTSP-1 did
notadsorb due to theabsence ofphage receptor
sites, it should be rapidly neutralized by TSP-1 antiserum, and the culture would be protected frominfection. Theseresultsclearly showthatthis
is thecase, forin allculturestowhich antiserum
was added the cultures did not manifest any
evidenceofphage infection (Fig. 2). All cultures
towhichnonspecificserum wasaddedlysed about
60to70minafter transferto53C.
Estimation of per cent phage adsorbed to B. subtilis W168 growing at53 and 45 C. A simple
method to quantitate the number of bacterio-phage particleswhichadsorbtoa suitablehost is toaddafewdropsofCHC1sto the
bacteria-bac-teriophage suspension andthentoassaythe resul-tant aqueous supernatant fluid for surviving
in-fectiouscenters(2).Thisprocedure actually meas-uresthe number of particles thatareirreversibly
adsorbed to bacteria, since it kills uninfected as
wellasinfected bacteriabut has noeffect on
un-adsorbed particles. The number of adsorbed
phagecanthen be determined by subtractingthe
titer obtained with CHCI3 treatment (i.e., free
phage), from the titer obtained without CHCI3
treatment (i.e.,freephage plusinfectedbacteria). However, when this experiment was done with
TSP-1 andB. subtilis W168 (MOI = 1.0)at53C,
the numberof infectiouscentersobtained withor
without CHC13 was identical. Moreover, if one compares these titers with the titer calculated
from the initial phage inoculum, it is apparent
thatabout 95% of the input phageisinactivated
very soonafteraddition of TSP-1 (Fig. 3B).This
coincidence inphagetiters indicates that infected cellsexposedtotemperaturesbelow 50 C willnot
formplaques. This loss inphage may be due to
the temperaturechangewhichaccompanies
dilu-tionandsample platingbytheagaroverlay
tech-nique. To test the possibility that extracellular
phage particles may be inactivated by a rapid
temperature shift, a TSP-1 suspension was
incu-B
A / *-_r
I
_-~~
C
_,
0. o
'00_
8C
V
cr4G
C.L
a 30
20 10
80
10
60
50
,0v
i
0
a.
.20
10
654 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.500.122.407.64.353.2]
200A-zD~~
X
F 0 ' lio
'12,
2UO i lie@ 2Uf * t'1@ 2i2~20
~
DEL''C
r;
t
\
200-~ ~
~
1/Ti0
200
100
200
100
0 120 240 0 120 240 0 120 240
[image:4.500.101.388.62.386.2]TIME AFTER INFECTION (min)
FIG. 2. Susceptibility ofTSP-I toneutralizationiby phage antiserum ifTSP-I isaddedto B. subtilis W168
growingat 37C. TSP-Iaddedattime(a) (MOI= 1.0), transferto53Cattime(b), andanitiserumornonspecific
serumaddedat lime (c). Key: uninfectedculturetransferredto 53 Cat T = 5mill (A); intfected culture
trans-ferredto53 Cat T = 5min, antiserum addedimmediately before transfer to 53 C(B); infectedculture
trans-ferredto53 Cat T 10 min, antiserum addedimmediately before transferto 53C(C); inifectedculture
trans-ferredto53Cat T =5miii (D); infectedculture transferred to 53 Cat T = 5 miii,nonzspecific serum added
immediately before tranisfer(E); infected culture transferredto53 Cat T = 10 miii, nionispecific serum added
imnmediately before transferto53 C(F).
bated at 53 C, rapidly transferred to 37 C, and
assayedatvarious times before and after transfer. The results of these experiments indicated that
TSP-1 particles per se are not inactivated by a
rapidtemperaturechangesincetherewas noloss ofinput after transfer to 37 C. It wouldappear
then thatreplicating phagearesensitivetoadrop
in temperature, andas aconsequenceplaqueswill
not be formed from infected bacteria. One can
then conveniently measureTSP-1 adsorption by assayinganinfectedculturewithoutCHCl3
treat-ment and comparingthis titer with the one
esti-mated from the phage inoculum. Repeated
ex-periments indicate that TSP-1 adsorption to B.
subtilis and B. licheniformis 9945Aoccursrapidly,
with 90to95%input phageadsorbed after 2min
at53C. Thislossofinput phage,however,isnot
evidentifoneaddsTSP-1 to B.subtilis incubated
at45 C(Fig. 3A) or37 C. Inthiscase,thephage
titers obtained withorwithoutCHC13arereduced
by about 40% oftheestimatedphage titer. There
isalsonoevidenceofphage replication (Fig. 3A)
comparable to the burst of new phage particles
observedat53C (Fig. 3B).Addition ofTSP-1to
cells incubated at 37 Cgives thesame results as
thecellsat45C (datanotshown), indicatingthat
irreversible phage adsorption and injection are reduced under these circumstances.
Temperature-dependent inactivation of replicat-ing TSP-1. The results presented above
demon-stratethatTSP-1 specificreceptorsitesareeither
not present or are not accessible on B. subtilis
W168cellsgrownat37 C. Inaddition,itappears
adsorbedphage canbe inactivatedbydecreasing
thetemperatureofincubation. These observations
imply that, although adsorption is a barrier to
on November 10, 2019 by guest
http://jvi.asm.org/
LAMONTAGNE AND McDONALD
20 40 0 20 40 60
TIME AFTER INFECTION
[image:5.500.69.260.66.297.2]min
FIG. 3. One-step growtAl ctirvesofTSP-1 doneoiiB.
subtilis W168growiiigat45 C (A) and 53 C (B). Tlte
claslhed linierepreseitts thlephage titercalcuilatedfroni
thephiage inoculione.P/lagetiters obtaiiied with CHCI3
(-) actcd withloit CHCI3 (O).PFU = plaquie-formniflg
units.
TSP-1
replication
below 50C,
it is not theonly
one. To define more
clearly
this apparenttem-perature-dependent
inactivation andits effect onphage
replication,
we undertook severalexperi-mentsinwhich TSP-1 wasaddedtoB.subtilisat
53 C
(to
allowadsorption), transferring
the in-fected cultureto37 Catvarious times after infec-tion anddetermining
ifthelytic cycle
could becom-pleted
at this temperature. Weemployed
twoindexesof
replication: (i)
theturbidimetricmeas-urementof host cell
lysis
and(ii)
themeasurementofnew
phage synthesis by
infectiouscenterassays.In the first
experiment,
TSP-1 was added toB.subtilis W168
(MOI
=1.0)
at53C.Theculturewastransferred to 37 C
immediately
afterinfec-tion,
and the number of infectious centers wasdetermined with or without
CHC13
treatment atvarious times after infection. These titers were
compared
with the calculatedinput phage
titer andplotted
against time. Over 95 'S- of thephage
inoculumwas
destroyed,
and thesurviving
phage
titerwasmaintained for the duration ofthe
experi-ment. These results indicate that
replicating
phage
weredestroyed
and that furtherphage
adsorption, injection,
andreplication
did notoccur at 37 C. In contrast, infected cultures at
53 Cshowedanidentical loss of
input phage
butwascharacterized
by
a burst ofnewphage
parti-cles about 30min after infection
(Fig. 3B).
To determine if it was possible to inactivate
replicatingTSP-1 atvarious times after
infection,
theeffectsofatemperatureshift from53 to37C
on the turbidity of infected and uninfected B.
subtilis W168 were observed. Ten flasks ofBHI
broth were inoculated withB. subtilis W168 and incubated at 53 C until they reached Klett 75
(5 X 107 bacteriaperml). TSP-1 wasthen added (MOI = 1.0) to nine of these flasks. At 5-min intervals through 35 min, cultures were
trans-ferredto 37C alongwith the uninfectedcontrol.
Inthis case, itwas possibleto determine atwhat timethetemperature requirementwas overcome.
Those infected flasks which were transferred to
37 Catless than 30minafterinfection continued
toincreased in turbidityataboutthe same rateas
theuninfected control culture. Flaskstransferred
attimesafter 30 min eventuallylysed,
suggesting
thattherequirement for 53 Cwas overcomeafter 30 min at 53 C. However, this experiment
only
measured changes in turbidityand did not
indi-cate if phage replication was actually taking
place, evenin those cultures transferred to 37 C after 30 min at 53 C.We tested this directlyby
transferring samples ofaninfected cultureto37C
atvarious times after infection, incubating them
at 37 C for 1 hr and thenassaying for infectious
centers. Thetiters of the transferred cultureswere
compared to the titers ofthe mother culture at
the time of transfer. If the transferred culture showed ahigher titer than the mother culture at
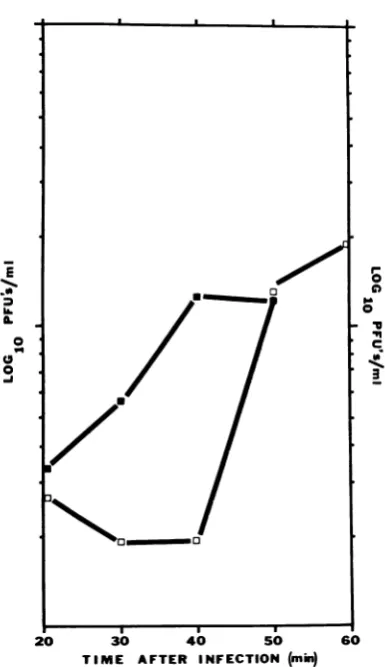
thetimeof transfer, thenonecouldconclude that in that sample replication had continued after transfer to 37 C. The results of one of these
ex-periments are shownin Fig. 4 and confirm that infectedcultures must bemaintained at 53 C for
atleast40minafter infectiontoobtainreplication
at 37 C in this experiment.
To determine how long anincubation at37 C wasrequiredtoobtain theirreversibleinactivation ofreplicating phage, infected culturesat53 Cwere
transferredto 37 Cfor2-minintervals atvarious times after infection, phageantiserum was
added,
and the culture was returned to 53 C. This time interval was used because it is just sufficient to
bring thetemperature of the medium from 53 to
37C. Theresultsdemonstrate that 2 min at37 C issufficient to prevent the lysis of infected cultures provided thattheexposure at 37 C occurs during
the first35minafterinfection.
DISCUSSION
The data presented above demonstrate that
there are at leat twobarrierstoTSP-1 replication
attemperatures below 50 C. One obstacle is the
absence of phage receptor sites on B. subtilis grown at 37 C or 45 C. This observation is
656 J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
E~~~~~~~
./.
1 16
L~~~~~~~~~~
0 0
20 30 40 50 60
[image:6.500.47.240.68.402.2]TIME AFTER INFECTION (min)
FIG. 4. Effect of trantsfer from53to37 ConiTSP-I
replicationinB. subtilis W168. At T = 0, TSP-1 was
added(MOI = 5); after20miii ofincubationiat53C,
3.0-misampleswereremoved andin2cubatedat37 Cfor
1 hr. At theenid ofthle 37 C inicubationiperiod, these
sampleswereassayed forTSP-I. Thephagetitersafter
1hrat37 C(a) werecomparedtothe phage titers in
theoriginalculture at thetimeof tranlsfer(0) to37 C.
strongly supported byevidence obtained through
four different experiments. Kinetic data indicate thatphage adsorptionand phageDNAinjection
occurredonly with cells grown and incubated at
53C,ortoalesser extent, with cells that hadbeen
grownat53 C untilshortly (10min) beforephage
addition and then transferred to 37 C. Phage
DNA injection was shown not to be a
thermo-philic eventsinceit couldoccurwith 53 C grown
cells incubated at37 C. Electron micrographsof
infected cellsgrownat53 and 37Cconfirmedthe
absence of irreversible TSP-1 adsorption. Addi-tional evidence was provided by quantitative
measurementsof the percentphageadsorbedto
cellsgrowingat53, 37,or45 C. Thedisappearance
ofTSP-1receptor sitesmaybe another
manifesta-tion of the cell wallchanges observed by Dul and
McDonald (4) in B. subtilis growing at 53 and 37 C. They observed a thicker cell wall and a greaterthermal resistance in B. subtilis incubated at 53 C. Additionally, the thermal resistance in 37 C grown cells can be induced by antibiotics which inhibit the final cross-linking of the peptido-glycan. It is possible that the phage receptor site for TSP-1 isassociatedwith this non-cross-linked peptidoglycan. Young et al. have shown that the specificity for phage receptor sites resides in the glucosylation of the polyglycerol teichoic acid (11). Perhaps the absence of a cross-linked pepti-doglycan is required for the suitable exposure of phage receptors on the teichoic acid. Another
possibilityis that theteichoic acidpresentin cells grown at 53Cisqualitatively different from that at 37C.
Apparently the absence of TSP-1 receptor sites
on37 C grown cells is nottheonlyencumbrance tophagereplicationat37C. Our results reveal a temperature-dependentinactivation of replicating TSP-1. This becomes evident when one allows TSP-1 toadsorb to cells growing at 53 C, shifts these to 37 C, and observes the culture for evi-dence of TSP-1 replication. These resultsclearly
indicate that if infected cells are transferred to 37Cduringthe initial 30 to 35 minofinfection, theinhibition ofmaturephageparticle production occurs.Thistimeperiodissimilartothe observed latent period of TSP-1 inB.subtilis (8). One can offeratleasttwoexplanations for this effect. The
inhibition could be due to the presence of a
tem-perature-sensitive restriction mechanism.
Evi-dence hasbeenpresentedfor suchasystem in B. subtilis 168 by Gwinn and Lawton (6). They
observed thattwo serologically identical phages,
SP-10and SP-20, formed plaques onB. subtilis W23Srbutnot onB.subtilis 168.These
bacterio-phages could form plaques on B. subtilis 168,
however, if the bacteria were incubated at 53 C
for 10minbeforebeing infectedat37C. Further-more, their evidence suggested a temperature-sensitive repressor, as they were able to rescue
phagemarkersfrom infectedB.subtilis 168aslate
as 3 hr afterinfection. Ifthisrestriction mecha-nismplaysarole in theinhibitionof TSP-1 repli-cationat37C,thenpreincubationat53 Cshould be sufficient to allow phage
replication
at 37 C. This does not occur in TSP-1infections. The alter-nativeexplanationfor thistemperature-dependentinactivation may be that TSP-1 istrulya
thermo-philic bacteriophage. That is, its phage-specific
enzymesareinactive below50C.If this is thecase
theneitherphage-specific DNA,ribonucleic acid,
or protein synthesis might be
temperature-sensi-tive.
on November 10, 2019 by guest
http://jvi.asm.org/
LAMONTAGNE AND McDONALD
ACKNOWLEDGMENTS
This investigation was supported by Public Health Service
training grant STOl GM-00079 from the National Institute of General MedicalSciences.
We thank J. Oaks for assistance with electron microscopy. LITERATURE CITED
1. Adams, M. H. 1959. Bacteriophages. Wiley-Interscience Publishers,Inc.,New York.
2. Clowes, R. C., and W. Hayes, (ed.). 1968. Experiments in microbialgenetics.JohnWiley and Sons, Inc., New York. 3.Davison, P. F. 1963. The structure of bacteriophage SP-8.
Virology21:146-151.
4. Dul, M. J., and W. C. McDonald. 1971. Morphological changesandantibiotic-inducedthermalresistance in
vege-tativecells of Bacillus subtilis. J. Bacteriol. 106:672-678. 5.Eiserling,F.A.,and E.BoydelaTour. 1965.Capsomersand
otherstructuresobserved insomebacteriophages. Pathol. Microbiol. 28:175-180.
6. Gwinn, D. D., and W. D. Lawton. 1968. Alteration of host specificityinBacillus subtilis. Bacteriol.Rev.32:297-301. 7. Hershey, A.D.,and M. Chase. 1952. Independent functions
of viralprotein and nucleic acid in growth of bacteriophage. J. Gen.Physiol.36:39-56.
8. LaMontagne, J. R., and W. C. McDonald. 1972. A bacterio-phage ofBacillus subtilis which forms plaques only at
temperaturesabove 50 C.I.Physical and chemical
charac-teristics ofTSP-1.J. Virol. 9:646-651.
9. Spizizen, J. 1968.Transformation of biochemically deficient strains of Bacillussubtilis bydeoxyribonucleate.Proc. Nat. Acad.Sci. U.S.A.44:1072-1078.
10. Tikhonenko,A.S. 1970. Ultrastructure of bacterial viruses. Plenum Press, New York.
11. Young, F.E., C. Smith, and B. E. Reilly. 1969. Chromosomal location ofgenesregulating resistancetobacteriophage in Bacillussubtilis. J.Bacteriol. 98:1087-1097.