N. Vassilaki,aK. I. Kalliampakou,aI. Kotta-Loizou,aC. Befani,bP. Liakos,bG. Simos,bA. F. Mentis,cA. Kalliaropoulos,cP. P. Doumba,d D. Smirlis,eP. Foka,aO. Bauhofer,fM. Poenisch,fM. P. Windisch,gM. E. Lee,gJ. Koskinas,dR. Bartenschlager,fP. Mavromaraa
Molecular Virology, Hellenic Pasteur Institute (HPI), Athens, Greecea; Laboratory of Biochemistry, School of Medicine, University of Thessaly, Larissa, Greeceb; Laboratory of
Medical Microbiology, HPI, Athens, Greecec; Second Department of Internal Medicine, Medical School of Athens, Hippokration Hospital, Athens, Greeced; Molecular
Parasitology, HPI, Athens, Greecee; Department of Infectious Diseases, Molecular Virology, University of Heidelberg, Heidelberg, Germanyf; Applied Molecular Virology,
Institut Pasteur Korea, Seongnam-si, South Koreag
Low oxygen tension exerts a significant effect on the replication of several DNA and RNA viruses in cultured cells.In vitro prop-agation of hepatitis C virus (HCV) has thus far been studied under atmospheric oxygen levels despite the fact that the liver tissue microenvironment is hypoxic. In this study, we investigated the efficiency of HCV production in actively dividing or differentiat-ing human hepatoma cells cultured under low or atmospheric oxygen tensions. By usdifferentiat-ing both HCV replicons and infection-based assays, low oxygen was found to enhance HCV RNA replication whereas virus entry and RNA translation were not af-fected. Hypoxia signaling pathway-focused DNA microarray and real-time quantitative reverse transcription-PCR (qRT-PCR) analyses revealed an upregulation of genes related to hypoxic stress, glycolytic metabolism, cell growth, and proliferation when cells were kept under low (3% [vol/vol]) oxygen tension, likely reflecting cell adaptation to anaerobic conditions. Interestingly, hypoxia-mediated enhancement of HCV replication correlated directly with the increase in anaerobic glycolysis and creatine kinase B (CKB) activity that leads to elevated ATP production. Surprisingly, activation of hypoxia-inducible factor alpha (HIF-␣) was not involved in the elevation of HCV replication. Instead, a number of oncogenes known to be associated with gly-colysis were upregulated and evidence that these oncogenes contribute to hypoxia-mediated enhancement of HCV replication was obtained. Finally, in liver biopsy specimens of HCV-infected patients, the levels of hypoxia and anaerobic metabolism mark-ers correlated with HCV RNA levels. These results provide new insights into the impact of oxygen tension on the intricate HCV-host cell interaction.
H
epatitis C virus (HCV) infection causes a wide range of clin-ical manifestations, from a healthy carrier state to acute and chronic hepatitis that can lead to fibrosis, cirrhosis, and hepato-cellular carcinoma. Nearly 3% of the world’s population is chron-ically infected with HCV (1, 2), and current therapeutic ap-proaches are not broadly effective (3).HCV is a positive-strand RNA virus with a 9.6-kb genome that is flanked at both termini by conserved, nontranslated regions (NTRs), required for RNA translation and replication. The 5=
NTR comprises an internal ribosome entry site (IRES) that directs the expression of a polyprotein precursor (4,5). The polyprotein is cleaved into structural (core, E1, E2) and nonstructural (p7, NS2, NS3, NS4A, NS4B, NS5A, NS5B) proteins that, in association with cellular factors, form a membrane-associated replicase complex. This copies the viral positive-strand RNA into a negative-strand intermediate that serves as the template for the synthesis of prog-eny genomes. The alternative reading frame (ARFP) or core⫹1 and minicore proteins, with as-yet-unknown functions, appear to be synthesized from the core region by alternative translation mechanisms (6,7).
Studies of the HCV replication cycle have first become possible in 1999 with the development of the replicon system (8). With the identification of a particular HCV genotype 2a isolate (JFH1) that replicates very efficiently in cell culture, a fully permissive HCV culture system was established (9). This system was improved upon development of intragenotypic chimeras consisting of the JFH1 replicase (NS3 to NS5B) fused to the J6 core-to-NS2 region (10,11) as well as of cell culture-adapted variants of the wild-type (wt) JFH1 virus (JFH1/adpt) (12).
To date, HCV proliferation has been studied exclusively under atmospheric oxygen tension (20% [vol/vol] O2) (13). However,
liver normoxic conditions range from 12% O2around the portal vein to 1% O2near the central vein (14), with a median value of 3% O2(15). This oxygen gradient is important for a metabolic activity zonation (16,17) that is reflected by an asymmetric distri-bution of key enzymes. The capacity for oxidative energy metab-olism, glucose release, and oxidation protection is higher in the periportal area, whereas the capacity for glucose uptake, glu-tamine formation, and fatty acid synthesis is higher perivenously. In tissue culture, low oxygen triggers an adaptive reprogram-ming of cellular homeostasis and bioenergetics (18). Concerning hepatocytes, low oxygen is essential for the preservation of their structure and metabolism (19). Moreover, hypoxia affects the rep-lication of several viruses (20–26).
Based on these observations, we studied the impact of oxygen tension on HCV replication and virus production in human hepa-toma (Huh7) cells. We show that low oxygen selectively enhances HCV RNA replication at an early stage and in a hypoxia-inducible factor (HIF)-independent manner. We provide evidence that
onco-Received22 September 2012Accepted19 December 2012
Published ahead of print26 December 2012
Address correspondence to P. Mavromara, [email protected], or N. Vassilaki, [email protected].
N.V. and K.I.K. contributed equally to this article and should be regarded as joint first authors.
Supplemental material for this article may be found athttp://dx.doi.org/10.1128 /JVI.02534-12.
Copyright © 2013, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.02534-12
on November 7, 2019 by guest
http://jvi.asm.org/
genes associated with increased anaerobic energy metabolism, as well as creatine kinase B (CKB), are upregulated under hypoxia and that they are responsible for the observed HCV RNA replication enhance-ment.
MATERIALS AND METHODS
Cell culture.Huh7.5 (27) and Huh7-Lunet (28) cells were grown in high-glucose (25 mM) Dulbecco’s modified minimal essential medium (Invit-rogen) supplemented with 2 mML-glutamine, 0.1 mM nonessential amino acids, 100 U/ml penicillin, 100g/ml streptomycin, and 10% (vol/ vol) fetal calf serum (complete DMEM). To create oxygen tensions lower than the atmospheric one, cells were cultured in a fully humidified incu-bator supplied with pure nitrogen gas to reduce oxygen as well as with 5% (vol/vol) CO2at 37°C (New Brunswick CO2incubator; Innova) (29). For
dimethyl sulfoxide (DMSO) differentiation, 8⫻104Huh7.5 cells were
seeded per well of 12-well collagen-coated plates; 24 h later (at 90% con-fluence), the medium was supplemented with 1% (vol/vol) DMSO, and cells were further cultured for 14 days.
Human liver biopsy specimens. Human liver fine-needle biopsy specimens were obtained from patients with chronic HCV infection fol-lowing consent. Sample collection was approved by the Ethical and Sci-entific Committee of the Hippokration Hospital of Athens.
Plasmid construction.All amino acid and nucleotide numbers refer to the JFH1 genome (GenBank accession no.AB047639[30]). Plas-mids pFK-JFH1wt, pFK-JFH1/adpt1/mut4-6 (referred to as JFH1/ adpt1), I389RLuc2ACore-3=-Jc1 (JcR2a), Jc1, and pFK-I389LucUbH77Core-EIJFH1NS3-3= (LucCore-NSJFH1) have been described previously (9,11, 12, 31,32). pFK-I389Luc-EIJFH1NS3-3=_delGDD (Luc-NSJFH1/delGDD) has an in-frame amino acid dele-tion (MLVCGDDLVV) encompassing the GDD motif of NS5B. pEGFP-HIF-1␣has been described previously (33). pEGFP-HIF-2␣carries the human HIF-2␣cDNA (obtained from pcDNA-HIF-2␣; kindly provided by S. L. McKnight) fused carboxy-terminally of the enhanced green fluo-rescent protein (EGFP) sequence of pEGFP-C1 (Clontech). p9xHRE-Luc carries nine copies of hypoxia response element (HRE) and rat prolactin minimal promoter upstream of the firefly luciferase gene (kindly provided by R. Hernandez-Alcoceba, University of Navarra, Pamplona, Spain). The cytomegalovirus (CMV)-driven JUN-FOS expression vector and the re-porter plasmidMMP1-luc, carrying a consensus AP-1 binding site, were kindly provided by L. Bakiri (Fundación Banco Bilbao Vizcaya, Madrid, Spain) (34).
In vitrotranscription.Full-length and bicistronic HCV constructs were linearized with MluI and used forin vitrotranscription as described previously (12).
In vitro-transcribed RNA and plasmid DNA transfection. Electro-poration with viral RNAs into Huh7-Lunet cells was performed as de-scribed elsewhere (32). For transfection with Luc-NSJFH1/delGDD, Huh7-Lunet cells seeded at 50 to 60% confluence were treated with Tran-sIT-mRNA transfection reagent and mRNA Boost reagent (Mirus) as rec-ommended by the manufacturer. Plasmid DNA transfections were per-formed at 40 to 50% cell confluence usingTransIT-LT1 transfection reagent (Mirus).
siRNA transfection.The small interfering RNAs (siRNAs) targeting HIF-1␣ (5=-AGGAAGAACTATGAACATAAA-3=; NM-001530) and HIF-2␣(5=-CCCGGATAGACTTATTGCCAA-3=; NM-001430) and the AllStars negative-control siRNA were obtained from Qiagen. The siRNA tar-geting CKB (5=-CGUCACCCUUGGUAGAGUUTT-3=; NM_001823) and the scramble negative-control siRNA to CKB (5=-GGCGUACUAGCUU AUUCGCTT-3=) were purchased from Sigma. Huh7.5 cells seeded at 40 to 50% confluence were transfected with siRNA using Lipofectamine 2000 transfection reagent (Invitrogen) as recommended by the manufacturer. Preparation of HCV cell culture (HCVcc) virus stocks and infection assays.Virus stocks were generated as described elsewhere (32) and used to infect naive Huh7.5 cells. Culture medium was exchanged 4 h after virus inoculation.
Virus titration in cell culture supernatants.HCV was titrated essen-tially as described elsewhere (10). Infectivity titers were determined using the JFH1 NS5A-specific mouse monoclonal antibody 9E10 (kindly pro-vided by C. Rice, The Rockefeller University, NY) (10) and expressed as the 50% tissue culture infective dose (TCID50)/ml.
Western blot analysis.Western blotting was performed as described elsewhere (35). Dilutions of 1:1,000 for JFH1 NS5A monoclonal antibody (9E10 [10]), 1:500 for human HIF-1␣mouse monoclonal antibody (BD Biosciences), 1:2,000 for human-actin mouse monoclonal antibody, 1:100 for GFP rabbit polyclonal antibody (Santa Cruz Biotechnology), and 1:1,000 and 1:500 for human phosphorylated and total AKT rabbit monoclonal antibodies (Cell Signaling), respectively, were used.
Indirect immunofluorescence.Indirect immunofluorescence analy-sis of JFH1 NS5A was performed as described elsewhere (32). DNA was stained with propidium iodide (Sigma). Images were acquired with the Leica TCS-SP four-channel confocal microscope equipped with an argon ion laser and helium-neon laser.
Luciferase assays.Firefly and renilla luciferase (F-Luc and R-Luc) activities in cell lysates were measured using the respective chemilumines-cent assay kit (Promega), as recommended by the manufacturer, in a GloMax 20/20 single-tube luminometer (Promega) for 10 s. Luciferase activities were normalized to the total protein amount determined using the Bradford assay reagent (Pierce).
Measurement of intracellular ATP levels.ATP was measured using the ViaLight HS BioAssay kit (Lonza) according to the manufacturer’s protocol in a GloMax 20/20 single-tube luminometer (Promega) for 1 s. ATP levels were normalized to total protein amounts.
RNA quantification by quantitative reverse transcription-PCR (qRT-PCR) and a branched DNA assay.Total cellular RNA was extracted using the NucleoSpin RNA II kit (Macherey-Nagel). cDNA synthesis was performed with Moloney murine leukemia virus reverse transcriptase (Promega) according to the manufacturer’s protocol and with a mixture of the specific primers (see Table S1 in the supplemental material) JFH1-5=NTR-354R and YWHAZ-R for the HCV positive-strand RNA and the 14-3-3-zeta polypeptide (YWHAZ) mRNA, respectively, JFH1-5=NTR-276F and YWHAZ-R for the HCV negative-strand RNA (3.5 pmol/l of each primer), or pd(N)6random hexamer primers (GE Healthcare Life
Sciences) for the cellular transcripts. Real-time quantitative PCR was per-formed using the SsoFast EvaGreen supermix (Bio-Rad) as well as primer pairs specific for the JFH1 5=NTR or the cellular YWHAZ, vascular en-dothelial growth factor A (VEGFA), glucose transporter 1 (GLUT1), hexokinase 2 (HK2), enolase 1 (ENO1), lactate dehydrogenase A (LDH-A), CKB, FOS, MYC, SRC,␣1-antitrypsin (A1AT), and hepatocyte nu-clear factor 4 alpha (HNF4-␣) (see Table S1 in the supplemental material). The YWHAZ housekeeping gene was selected as a normalization control, as it was confirmed that its expression was not affected under low-oxygen conditions (36). Total cellular RNA from human liver biopsy (LB) speci-mens was prepared using the RNeasy Plus Mini total RNA isolation kit (Qiagen), and HCV positive-strand RNA was quantified using the Ver-sant HCV RNA 3.0 (branched DNA [bDNA]) assay as recommended by the manufacturer.
Chemicals.AKT inhibitor VIII (AKTi-1/2) was obtained from Calbi-ochem.
Statistical analysis.Only results subjected to statistical analysis using Student’sttest with aPvalue ofⱕ0.05 were considered statistically sig-nificant and presented. Statistical calculations were carried out using Ex-cel in Microsoft Office.
RESULTS
Low oxygen tension induces HCV production in hepatoma cul-tured cells.HCV naturally infects hepatocytes that are exposed to an oxygen concentration ranging from 1 to 12% (14), with a me-dian value of 3% (vol/vol) O2(15). However, to date, the HCV replication cycle has been studied exclusively under atmospheric oxygen conditions (20% O2) in all cell culture systems (13).
Vassilaki et al.
on November 7, 2019 by guest
http://jvi.asm.org/
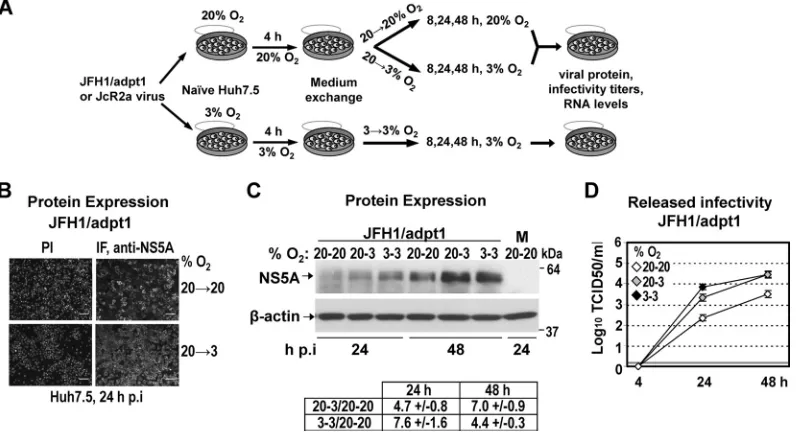
To investigate the effect of low oxygen (hypoxia and 3% O2) on the efficiency of HCV replication, Huh7.5 cells were infected with a highly assembly-competent JFH1 isolate (JFH1/adpt1) (Fig. 1), and virus production was evaluated at 3% and 20% O2(Fig. 2A), as follows: subconfluent Huh7.5 cells preincubated for 18 h at
20% or 3% O2 were inoculated with HCV (under the oxygen
conditions of preincubation), the virus inoculum was withdrawn
4 h later, and cells were further incubated at 20% O2(a condition hereafter referred to as 20¡20), transferred immediately to 3% O2(20¡3), or further incubated at 3% O2(3¡3) for the indi-cated time points. The last condition (3¡3) was intended to in-duce low-oxygen-mediated cellular changes prior to infection. HCV production was assessed by evaluating viral protein expres-sion and release of infectious HCV particles by using indirect im-FIG 1Schematic representation of HCV constructs used in this study. From top to bottom: JFH1 wild-type (wt) virus genome; JFH1/adpt1, cell culture-adapted JFH1 genome containing virus titer-enhancing mutations (indicated with asterisks) (12); Jc1, chimeric virus genome composed of the J6CF structural and JFH1 nonstructural regions (gray and white boxes, respectively); JcR2a, a Jc1 derivative containing the R-Luc gene (striped box) fused N-terminally to 16 codons of the core gene (white box) and C-terminally to the FMDV 2A protease-coding sequence (gray striped box); LucCore-NSJFH1, a bicistronic replicon encoding a firefly luciferase (F-Luc)-ubiquitin (Ub)-core fusion protein (N-terminal 161 residues of the H77 isolate); Luc-NSJFH1/delGDD, containing a three-codon deletion in the NS5B region (delGDD) gene. Black bars in all panels indicate HCV NTRs. EI, IRES of the encephalomyocarditis virus.
FIG 2Low oxygen tension enhances the production of JFH1-derived viruses in Huh7.5-infected cells. (A) Schematic representation of the experimental procedure. Supernatants of Huh7-Lunet cells electroporated with JFH1/adpt1 or JcR2a RNA (10g/4⫻106cells) and incubated at 20% O
2were collected and used for infection of naive Huh7.5 cells (1 TCID50/cell) that were seeded at 30% confluence (to avoid pericellular hypoxia) and preincubated for 18 h at 20% or 3% O2, respectively. Cells preincubated at 20% O2were cultured at either 20% or 3% O2until harvest (20¡20 or 20¡3, respectively). Cells preincubated at 3% O2were incubated at 3% O2(3¡3) until harvest. (B) Indirect immunofluorescence for NS5A in JFH1/adpt1-infected cells incubated as specified on the right of the panels. Nuclei were stained with propidium iodide (PI; left column). Bar, 80m. A representative experiment of two independent repetitions is shown. (C) (Top) Western blot analysis of NS5A (top) in JFH1/adpt1-infected cells incubated as specified in the top of each lane. M, mock-infected cells. (Bottom)-Actin served as a loading control. Numbers on the right refer to the positions of molecular mass marker proteins. A representative experiment of 3 independent repetitions is shown. (Bottom) Imaging quantification of NS5A signals, normalized by using-actin signals. Quantity I Bio-Rad software was used to quantify the respective signals from the 3 independent repetitions of the Western blot analysis. Mean values are expressed relative to that obtained at 20¡20% O2. (D) Enhanced release of infectious HCV at 3% O2from JFH1/adpt1-infected Huh7.5 cells, as determined by a TCID50assay. The gray horizontal line indicates the cutoff of the assay (⬃2 TCID50/ml). A representative experiment of three independent repetitions is shown.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:3.585.114.483.64.216.2] [image:3.585.95.499.396.612.2]munofluorescence or Western blotting and TCID50assays, re-spectively.
As shown inFig. 2B, at 24 h postinfection (h p.i.), the NS5A immunofluorescence staining of JFH1/adpt1-infected Huh7.5 cells was noticeably increased at 3% O2(20¡3) compared to that at 20% O2(20¡20). Accordingly, an enhanced accumulation of NS5A protein (5- to 7-fold) was also detected by Western blot analysis in lysates of JFH1/adpt1-infected Huh7.5 cells cultured under low-oxygen conditions (20¡3 or 3¡3) (Fig. 2C). In addi-tion, a significant titer increase was observed with cells cultured at 3% O2(about 10-fold for the 20¡3 condition and up to 30-fold for 3¡3) compared to cells cultured at 20% O2both 24 h and 48 h p.i. (Fig. 2D), suggesting that low oxygen provides an advantage to HCV propagation in Huh7.5 cells.
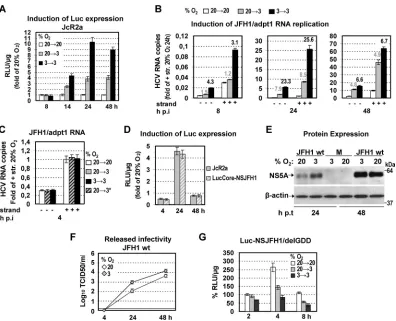
Low oxygen tension induces HCV genome replication dur-ing the early stages of infection.We next investigated the partic-ular step of the viral replication cycle that is affected by low O2. We first determined viral RNA replication by using the JcR2a reporter virus (Fig. 1) containing the R-luc gene. An enhancement of lucif-erase activity was found with cells cultured at 3% O2, arguing for elevated viral RNA replication under hypoxia (Fig. 3〈; see also Fig. S1〈in the supplemental material for 20% O2). This result was supported by analysis of cells cultured under the same conditions and infected with the JFH1/adpt1 virus, thus excluding the possi-bility that the observed enhancement is caused by effects on lucif-erase (Fig. 3B). Importantly, independent of the readout (lucifer-ase assay, negative- or plus-strand RNA by qRT-PCR), replication was stimulated most in cells cultured consistently at low O2 FIG 3Three percent oxygen positively regulates the HCV genome replication. (A and B) Enhanced HCV RNA replication in Huh7.5 cells infected (1 TCID50/cell) with one of two viruses. (A) JcR2a virus. Cells were cultured as specified in the upper left, and replication-derived luciferase activity is expressed as relative light units (RLU) perg of total protein amount. Values obtained with 20¡20 cells were set (each time) to one. (B) JFH1/adpt1 virus. qRT-PCR analysis of the intracellular HCV-negative (⫺) and -positive (⫹) strand RNA copies from cells incubated at 20¡20, 20¡3, or 3¡3% O2, expressed relative to the positive-strand RNA obtained at 20¡20% O2at 24 h p.i. YWHAZ mRNA levels were used for normalization. Above the respective bars, ratios of RNA copies at 20¡3 and 3¡3 versus 20¡20% O2for each time point are shown. (C to F) Low oxygen tension does not affect virus entry. (C) qRT-PCR analysis of the intracellular HCV-negative (⫺) and -positive (⫹) strand RNA copies from Huh7.5 cells inoculated with JFH1/adpt1 virus (1 TCID50/cell) and incubated for 4 h as specified. 20¡3* refers to cells that were preincubated at 20% O2and transferred immediately after virus addition from 20% to 3% O2. Values (normalized as described above) are expressed relative to the positive-strand RNA obtained at 20¡20% O2at 4 h postinoculation. (D) Luciferase activities, derived from JcR2a and LucCore-NSJFH1 RNA-transfected Huh7-Lunet cells (10 and 5g RNA/4⫻106cells, respectively). After transfection, the cells were further cultured at 20% or 3% O2, and luciferase activities at 3% O2are expressed as fold inductions of those obtained at 20% O2. (E and F) Enhanced viral protein expression and release of infectivity at 3% O2upon JFH1 RNA electroporation (10g RNA/4⫻10
6cells) of Huh7-Lunet cells. (E) (Top) Western blot analysis of NS5A production in cells incubated at 20% or 3% O2. (Bottom)-Actin served as a loading control. M, mock nontransfected cells. A representative experiment of two independent repetitions is shown. (F) Kinetics of release of infectivity from the viral RNA-transfected cells incubated at 20% or 3% (TCID50assay). A representative experiment of three independent repetitions is shown. (G) Three percent oxygen does not positively affect HCV RNA expression. Huh7-Lunet cells preincubated at 20% or 3% O2for 18 h were transfected with Luc-NSJFH1/delGDD RNA and further incubated as specified on the upper right. Luciferase values measured with 20¡20% O2cells harvested 4 h p.t. were set to 100%. In all panels, bars represent mean values from at least three independent experiments in triplicate. Error bars indicate standard deviations.
Vassilaki et al.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.95.491.63.384.2](3¡3). We therefore concluded that low O2has a positive effect on HCV RNA replication, best visible at early time points. The profound difference between RNA levels obtained with cells cul-tured at 3¡3 and those obtained with cells cultured at 20¡3% O2 conditions argues for a replication-promoting effect by the preex-isting cellular response to low oxygen.
In order to examine whether the significantly higher levels of viral RNA are due to enhanced viral entry, we determined the amounts of JFH1/adpt1 RNA 4 h postinoculation in cells cultured at 20¡20 or 3¡3% O2or in cells that were transferred immedi-ately after virus addition from 20% to 3% O2(20¡3*). As shown inFig. 3C, viral RNA levels were similar in all three conditions, suggesting that virus entry is most likely not affected by low oxy-gen in our system.
These results were substantiated by transfecting the bicistronic reporter replicon LucCore-NSJFH1 and the reporter virus JcR2a as well as the JFH1 wild-type (wt) genome (Fig. 1) into Huh7-Lunet cells that were subsequently incubated at 20% or 3% O2for various time points posttransfection (p.t.). At 4 h p.t., luciferase values attained with the reporter constructs were reduced around 2-fold at 3% O2compared to values at 20% O2, whereas at 24 h p.t., a significant rise of about 4.5-fold was detected (Fig. 3D). Similarly, at 24 h p.t. of JFH1 wt RNA, Western blot analysis re-vealed a higher accumulation of NS5A protein (5- to 7-fold) (Fig. 3E) and the TCID50assay showed a significant increase (7- to 8-fold) in the production of infectious HCV (Fig. 3F) at 3% O2. However, at 48 h p.t., the 3% O2-related enhancement was signif-icantly lower for the transfected cells than for the infected cells. We hypothesized that this may be related to the different kinetics of HCV RNA replication in transfected cells compared to infected cells (37) or to stronger exhaustion of host factors in transfected cells than infected cells (38). In fact, a more detailed kinetics anal-ysis using different amounts of the bicistronic replicon LucCore-NSJFH1 showed that under low-oxygen conditions, RNA replica-tion enhancement reaches maximum levels 24 h p.t. and gradually declines thereafter (see Fig. S1B in the supplemental material). Overall, these results confirm that hypoxia facilitates HCV RNA replication during early stages of viral infection independent from virus entry.
We next investigated whether low oxygen tension has a posi-tive effect on HCV RNA stability/expression, which may contrib-ute to the enhancement of viral replication. Indeed, it has been reported that under hypoxia, the rate of cellular protein synthesis is reduced (39). Consistently, we found that upon transfection of Huh7-Lunet cells with a capped and polyadenylated reporter mRNA, expression of F-Luc was diminished at 3% O2compared to normoxia (data not shown). However, HCV RNA translation is IRES dependent, raising the question of whether low oxygen may favor IRES-mediated translation. For this, a nonreplicative bicis-tronic JFH1 reporter RNA (Luc-NSJFH1delGDD) (Fig. 1) was electroporated into Huh7-Lunet cells that were further cultured at 20% or 3% O2for various time points p.t. Interestingly, a signifi-cant impairment of luciferase activity was detectable with cells transferred to 3% O2right after transfection (20¡3) or with cells preincubated for 18 h at 3% O2prior to transfection (3¡3) (Fig. 3G). Furthermore, no increase in the HCV IRES-dependent ex-pression of luciferase was observed using a bicistronic reporter plasmid under infection conditions (40) at 3% O2(see Fig S1C in the supplemental material). Thus, enhanced HCV RNA
replica-tion at low O2concentrations is not due to elevated RNA transla-tion.
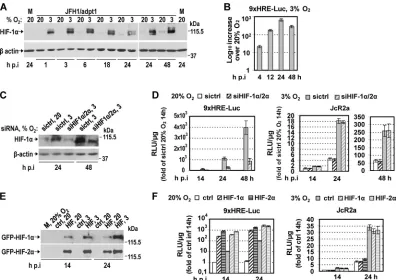
Low-oxygen-mediated enhancement of HCV replication during early infection is not facilitated by HIF-␣. Hypoxia-in-ducible factors (HIFs) are important transcription factors for cell adaptation to hypoxia (18) which are stabilized and acti-vated below 5% O2(41). The oxygen-regulated HIF-␣subunit (1␣/2␣/3␣) and the ubiquitous HIF-1interact as a complex with HRE-containing gene promoters. However, HIF-1␣ acti-vation under normoxia is a general phenomenon in infection with bacteria, protozoa, and pathogenic viruses (29). HCV in-duces HIF-1␣stabilization at late time points postinfection (42–44). Thus, we examined if the induction of HCV replica-tion, observed at 3% O2, was related to HIF activation. To this end, we performed the following experiments.
First, we confirmed that HIF-1␣was stabilized already 1 h after transferring JFH1/adpt1-infected Huh7.5 cells to 3% O2and re-mained at high levels up to 48 h p.i. (Fig. 4A). HIF activation at 3% O2(⬃20-fold) (Fig. 4B) was verified by measuring luciferase ac-tivity in cells transfected with the plasmid 9xHRE-Luc, infected with JFH1/adpt1, and then incubated at 20% or 3% O2(20¡20 or 20¡3). Consistent with the HIF-1␣protein (Fig. 4A), HRE activ-ity (see Fig. S2 in the supplemental material) was similar between HCV- and noninfected cells at early hours p.i., whereas at 72 h p.i., an⬃2-fold increase was detected in infected cells, in agreement with studies (42–44) showing HIF-1 activation at late stages of HCV infection.
Second, we evaluated the role of HIFs in the enhancement of HCV replication under low-oxygen conditions by silencing the two most well-studied HIF-␣isoforms, 1␣and 2␣(45). A mixture of two siRNAs (siHIF1␣/2␣) efficiently reduced HIF-1␣levels at 3% O2, as shown by Western blotting of JcR2a-infected cells (Fig. 4C), without affecting cell viability (see Fig. S3D in the supple-mental material). Also, silencing of HIF1␣/2␣ was confirmed upon cotransfection of the 9xHRE-Luc plasmid, as HRE activity was downregulated⬃5-fold (Fig. 4D, left). Surprisingly, silencing had no effect on the enhancement of HCV RNA replication at 3% O2(Fig. 4D, right). Third, we overexpressed HIF-1␣and -2␣(Fig. 4E), leading to upregulation of HRE comparable to that observed at 3% O2(Fig. 4F, left) and induction of HIF-regulated genes (see Fig. S3A in the supplemental material), albeit lower than that ob-served at 3% O2; however, this had no effect on HCV replication (Fig. 4F, right) or particle production (see Fig. S3B in the supple-mental material). Finally, we pharmacologically induced HIF sta-bility and/or activity (46) by treating Huh7.5 cells with CoCl2, desferrioxamine (DFO), or dimethyloxallyl glycine (DMOG). This enhanced HRE activity (see Fig. S3E, left, in the supplemental material) again had no effect on viral replication (see Fig. S3E, right, in the supplemental material). These results suggest that hypoxia-related enhancement of HCV replication, observed at the early stages of infection, occurs independently from HIF-1 and HIF-2 activation.
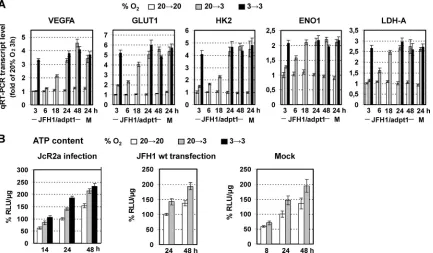
Low-oxygen-mediated enhancement of HCV replication is linked to hypoxia-induced reprogramming of cellular energet-ics.Since the activation of HIFs did not contribute to the detected upregulation of HCV replication at 3% O2, we examined global cellular gene expression changes in mock- and JFH1/adpt1-in-fected Huh7.5 cells using hypoxia signaling pathway-focused DNA microarrays. Cells cultured for 24 h at 3% O2showed (18, 47) an upregulation (by at least 2-fold) of several hypoxia-related
on November 7, 2019 by guest
http://jvi.asm.org/
genes (see Table S2 in the supplemental material), such as HIFs, directing transcriptional targets (VEGFA, EPO), genes encoding stress-related proteins, and genes involved in oxidative stress and reactive oxygen species (ROS) production, glucose transport (GLUT1, GLUT8) and metabolism (HK2, ENO1), cell growth, proliferation, and cell cycle regulation, including the FOS, MYCN, and KIT oncogenes. Notably, the respective transcript levels in uninfected and infected cells were similar. Moreover, qRT-PCR analysis of selected genes revealed an early response of transcrip-tional reprogramming to low oxygen (Fig. 5A) and confirmed a metabolic shift toward a higher rate of anaerobic glycolysis. Cell cycle distribution experiments using hydroxyurea (HU) treat-ment and fluorescence-activated cell sorting (FACS) analysis (see Table S3 in the supplemental material) also supported an acceler-ation of the cell cycle kinetics at 3% O2, consistent with previous studies (48,49). Interestingly, HCV replication enhancement at 3% O2was also observed in the HU-arrested cells, suggesting that
the effect on HCV is related neither to cell cycle acceleration nor to a specific cell cycle phase (data not shown).
Given that viral replication processes depend strongly on cell energetics, we assessed the effect of 3% O2on the intracellular levels of ATP in JcR2a-infected (Fig. 5B, left), JFH1 RNA-trans-fected (Fig. 5B, middle), and mock-transfected (Fig. 5B, right) Huh7 cells. Incubation of JcR2a-infected Huh7.5 cells at 3% O2 resulted in an increase of ATP levels by 40 to 45% (20¡3) or 50 to 80% (3¡3) at 24 and 48 h p.i. No significant difference in the ATP content was observed between infected and control cells up to 48 h p.i. (data not shown). Notably, the ATP ratio at 3% versus 20% O2was cell confluence dependent, most likely due to the develop-ment of pericellular hypoxia (50) in high cell densities (see Fig. S4 in the supplemental material).
To test the relationship between cell energetics and HCV rep-lication, we modulated ATP production of cells by feeding with high or low glucose, galactose, or glutamine, followed by incuba-FIG 4Hypoxia-mediated enhancement of HCV replication is not facilitated by HIF-1␣/2␣. (A and B) HIF-1␣is stabilized and HIFs are activated at 3% O2. (A) (Top) Western blot analysis of endogenous HIF-1␣protein using lysates of JFH1/adpt1-infected Huh7.5 cells incubated at 20% or 3% O2. M, mock-infected cells. (Bottom)-Actin served as a loading control. (B) HRE-dependent expression of F-Luc in Huh7.5 cells transfected with the 9xHRE-Luc construct, infected with JFH1/adpt1 (1 TCID50/cell) 18 h later, and subsequently cultured at 20% or 3% O2until harvest. Values were normalized to those obtained with cells cultured at 20% O2. (C and D) HIF-1␣and HIF-2␣silencing does not affect low-oxygen-mediated enhancement of HCV replication. Huh7.5 cells cotransfected with the 9xHRE-Luc construct (0.05g/4⫻104cells) and a mixture of siRNAs targeting HIF-1␣and HIF-2␣(20 nM each; siHIF1␣/2␣) or a control siRNA (40 nM) (sictrl) were inoculated with JcR2a (1 TCID50/cell) 18 h p.t. and further incubated at 20% or 3% O2. (C) (Top) Western blot analysis of endogenous HIF-1␣ protein. (Bottom)-Actin served as a loading control. (D) Effect of HIF-1␣/2␣knockdown on the following. (Left) HIF activation, as determined by HRE-dependent F-Luc activity. (Right) HCV replication, as determined by R-Luc activity. Mean values were normalized to the reporter activity measured in cells that were cultured at 20% O2. (E and F) HIF-1␣and HIF-2␣overexpression is not sufficient to enhance HCV replication. Huh7.5 cells cotransfected with the 9xHRE-Luc construct (0.05g/4⫻104cells) and one plasmid expressing GFP-HIF-1␣, GFP-HIF-2␣, or GFP (0.4g/4⫻104cells) were inoculated with JcR2a (1 TCID50/cell) 18 h p.t. and subsequently incubated at 20% or 3% O2. ctrl, cells transfected with the plasmid expressing GFP. Transfection efficiency was 60 to 70%. (E) HIF-1␣and -2␣overexpression as determined by GFP-specific Western blot analysis. (F) Effect of HIF-1␣or HIF-2␣overexpression on the following. (Left) HIF activation, as determined by HRE-dependent luciferase expression. (Right) HCV replication, as determined by R-Luc expression. Mean values were normalized to the reporter activity detected in cells cultured at 20% O2. For Western blot analysis, at each time, a representative experiment of at least two independent repetitions is shown. In all panels, bars represent mean values from at least three independent experiments in triplicate. Error bars indicate standard deviations.
Vassilaki et al.
on November 7, 2019 by guest
http://jvi.asm.org/
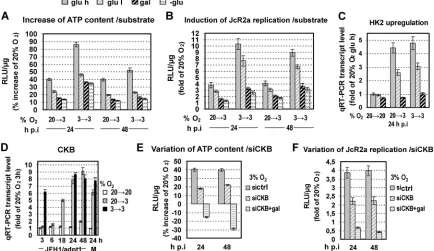
[image:6.585.94.492.65.345.2]tion of cells at different O2concentrations. As shown inFig. 6A andB, glucose reduction or substitution resulted in a gradual at-tenuation of the increase of both the intracellular ATP content and HCV replication observed under low oxygen conditions. qRT-PCR analysis confirmed a concomitant strong reduction of HK2 gene expression at 3% O2(Fig. 6C), suggesting an impairment of glycolysis. We therefore concluded that HCV replication en-hancement at 3% O2correlates primarily with an ATP increase due to the induction of anaerobic glycolysis.
Low-oxygen-mediated HCV replication enhancement is also linked to an increase in CKB-dependent ATP synthesis.Based on previous studies (51–53), another important metabolic route that can still produce ATP under low-oxygen conditions in Huh7 cells is catalyzed by creatine kinase (CK) (54). The brain-type iso-form of this enzyme (CKB) has been shown to promote HCV replication (55). Interestingly, qRT-PCR analysis demonstrated a significant (8- to 9-fold) increase in CKB mRNA at 3% O2, indi-cating directly that CKB is a hypoxia-induced gene (Fig. 6D). To determine the possible effect of this kinase on HCV replication induction at 3% O2, we silenced CKB expression. This reduced the ATP increase observed at 3% O2by 50% (Fig. 6E) and resulted in a lower enhancement of JcR2a RNA replication (Fig. 6F). Replace-ment of glucose by galactose simultaneously with CKB silencing resulted in lower intracellular ATP levels and viral replication at 3% than at 20% O2. These data suggest that under low-oxygen conditions, glycolysis and CKB are the main contributors to ATP production and play a key role in the observed enhancement of HCV replication.
At 3% O2, virus replication is induced in DMSO-treated
Huh7.5 cells.To investigate the effect of hypoxia on HCV repli-cation in a cellular system that better mimics thein vivosituation and to exclude effects caused by cell proliferation, we produced highly differentiated, growth-arrested Huh7.5 cells (Huh7.5dif) by DMSO treatment (see Fig. S5A and C, left, in the supplemental material) (56). These cells were readily infectible with HCV, albeit with lower efficiency than Huh7.5 cells (see Fig. S5B and C in the supplemental material).
Under hypoxia, HCV replication enhancement was also ob-served in Huh7.5difcells, detected early after infection (Fig. 7Aand B), and sustained throughout the 2-week observation period (Fig. 7A; see also Fig. S5D in the supplemental material). Consistently, an upregulation of the glycolysis-related genes GLUT1 and HK2 was detected at 20¡3 and 3¡3% O2(Fig. 7C).
Correlation of VEGF, HK2, LDH-A, and CKB with HCV RNA levels in liver biopsy specimens from chronically infected patients.In an attempt to correlate results obtained in cell culture with thein vivosituation, we obtained liver fine-needle biopsy specimens from 12 patients with liver fibrosis due to chronic HCV infection. Total RNA was extracted and subjected to qRT-PCR analysis for quantification of mRNA levels of the energy produc-tion-related genes HK2, LDH-A, and CKB as well as the hypoxic stress-responsive gene VEGFA. HCV positive-strand RNA was quantified by a branched DNA assay. Importantly, liver biopsy (LB) samples 10 to 12, containing the largest amounts of the cel-lular transcripts (Fig. 8A), also contained the largest HCV RNA amounts (Fig. 8B). Moreover, LB samples 1 to 6, containing the FIG 5Early induced reprogramming of cellular energetics by hypoxia. (A) Upregulation of VEGFA and GLUT1, HK2, ENO1, and LDH-A genes in JFH1/adpt1-infected Huh7.5 cells that were cultured at 20¡3 or 3¡3% O2relative to 20¡20% O2. Values were normalized to those obtained with cells kept at 20% O2and harvested 3 h p.i. For 3¡3 infected cells, values were measured only at 3, 24, and 48 h p.i. J, JFH1/adpt1-infected cells; M, mock-infected cells. (B) Three percent oxygen increases the ATP content of hepatoma cells. Intracellular ATP levels in JcR2a-infected Huh7.5 cells (1 TCID50/cell) (right), JFH1 RNA-electroporated Huh7-Lunet cells (10g RNA/4⫻106cells) (middle), or mock-transfected Huh7-Lunet cells (left). Cells were incubated as specified at the top and harvested at given time points. Mean values were normalized to the ones detected in cells cultured at 20¡20% O2and harvested 24 h postinfection/posttransfection. In all panels, bars represent mean values from at least three independent experiments in triplicate or quintuplicate (ATP content). Error bars indicate standard deviations.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.78.511.68.321.2]smallest amounts of the selected cellular genes, also contained the smallest HCV RNA amounts. Intermediate values of RNAs were detected in LB samples 7 to 9. Despite the limited number of clinical samples, these results argue for anin vivocorrelation
be-tween the degree of metabolic activity and response to hypoxic stress and the HCV replication level.
Possible role of oncogene induction for hypoxia-related en-hancement of HCV replication in Huh7.5 cells.ln addition to FIG 6Replacing the glycolytic substrate with oxidative ones, as well as CKB silencing, reduces low-oxygen-mediated HCV enhancement. (A to C) Huh7.5 cells preincubated at 20% or 3% O2were inoculated with JcR2a (1 TCID50/cell) and cultured at the given conditions in complete DMEM containing high glucose (glu h; 25 mM), low glucose (glu l; 5.56 mM), galactose instead of glucose (gal; 10 mM), or no glucose (-glu). (A) Bioluminescent measurement of intracellular ATP levels. Mean values (%) were normalized to ATP levels detected in cells cultured at 20¡20% O2. (B) JcR2a-derived luciferase activity expressed as fold induction relative to values obtained with cells cultured at 20¡20% O2. (C) qRT-PCR-based quantification of HK2 mRNA in cells treated as described above. Results are expressed as fold induction of gene expression relative to that in cells cultured at 20¡20% O2in DMEM with high glucose. (D) Upregulation of CKB expression at 3% O2. qRT-PCR analysis of CKB mRNA amounts in JFH1/adpt1-infected Huh7.5 cells cultured as specified on the right. Data were normalized to values obtained with cells cultured at 20¡20% O2and harvested 3 h p.i. For 3¡3 infected cells, values were measured only at 3, 24, and 48 h p.i. J, JFH1/adpt1-infected cells. M, mock-infected cells. (E and F) Huh7.5 cells transfected with an siRNA targeting CKB (siCKB) or a control siRNA (sictrl) (100 nM) were inoculated 18 h p.t. with JcR2a (1 TCID50/cell), supplied with DMEM containing either high glucose or galactose (gal), and incubated at 20% O2or transferred to 3% O2. (E) Intracellular ATP levels as determined by a bioluminescent assay. (F) JcR2a-derived luciferase activity. Values were normalized to those obtained with cells cultured at 20¡20% O2and transfected with the same siRNA. In all panels, bars represent mean values from at least three independent experiments in triplicate or quintuplicate (ATP content). Error bars indicate standard deviations.
FIG 7Three percent oxygen enhances HCV replication in Huh7.5difcells. (A) Effect of low oxygen tension on JcR2a-infected Huh7.5difcells that were incubated as specified at the top and harvested at various time points. Mean values are expressed relative to that obtained with cells cultured at 20¡20% O2. (B) Effect of low oxygen on Jc1 RNA replication as determined by quantification of intracellular positive-strand RNA copies. Values are expressed relative to those obtained at 20¡20% O224 h p.i. Numbers above the bars refer to ratios of RNA copies. 20¡3*, cells that were transferred immediately after virus addition from 20% to 3% O2. (C) Effect of 3% O2on GLUT1 and HK2 genes in Jc1-infected Huh7.5difcells. mRNA amounts were quantified by qRT-PCR and expressed relative to values obtained with cells cultured at 20¡20% O2and harvested 24 h p.i. M, mock-infected cells. In all panels, bars represent mean values from at least three independent experiments in triplicate. Error bars indicate standard deviations.
Vassilaki et al.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:8.585.75.509.69.320.2] [image:8.585.96.492.540.655.2]HIF, reprogramming of bioenergetics during hypoxia is also known to be affected by certain oncogenes and tumor suppressor genes (47, 57–62). Hypoxia-mediated upregulation of AKT1, MYC, FOS, H-RAS, SRC, BCR/ABL, and p53 has been reported to enhance glucose uptake and glycolysis through direct activation of related genes, including GLUT1, HK2, ENO1, and LDH-A. Since our results (see Table S2 in the supplemental material) suggested the upregulation of certain oncogenes (FOS, MYCN, and KIT) in HCV-infected cells under hypoxia, we investigated further the in-duction of selected oncogenes by using qRT-PCR. As shown in Fig. 9A, at 24 h of cell incubation at 20¡3 or 3¡3% O2, FOS, MYC, and SRC were upregulated 3- to 4-fold compared to levels after incubation at 20¡20% O2. Notably, mRNA levels in nonin-fected (mock) cells were similar to those in innonin-fected cells.
Expres-sion of AKT1, H-RAS, and BCR/ABL genes was unaffected (data not shown). However, consistent with previous studies (63), Western blot analysis revealed a significant increase of AKT phos-phorylation levels in HCV-infected Huh7.5 cells cultured at 3% O2compared to cells at 20% O2(Fig. 9B). Notably, AKT phos-phorylation was also induced early during infection of Huh7.5 cells at 20% O2(compare mock- and JFH1-infected cells), as pre-viously described for other flaviviruses (64,65), and this increase was higher at 3% O2, even after elimination of the infection-re-lated enhancement of AKT (63). Interestingly, in the nondividing Huh7.5difcells, FOS was also upregulated at 20
¡3 and 3¡3% O2 24 h p.i. (Fig. 9A) whereas MYC and SRC were not changed (data not shown). Moreover, FOS and SRC were not induced upon HIF-1␣/2␣overexpression (see Fig. S6A in the supplemental ma-FIG 8Analysis of human liver biopsy specimens reveals a correlation between elevated HCV RNA amounts and increased expression of genes involved in anaerobic metabolism. (A) qRT-PCR quantification of HK2, LDH-A, CKB, and VEGF mRNA amounts in total RNA isolated from 12 liver samples (LB 1 to 12) from patients with chronic hepatitis C. Bars represent mean values from three independent repetitions, and error bars indicate standard deviations. YWHAZ mRNA levels were used for normalization. (B) HCV positive-strand RNA copy levels from the same samples, as quantified using the branched DNA assay. Values were normalized to those obtained with sample LB 1.
FIG 9Low-oxygen-mediated oncogene upregulation is linked to enhancement of HCV replication. (A) Upregulation of FOS, MYC, and SRC genes in JFH1/adpt1 (J)-infected Huh7.5 or Huh7.5difcells. M, mock-infected cells. (B) Elevated levels of AKT phosphorylation in infected Huh7.5 cells cultured under hypoxia. Western blot analysis of endogenous levels of phosphorylated (top) and total (bottom) AKT. M, mock-infected cells. A representative experiment of two independent repetitions is shown. (C) JUN-FOS overexpression enhances intracellular ATP levels and HCV replication. Huh7.5 cells cotransfected with the MMP1-Luc (0.05g/4⫻104cells)- and the JUN-FOS (0.4g/4⫻104cells)-encoding plasmids or the respective vector (control) were inoculated 18 h p.t. with JcR2a (1 TCID50/cell) and further incubated at 20% O2for 24 h. JUN-FOS overexpression enhances TRE-dependent expression (left), increases HCV RNA replication (middle), and results in a detectable upregulation of the intracellular ATP (right). For all panels, values were normalized to those obtained with control-transfected cells. (D) AKT inhibition reduces hypoxia-mediated HCV enhancement. Huh7.5 cells were inoculated with JcR2a and 4 h later were untreated or treated with the AKT inhibitor VIII (Akti-1/2) at the following concentrations: C1, 2.5⌴; C2, 5⌴; C3, 10⌴. After 24 h of incubation of cells at 20% or 3% O2, luciferase activities were determined. Values were normalized to those obtained with untreated cells cultured at 20% O2. In all panels, bars represent mean values from at least three independent experiments in triplicate or quintuplicate (ATP content). Error bars indicate standard deviations.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:9.585.111.473.422.619.2]terial). Therefore, we hypothesized that certain hypoxia-regulated oncogenes can operate separately from HIFs to modulate cell en-ergetics favoring HCV RNA replication.
To challenge this hypothesis, we evaluated the role of FOS and AKT in HCV replication. FOS was chosen because its expression was upregulated in both normal and DMSO-treated Huh7.5 cells. As shown inFig. 9C, overexpression of a JUN-FOS tethered dimer in cells cultured at 20% O2enhanced virus replication⬃2-fold (middle panel), concomitant with an⬃20% elevation of intracel-lular ATP levels (right panel). As expected, the consensus tissue plasminogen-activator response element (TRE) within the human matrix metalloproteinase 1 (MMP1) promoter, known to be rec-ognized by JUN-FOS, was strongly activated (⬃10-fold) (left panel). Concerning AKT, its role in the induction of glycolysis is well documented. Treating Huh7.5 cells with the AKT inhibitor VIII (AKTi-1/2) reduced JcR2a replication enhancement at 3% O2 (Fig. 9D), even at an inhibitor concentration (C1 inFig. 9D) that does not significantly alter cellular ATP content (data not shown). These results suggest that activator protein 1 (AP-1) (FOS/JUN or JUN/JUN dimers) and AKT are factors responsible for the en-hancement of HCV replication under low-oxygen conditions.
In support of this assumption, incubation of cells at 12% O2, a condition prevailing in the liver, did not stabilize or activate HIF in Huh7.5 cells (see Fig. S6B and C, respectively, in the supple-mental material), consistent with HIF activation at oxygen levels lower than 5% (vol/vol) (41). However, transcription of glycoly-sis-related genes and CKB (see Fig. S6D, left, in the supplemental material) and expression of the FOS oncogene (see Fig. S6D, right, in the supplemental material) were enhanced. Consistent with these results, JcR2a replication (see Fig. S6E in the supplemental material) and intracellular ATP levels (see Fig. S6F in the supple-mental material) were also enhanced at 12% O2, albeit with de-layed kinetics and to a lower extent compared to levels at 3% O2. To summarize, certain oncogenes, including FOS and AKT1, in combination with other cellular factors, such as CKB, modulate cell energetics and, thus, HCV RNA replication.
DISCUSSION
Recent studies have shown that oxygen tension exerts a significant effect on the replication of several viruses. In general, hypoxia restricts the replication of viruses that naturally infect tissues ex-posed to ambient oxygen (simian virus 40, adenovirus) (25,26) and induces the propagation of viruses that naturally target tissues exposed to low oxygen concentrations (vesicular stomatitis virus, herpesviruses, human immunodeficiency virus, parvovirus B19) (20–24).
To date, HCVin vitroinfection has been studied exclusively under atmospheric oxygen conditions using mostly Huh7 and derivatives, although the liver microenvironment is hypoxic (14, 15) and hepatocytes are highly differentiated (66). Here, we inves-tigated HCV replication efficiency at 3% (vol/vol) O2to mimic the oxygen tension sensed by most hepatocytes in the liver or at 12% O2, which is present around the liver portal vein. Our results reveal the following.
Low oxygen tension (3% O2) enhances HCV replication in
cultured cells.This was most pronounced by preincubation of cells at 3% O2prior to infection (3¡3% O2), arguing that a pre-existing hypoxia-induced cellular factor(s) promotes HCV repli-cation. This enhancement was due to an RNA replication increase at early hours of infection, whereas virus entry, RNA translation,
assembly, and release were not detectably affected (Fig. 2and3). Several mechanisms responsible for replication enhancement can be envisaged.
Low-oxygen-mediated increase of HCV replication is inde-pendent of HIF-1␣/2␣activation.HIFs are fundamental control transcription factors of the cellular metabolic state under low-oxygen conditions (18). Although low-oxygdependent en-hancement of HCV replication may reflect HIF-1-mediated alter-ations of cellular homeostasis, overexpression and silencing studies revealed that HIF-1␣/2␣activation did not correlate with the hypoxia-related increase of HCV replication observed at the early stages of infection (Fig. 4; see also Fig. S3 in the supplemental material). In agreement, chemically induced hypoxia failed to el-evate HCV replication, whereas 12% O2, which fails to activate HIF-1␣(41) (as well as HIF-2␣), positively affected HCV replica-tion (see Fig. S6 in the supplemental material). Notably, HIF-1␣ activation from HCV at late stages of viral infection has been re-ported under normoxia and has been linked to mitochondrial dysfunction and oxidative stress caused by HCV proteins (42–44). In agreement with these studies, we also detected an increase in HIF-1␣activation, but only 72 h p.i. (see Fig. S2 in the supplemen-tal material). Interestingly, a HIF-1-independent increase of ge-nome replication under hypoxic conditions has been found also for B19V (67).
Low-oxygen-mediated increase in HCV replication in cul-tured cells is directly linked to cellular energetic changes. Hyp-oxia is associated with an adaptive cell metabolic reprogramming, including a shift in glucose metabolism from oxidative phosphor-ylation to anaerobic glycolysis and lactic acid production (18,47). Huh7.5 cells, like most cancer cells, exhibit high rates of glycolysis and lactate production under normoxia (Warburg effect) (58,62) and even higher rates under hypoxia conditions (Pasteur effect) (68). For highly proliferating cells, the advantage of such a meta-bolic switch is to combine energy (ATP) from enhanced glycolysis with production of nutrients/intermediates for cell growth and division (69). Oncogenes and tumor suppressor proteins are known to regulate this phenomenon in addition to HIF activation (60).
Indeed, transcriptome analysis of Huh7.5 cells confirmed that several oncogenes, as well as genes involved in glucose uptake (GLUT1) and anaerobic glycolysis (LDH-A, HK2, ENO1), were upregulated at 3% O2(Fig. 5A; see Table S2 in the supplemental material). A comparison of HCV-infected and mock cells up to 24 h p.i. suggested that during the early stages of infection, HCV does not affect the cellular transcription profile, as previously shown (70). As expected, enhanced glucose uptake and use compensated for the ATP levels under low-oxygen conditions (Fig. 5B) and promoted cell proliferation (see Table S3 in the supplemental ma-terial), consistent with previous reports (48,49).
Interestingly, the ATP gain correlated well with the low-oxy-gen-mediated HCV replication enhancement and was directly linked to hypoxia-induced glycolysis, as revealed by experiments comprising the use of different energy substrates in the culture medium of infected Huh7.5 cells (Fig. 6AtoC). Consistently, previous studies have directly implicated two glycolytic enzymes in the regulation of HCV replication (71, 72), hexokinase III, which is transcriptionally activated by both HIF and MYC under hypoxia, and pyruvate kinase, which is enhanced in a HIF-inde-pendent manner (73–75). Thus, hypoxia-mediated upregulation of glycolysis represents an HCV replication-promoting metabolic
Vassilaki et al.
on November 7, 2019 by guest
http://jvi.asm.org/
modification. Moreover, oxidative phosphorylation, although ac-tive at 3% O2, had no role in the observed gain of energy, as detected by oligomycin treatment (mitochondrial ATP synthase inhibitor) of the infected cells (data not shown).
Oncogene induction may be implicated in hypoxia-related HCV replication enhancement.Oncogenes and tumor suppres-sor proteins, such as AKT, AP-1 (FOS/JUN or JUN dimers), MYC, SRC, and p53, are known to coordinate glycolysis under both anaerobic and aerobic (Warburg) conditions (47,58–62). Fur-thermore, induction of oxidative stress- and ROS-related genes (76) or other HIF-1-independent pathways, such as that of RAS and intracellular Ca2⫹effector pathways (77,78), may modulate oncogene expression under certain cellular conditions. Inasmuch as our results indicated a HIF-independent hypoxia-induced HCV replication, we hypothesized that certain oncogenes and/or tumor suppressor genes may play a critical role in cellular changes that favor virus replication enhancement under low-oxygen con-ditions.
Indeed, several oncogenes were found to be upregulated (Fig. 9A; see Table S2 in the supplemental material) or activated (Fig. 9B) at 3% O2, together with oxidative stress- and ROS-related genes (see Table S2 in the supplemental material) that may con-tribute to oncogene induction. Among the induced oncogenes, FOS, MYC, SRC, and AKT have been already shown to directly activate glucose-related transporter and glycolytic gene promoters independently of HIFs (47,57–62) and, in addition, have been reported to stimulate HCV replication through either association with viral nonstructural proteins or other ways of action not com-pletely defined (71,79–83). In fact, our results confirmed the HIF-independent transcriptional activation of FOS and SRC (see Fig. S6A and D in the supplemental material) under hypoxia and showed that JUN-FOS overexpression was sufficient to increase HCV replication and intracellular ATP (Fig. 9C). In contrast, no ATP increase was detected during HIF overexpression, even though the expression of certain glycolytic enzymes was affected (see Fig. S3A and C in the supplemental material), or upon chem-ical-induced hypoxia (data not shown), whereas no decrease was observed after HIF-1␣/2␣silencing (see Fig. S3D in the supple-mental material). Moreover, the enhanced FOS expression in the DMSO-differentiated Huh7.5 cells at 3% O2(Fig. 9A) argues for a cellular proliferation-unrelated role of this oncogene in HCV rep-lication enhancement. In addition, although expression of the AKT1 gene was found unaffected at 3% O2in infected cells, AKT phosphorylation/activation was increased under hypoxia (Fig. 9B). Interestingly, the addition of the AKT inhibitor VIII (Akti-1/2) had a negative effect on the HCV replication enhancement at 3% O2(Fig. 9D), even at concentrations that do not significantly alter intracellular ATP, implicating AKT activity in the hypoxia-mediated increase of HCV propagation.
Finally, supportive of our hypothesis that oncogenes are cru-cial for the hypoxia-mediated HCV enhancement, our results from 12% O2show that although, as expected (41), 12% O2does not support HIF stabilization/activation, it enhances HCV repli-cation and, concomitantly, the expression of glycolytic enzymes and oncogenes (see Fig. S6B to E in the supplemental material).
Thus, at least in hepatoma cells, oncogenes appear to be im-portant players in the hypoxia-driven cellular metabolic shift, en-hancing glycolysis and ATP production and, concomitantly HCV replication in a HIF-independent manner. Interestingly,
hypoxia-mediated induction of oncogenes has been reported also for pri-mary liver cells (49).
CKB induction contributes to hypoxia-related HCV replica-tion enhancement.Creatine kinase covers the immediate ener-getic needs of the cell by catalyzing the reversible reaction of phos-phocreatine (PCr) and ADP to creatine and ATP (54). To date, CKB upregulation under hypoxia has been reported in a three-dimensional hepatocellular culture model (53) as well as in the rat liver (51), and CKB overexpression in the liver of transgenic mice has been shown to stabilize the energy metabolism protecting against low-oxygen stress (52). Moreover, CKB, the main isotype expressed in the liver (84), is known to regulate HCV replication by forming a complex with NS3-4A and locally delivering the energy required for the helicase activity (55).
In our study, the CKB mRNA levels were strongly induced at 3% O2(Fig. 6D), and this contributed significantly to both HCV replication enhancement and higher intracellular ATP levels at 3% O2(Fig. 6EandF), as revealed by silencing experiments. In-terestingly, CKB upregulation observed at 12% O2argues for a HIF-independent transcriptional activation under low-oxygen conditions (see Fig. S6D in the supplemental material). Onco-gene-mediated regulation of the CKB promoter has been sug-gested only for the case of p53 (repression) (54). However, the details of CKB modulation by oxygen tension remain elusive. Sp1 and cyclic AMP (cAMP)/protein kinase A (PKA) response ele-ments of CKB promoter may be involved, as shown for glucose metabolism genes during low-oxygen conditions (73,85).
Overall, our results indicate that, in addition to glycolysis, CKB may act to enhance the intracellular ATP levels under low-oxygen conditions in a HIF-independent manner. Thus, oncogene-medi-ated glycolysis and CKB activation are the main contributors to ATP production, at least in Huh7.5 cells, and the observed hypoxia-related enhancement of the HCV replication is directly linked to the activity of those processes. Therefore, it is tempting to speculate that HCV-infected patients may benefit by combin-ing the current therapy with the use of glycolysis or CKB inhibi-tors, used already in cancer treatment (62,86,87).
Low oxygen tension promotes a sustained induction of HCV infection in Huh7difcells.A low-oxygen-mediated increase in
HCV replication was also observed in Huh7.5difcells (differenti-ated by DMSO) that better mimic the conditions of HCV infec-tionin vivo(56). Interestingly, HCV replication enhancement at 3% O2was maintained up to 2 weeks in this system (Fig. 7), con-sistent with its ability to support long-term HCV replication. Moreover, this system confirmed that hypoxia selectively affects the establishment of HCV replication, since preincubation of cells at 3% O2determined the level of viral RNA throughout a 2-week observation period.
Importantly, bioenergetics reprogramming and HCV replica-tion enhancement in the nondividing Huh7.5difcells at 3% O2can be exclusively linked to hypoxia adaptation, excluding confound-ing effects caused by increased cell proliferation (47,58,62,69, 88), similarly to the HU-arrested Huh7.5 cells.
Increased HCV RNA levels correlate with increased activity of anaerobic metabolism in liver biopsy specimens.Liver cells regulate their metabolism and structure in response to oxygen tension (16,19). A correlation between low oxygen and increased glycolysis prevails in the liver (16,17). Also, at least in mouse liver, CKB overexpression stabilizes energy metabolism upon hypoxic stress (51,52).
on November 7, 2019 by guest
http://jvi.asm.org/
Analyses of liver biopsy specimens derived from HCV patients with liver fibrosis due to chronic infection showed a significant association between the expression levels of anaerobic energy me-tabolism (HK2, LDH-A, CKB)- and hypoxia (VEGFA)-related genes and the HCV RNA load (Fig. 8). These results are in perfect agreement with our cell culture data, indicating a strong link among hypoxia, increased glycolysis rate and CKB activity and higher HCV RNA replication.
In summary, we report that HCV infection of Huh7-derived cell lines cultured under low-oxygen conditions (3% or 12%) pro-motes the early stages of HCV infection/replication compared to standard cell culture conditions (20% O2). This enhancement di-rectly correlated with hypoxia-induced changes in cell energetics, including an increase in anaerobic glycolysis and CKB expression that leads to elevated ATP production. Surprisingly, activation of HIF-1␣/2␣was dispensable for this enhancement, whereas acti-vation of certain oncogenes appears to be the central event in the HCV RNA replication induction. This study opens new possibil-ities in optimizing conditions forin vitropropagation of HCV and defining novel therapeutic targets.
ACKNOWLEDGMENTS
We are grateful to T. Wakita for the JFH1 isolate, C. Rice for Huh7.5 cells and the 9E10 antibody, R. Hernandez-Alcoceba for the p9xHRE-Luc plas-mid, and L. Bakiri for the JUN-FOS andMMP1-luc vectors. We acknowl-edge H. Boleti for scientific advice and editing as well as E. Xingi for technical assistance in the microscopy experiments.
This work was funded by the Hellenic General Secretariat of Research and Technology and partially by Institute Pasteur, Paris, France. Work in the laboratory of R.B. was supported by grants from the Deutsche For-schungsgemeinschaft (FOR1202, TP1, and SFB/TRR77, TP A1).
REFERENCES
1.Levrero M.2006. Viral hepatitis and liver cancer: the case of hepatitis C. Oncogene25:3834 –3847.
2.Seeff LB.2002. Natural history of chronic hepatitis C. Hepatology36:S35– S46.
3.Feld JJ, Hoofnagle JH.2005. Mechanism of action of interferon and ribavirin in treatment of hepatitis C. Nature436:967–972.
4.Moradpour D, Penin F, Rice CM.2007. Replication of hepatitis C virus. Nat. Rev. Microbiol.5:453– 463.
5.Tang H, Grise H.2009. Cellular and molecular biology of HCV infection and hepatitis. Clin. Sci. (Lond.)117:49 – 65.
6.Eng FJ, Walewski JL, Klepper AL, Fishman SL, Desai SM, McMullan LK, Evans MJ, Rice CM, Branch AD.2009. Internal initiation stimulates production of p8 minicore, a member of a newly discovered family of hepatitis C virus core protein isoforms. J. Virol.83:3104 –3114. 7.Vassilaki N, Mavromara P. 2009. The HCV ARFP/F/core⫹1 protein:
production and functional analysis of an unconventional viral product. IUBMB Life61:739 –752.
8.Lohmann V, Korner F, Koch J, Herian U, Theilmann L, Bartenschlager R.1999. Replication of subgenomic hepatitis C virus RNAs in a hepatoma cell line. Science285:110 –113.
9.Wakita T, Pietschmann T, Kato T, Date T, Miyamoto M, Zhao Z, Murthy K, Habermann A, Krausslich HG, Mizokami M, Bartenschlager R, Liang TJ.2005. Production of infectious hepatitis C virus in tissue culture from a cloned viral genome. Nat. Med.11:791–796.
10. Lindenbach BD, Evans MJ, Syder AJ, Wolk B, Tellinghuisen TL, Liu CC, Maruyama T, Hynes RO, Burton DR, McKeating JA, Rice CM.
2005. Complete replication of hepatitis C virus in cell culture. Science
309:623– 626.
11. Pietschmann T, Kaul A, Koutsoudakis G, Shavinskaya A, Kallis S, Steinmann E, Abid K, Negro F, Dreux M, Cosset FL, Bartenschlager R.
2006. Construction and characterization of infectious intragenotypic and intergenotypic hepatitis C virus chimeras. Proc. Natl. Acad. Sci. U. S. A.
103:7408 –7413.
12. Kaul A, Woerz I, Meuleman P, Leroux-Roels G, Bartenschlager R.2007.
Cell culture adaptation of hepatitis C virus and in vivo viability of an adapted variant. J. Virol.81:13168 –13179.
13. Ashfaq UA, Khan SN, Nawaz Z, Riazuddin S. 2011. In-vitro model systems to study hepatitis C virus. Genet. Vaccines Ther.9:7.
14. de Groot H, Noll T.1987. Oxygen gradients: the problem of hypoxia. Biochem. Soc. Trans.15:363–365.
15. Fleckenstein W, Weiss C.1984. Ein neues Gewebe-pO2-Messverfahren zum Nachweis von Mikrozirkulationsstorungen. Focus Med. Hochschule Lubeck.1:74 – 84.
16. Jungermann K, Kietzmann T.2000. Oxygen: modulator of metabolic zonation and disease of the liver. Hepatology31:255–260.
17. Torre C, Perret C, Colnot S.2010. Molecular determinants of liver zonation. Prog. Mol. Biol. Transl. Sci.97:127–150.
18. Semenza GL.2011. Oxygen sensing, homeostasis, and disease. N. Engl. J. Med.365:537–547.
19. Martinez I, Nedredal GI, Oie CI, Warren A, Johansen O, Le Couteur DG, Smedsrod B.2008. The influence of oxygen tension on the structure and function of isolated liver sinusoidal endothelial cells. Comp. Hepatol.
7:4.
20. Aghi MK, Liu TC, Rabkin S, Martuza RL.2009. Hypoxia enhances the replication of oncolytic herpes simplex virus. Mol. Ther.17:51–56. 21. Connor JH, Naczki C, Koumenis C, Lyles DS.2004. Replication and
cytopathic effect of oncolytic vesicular stomatitis virus in hypoxic tumor cells in vitro and in vivo. J. Virol.78:8960 – 8970.
22. Haque M, Davis DA, Wang V, Widmer I, Yarchoan R.2003. Kaposi’s sarcoma-associated herpesvirus (human herpesvirus 8) contains hypoxia response elements: relevance to lytic induction by hypoxia. J. Virol.77: 6761– 6768.
23. Pillet S, Le Guyader N, Hofer T, NguyenKhac F, Koken M, Aubin JT, Fichelson S, Gassmann M, Morinet F.2004. Hypoxia enhances human B19 erythrovirus gene expression in primary erythroid cells. Virology327: 1–7.
24. Polonis VR, Anderson GR, Vahey MT, Morrow PJ, Stoler D, Redfield RR.1991. Anoxia induces human immunodeficiency virus expression in infected T cell lines. J. Biol. Chem.266:11421–11424.
25. Riedinger HJ, van Betteraey M, Probst H.1999. Hypoxia blocks in vivo initiation of simian virus 40 replication at a stage preceding origin un-winding. J. Virol.73:2243–2252.
26. Shen BH, Hermiston TW. 2005. Effect of hypoxia on Ad5 infection, transgene expression and replication. Gene Ther.12:902–910.
27. Blight KJ, McKeating JA, Rice CM.2002. Highly permissive cell lines for subgenomic and genomic hepatitis C virus RNA replication. J. Virol.76: 13001–13014.
28. Friebe P, Boudet J, Simorre JP, Bartenschlager R.2005. Kissing-loop interaction in the 3=end of the hepatitis C virus genome essential for RNA replication. J. Virol.79:380 –392.
29. Werth N, Beerlage C, Rosenberger C, Yazdi AS, Edelmann M, Amr A, Bernhardt W, von Eiff C, Becker K, Schafer A, Peschel A, Kempf VA.
2010. Activation of hypoxia inducible factor 1 is a general phenomenon in infections with human pathogens. PLoS One 5:e11576. doi:10.1371 /journal.pone.0011576.
30. Kato T, Furusaka A, Miyamoto M, Date T, Yasui K, Hiramoto J, Nagayama K, Tanaka T, Wakita T.2001. Sequence analysis of hepatitis C virus isolated from a fulminant hepatitis patient. J. Med. Virol.64:334 – 339.
31. Schmitt M, Scrima N, Radujkovic D, Caillet-Saguy C, Simister PC, Friebe P, Wicht O, Klein R, Bartenschlager R, Lohmann V, Bressanelli S.2011. A comprehensive structure-function comparison of hepatitis C virus strain JFH1 and J6 polymerases reveals a key residue stimulating replication in cell culture across genotypes. J. Virol.85:2565–2581. 32. Vassilaki N, Friebe P, Meuleman P, Kallis S, Kaul A, Paranhos-Baccala
G, Leroux-Roels G, Mavromara P, Bartenschlager R.2008. Role of the hepatitis C virus core⫹1 open reading frame and core cis-acting RNA elements in viral RNA translation and replication. J. Virol.82:11503– 11515.
33. Mylonis I, Chachami G, Samiotaki M, Panayotou G, Paraskeva E, Kalousi A, Georgatsou E, Bonanou S, Simos G.2006. Identification of MAPK phosphorylation sites and their role in the localization and activity of hypoxia-inducible factor-1␣. J. Biol. Chem.281:33095–33106. 34. Bakiri L, Matsuo K, Wisniewska M, Wagner EF, Yaniv M. 2002.
Promoter specificity and biological activity of tethered AP-1 dimers. Mol. Cell. Biol.22:4952– 4964.
35. Vassilaki N, Boleti H, Mavromara P.2008. Expression studies of the Vassilaki et al.