Copyrighti 1975 AmericanSociety for Microbiology Printed in U.S.A.
Structure, Subunit
Composition, and Molecular Weight
of
RD-114
RNA1
HSING-JIEN KUNG, JAMES M. BAILEY, NORMAN DAVIDSON,* MARGERY 0. NICOLSON,
AND ROBERT M. McALLISTER
Department ofChemistry, California Instituteof Technology, Pasadena, California91125,*andDepartment of Pediatrics, University of Southern California Schoolof Medicine, Childrens Hospital ofLosAngeles,Los
Angeles, California90054 Receivedforpublication9April 1975
The properties and subunit composition of the RNA extracted from RD-114
virions have been studied. The RNA extracted from the virion hasa
sedimenta-tion coefficient of 52S in a nondenaturing aqueous electrolyte. The estimated
molecular weight by sedimentation in nondenaturing and weakly denaturing media is in the range 5.7 x 106 to 7.0 x 106. By electron microscopy, under
moderately denaturing conditions, the 52S molecule is seento be an extended
single strand withacontourlength of about 4.0,umcorrespondingtoamolecular
weight of 5.74 x 106.Itcontainstwocharacteristic secondarystructurefeatures:
(i)acentral Y- orT-shaped structure(the rabbit ears) withamolecularweightof
0.3 x 106; (ii) two symmetrically disposed loops on each side of and at equal distance from the center. The 52Smolecule consists oftwohalf-size molecules, with molecular weight 2.8 x 106, joined together within the central rabbit ears
feature. Melting of the rabbit ears with concomitant dissociation of the 52S
molecule into subunits, has been caused by eitheroneoftwostrongly denaturing
treatments: incubation in a mixture of CH3HgOH and glyoxal at room
temperature, or thermal dissociation in a urea-formamide solvent. When
half-size moleculesarequenched from denaturingtemperatures, a newoff-center
secondary structure feature termed the branch-like structure is seen. The
dissociation behavior of the 52S complex and the molecular weight of the subunits have been confirmed by gel electrophoresis studies. The loopstructures
meltatfairly lowtemperatures;thedissociation of the52S molecule into itstwo
subunits occurs ata highertemperature correspondingtoa basecomposition of
about 63% guanosine plus cytosine. Polyadenylic acid mapping by electron
microscopy shows that the 52S molecule contains two polyadenylic acid
segments,oneateach end. It thusappearsthat52SRD-114RNA consistsof two
2.8 x 106 dalton subunits, eachwith a characteristic secondary structure loop,
andjoinedatthe 5' endstoformtherabbitearssecondarystructurefeature. The
observations are consistent with but do notrequiretheconclusion that the two
2.8 x 106dalton subunits of52SRD-114 RNA areidentical.
The organization of RNA extracted from RNAtumor viruseshas been
extensively
stud-iedduring thepast few years. Inmostinstances, the principal RNA species isolated from the virion is a complex sedimenting at 60 to 70S with a molecular weight of approximately 107. Upon exposure to denaturing conditions, this complex dissociates into a major component with a sedimentation coefficient of about 35S (and a molecular weight of2.5 x 106 to 3.3 x106) and into severalsmall(4 to10S) species.
IContribution no.5076 fromtheDepartment of
Chemis-try, California Institute of Technology, Pasadena, Calif. 91125.
In earlier papers we described our prelimi-nary electron microscope characterization of total RNA from the endogenous feline C type virus, RD-114 (9, 10). We
reported
that the major RNA component, when mounted for electronmicroscopyby procedures which extend the RNAsufficiently well fortracing, is a mol-ecule of 3.7gm
contourlength,
corresponding
to a molecularweight
of 5.0 x 10.. Further-more, these molecules all contained a charac-teristic Y- orT-shaped
secondary
structurefeature near the middle ofthe molecule. Such anobservation isconsistent with the
hypothesis
that all 5 x 101 dalton moleculesare identical in sequence. However, the molecularweight
of397
on November 10, 2019 by guest
http://jvi.asm.org/
this molecule is quite high in comparisontothat of the major RNA component of other RNA tumor viruses. The presence of the characteristic secondary structure feature at the middle of the molecule therefore raises the possibility that the 5.0 x 106 dalton molecule is not a con-tinuous polynucleotide chain but is instead two approximately 2.5 x 106 dalton molecules joined by base pairing within the central
sec-ondarystructure feature.
We were initially inclined to reject this hy-pothesis because in electron microscope spread-ings ofRNA molecules exposed to rather stongly denaturing conditions, we were able to identify full length (3.7 um) molecules in which the secondary structure feature appeared to be absent. We stated at the time that we felt that this evidence was notconclusivesince only 50% ofthe full length molecules were in this class.
We have accordingly continued these studies and searched for denaturing conditions which would either denature the secondary structure feature in all molecules or cause dissociation into smaller subunits. Our further electron microscope, sedimentation, and gel electropho-resis investigations of the structure subunit composition ofRD-114 RNA are reported here.
MATERIALS AND METHODS
Virus and RNApreparations. SindbisvirusRNA was prepared as previously described (6). RD-114 virus was grown as described by Filbert et al. (3). Viruses werepurifiedby isopycnic banding ina 24 to 48% sucrose gradient in NTE buffer (0.1 M NaCl, 0.01 M Tris, pH 7.2, 0.001 M EDTA) in an SE50.1 tubeat 44Krpm for3hat 4C. Virus fractionswere
pooled and treated with self-digested Pronase (-500
/g/ml)
in the presence of 0.5% sodium dodecyl sul-fate for 30 minat 37 C. The solution was then ad-justedto 1%sodiumdodecyl sulfate and 1% mercapto-ethanol. After repeated phenol extraction, the RNA samples were ethanol precipitated and resuspended in -100IAl
ofNTEsolution. In several experiments, the Pronase step was omitted and identical results were obtained. Rous sarcoma virus was a gift from Peter Vogt. HeLa 28S rRNA was generously provided by James Casey.Sedimentation. (i) NTE-sucrose gradient. A so-lution (100A) of phenol-extracted 3H-labeledRD-114 RNA (inNTE)waslayered directly onto a5-mi 10 to 30%sucrose gradient in NTE buffer. Centrifugation wascarried out in anSW50.1 rotor at 45 K rpm at 4 C for1.75 h.Sindbis RNA and 28S HeLa rRNA were run inparallel as externalmarkers.
(ii) Glyoxal-sucrose gradient. The phenol-extracted "H-labeled RD-114 RNA was dialyzed against 1 M glyoxal in 0.01 M phosphate buffer, pH 7.0, for 1 h at 37C, then dialyzed against 0.1 M glyoxalinthe samebuffer for approximately 30min at 4 C, all aspreviously described (6, 10). The sample thus treated was sedimented through a 10 to 30%
sucrosegradient in the presence of 0.1 Mglyoxal, 0.01 Mphosphate buffer, pH 7.2,at 4C,45Krpm for 5 h in an SW50.1 rotor. The peak fractionswereused for electron microscope studies. Sindbis and Rous sar-coma virus RNAs weresimilarly treated and run in parallel as external markers.
(iii) Low salt-sucrosegradient. Phenol-extracted RD-114RNA sampleswereincubated in 50% formam-ideand 50% NTE at 37 C for 10 min. This treatment wasdesigned to expose the hidden nicks of the RNA complex so as to give better fractionation. After removal of the formamide by dialysis at 4C, RNA samples were loadedonto a 10 to 30% sucrosegradient containing 1 mM Tris, 0.2 mM EDTA, pH 7.2. Centrifugationwasdonein anSW50.1rotor at 4C,41 K rpmfor 3.5h. Peak fractionswerepooledforother studies. Rous sarcoma virus 60 to 70S RNA was similarly treated andruninparallel.
Electron microscopy.(i) Drop andhigh tempera-ture spreading. In the present study, both the standard dish-spreading technique described by
Davisetal. (1) andadrop-spreadingtechniquewere applied. The latter is a modification ofthe method described by Inmanand Schnos (7). This technique requiresonly 1 to 5 ng of RNAsample perspreading and is thus usefulfor studying small quantities and nucleicacids. Itisalso convenientinthat it allowsa spreading tobe performedat auniformhigh tempera-tureinan oven.A Teflonblock (10by10 cmby1.2cm) which contains nine evenly spaced indentations (1.9 cmindiameter and0.1cm deep)wasprepared. One dropofhypophase (-0.9 ml)wasplacedonone ofthe indentations. A Pasteur pipette with the narrow end sealed was inserted at an angle of 60C into the hypophase. Five microliters of the spreading solution was then applied through the outer surface of the narrowend of the Pasteur pipette onto thehypophase.
Samples were picked up by touching a parlodion-coatedgridtothe surfaceofthedrop within30 safter
spreading. Thegridwas then rinsed in 95%ethanol and rotaryshadowed with platinum-palladium alloy.
Forhigh temperature spreading, the Teflon block, the Pasteur pipette, the micropipettes, and the hypo-phase solution were allpre-equilibratedinanoven at the desired temperature foraleast30min.The RNA samplewasheated for30 to 60 sby immersing10
gl
of spreading solution in a beaker ofwater pre-equili-brated in the oven. After heat treatment, 0.5 ,ul of cytochrome c solution (1 mg/ml) was added to the spreading solution, which was incubated in the hot water bath for another 30 s. The spreading was quickly performed inside the oven with the door partially opened. The whole procedure from applying the spreading solution onto the hypophase until picking up the thefilmtookapproximately 30 s.Forexperiments to study the reassociated second-ary structure, the spreading solution after heat treat-ment was quickly chilled on ice for 15 to 30 s. The cytochrome cwasadded and thesolution was spread at roomtemperature.
Preparation for spreading. (i) Glyoxal-form-amide method. RNA samples taken from the gly-oxal-sucrose gradient peak fraction werediluted into the spreading solution to give a final concentration
on November 10, 2019 by guest
http://jvi.asm.org/
399
of 0.2 to 0.5 ug of RNA perml, 40% formamide, 0.1 M Tris, pH 8.2, 0.01 M EDTA, and '30 jig of cyto-chromecperml. Five microliters of the above spread-ing solution wasused in onedrop spreading, whereas 50 pi was required in theconventional dish spreading. The hypophase contained 10% formamide, 0.01 M Tris, pH 8.2,and 0.001 M EDTA.
(ii) Urea-formamide method. RD-114 RNA sam-ples purified either by NTE or low salt-sucrose gradient centrifugation wereused. The RNA samples were diluted into the spreading solution which con-tained -30Agofcytochromecper ml the desired con-centration of urea-formamide and electrolyte.
The urea-formamide solvents used for electron microscope spreadings and for other denaturation studies were prepared as follows. Formamide was purified by recrystallization (11). A solution was preparedby dissolving480g (8mol)of urea(Schwarz/ Mann ultra-pure grade) per liter of formamide. The conductanceofthis solutionat 4C was2x 104ohm-1
cm-1, corresponding to an estimated electrolyte concentrationofabout6mM. We observeda1.35-fold volume increase of the solution relative to the forma-mide. (Thus, the estimated concentrations of com-ponents inthe solventare74%volume percent form-amide and 5.9 M urea, but this calculation is not used in characterizing the mixed urea, formamide, aqueous solutions as discussed below.) A solution preparedfrom pvolumes oftheurea-formamide and (100-p) volumesofaqueous solution is described as a p% (U + F) solution. Unless otherwisespecified, the aqueous electrolyte mixed withurea-formamide con-tained y M (TrisOH + HCl), pH 8.5, 0.1 y M Na8EDTA, with an estimated univalent cation con-centrationof 0.6y M(basedonpKaofTrisH+of8.1). The cation concentration, after dilution with urea-formamide, is reported for each experiment. Thus, forexample, if the aqueous electrolyte had y=0.24, it contained0.24M(TrisOH+HCl),0.024MNa8EDTA.
If40 volumes ofthis solution aremixed with60 vol-umes of(U + F), the final concentrationsare0.096 M (TrisOH + HCl), 0.0096 M
Na3EDTA.
Using8.1 for thepK.
ofTrisH+,wecalculate (TrisH-) =0.027M.The100%(U + F)solvent had an absorbanceat 275 nm of 0.5. The denaturing powerofthe solvent was determinedby optical melting experiments with calf thymus DNAinaqueousU +Fsolutions containing the standard0.06 Melectrolyte. We observe that
Tm = 73 to 0.65C x p(U + F).
For electron microscope spreading from (U + F) solutions, the hypophasewasdistilledwater.
(iii)
CHXHgOH-glyoxal-formamide.
We have also used a modification of theglyoxal technique which veryeffectively extends RNA. Thisprocedureconsists ofthedialysisofthe RNAagainst 1Mglyoxal, 0.045 M sodium phosphate buffer, pH 8, and 10 mM methylmercuric hydroxide for 1 hat room tempera-ture, followedby dialysis against0.1 Mglyoxal,0.045 Msodiumphosphatebuffer, and0.05MNaClfor1h at roomtemperature.Methylmercurichydroxideisaneffective denaturant that disrupts secondary struc-ture features at room temperature (5, 13). The CH3HgOH-extended RNA is easily fixed by glyoxal treatment and theCH3HgOH is then removed by the second dialysis against NaCl and glyoxal.
Agarose gelelectrophoresis. Gels (10 by 0.8 cm) wereformedby melting 0.8 or 1.0% agarose inEbuffer [0.05 M boric acid, 0.005 M sodium borate
(Na2B4O7.10H20), 0.01 M sodium sulfate, and 0.001 MEDTA, pH 8.2]and pouring the hotsolution into an 11-cm glasstube covered at one end with dialysis membrane. To conduct electrophoresis under condi-tionswhich are denaturing for RNA, methylmercuric hydroxide was added to the hot gel solution to the desired concentration. This technique will be de-scribed indetail elsewhere (J. Bailey, personal com-munication).
After solidification of the agarose, the gels were electrophoresed in a vertical tube apparatus contain-ing electrophoresis E buffer in both the upper and lower chambers (the denaturing gel system used in thisstudy doesnotrequirethe presence of methylmer-curichydroxideinthebufferchambers). The samples wereapplied in 50 X of a twofold dilution ofEbuffer containing 10% glycerol, and 5 mM CHsHgOH for denaturing gel electrophoresis. For other experiments, the sampleswere applied in an electron microscope spreading solution containing65% (U + F), 0.026M NaCl, 0.042 M Tris, pH 7.9, and0.5mM EDTA(the total cation concentrationis 0.06M). Electrophoresis wasperformed at 5 mA per tube for periods of2to 3 h at roomtemperature. Withradioactive RNA, the gels were sliced into 2-mm fractions with a Mickel gel slicer and each fractionwasincubated under10ml of Aquasol for 16h. Radioactivity wasdetermined in a Beckman LS-250 liquid scintillationcounter. In sev-eral experimentsunlabeled RNAwasused, in which casebandswerelocatedbyethidiumbromide staining (12). After electrophoresis gels were incubated in 1 ug of ethidium bromide, 0.5 M NH4Ac (to remove
CH.HgOH
and enhance the dye binding) for30min and then examined by illumination with short wavelength ultraviolet light.RESULTS
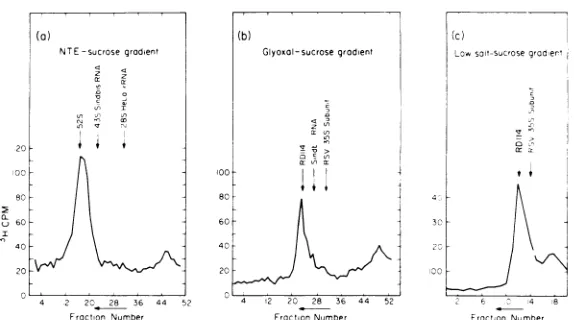
Sedimentation analysis of RD-114 RNA. The sedimentation properties of the high-molecular-weight RNA component extracted from the RD-114 virion has been studied in sucrose gradients in nondenaturing (high salt, NTE), moderately denaturing (low salt), and more strongly denaturing (glyoxal) solvents. There is a high-molecular-weightRD-114 RNA complex which has asedimentation coefficient of52Srelativetomarkers ofSindbis RNA(43S)
and HeLa (28S rRNA) in the nondenaturing
NTEsolvent (Fig. 1). A plot of log M versus log (distance sedimented) in these experiments is
shown in Fig. 2. If a linear relation between these two variables is assumed, the molecular
on November 10, 2019 by guest
http://jvi.asm.org/
(a)
NTE-sucrosegrodient
z c
I T
"I 0
20- i
20
-60
20
4 .2 20 28 36 44 52
Froction Nurmber
100 80 60 40 20
(b)
Glyoxal-sucrosegrodient
4 12 20 28 36 44 52
FroctionNumber
FIG. 1. Sedimentationprofilesof RD-114 RNAasextractedfromthe virion in(a)NTE-sucrosegradient;(b) glyoxal-sucrosegradient; (c) low salt-sucrosegradient.Allproceduresaredescribed in Materials and Methods.
weight of the RD-114 RNA complex is
calcu-lated by extrapolation to be7.0(+0.6) x 106. Boththesecondarystructureand the molecu-lar weight of a polynucleotide chain affect its sedimentation coefficient. We have therefore attempted to obtain a molecular weight esti-mate after disrupting the weaker secondary structurefeatures ofRD-114 RNA and of suita-ble marker RNAs. As a controlled denaturing agent, we have used the reagent,
glyoxal.
As described previously (4, 6, 10) glyoxal, under proper conditions, binds quasi-irreversibly toguanosine residues and disrupts some of the weaker secondary structure features in a poly-nucleotidechainwithoutcausingdissociationof long well-paired duplex segments. The sedi-mentation profile ofRD-114 (in this case with glyoxal-treated SindbisRNAand Rous sarcoma virus 35S subunits as reference RNAs) after glyoxal treatment is shown in Fig. lb. After modification with glyoxal, RD-114 andSindbis RNA both sediment at 0.30times the velocity observed in the nondenaturing (NTE) solvent. A linear extrapolation of the log M versus log (distance sedimented) plotforthetworeference RNAsgivesamolecularweightforRD-114 RNA
of 5.7
(o40.3)
x 106(Fig. 2). As will be reportedin the next section, this value is in good agreement with electron microscope measure-ments.
For preparative purposes we wished to use a weakly denaturing sedimentation medium which did not involve chemical modification of the RNA, but which would cause dissociation
from the high-molecular-weightcomplex of any weakly bound, low-molecular-weight
compo-nents and which might cause dissociation of molecules containing internal nicks. Our proce-dure was to incubate the RNA samples at 37 C
9 8 7
6
5
4
I60
0 x
2
10 20 30 40 50
Distance Moved (Froc. Number)
FIG. 2. Empirical relations between molecular weight and distance traveledas determinedby
sedi-mentation analysis in Fig. 1. Symbols: (A)
NTE-sucrose gradient; (0) glyoxal-sucrose gradient; (0)
lowsalt-sucrosegradient. Calibrationcurvesfor NTE
andglyoxal-sucrose gradient were constructed from the resultsof marker RNAs and plottedonalog-log
scale. (The molecular weight values used here for Sindbis and Rous sarcoma virus RNA have been
presentedpreviously [6, 10].) The molecular weight of RD-114 is estimatedfrom thesecurves. The
horizon-tal error bars correspond to + one fraction. The
verticalerrorbarsfor the molecular weightof RD-114 arecorresponding estimates of the extrapolation
un-certainty.
6 C 4 8
Fraction Number
*28SHeIl rRNA
7
A
A
I
c
2
a-u(
T 1
I
z a:0
I 'It 0 1o
cr ct I ii
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.503.118.405.71.231.2] [image:4.503.267.450.268.520.2]STRUCTURE OF RD-114 RNA 401
in 50% formamide, 50% NTE, andto sediment the product through alowsalt (1 mM Tris, pH 7.0) aqueous sucrose gradient. The peak frac-tions from these preparative runs were pooled for other studies. As shown in Fig. lc, RD-114 RNA againsedimentsat a positionexpectedfor a 5 x 106to 6 x 106daltonRNA species.
We presume that essentially the same high-molecular-weight complex is beingobserved by sedimentation in the three different systems described above. We refer tothis entity as the 52S RD-114 RNA
complex.
Itmaybenotedthat incubationat 37 Cin 50%formamide,50%NTE causes dissociation of 60 to 70S avian RNA tumor virus RNA into 35S subunits(10,
14).
However, the 52S RD-114 RNA is not disso-ciatedby
this treatment.Electron microscopestudies.We have stud-ied the molecular weight, the secondary struc-ture, and the subunit composition of the RD-114 52S RNA complex by electron
micros-copy in experiments in which the RNA is
exposed to a set of conditions of increasing
denaturing
power. It may be recalled that single-strand RNA molecules are not well ex-tended under the usual 40 to 60% formamide, 0.1 M Tris electrolyte, spreading conditions that are effective for extending single-strand DNA(1).Several different solvent systems that are effective for extending RNA have been used in
the present studies. (i) Urea-formamide
spreadings.
Aspreading
solution containing urea, formamide, anda low electrolyte concen-tration is useful for extending RNA molecules undercontrolleddenaturing conditions(15, 16). Inthepresentinstance, wehave usedaseries of solutions with afixedelectrolyte
concentration as described in Materials andMethods. When52S
RD-114 RNA isspread
from 30% form-amide (0.06 M univalentcations,
nourea)
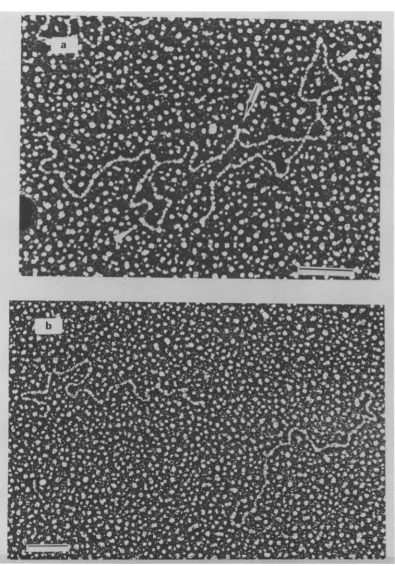
it has a highly condensed structure (Fig. 3a), and the detailed topology of the molecules cannot be discerned.Inspreadingsfrom55% (U + F) (with0.12M univalent cations), the RNA is, in general, still very tangled. However, in somemolecules,
suchasthose showninFig.3b,
thesecondary
structure features described in detailbelow canberecognized. Whenthe (U +F) concentration is raised to 70% (0.06 M
cations), many ofthe molecules aresufficiently wellextendedsothattheycanbe traced. There are two characteristic secondary structure fea-tureswhich arepresent in almostall of the full length traceable molecules: (i) a Y- (or T-) shaped structure located close to the middle, which we refer to as the rabbit ears(RE) struc-ture, (ii) two symmetrically disposed loops, on
each side of and at equal distances from the central RE feature. The micrograph in Fig. 3c illustrates several molecules spread from 70% (U + F). One is quite tangled but the RE can be recognized; intheother two, theREand the loopscanbeseen.
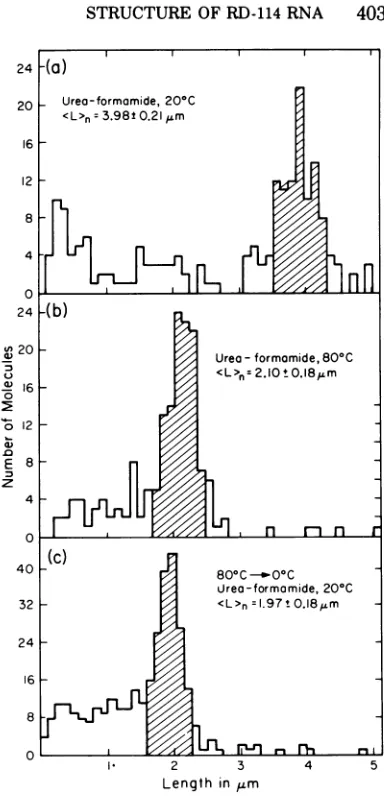
Ifthe RNA isspread from80% (U + F) (0.05 M cations), all ofthe molecules are extended andsuitable forlength measurements (Fig.3d). A histogram of these length measurements is
shown in Fig. 4a, with the number average
lengthof3.98 i 0.21
Am.
We believe that the molecule of molecular length3.98
,m
withtheREandthetwoloops is the 52S RD-114 RNA complex. The length measurementscorrespondto amolecular weight ofabout 5.7 x 106. However,wedeferadetailed discussionoflengths andmolecular weightsand ofthe positions ofthe several secondary struc-turefeatures until alatersection.Amongfull length molecules (shaded area in Fig. 4a) over 97% contained the RE. Ofthese, 46% had one loop and 28% (including the molecule shown in Fig. 3d) had both loops at symmetrical positions.Theseobservations, and others reported below, are consistent with the view that the two loops and the RE are native secondary structure features of 52S RD-114 RNA. Spreading conditions, suchasthose used in Fig. 3d, which are useful for extending the molecules for good length measurements, are sufficiently denaturing to causedissociation of about50%ofthe loops butnotthe RE. Itshould also be noted that the RE inRD-114 RNAwas described in our
preliminary
papers (8, 9), but thereproducibleloopstructures werenot recog-nized.We wished toask whether the RD-114 RNA molecule of molecular weight 5.7 x 106 is one
continuous
polynucleotide
chain or consists of twosubunits, eachofmolecularweight
approxi-mately 2.8 x 106, held togetherby some sortof cohesion within the RE. We therefore sought procedurestoexposethe RNAtostrongly
dena-turing conditions whileminimizing
the risk of covalentchainbreakage.
Asreportedinthisand
following sections,
we have foundtwodifferentdenaturing
treatmentswhich cause the 52S RD-114 RNA molecule to
bedissociated intotwohalf-size
molecules,
with a concomitantdisappearance oftheRE.The structure of the RNAwasobserved when spread from 65% (U +
F)
(0.06 M univalent cations),ontodistilledwater atseveral elevated temperatures as described in Materials and Methods. Below 50C,
3.98gm
molecules with an RE are observed. At60 C andabove,
many half-size moleculeswithoutanREareobserved. 16, 1975on November 10, 2019 by guest
http://jvi.asm.org/
FIG. 3. Electron micrographs of RD-114 spread by theurea-formamide technique. (a) In 30% formamide (no urea, 0.06M cations) at 20 C; (b) in 55% (U + F) (0.12 M cations) at 20 C; (c) in 70% (U + F) (0.06 Mcations) at 20C; (d) in80%o(U + F) (0.05 M cations) at 20 C; (e) in 65% (U + F)(0.06 M cations) at 80 C. Arrows indicate the central RE structure. Triangles point to the loop features. The urea-formamide- and high temperature spreading technique are described in Materials and Methods. The length marker is0.2,m.
A histogram of the size distribution from an
80C experiment is shown in Fig. 4b. The
average molecular length was 2.10 ,um. Over 98% of themoleculesobservedweresmooth and extended without any secondary structure fea-ture, asshowninexamplesinFig.3e. About2% of the molecles were undissociated, with a lengthofabout
4.0,um
and an RE.A solution of RD-114 RNA in the
urea-form-amide solvent was heated to 60 or 80 C,
quenched in ice water, and promptlyspread at
room temparature. The resulting molecules
were half size without an RE. The length
distribution for these molecules presented in Fig. 4c corresponds to an averagelengthof1.97
Am,
in good agreement with the lengthson November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.64.456.74.522.2]STRUCTURE OF RD-114 RNA 403
served in the high temperature spreadings. After heating and quenching, there was a new secondary structurefeature which isillustrated in the micrographs inFig. 5. The structure can bedecribedashavinganunsymmetricalbranch over itsstem, and is referred to as abranch-like structure (BLS). It is morphologically quite different in appearancefrom the RE. The con-tourlengthofthisfeature (tracedassumingit is duplex) is 0.31 ± 0.06 ,um and it occurs at a distance of0.21 ±0.05 jm from one end ofthe otherwise linear 1.97-jm molecules. About 30% of the half-size molecules in the quenched sample had the BLS; the remaining 70% were extended, although slightly knobby.
Between 1 and 2% ofthe molecules were not dissociated by heating to 60 C and quenching
and were full size (4.0
Mm)
with an RE. Arepresentative molecule is shown in Fig. 5a. It contains two BLS, symmetrically disposed at a distance of 0.2
gm
from the RE. This resultstrongly supports the model that RD-114 RNA consistsoftwochainsofmolecular weight 2.8 x 106joined together within the RE. The BLS appears tobe a base-paired structure due tosequencesextendingfrom apointclosetoRE to a point slightly within the loop structure. It is formed after quenching when the loop structures seen inthenative52S RNA complex aredissociated.(ii) Glyoxal-formamide spreading. Reac-tion withglyoxal under the conditionsin Mate-rials and Methods modifies single-strand RNA sothatit iswellextended informamide spread-ings, but does not cause denaturation of well-matched duplex structures. We prefer glyoxal spreading to urea-formamide spreading for quantitative length measurements because in our hands it gives more constant and narrow length distributionsfor a homogeneous RNA.
Glyoxal-treated
RD-114 RNA from thepeak
fractions of the glyoxal-sucrose gradient (Fig. lb), whenexamined inthe electron microscope understandard 50%formamidespreading
con-ditions, appears as a linear extended filament withthe RE closetothemiddleofthemolecule. The loop structuresobserved in the urea-form-amide spreads were also observed in glyoxal spreadings. An electron micrograph of a full length moleculewiththeRE and the twoloops, symmetrically disposed relative to the RE, is showninFig. 6a. Asshown in the histogram in Fig. 7a, thesizedistributionoftheRNA mole-cules isreasonably homogeneouswithanumberaverage length of 4.27 + 0.17 ,um. We use
Escherichia coli23SrRNAas anexternal
length
standard((L),
= 0.80 0.04gm,
molecular weight = 1.08 x106)
andcalculate themolecu-24
-(a)
20 Urea-formamide,20°C
20 ~ ~ ~ rafommd,2°
16 12
8
4 2 3 4
0 24
-(b)
RD-11420 A spread by the
Urea
-formamide,800CMcation) at 0 C; () RD-14RN<L>A=2.iot0.18%m 16-6
0 0 12-E8 z
4
0
40-(c) 80C -14Ni
wrea-formamide, 200C
3?- <L>nI=.97t0.18MLm
24- 16-8 0._
2 3 4 5
Lengthin Mm
FIG. 4.
Histogram
of thelength distribution
of RD-114 RNA spread by the urea-formamidetech-nique.
(a)
RD-114 RNAspread in 80% (U+F) (0.05M
cations)
at
20C; (b)RD-114 RNAspread
in65%(U+F)(0.06Mcations)at80C; (c)
RD-1J14
RNA in 65%(U +F) (0.06M
cations), heated
at80C, chilled onice, and
spreadat20C. The numberaverage lengths
were calculatedfrom molecules
intheshadedarea.lar
weight
of this RD-114molecule
as 5.74 0.23 x 106. In theselength
measurements the RE structure is treated asentirely duplex,
and itssingle-strand length
measuredby going
back and forthoverthe entire feature.Over97% ofthefull
length
molecules(defined
by
the shaded area inFig.
7a) contain the RE.By tracing
upanddown,
asindicatedabove,
itssingle-strand length
was estimated as 0.21±i
0.04 um.Approximately
28% of theglyoxal-treated full
length
molecules contain two sym-metricalloops,
52% contain oneloop,
and the VOL.16, 1975on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.503.257.449.61.461.2]b4".
~~~~~~
"~~L
14C
.4
"40:hj
Jo~
h.Ahgp40
a0,~~~~~~~~~~~~~
FI.5Eetonmcorah f etd n hlldR-14RAspedbyue-fraie ehiqe 2
RD11RAamle i 6% U+ ) er hatd o 0 ,thn uikl qenhe o cean sred t 0 . a
An~~~~~euniscae0.0 atnR-1 N.Temlcl otisteR arw n h w L
(triaglessymetriallylocaed o eac sid of he RE (b nd c Eac sho a 28 x 06 dltonRD-14 RN
wit
thILS(ranl).Teleghmakr
s02 .mon November 10, 2019 by guest
http://jvi.asm.org/
OF RD-114 RNA
FIG. 6. Electron micrographs of RD-114 RNA spread by the glyoxal-formamide technique. (a)
Glyoxal-treated RD-114RNA;(b)
CHJHgOH-glyoxal-treated
RD-114 RNA. An arrow indicates the central RE structure.Triangles pointtothetwoloop features.ProceduresforRNA treatmentand electronmicroscopy spreadingare detailed in Materials and Methods. The length marker is 0.2um.
16,1975
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.503.46.441.61.627.2]2 ~~~~~~~~~~~~~8
0~~~~~~~~~~~~~~~~~
2
L
n Finf
n
n
d
(3b 40 __ -<Loop( Jointto REdistonce)n
z CH3HgOH/glyoxal-formomide
<L>n 2.07t0.l5im
12 M.W. 2.79'0.200x06 doltons 32
10
24
8-6
16-
4-8
2 3 4 5 2 3 4 5
[image:10.503.116.408.70.327.2]LengthinFm LengthinLLm
FIG. 7. Histogram of the length distribution ofRD-114 RNA as studied by the glyoxal-formamide and
CH,HgOH-glyoxal-formamide
techniques. (a) The totallength of glyoxal-treated RD-114RNA; (b) the total length ofCH8HgOH-glyoxal-treated52S RD-114 RNAfromalow saltgradient; (c)endtocentral REstructure distanceofglyoxal-treatedRD-114RNA; (d)theloopjointtocentral REstructuredistance and theloopsizeof glyoxal-treatedRE-114 RNA.remaining 20% do not have any. As shown by the histogram in Fig. 7d, these loops are mapped reproducibly at a distance0.44 + 0.06
Am
from the RE and have a homogeneous loop length of 0.78 ± 0.09 ,m. One would suspect that theloopstructures areduetoshort comple-mentary sequences at the baseofthe loop. Ifso, the lengthofthisduplexsegment appears tobe less than200nucleotidepairs and thustooshort tobepositively identifiedassuchinthe electron micrographs. (One can, however, imagine that suchaduplex segmentdoesexistonexamining micrographs suchasthat showninFig. 6a.) The observation that not all molecules contain the loop structure suggests that the glyoxal dena-turingconditionsfrequentlycausedenaturation ofthe shortsequence ofbase pairsinvolved.Thus, the results from theglyoxalspreadings confirm the structures observed in urea-form-amidespreadingsand supporttheviewthat52S RD-114 RNA isolated either by NTEgradients orby glyoxal gradients has essentially the same structure; i.e., a molecule of molecular weight
5.7 x 106,withthe central REstructureandthe twosymmetricallydisposed loop structures.
We wish to ask whethertheRE isactually in
thecenter orslightlyoff-center of the molecule. LetL2and
L,
be distancefrom hypotheticalleft and right ends of an asymmetrical molecule of molecular weight 5.7 x 106with an RE feature close to but not necessarily at the center. We cannot a priori distinguish L2 fromL,
for any molecule measured. We have calculated the root mean squeare value oftheobserved distri-bution of values of the intrinsically positive (and measurable) quantity,IL2
-L11,
and find that it is 0.16 ,m. For the same sample, the standarddeviation of the distribution of values of(L,
+L2)wasmeasuredas 0.17gm.
IftheRE were at the center of the molecule, the average value of the unobservable quantity (L2 -Lj)
wouldbezero andthe,rootmean square value of
JL2
- L ispredicted
tohave thesamevalueasthe standard deviation in values of
(L,
+ L2). Thus, the observations support the view that the RE is atthe center of the molecule.(iii) Methylmercury-glyoxal spreadings. We wished tostudythe possibledissociation of the52SRNAcomplexintoits twosubunitsbya strongly denaturing treatment which did not involve elevated temperatures as in the urea-formamide thermal dissociation procedures. We
on November 10, 2019 by guest
http://jvi.asm.org/
STRUCTURE OF RD-114 RNA 407
have found that treatment with glyoxal in the presence of methylmercuric hydroxide is effec-tive forthispurpose.
CH3HgOH is areversible denaturing reagent for nucleic acids because it reacts with the imino NH bonds of G, U, and T (5, 13). It is a powerful denaturant. For example, at room
temperature in 0.1 M
Na2SO,
at pH 8.65Micrococcus luteus DNA which has a guanosine plus cytosine content of 72% is denatured at a CH3HgOH concentration of 2.9 mM (5). We therefore anticipated that a sufficiently high concentration of
CH3HgOH
would denature any base pairing within the RE. As described in Materials and Methods the guanosine residues exposed by denaturation werethenfixed in the nonhydrogen bonding state by treatment with glyoxal. Accordingly, 52S RD-114 RNA from a lowsalt gradientwastreated with glyoxal in the presence of 10 mM methylmercuric hydroxide, as described in Materials and Methods. Note that the methylmercuric hydroxide is removed before spreading. Figure 6b shows an electron micrographofthe resulting RD-114RNA mole-cules.Asshownby the histograminFig. 7b, the resulting molecules have an average length of 2.07 + 0.15 ,um corresponding to a molecularweightof 2.8 i 0.2 x 106. They donothaveany of the characteristic secondary structure fea-tures of the 5.7 x 106 dalton molecules. It therefore appears that the treatment described dissociates the 5.7 x 106 dalton RD-114 RNA into two subunits, each of molecular weight one-half the valuefor the starting molecule.
Gel
electrophoresis
of RD-114 RNA. The dissociation of 52S RD-114 RNA into two half-size subunits under suitable denaturation con-ditionshasbeen confirmedby
gel electrophore-sis inseveraldenaturing solvent systems.We have recently developed a methylmercu-richydroxide-agarosegelsystem for electropho-resisunder denaturing conditions. (Bailey and Davidson, personal communication). 52S RD-114 RNA
electrophoreses through
a 1% agarosegel
containing5 mMCH3HgOH
in the boratebufferelectrolyte
described inMaterials and Methods at avelocity
that isgreater than that ofSindbis
RNA but less than that of28S HeLarRNA,
and23S
E.coli rRNA.Typical gel
patterns are illustrated in Fig. 8a. The log M versus(distance migrated) plotforseveral refer-ence RNAs in this gel system is linear (Fig. 9) fromwhichwe interpolatethe molecularweight ofRD-114 RNA as2.64(±0.1)
x 106. J. Bailey had observed that nicked circularduplex PM2 DNA is dissociated intosingle-strand
compo-nents inthe agarosegel-methylmercurysystemat methylmercury hydroxide concentrations
greater than 3 mM. Therefore, the concentra-tion of 5 mM usedin the RD-114 RNA experi-ments is strongly denaturing. The observed molecular weight is approximately one-half of that (5.7 x 106) of the 52S RD-114 complex. Therefore we conclude that RD-114 RNA is dissociated into half-size subunits inthe dena-turingmercurial gel.
A more precisestudyofthe thermal dissocia-tion of52S RD-114 RNA (molecular weight5.7
X 106
by
electron microscopy and sedimenta-tion analysis) into half-size subunitswas made using a gel electrophoresis assay to observe dissociation. Labeled 52S RD-114 RNA in the 65% (U + F), 0.06M cationsolvent usedforthe thermal dissociation electron microscopestud-ies, was heated to various temperatures,
quenched on ice, and
subjected
togel
electro-phoresis inthenondenaturingEbuffer at room temperature. It may be seen fromthe radioac-tivity profiles shown in Fig. 8b that there is a pronounced increase in the distance migrated for a sample heated to 60C as compared to a sample heated to 55 C. Samples heated to 25, 40, and50Callmigratedwith the samevelocity as the 55C heat treatment; samples heatedto 70 and 80 C migrated like the 60C sample. Furthermore, asampleofunlabeled 52S RD-114 RNA from an NTEgradient
was diluted di-rectly into a twofold dilution of the E buffer,subjected
toelectrophoresis
in the standardbuffer,
and locatedby
ethidium bromide stain-ing. ItspositionasshowninFig.8isthesame as thatofthesample
heatedto55 C. Thereisthus a structural transition with a marked increase in mobility between 55 and60 C.By interpolation from the values for the electrophoretic mobilities of HeLa 28S rRNA and Sindbis RNA in the same gel system, the molecularweightsofthe low andhigh tempera-tureforms ofRD-114 RNAareestimatedas6.7 x 106 and 4.2 x 106. These values are
higher
thantheexpected valuesof 5.7 x 106 and 2.8 x 106.A sampleofRD-114 RNAwastreated with 10 mMCH3HgOH
inNa2SO4,
boratebuffer,
and then dialyzed against 0.1 MNH,Cl
toremove the bound material and studied
by
electrophoresis
in thenondenaturing
gel.
We believe, from the electrophoresis studies in 5mM
CH3HgOH
that this treatment will cause dissociation of RD-114 RNA into 2.8 x 106dalton subunits (actually measured as 2.64
0.1 x 106by electrophoresisin amercurialgel). Themobilityofthissample
corresponded
to anapparent molecularweight of4.2 x 106. Thus, we believe that the 52S RD-114 RNA
VOL.16, 1975
on November 10, 2019 by guest
http://jvi.asm.org/
150
100o E
0.
C-50
200
E
ca-5 10 15 20 25 30 35 40
Fraction Number
5 10 15 20 25 30 35 40 45
[image:12.503.122.409.66.443.2]Fraction Number
FIG. 8. Electrophoretic gel patterns. Electrophoresis is from left to right. (a) RD-114 RNA in a 5 mM CHJHgOHagarosegel.Thearrowmarks theposition ofSindbis 43SRNA; (b) gelpatternsafterheatingto55C and to 60C. Electrophoresis is in electrophoresis buffer at 22C. The arrow is the migration position of unlabeled 52S RD-114directly from anNTEgradient, asdetected byethidium bromidestaining.
with a molecular weight of 5.7 x 106 and its
half-size subunit with amolecularweightof 2.8
x 106 show anomalously low electrophoretic
velocities dueto relatively extended configura-tions, as compared to HeLa 28S rRNA and Sindbis RNA in the nondenaturing
electropho-resis buffer. The data presented here therefore
fix the melting temperature of the central RE feature, which is mainifested bydissociation of
52SRNA intohalf-size subunits, asbetween55 and 60 C in the 65% (U + F), 0.06 M cation solvent.
As reported in Materials and Methods, the melting temperatureofcalfthymus DNAinthe
same solvent was measured as 33 C in an
opticalmelting experiment.
DISCUSSION
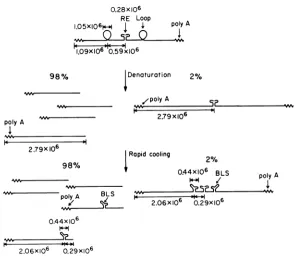
Figure 10summarizesourobservationsofthe
structure and properties of RD-114 RNA. The high-molecular-weight component extracted from the virion has asedimentation coefficient
of 52S anda molecularweightof 5.74 0.17 x
106.It containsacharacteristicsecondary struc-turefeature thatwe describe asthe centralRE
withamolecularweightofabout 0.3 x 106.The rest ofthe molecule is mainly single stranded, but there are two loops at the symmetrical positions around the RE and with the
dimen-sions indicated in Fig. 10.
The 5.74 x 106 dalton molecule can be
dissociated into half-size molecules by
suffi-a.
\V/
I
on November 10, 2019 by guest
http://jvi.asm.org/
STRUCTURE OF RD-114 RNA 409
ciently denaturing conditions (methylmercury-glyoxal, or thermal dissociation in urea-form-amide). TheRE dissappears upon dissociation.
We presume, that there is some sort of a
base-paired structure betweenthe two subunits giving rise to the RE (Fig. 10). By using a gel electrophoresis assay, we have concluded that
0E
0
E E 0
0.5 1.0 1.5 2.0 2.5 3.0 3.5 4.0 4.5 mobility (arbitrary units)
FIG. 9. Aplot of RNAelectrophoretic mobility in 5 mM CH3HgOH 0.8% agarose gels as a function of
molecular weight. Marker RNAs, indicated by the filledcircles, are16S and 23S E. coli rRNA, 28S HeLa rRNA, and Sindbis virus RNA with respective molecular weights of 0.56, 1.10, 1.90, and 4.67 mega-daltons. The mobility and interpolated molecular weightof RD-114 RNA are shown.
0.28xlO6 RE Loop
1.05X106"
the melting temperature for dissociation into half-size molecules in 65% (U + F), 0.06 M univalent cation, is between 55 and 60 C. By comparison with the melting temperature of 31 C forcalf thymus DNA inthesame solvent, weestimatethat the meltingtemperature ofthe RE is 108 to 113 C in an aqueous electrolyte containing 0.2 M Na+. The data collated by Kallenbach (8) on the melting temperature of duplex RNAsas afunction of their base compo-sition would then suggest that, if the RE is a simple RNA duplex, its base composition is about63%guanosine plus cytosine.
W. Bender at this laboratory has developed and applied a method of electron microscope mapping of polyadenylic acid
[poly
(A)]
stretches on polynucleotides byhybridization
withpoly (dT) attachedtoduplexsimian virus 40 circles (as anelectron microscopelabel)
(W. Bender, personal communication).This
method, which is based on a suggestionby
J. Carbon, is an extension of onepreviously
re-ported (6). Bender finds that therearepoly (A)
stretches onboth outside termini ofthe 5.74 x 106 RD-114 molecule. We presume that the poly(A) stretches arelocatedatthe3'-termini of the half-size subunits; therefore the two 5'-termini are contained within the RE(Fig. 10).
Note that theobservationofapoly(A)
stretchatpolyA
I,09X106 0.59x106 98%
poly A
2.79X 106
tDenaturation 2%
/polyA 2.79x106
Rapid cooling
QR0/
0.44X106 BLS polyA
polyA BL 6o
poy/ 2.06x106 0.29xlo6
0.44x106
2.06X106 0.29X106
FIG. 10. Structure and interconversion between the severalforms ofRD-114 RNA described in the text
(molecular weightsin daltonsareshown,andarebasedonelectronmicroscopylengthmeasurements). 2%
VY%
F- m
VOL.16, 1975
on November 10, 2019 by guest
http://jvi.asm.org/
[image:13.503.48.240.161.296.2] [image:13.503.99.396.378.635.2]bothofthe outside ends demands that the5.74 x 106 dalton molecule be
composed
of two subunits, ifpoly(A) stretches occur only at 3' ends.We wish to note at this point that in our earlier preliminary reports (9, 10) we drew the tentative conclusion that the5.74 x 106dalton molecule was one continuous
polynucleotide
chain. We now believe that the conclusionwas incorrect. Atthattime we also failed to notice thereproduciblepresence oftheloopstructures. It should also be noted that J. L. East et al. (2) had reported that the sedimentation coeffi-cient of the high-molecular weight-component of RD-114 RNA, as extracted from the virion, was50S
and that thermal dissociation in aqueous electrolyte solution led to the forma-tion of28Ssubunits, allin approximate agree-mentwith our present interpretations.WhenRD-114
RNA,
in 65%(U+ F),isheated to temperaturesabove60 C and then quenchedon ice so that the loops disappear a new
secondary structure feature, the BLS, is ob-served. This featureisclose tothecenter, buta few molecules are observed that contain both theBLS and theRE (Fig. 10). Thisobservation strongly suggests that the BLS is a distinct sequenceandnotpart ofthe RE. The quantita-tive dimensions indicate that theBLS involves somesequenceclosertothecenterthan thetwo loops, but also include some sequences just within the loop (Fig. 10).
In spreadings atroom temperature from 70% (U + F), almostall full length traceable mole-cules show the two loops symmetrically dis-posed around the RE. In 80% (U + F), about 50% ofthe loops are seen. A simple interpreta-tion of these results is that the loop is held together by short complementary sequences which are about 50% melted in 80% (U + F) at room temperature. If a loop is not present, the RNAappears tobeasimple singlestrand in this region; i.e., the BLS is not seen. After heating and cooling in 65% (U + F), the BLS isseen. Since the BLS involves sequences closer to the center of the molecule than is the loop and sequences within the loop, and since the BLS is not seen when a loop is present, wetentatively conclude that theformation of the BLS requires an interaction between sequences within the loop with sequences closer to the center. An alternativepossibility is that the loop structure is somehow held together by an additional low-molecular-weight molecule; the BLS can-not form until this molecule is removed by thermal dissociation.
We propose that two 2.8 x
106
molecules of RD-114 RNA are joined together at their 5'3" ' o'I..l.a 5 uo
FIG. 11. Simplestpossible structure fortwo RNA strands heldtogether byanti-parallel basepairingat their5'ends.Ifthetwostrandsareidentical,eachone mustcontainaanditscomplementa' asindicated. ends to form the RE feature. Watson-Crick base pairing requires that the two strands be oriented antiparallel with respect to each other.Inthesimplestcase, this would leadto a linear structure at the junction of the two strands (Fig. 11). We have been unable to conceive of any simple,
antiparallel
scheme of base pairing between two strands that would lead to a Y- orT-shapedstructure.The possibil-ity that an additionallow-molecular-weight
RNA component is involved in the cohesion within the RE shouldalsobe considered. Since theRNA had been extractedby
treatmentwith phenol, sodiumdodecyl sulfate, and Pronase, it isunlikely(but notexcluded) thatanyproteins are involvedin the cohesion.Weobserve that: (i) the molecularweight of the two 2.8 x 106 subunits into which the 52S RD-114 RNA dissociates are identical within experimental error, as indicated
by
asingle
sharp peak in both the electron microscope length histogram and the gelelectrophoresis
pattern ofthe dissociated subunits; (ii) all the secondary structure features observed in these two subunits are identical in position and shape; (iii) both subunits contain apoly(A)
stretch at theend.One simple interpretation ofthese results is that the 52S RD-114 molecule is a dimer
con-sisting of twoidenticalsubunits, joined
together
somehowattheir 5' endswithin the RE. These observations areconsistent with results for the RNAfrom othertumor viruses,indicating
that the60 to70S
complexcontainstwo orthree sub-units ofmolecular weightinthe range 2.5 x 106 to 3.3 x 106, and that theseseveral subunitsare identical in sequence (M. A. Baluda, M.Shoyab,
P. D. Markham, R. Wans, and W. N. Drohan, Cold Spring Harbor Symp. Quant. Biol., in press; H. Delius, P. Duesberg, and W. Mangel, Cold Spring Harbor Symp. Quant. Biol., in press; Duesberg, P. K. Vogt, M. Lai,and K. Beeman, Cold Spring Harbor Symp.
Quant. Biol., inpress).
ACKNOWLEDGMENTS
This research has been supported by contracts NO-1-CP-43306 and PH 43-68-1030within theVirus-Cancer Pro-gram ofthe National Cancer Institute. We thank Welcome Benderforpermissiontoquotehisunpublished poly(A) map-ping results. We also thank James Casey for helpful dis-cussions.J.M.B.istherecipientof a HelenHay Whitney Fel-lowship.
on November 10, 2019 by guest
http://jvi.asm.org/
STRUCTURE OF RD-114RNA 411
LITERATURE CITED
1. Davis, R., M. Simon,and N. Davidson. 1971. Electron microscopicheteroduplex methodsfor mapping regions ofbasesequencehomology,p.413-428. In L.Grossman
and K. Moldave (ed.), Methodsinenzymology, vol.21. Academic Press Inc., NewYork.
2. East, J. L., J.E.Knesek,P.T. Allen,and L. Dmochow-ski. 1973. Structural characteristics and nucleotide
sequenceanalysisof genomicRNAfromRD-114virus andfeline RNAtumorviruses.J. Virol. 12:1085-1091. 3. Filbert,J.E.,R. M.McAllister, M.0.Nicolson, and R.
V. Gilden. 1974. RD-114 virus, infectivity assay by
measuirements ofDNApolymeraseactivity andvirus
group specific antigens. Proc. Soc. Exp. Biol. Med.
145:366-370.
4. Forsheit,A.B.,N.Davidson, and D. B. Brown.1974.An electron microscope heteroduplex study ofthe ribo-somal DNAs inXenopus laevis and Xenopus mulleri. J. Mol. Biol. 90:301-314.
5. Gruenwedel,D.W., andN.Davidson. 1966.Complexing and denaturation of DNAby methylmercuric hydrox-ide. J. Mol. Biol. 21:129-144.
6. Hsu, M. T., H. J. Kung, and N. Davidson. 1973. An electron microscopestudyofSindbisvirusRNA.Cold Spring Harbor Symp. Quant. Biol.38:843-950. 7. Inman, R. B., and M. Schnos. 1970.Partial denaturation
of thymine-and 5-bromouracil-containing A DNA in
alkali. J.Mol.Biol.49:93-98.
8. Kallenbach, N. R. 1968. Theoryofthermaltransitions in low molecular weight RNA chains. J. Mol. Biol. 37:445-466.
9. Kung, H. J., J. M.Bailey, N. Davidson, M.0.Nicolson,
and R. M.McAllister. 1974.Structure and molecular length of the large subunitsofRD-114 viral RNA. J. Virol. 14:170-173.
10. Kung, H. J., J. M.Bailey, N. Davidson, P. Vogt, M.0.
Nicolson,and R. M. McAllister.1974.Electron
micro-scopestudiesoftumorvirus RNA. ColdSpring.Harbor Symp. Quant. Biol. 39:827-834.
11. Robberson, D.,Y.Aloni, G. Attardi, and N. Davidson. 1971.Expressionofthe mitochondrialgenomeinHeLa
cells. XI. Size determination ofmitochondrial ribo-somal RNA by electron microscopy. J. Mol. Biol. 60:473-484.
12. Sharp,P.A., B.Sugden, and J. Sambrook. 1973. Detec-tion oftworestrictionendonucleaseactivities in Hae-mophilusparainfluenza using analyticalagarose ethid-ium bromide electrophoresis. Biochemistry 12:3055-3063.
13. Simpson, R. B. 1963. Association constants of methyl-mercuricand mercuric ions withnucleosides.J. Am. Chem. Soc.86,2059-2065.
14. Travnicek, M., and Riman, J. 1973.Subunits of
oncor-navirus high-molecular-weight RNA. I. Stepwise
con-verstion of 60S AMV RNA to subunits. Biochem. Biophys. Res. Commun. 53:217-223.
15. Wellauer, P. K., and I. B. Dawid. 1973. Secondary structuremapsof RNA:processingof HeLa ribosomal RNA.Proc.Natl.Acad. Sci. U.S.A. 70:2827-2831. 16. Wellauer, P. K.,I. B. Dawid, D. E. Kelley, and R. P.
Perry. 1974.SecondarystructureofmouseL-cell
ribo-somal RNA andvariations inprocessing pathway.J. Mol.Biol. 89:397-407.