0022-538X/11/$12.00 doi:10.1128/JVI.01612-10
Copyright © 2011, American Society for Microbiology. All Rights Reserved.
Structural Comparison of HIV-1 Envelope Spikes with and
without the V1/V2 Loop
䌤
†
Guiqing Hu,
1,2Jun Liu,
3Kenneth A. Taylor,
1,2and Kenneth H. Roux
1,2*
Department of Biological Science1and Institute of Molecular Biophysics,2Florida State University, Tallahassee,
Florida 32306-4295, and The University of Texas—Houston Medical School, Department of Pathology & Laboratory Medicine, 6431 Fannin, MSB 2.228, Houston, Texas 770303
Received 2 August 2010/Accepted 21 December 2010
We have used cryoelectron tomography of vitreous-ice-embedded HIV-1 virions to compare the envelope (Env) spikes of a wild-type strain with those of a mutant strain in which the V1/V2 loop has been deleted. Deletion of V1/V2 results in a spike with far more structural heterogeneity than is observed in the wild type, likely reflecting greatly enhanced gp120 protomer flexibility. A major difference between the two forms is a pronounced loss of mass from the “peak” of the native Env spike. The apparent loss of contact among three gp120 protomers likely accounts for the more open structure, heterogeneity in configuration, and previous observations that broadly neutralizing epitopes and reactive sites on other structural elements are more exposed in such constructs.
Viral tropism and cellular entry by human immunodefi-ciency virus type 1 (HIV-1) and simian immunodefiimmunodefi-ciency virus (SIV) are mediated by heterotrimeric viral envelope (Env) spikes which interact initially with cellular CD4 and subse-quently with either the CXCR4 or the CCR5 chemokine re-ceptor (15). A variety of structural and functional data indicate that these receptor-ligand interactions induce a dramatic struc-tural reorganization of both the gp120 spike head and the gp41 spike stalk, culminating in fusion of the viral and cell mem-branes. Gp120 consists of conserved segments which form a core structure and four variable regions. The V1/V2 and V3 variable regions constitute extended loop structures which likely comprise a significant portion of the gp120 surface fea-tures (reviewed in references 63 and 89). The large V1/V2 loop complex, which is subdivided into V1 and V2 by a disulfide bridge, is involved in modulating viral tropism and resistance to neutralization (7, 13, 31, 34–36, 56, 63, 72, 78).
Both unliganded and liganded soluble recombinant forms of Env gp120 have been described at the atomic level but only as isolated monomer core structures (11, 27, 30, 38, 39) or, more recently, the core structures with V3 loop (26) and N- and C-terminal peptides (62). The unliganded atomic structure of an SIV gp120 core devoid of the V1/V2 and V3 loops has also been described (11). No comparable HIV-1 unliganded atomic structure has been reported. Also unknown is the precise man-ner by which Env gp41 and gp120 associate or how the three gp41/120 subunits are assembled in their native trimeric struc-ture.
Artificially trimerized constructs with or without the gp41 ectodomain have been produced, but these do not fully mimic the antigenic profile or, presumably, the structure of native
spikes (3, 5, 14, 18, 21, 66, 91) and have proven refractory to crystallization attempts. However, the available evidence de-rived from the atomic structures of gp120 monomers is suffi-cient to permit the construction of hypothetical trimer models (11, 26, 30, 38, 39, 79).
One such model, based on the crystal structure of unligan-ded SIV gp120, positions the V1/V2 and V3 stalks so as to project in opposite directions (11). This aspect of the model was rather unexpected since there is considerable evidence for an intimate association between the two loops both before and after CD4 binding (54, 58, 64, 67, 80). This apparent dilemma can be resolved by proposing a model in which the V1/V2 loop of each subunit reaches across to interact with the V3 loop of a neighboring protomer (11). This mode of association would likely contribute to a stabilization of the native trimer.
The V3 loop is typically a prime target for neutralizing antibodies (13, 28, 42, 70). The apparent accessibility of the V3 loop varies considerably between viral strains; some viruses, termed tier 1, are readily neutralized by anti-V3 antibodies, whereas tier 2 viruses are neutralization resistant (30, 44, 51, 75). It is generally assumed that neutralization resistance is due to physical shielding of V3 by the V1/V2 loop (7, 13, 31, 35, 36, 56, 63, 72, 78), though less direct epitope-modulating mecha-nisms are possible.
The V1/V2 loop, in contrast to V3, appears much less im-munogenic, though it can express neutralization determinants, typically of lower affinity (14, 16, 24, 36, 87). Whereas V1/V2 may shield V3, V1/V2 itself is substantially shielded and may be conformationally influenced by extensive glycosylation (39, 49, 82, 89).
Because of the proposed shielding of potentially broadly neutralizing epitopes on V3, including the coreceptor binding site, and other regions of the Env spike (7, 31, 35, 61, 78, 90), constructs in which all or portions of V1/V2 have been deleted have been extensively evaluated for potential use in Env-based HIV-1 vaccines (1, 2, 14, 18, 22, 32, 77). The hope is that removal of the shielding action of the V1/V2 loop will allow fuller exposure to cryptic but potentially broadly neutralizing
* Corresponding author. Mailing address: Department of Biological Science, Florida State University, Tallahassee, FL 32306-4295. Phone: (850) 644-5037. Fax: (850) 645-8447. E-mail: [email protected].
† Supplemental material for this article may be found at http://jvi .asm.org/.
䌤Published ahead of print on 29 December 2010.
2741
on November 7, 2019 by guest
http://jvi.asm.org/
epitopes. Thus far, most studies have failed to generate signif-icant levels of broadly neutralizing antibodies following immu-nization (43); however, one recent report shows significant protection in a macaque model using an HIV-1 trimer with V2 deleted and a relevant SHIV challenge (76).
In this report, we compare the overall structures of wild-type (WT) HIV-1 Env spikes on intact virions to the structure of ⌬V1/V2 Env spikes by cryoelectron tomography (cryo-ET). The results reveal a dramatic difference in morphology wherein the⌬V1/V2 spikes are missing considerable density at the apex and the individual gp120 subunits display variable morphology indicative of pronounced subunit flexibility.
MATERIALS AND METHODS
Viruses.Strains R3A WT (GenBank accession no. AY608577) (52) and R3A
⌬V1/V2 (GenBank accession no. EF613595) (40) were provided as highly
puri-fied virus samples treated with AT-2 by the AIDS Vaccine Program (SAIC Frederick, NCI, Frederick, MD).
Cryoelectron microscopy (cryo-EM) preparation.Viruses were mixed with 10-nm colloidal gold (used for better tracking during tilt series data collection) and deposited on homemade, carbon-coated, holey carbon grids for 1 min. Excess virus and buffer were removed by blotting with filter paper, and the preparation was rapidly vitrified in a homemade cryoplunger.
Cryo-ET.The frozen virus specimens were examined under low-dose condi-tions at liquid nitrogen temperature using an FEI G2 Polara electron microscope equipped with a field emission gun and TemCam-F415 4,096-by-4,096-pixel charge-coupled device (CCD) camera (TVIPS, Gauting, Germany). The
micro-scope was operated at 300 kV and a magnification of⫻39,000, resulting in an
effective pixel size of 0.46 nm after 2-by-2 binning. Single-axis tilt series were collected using the FEI “batch tomography” capability of the Xplor3D software.
Defocus was set to 4 to 5m with a cumulative dose of⬃100 e⫺/Å2
. Each tilt
series consisted of⬃120 images recorded over an angular range of⫺65° to⫹65°.
Twenty-four WT and 26 mutant tilt series suitable for tomography were col-lected. Tilt angles for data collection were determined according to the Saxton scheme (73). After computation of the tomogram, the specimen thickness was found to range from 170 to 250 nm for the WT and from 140 to 197 nm for the
⌬V1/V2 mutant.
Image analysis.The tilt series were aligned based on the intrinsic features of the individual projection images (rather than the gold fiducials) using the PRO-TOMO program package (85). Consequently, before alignment, the images of the colloidal gold particles were masked out and replaced with areas which had the density of the mean value of the whole image. Tomograms were computed by weighted back projection (85).
The individual spike volumes were manually picked from one virion at a time. Initial Euler angles for each spike were assigned by assuming that the spike was
oriented perpendicular to the viral surface and aligning thezaxis with the vector
between the center of the virus particle and the center of the extracted spike. Each spike volume was then transformed based on these initial Euler angles, and a global average was generated and used as an initial reference to align the raw spike volumes. The initial average had no regions that were data poor due to the missing wedge since the spikes were randomly oriented with respect to one another over the entire surfaces of the virions. The initial alignment was the only step where the raw spike subvolumes were themselves aligned. In this step, only translation alignment was permitted; no rotational alignment was performed in
the first cycle. The aligned particles were then grouped into⬃10 to 30 classes by
multivariate data analysis and hierarchical ascendant classification (84). Class averages were generated and aligned with one another by translation and rota-tion. The translation and rotation obtained for each class average were then applied to the stored alignment parameters of the constituent members of each class, and the resulting new set of parameters was stored for the next cycle. Thus, classifications that produce the class averages were performed on the raw spike subvolumes, but alignments were performed using only the class averages and the results applied to the individual spike subvolumes. This procedure was cycled
⬃12 times until the appearance of the classes no longer changed. For the HIV-1
V1/V2 loop deletion mutants, around 10% of the particles with high variance in the multivariate data analysis were discarded. No WT spikes were discarded. In the final cycle, spike volumes were grouped into 10 classes. For the spike align-ment, the mask included the spike and a small portion (outer leaflet) of the membrane. For classification, the mask included only the spike; the membrane
was excluded. Previous work has shown that including the density corresponding to the lipid bilayer in the classification introduces a bias that separates spikes extending out the sides of the virions, where the membrane densities are prom-inent, from those that extend out the top, where the bilayer is not visible or has low contrast (86).
The signal-to-noise (S/N) ratio for the WT and mutant spikes was estimated
using the equationvar(p[N])⫽var(p)⫹1/N var(n), wherevar(p[N]) is the
variance of the average ofNspikes,var(p) is the signal variance of an average of
an infinite number of spikes, andvar(n) is the noise variance, which decreases
inversely with the number of spikes in an average (69). The S/N ratio of the raw WT spike is 0.1, and that of the raw mutant spike is 0.18. Each class average has
⬃150 raw particles, so the S/N of the corresponding class averages should be 150
times that of the raw image, 15 for the WT and 27 for the mutant.
Statistical analysis of difference maps.To obtain an estimate of the signifi-cance of the difference in density between WT and mutant Env spikes, a cylin-drical mask which excluded the membrane densities was applied to both struc-tures. The structures were low pass filtered to 3.3-nm resolution and then normalized by subtracting the mean value and subsequently dividing by the root mean square density. The difference between the two normalized maps was calculated using the “DR DIFF” procedure of the SPIDER software package (19). The maps were scaled during this procedure. The standard deviation of the map of the difference between WT and mutant symmetrized global averages was used to determine the contour thresholds for display (41, 53). This sigma was much larger than that computed between odd and even half-data set global averages of WT HIV-1 and thus represents a conservative estimation of signif-icance.
Quasiatomic model building.The HIV-1 atomic model was built based on Protein Data Bank (PDB) entry 3JWD (62) using the MMTSB tool kit http: //feig.bch.msu.edu/mmtsb/Main_Page (17). The V3 loop was added into this structure from PDB 2B4C (26) in the following manner. First, the V3 loop of the gp120 core of 2B4C was extracted along with five extra residues at each end to facilitate the alignment step. The artificial connecting fragment of the V3 loop of 3JWD was then removed. The MMTSB command lsqfit.pl was used to align these two fragments using the sequences in common within the overlapping ends. The two best-aligned residues at both ends of the V3 loop were determined and left unchanged, and the additional V3 loop residues between them were then added into the gp120 core of 3JWD using the convpdb.pl command.
The SIV model was built in a similar fashion based on the PDB 3FUS core structure. The V3 loop was again obtained from PDB 2B4C and aligned as described above. After the aligned V3 loop fragment of 2B4C was placed into the gp120 core of 3FUS, the sequence of the V3 loop from 2B4C was replaced with the corresponding SIV sequence using the command mutate.pl.
The two atomic models were placed manually inside the cryo-ET reconstruc-tions at the envelope cutoff appropriate to a mass of 420 kDa using the find_3d_threshold command in SPARX in conjunction with CHIMERA (25, 65). The gp120 model was arranged so that V3 and the V1/V2 stump were oriented toward the top of the spike and the N and C termini at the bottom. The glycosylated residues were placed facing out and the CD4 molecule (in the liganded model) oriented upward, roughly parallel to the spike axis. The trimer was generated by applying a 3-fold rotation to the individual spike atomic model. For comparison, the atomic model of Liu et al., PDB 3DNN (46), was aligned with our WT density map using CHIMERA (65). Density maps in the figures were displayed using CHIMERA (65).
Accession numbers.The reconstructed three-dimensional (3D) averages have been deposited in the Electron Microscopy Data Bank (EMDB) under accession codes EMD-1813 and EMD-1814.
RESULTS
Data acquisition and processing. Similar to our previous efforts, we used a marker-free alignment method (85) that aligns the individual images of the tilt series based on intrinsic features of the tomogram. The global distribution of spike orientations for the selected spikes of both WT (see Fig. S1 in the supplemental material) and mutant (see Fig. S2 in the supplemental material) virions was not random, showing a slight bias toward spikes extending out the side of the virion and with fewer spikes extending out the top and bottom sur-faces. Each of the class averages reflected this bias but other-wise showed no additional bias with respect to the orientation
on November 7, 2019 by guest
http://jvi.asm.org/
of the tilt axis other than that reflected in the total distribution. This observation indicates that the spike averages derive from the actual structure without a missing wedge bias. The resolu-tion by Fourier shell correlaresolu-tion in the WT individual classes ranged from 4.1 to 4.5 nm and was 3.3 nm in the global average. For the⌬V1/V2 mutant, the values were 4.2 to 4.6 nm for the class averages and 3.3 nm for the global average.
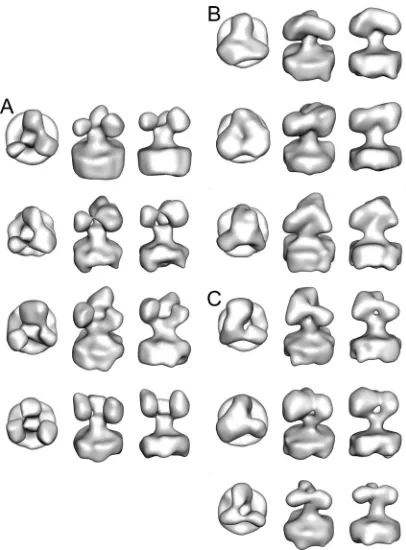
WT HIV-1 spikes.We collected a total of 1,463 spikes from 138 WT R3A virions. On average, each virion had 10.6 spikes, which is in line with our previous EM observations (97, 98) and other data (12). We computed 10 classes from the WT spikes to obtain an estimate of the native variation (Fig. 1). Although the classes are not identical, they are all thematically similar in having a pronounced peak of density at the top, an open interior, and a strong density for the stalk. The stalk broadens as it approaches the bilayer. The minor differences in the WT spike structure could be due to technical issues relating to the comparatively high noise due to low electron counts (the class averages have 10-fold fewer members than the global average) characteristic of cryo-EM tomography or could represent in-herent segmental flexibility.
Because there are few pronounced differences among the WT classes, the global average of all classes can be taken as a representation of the native spike structure (Fig. 2A). The spike dimensions are 14.3 nm high and 12.0 nm wide, which is in general agreement with previous measurements (46, 95, 98). The spike itself has a very open structure with three well-resolved Env protomers. Enforcement of 3-fold symmetry changes the spike appearance very little (Fig. 2B), a further indication of structural homogeneity. The three protomers of the spike come together to form a pronounced “peak” in the center at a point farthest from the membrane. The connection to the membrane is robust and flared. The chirality for the arms is slight.
We have aligned and overlaid our structure on the native spike from the recently published HIV-1 strain BaL structure of Liu et al. (46) to facilitate comparison (Fig. 2C). Our global average is very similar, especially with regard to the openness of the center, but has the following apparent differences. (i) The chirality in our spike structure is very slight compared to that of the Liu et al. structure (46). (ii) The stalk density, which connects the head of the spike to the membrane, is more robust in our structure, and the stalk is more flared at the membrane surface. (iii) The peak of the spike at its furthest point from the membrane appears higher (by 1.2 nm) and more pronounced.
[image:3.585.60.266.69.305.2]V1/V2 deletion mutant. The ⌬V1/V2 mutant was derived from the R3A WT strain by deletion of 71 amino acids of the V1/V2 loop (40). Only the basal disulfide bond and a five-amino-acid linker strand remain. Unlike the WT spike, the spikes of the V1/V2 deletion mutation are quite variable in structure. We obtained a total of 1,860 spikes from 220 mutant virions, for an average of 8.5 spikes per virion, using the same equipment and methods used for WT data acquisition and analysis. Of the⌬V1/V2 mutant spikes, 10% or 156 were dis-carded based on high variance so that the final spike count was 1,674 and more similar to the WT spike count. We computed 10 class averages which, although probably representing a con-tinuum, for ease of discussion, we have divided into three groups (Fig. 3). About 40% of the spike subclasses show the three arms well separated with little connecting density in the “peak” on the 3-fold axis (Fig. 3A). Another 30% of the spike subclasses have the arms coming together, but with some den-sity on the 3-fold axis (Fig. 3B). The remaining 30% are in an intermediate state with a robust connection between two of the arms with the third arm appearing smaller and weakly con-nected to one of the other arms (Fig. 3C). The⌬V1/V2 mutant
[image:3.585.336.506.72.291.2]FIG. 1. Class averages for 10 classes (A to J) from the WT spikes with the top, oblique, and side views.
FIG. 2. WT spike structure shown in top (left), oblique (middle), and side (right) views. (A) Global averages for the native spikes with-out (A) and with (B) symmetry enforced. (C) Fitting between the structure in panel B with the structure from Liu et al. (46) shown in yellow after low-pass filtering to a 33-Å resolution.
on November 7, 2019 by guest
http://jvi.asm.org/
global average and the averages of the three mutant subclasses show that they lack the pronounced peak at the top and have various amounts of connectivity between the gp120 arms (Fig. 4B to E). As with the WT, the class averages for the⌬V1/V2 mutant did not show a strong global orientation bias that could explain these differences (see Fig. S2 in the supplemental ma-terial).
Comparison of the WT and the V1/V2 deletion mutant.
Comparison of the WT and the three mutant subclasses, with or without symmetry enforced, reveals that the single largest difference lies in the height of the peak of density on the 3-fold axis (Fig. 4). As a more quantitative comparison, we computed density difference maps between the global averages of the WT and mutant spikes (Fig. 5) at different standard deviation cut-offs for several different class averages (see Fig. S3 to S6 in the supplemental material). Above aof 2.0, the difference den-sities change little except in size and are similar with or without symmetrization.
The difference maps show significant densities at three lo-cations. Positive differences (red in Fig. 5B) are due to density in the WT spike average that is not present in the mutant average. One of the positive differences is found at the peak of the spike, a pattern consistent with the visual comparison of the mutant and WT spikes (Fig. 4). Another positive difference peak is located at the lower part of each of the arms. When contoured at 2.5, the WT-⌬V1/V2 difference map reveals that there is a loss or displacement of mass from the peak upon
V1/V2 deletion. Another mass reduction is observed in the proximal region of the gp120 spike head (Fig. 5B). These density losses are partially offset by a density gain (negative density in blue, Fig. 5C) in the central void of the spike head. This negative difference represents densities found in the mu-tant spike average but not in the WT spike average. Because the mutant spikes have less mass than the WT and would not have net overall negative differences, the negative densities would be due to conformational differences between WT and mutant spikes. The negative density difference in the center of the spike below the location of the WT peak implies that the gp120 arms and/or elements of gp41 of the mutant can ap-proach closer to the 3-fold axis than is permissible for the WT spike (blue in Fig. 5C).
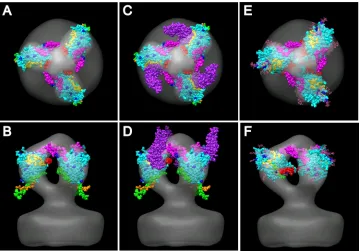
[image:4.585.61.264.67.342.2]Atomic model of the WT spike.We built atomic models into the WT spike average using published crystal structures of gp120. The liganded HIV-1 atomic model was built based on PDB 3JWD, which includes the extended N and C termini (62), onto which the V3 loop from PDB 2B4C (26) was grafted. The unliganded SIV core structure, PDB 3FUS (B. Chen et al., unpublished data) was also used for some fitting exercises. When the composite liganded HIV-1 atomic model (PDB 3JWD plus V3 of 2B4C, Fig. 6) is viewed by looking down the spike axis toward the membrane, the qualitatively best fit was obtained by placing the V3 loop (magenta) at the top pointing clockwise to connect with the adjacent gp120 arm (Fig. 6A). This places the V1/V2 “stump” (red) on the opposite side of the core structure, facing counterclockwise and more proximal to the membrane surface than V3. CD4 (purple), when placed on its binding site, is oriented more perpendicular to the mem-brane than observed in the liganded structure, where it is in a
FIG. 3. Ten class averages from the⌬V1/V2 mutant spikes shown in top (left), oblique (middle), and side (right) views. (A) Four class averages which form an “open” conformation. (B) Three class aver-ages having the “closed” conformation. (C) Three class averaver-ages hav-ing a conformation roughly intermediate between the open and closed conformations.
FIG. 4. Comparison of unsymmetrized and symmetrized class av-erages. (A) The global WT average. (B) The global⌬V1/V2 mutant average. (C) The mutant open subaverage. (D) The mutant closed subaverage. (E) The mutant intermediate subaverage.
on November 7, 2019 by guest
http://jvi.asm.org/
more tangential position (Fig. 6A and B) (46). Placement of the CD4 binding site to the side of the core structure is, however, consistent with the liganded spike structure as deter-mined by cryo-ET (Fig. 6C and D) (46).
This position places the N- and C-terminal peptides (green and orange, respectively) proximal to the membrane but ex-tending outside of the spike envelope. No orientation was found that could place them inside the envelope and at the same time place the V1/V2 stump, the V3 loop, and the CD4 binding site in reasonable positions. The residues that are known to be glycosylated all point toward the outside of the spike envelope; no glycosylated residue faces the adjacent gp120 protomer or the inside of the spike.
Our model is quite similar to the recently published model of liganded HIV-1 (46), which was built from a different start-ing structure (PDB 2NY7) (96). Both models place V3 at the top on one side of the gp120 core and the V1/V2 stump on the other side. Consequently, our V1/V2 stump is slightly higher than that of Liu et al. Our position of V3 is very similar but with theirs rotated upward slightly. The N and C termini of the Liu et al. models, T90 and G492, are located 26 and 29 Å closer to the spike axis, respectively, than in our model (46).
[image:5.585.77.248.69.289.2]Fitting the unliganded SIV atomic model (3FUS), which is devoid of all V loops, into our gp120 WT density map was more problematic. We built on a V3 loop based on the HIV-1 Env structures, but the composite SIV structure is a poor match for the WT spike density. The most reasonable model placed the V3 loop in a position similar to that in the HIV-1 model, at the top oriented clockwise from the core structure
FIG. 5. Map of the differences between the global averages of native and mutant spikes. Global averages have symmetry enforced, and the difference map is shown at a contour threshold of 2.5. (A) Global average of the ⌬V1/V2 mutant spikes. (B) Positive difference map (red) superimposed on the symmetrized global av-erage of the WT spike (gray). (C) Negative-difference map (blue) superimposed on the symmetrized global average of the WT spike (gray).
FIG. 6. Fitting of the HIV/SIV gp120 atomic structures into the symmetry-enforced global average of WT HIV spikes. The V3 loop is magenta; the yellow spheres are the CD4 binding site residues; blue spheres mark the glycosylated residues; white spheres are glycosylated residues near the CD4 binding site; CD4, when shown, is purple; the N-terminal peptide is green; the C-terminal peptide is orange; red marks the base of the⌬V1/V2 loop; and the remainder of the gp120 core is cyan. (A and B) HIV-1 gp120 core (PDB 3JWD) with an added V3 loop (from PDB 2B4C). (C and D) Same as panels A and B but with CD4 (purple) (from PDB 3JWD). (E and F) SIV gp120 core (PDB 3FUS) with an added V3 loop (from PDB 2B4C).
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.113.473.417.668.2]and facing the neighboring gp120 (Fig. 6E and F). The V1/V2 stump then was placed much closer to the membrane and oriented toward the open interior of the spike, where there was little density. The N and C termini were oriented facing the membrane and toward the spike axis. All of the glycosylation sites faced outward, and the residues involved in CD4 binding faced in the same direction as for the HIV-1 model, i.e., coun-terclockwise from the gp120 core structure. The SIV model had distinctly more main chain atoms extending outside of the envelope.
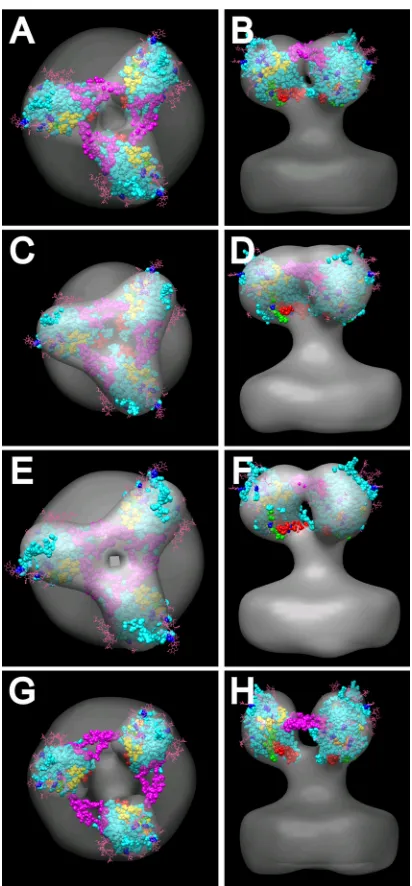
We also built the SIV crystal structure (3FUS), with an added V3 loop as described above, into our⌬V1/V2 averaged mutant spike. Since the SIV crystal structure itself lacks the V1/V2 region, it should be a better match to the shape of the ⌬V1/V2 mutant spike density than the WT density, which turned out to be the case (Fig. 7A and B). Although the spike density lacks much shape at this resolution, the requirements that the glycosylation sites face outward and that the N and C terminals face down restrict fitting options. Because of the relatively close proximity of the V1/V2 loop to the bases of the N and C termini in this atomic model, there was no way of positioning the base of the V1/V2 loop pointing toward the peak while maintaining the downward orientation of the ter-minal peptides. The V3 loop, which we added to the SIV gp120 core structure, could be placed into the density that bridges the individual spike arms. In this orientation, the position of the V1/V2 stump is located toward the bottom facing the inside of the spike and the N and C termini are facing down and con-tained within the envelope. Similar models can be built into the three symmetrized subaverages with reasonable though not identical fits for the key components (Fig. 7C to H). In the case of the open spike average (Fig. 7G and H), the V3 loop falls outside the gp120 core density envelope. If the V3 loop occu-pies such a position, the lack of density between spikes suggests that the loop can be disordered and hence not visible in the class averages.
DISCUSSION
We have directly compared the 3D structures of in situ
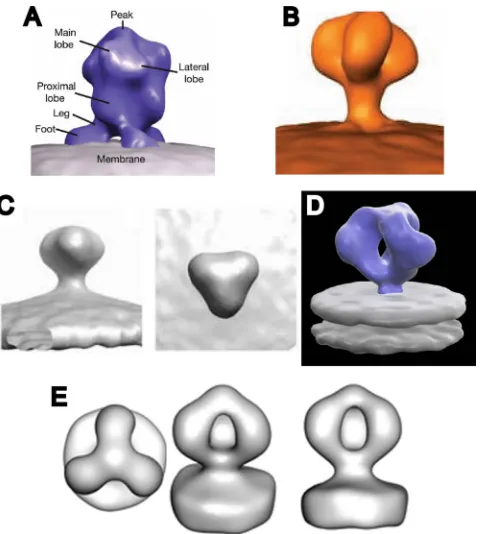
HIV-1 Env spikes in their WT and⌬V1/V2 mutant conforma-tions by cryo-ET. Our WT Env spike reconstruction is similar, though not identical, to a recently published cryo-ET-derived WT spike reconstruction (46). The five prior reports (46, 86, 95, 98, 99) on the structure of virion-associated WT SIV or HIV-1 Env spikes describe two general morphologies (Fig. 8). One type (Fig. 8A to C) has a generally closed conformation in which the presumptive gp120 protomers do not enclose a dis-tinct void at the published contour threshold (86, 95, 98, 99). The other reported structure (46) shows the arms enclosing a pronounced axial void (Fig. 8D). The WT structure reported here shares this characteristic (Fig. 8E). There are, however, several differences between our current WT structure and that of Liu et al. (46), namely, in the height of the central peak, the robustness of the gp41 stalk where it intersects with the lipid bilayer, and the degree of chirality, but otherwise, the two structures are in general agreement. The differences between the two open structures are likely due to the resolution.
What are the bases for these similarities and differences, and how do they impact the interpretation of the current study? In
[image:6.585.318.524.66.509.2]the case of Env spikes, differences in biological samples (e.g., HIV-1 versus SIV) must be considered, as well as the potential for structural heterogeneity resulting from innate subunit flex-ibility. However, there are also experimental factors that vary among the published structures: (i) specimen thickness, (ii) tilt angle increments, (iii) tilt series collection scheme (fixed in-crement or Saxton scheme inin-crements), (iv) microscope, (v) CCD camera, (vi) defocus, (vii) tilt series alignment method (marker or marker-free merging), and (viii) subvolume align-ment, classification, and averaging. All of these experimental
FIG. 7. Fitting of the SIV gp120 atomic structures (PDB 3FUS) with an added V3 loop (from PDB 2B4C) into the symmetry-enforced ⌬V1/V2 loop mutant averages. The color scheme is as follows: gp120 core, cyan; V3 loop, magenta; base of⌬V1/V2 loop, red; N terminus, green; C terminus, orange; CD4 binding site residues, yellow. Glyco-sylation is pink, and glycosylated residues are blue. A, C, E, and G, top views; B, D, F, and H, side views; A and B, global average; C and D, “closed” subaverage; E and F, intermediate subaverage; G and H, “open” subaverage.
on November 7, 2019 by guest
http://jvi.asm.org/
factors affect the resolution of the final average 3D spike struc-ture in a complicated way. The most likely variables are the specimen thickness and the tilt increment; all of the closed structures (86, 95, 98, 99) used a larger, 2 to 3°, tilt increment and were obtained from thicker specimens. Both thickness and tilt increment affect the resolution in raw tomograms in direc-tions other than the tilt axis.
Because the details of the final averaged images are so dependent upon technical parameters, we collected and pro-cessed the WT and⌬V1/V2 data for the current study in the same manner so that any variance in the resulting images could be ascribed to differences in molecular structure.
An obvious question raised by our findings is whether the peak which is present in the WT spike and missing from the ⌬V1/V2 spike comprises the V1/V2 loops. Our gp120 trimer models (Fig. 6A and B), based on extrapolation from X-ray crystallography data from soluble CD4-liganded gp120 core structures (89), show the V3 loop (magenta) occupying this general position. In this orientation, the base of the V1/V2 loop (red) is positioned toward the medial axis and pointing counterclockwise into unoccupied space. However, because of the significant mass and unknown configuration of the V1/V2 loop, it could conceivably curve upward to both interact with V3 and contribute to the peak density. The major caveat here is that we are fitting a liganded fusion intermediate structure into an unliganded prefusion density, and major structural rearrangement is known to accompany ligand binding (62).
A published trimer model that consists of unliganded (SIV) gp120 atomic cores places the stems of the V3 and V1/V2
loops (the loops are not present) pointing in nearly opposite directions (11). The V1/V2 loop in this model is suspected of reaching laterally to interact with the V3 loop from a neigh-boring protomer to help stabilize the spike. If this aspect of the SIV trimer model is correct, and if the HIV-1 prefusion spike is similarly configured, then our data indicate that the loss of V1/V2 would destabilize the basolateral coupling of the pro-tomers by V1/V2 and that this loss would permit enhanced subunit flexibility and, perhaps, V3 loop reorientation.
There is ample evidence for an interaction between the V1/V2 and V3 loops, including proposed molecular shielding of V3 epitopes by the V1/V2 loop (7, 13, 31, 35, 36, 56, 61, 63, 72, 78, 90). Consistent with this view are the characteristics of several broadly neutralizing monoclonal antibodies, including 2909, PG9, and PG16, that appear to target conformational epitopes consisting of V2 and V3 elements (20, 81).
In our previous cryo-ET studies involving SIV Env (98), the optimum fitting of the SIV prefusion monomer atomic struc-ture (11) into the averaged spike density positioned the V3 stalk pointed laterally and the V1/V2 stalk and the presumed mass of the V1/V2 loop pointed basolaterally, where it would be in a position to interact with neighboring protomers. In the Liu et al. cryo-ET spike reconstruction (86), which most closely resembles our current reconstruction, the authors were unable to fit the SIV unliganded model of Chen et al. into their density, choosing instead to use the CD4-liganded configura-tion, which displayed a better overall fit. In the Liu et al. model, as with a crystallography-based model of the liganded core plus V3 (26), the V3 loop constituted the peak and V1/V2 projected upward with a clockwise chirality. However, as indi-cated above, considerable evidence suggests a large-scale re-arrangement of the protein structure of gp120 following ligand binding (37, 57, 60, 62). Consequently, fitting exercises using liganded atomic structures with unliganded cryo-ET density maps can potentially misinform.
Despite these caveats, we attempted to fit both liganded and unliganded models into our WT and⌬V1/V2 mutant densities. The unliganded SIV core atomic structure, which lacks the V1/V2 loop (11) but contains a computationally grafted V3 loop, should be a good fit to the arms of the⌬V1/V2 mutant spikes, and this proved to be the case (Fig. 7). Although at the resolution of our cryo-ET reconstruction, the presumptive gp120 density lacks a pronounced shape, placement of the glycosylation sites to the outside and the N and C termini toward the membrane limits the range of possible orientations. The best placement positions the grafted V3 loop (magenta) so as to connect the gp120 protomers. The V1/V2 stump (red) is positioned toward the base of the gp120 protomer, where it could also reach neighboring protomers. However, in this po-sition, a direct contribution by V1/V2 to peak mass observed in the WT spike seems highly unlikely. When the composite li-ganded core structure, which included the V3 loop (26) and N-and C-terminal peptides (62), is fitted into the WT spike, a similar placement of the V3 loop is obtained but the base of the V1/V2 loop is positioned higher in the trimer and could conceivably both interact with V3 from a neighboring pro-tomer and extend upward to contribute to the peak density (Fig. 6A and B). But while the outcome of this fitting exercise may be satisfying, as indicated above, such attempts to place liganded atomic models into unliganded densities may be
in-FIG. 8. Comparison of published structures for the Env spikes from intact HIV and SIV virions. (A) SIV spikes from reference 98. (B) SIV spikes from reference 95. (C) HIV spikes from reference 99. (D) HIV spikes from reference 46. (E) HIV spikes from the present work.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.45.285.67.334.2]appropriate based on the current understanding of the struc-tural dynamics associated with CD4 binding.
It has previously been proposed that there is a certain amount of structural disorder not only in the variable loops but also within the structural core of the mature spike. This inher-ent inher-entropy would serve to frustrate antibody binding and neutralization by a mechanism dubbed “entropic” or “confor-mational masking” (37, 57, 60, 62, 93). All of the 10 classes derived from the WT data in our study show similar charac-teristics, though some minor variations can be noted (Fig. 1). It is not clear whether these low-level structural variations reflect actual structural heterogeneity or the inherent “noise” that is characteristic of cryo-ET. The three arms in the global average of WT spikes are similar and highly symmetric even before symmetrization (Fig. 2A). Because of the relative homogeneity of the cryo-ET WT classes, entropic movements may be so small as to be below the level of detection by our classification scheme.
In marked contrast to the WT, deletion of the 71-amino-acid V1/V2 loop has a dramatic effect on not only the overall struc-ture of the Env spike but also on its degree of structural heterogeneity (compare Fig. 3 to Fig. 1). The most obvious differences are the loss of density at the peak of the⌬V1/V2 mutant (compare the WT averaged image in Fig. 4A to the mutant average in Fig. 4B) and the considerable structural heterogeneity displayed by the various classes of the mutant (Fig. 3 and 4C to E). The increased heterogeneity is most probably due to greatly enhanced structural flexibility. If the V1/V2 loop deletion does not lead to structural reorganization of components with respect to the WT structure, then the simplest interpretation would be that the highlighted density at the peak indicates the location of a convergent set of three V1/V2 loops in the native structure. Because the basolateral positive density is also of a fairly significant mass, it could, in addition or instead, represent V1/V2 density. Alternatively, the data could be interpreted as indicating considerable conforma-tional changes in other gp120 elements resulting from V1/V2 loop deletion and/or greatly enhanced subunit flexibility, in which case the V1/V2 loop may not necessarily contribute directly to either of the positive densities.
The strategy of using constructs in which all or a portion of the V1/V2 loop has been deleted as a vaccine has been exten-sively explored because of the enhanced exposure of conserved potentially broadly cross-reactive epitopes associated with such constructs (1, 2, 18, 22, 32, 43, 76, 77). Our data suggest that although removal of the bulky V1/V2 loop may lead to epitope exposure by direct unmasking (7, 31, 35, 61, 78, 90), the greatly enhanced flexibility imparted to the gp120 subunits in the ⌬V1/V2 trimer likely plays a significant role in this process.
Several of the ⌬V1/V2 mutant class averages show gp120 subunits which appear to have lost contact with one another at the apical peak (Fig. 3A and C and 4C and E), raising the question of what is holding the spike together under such circumstances. A reasonable assumption is that either the re-sidual proximolateral gp120-gp120 interactions of the gp120 protomers (39, 50) with one another in⌬V1/V2 are sufficient to prevent gp120 loss or, in the absence of such interactions, that gp41-gp41 interactions within the⌬V1/V2 trimer are suf-ficient to stabilize the spike structure. In the latter case, the
gp120 protomers would remain in place solely because of their interaction with gp41 (23, 88, 92).
It is generally assumed that gp41-gp41 interactions contrib-ute significantly to spike structure assembly and stability (10, 47, 48, 83). However, much of the data underlying this assump-tion derive from structural studies of trimeric forms likely in the fusion intermediate state or the analysis of MPER peptides unconstrained by Env spike structure. Because of the consid-erable structural rearrangements associated with the transition from the prefusion to the fusion intermediate state, these data may have little bearing on the structure and stability of gp41 in the native spike.
The role of gp120-gp120 interactions in spike stabilization is not well defined, even though such interactions have been considered (8, 11, 39, 50). It is worth noting that efforts to produce soluble Env spikes comprising gp120 and the ectodo-main of gp41 have been fraught with issues of stability (re-viewed in references 63 and 66). One possible interpretation of these data is that intramembrane or cytoplasmic interactions, lacking in such constructs, make a significant contribution to spike stability, perhaps more so than gp120 or gp41 ectodo-main interactions (86, 94). On the other hand, the observations that soluble recombinant ⌬V1/V2 mutant constructs can be less stable than WT constructs and replication-competent ⌬V1/V2 mutant spikes are less stable than virion-associated WT Env spikes point to at least some direct or indirect stabi-lizing effect imparted by the V1/V2 loop (6). Interestingly, under selective pressure, HIV-1, HIV-2, and SIV⌬V1/V2 mu-tants can revert to replication competence and trimer stability through mutations in other regions of gp120 and even in gp41, implying that enhancement in other contact regions can com-pensate for the loss of V1/V2 loop structural interactions (6, 31, 45, 72, 87). It would be interesting to determine if reversion to enhanced fusion competence in such situations is concom-itant with the reestablishment of a more native structure and/or reduced gp120 flexibility. It is worth noting that one recombinant gp140 construct, designated SOSIP, does form stable soluble trimers which, by negative-stain EM, have reg-ular 3-fold symmetry and bind CD4 in a manner similar to that of WT spikes (29, 33). These constructs are stabilized by the addition of a disulfide bond bridging the gp120 and gp41 sub-units and have a point mutation in gp41 which enhances the affinity between the gp41 ectodomains (71, 74).
Moore et al. have provided considerable evidence for the expression of 3-fold axially dissociated spike elements (gp120-gp41 monomers and dimers) on HIV-1 virus-like particles (55). Other evidence suggests a similar presence of gp120-gp41 monomers on virions (9, 59, 68). Whether these forms reflect an equilibrium between intact and dissociated spike elements on virion surfaces (4) or the incorporation of incompletely assembled spike elements is not clear (55). If the association of the three gp120-gp41 subunits is weaker in the⌬V1/V2 mutant than in the WT spikes, the mutant form might be expected to dissociate more readily into gp120-gp41 dimers and mono-mers. However, it seems likely that any differences between the WT and the⌬V1/V2 mutant in this respect is not great since we observed that both WT and⌬V1/V2 virions displayed sim-ilar average numbers of intact Env spikes per virion (10.6 and 8.5, respectively). It should be noted that any monomer and dimer spike elements would likely not have been selected or
on November 7, 2019 by guest
http://jvi.asm.org/
even detected in our study because of their reduced mass. Had they been inadvertently selected, they would have segregated into separate distinct classes in our classification scheme, which was not the case.
While it is tempting to ascribe the enhanced flexibility of gp120 in⌬V1/V2 spikes to loss of the stabilizing influence of apical interactions, an alternative possibility is that the peak in the WT spike is formed by three gp120 loop projections which, though held in close proximity by inherent rigidity within gp120, do not actually form (significant) molecular bonds. In this scenario, V1/V2 would serve to stabilize the upward-arch-ing form of the gp120 protomers and/or stabilize basal (prox-imal) gp120-gp120 and/or gp120-gp41 associations either di-rectly or indidi-rectly through rigidification of V1/V2-associated elements.
ACKNOWLEDGMENTS
This work was supported by NIH grant 2R01 AI055461.
Virus samples were generously supplied by James Hoxie and the AIDS Vaccine Program (SAIC Frederick, NCI, Frederick, MD). We thank Elena Chertova and Julian Bess, Jr., for production and bio-chemical analyses of the viruses and Zhong Huang for discussions and suggestions.
REFERENCES
1.Barnett, S. W., et al.2001. The ability of an oligomeric human immunode-ficiency virus type 1 (HIV-1) envelope antigen to elicit neutralizing antibod-ies against primary HIV-1 isolates is improved following partial deletion of
the second hypervariable region. J. Virol.75:5526–5540.
2.Barnett, S. W., I. K. Srivastava, J. B. Ulmer, J. J. Donnelly, and R. Rappuoli.
2005. Development of V2-deleted trimeric envelope vaccine candidates from human immunodeficiency virus type 1 (HIV-1) subtypes B and C. Microbes
Infect.7:1386–1391.
3.Beddows, S., et al.2005. Evaluating the immunogenicity of a disulfide-stabilized, cleaved, trimeric form of the envelope glycoprotein complex of
human immunodeficiency virus type 1. J. Virol.79:8812–8827.
4.Bewley, C. A., J. M. Louis, R. Ghirlando, and G. M. Clore.2002. Design of a novel peptide inhibitor of HIV fusion that disrupts the internal trimeric
coiled-coil of gp41. J. Biol. Chem.277:14238–14245.
5.Binley, J. M., et al.2000. A recombinant human immunodeficiency virus type 1 envelope glycoprotein complex stabilized by an intermolecular disulfide bond between the gp120 and gp41 subunits is an antigenic mimic of the
trimeric virion-associated structure. J. Virol.74:627–643.
6.Bontjer, I., et al.2009. Optimization of human immunodeficiency virus type 1 envelope glycoproteins with V1/V2 deleted, using virus evolution. J. Virol.
83:368–383.
7.Cao, J., et al.1997. Replication and neutralization of human immunodefi-ciency virus type 1 lacking the V1 and V2 variable loops of the gp120
envelope glycoprotein. J. Virol.71:9808–9812.
8.Center, R. J., P. L. Earl, J. Lebowitz, P. Schuck, and B. Moss.2000. The human immunodeficiency virus type 1 gp120 V2 domain mediates
gp41-independent intersubunit contacts. J. Virol.74:4448–4455.
9.Center, R. J., et al.2002. Oligomeric structure of the human
immunodefi-ciency virus type 1 envelope protein on the virion surface. J. Virol.76:7863–
7867.
10.Chan, D. C., D. Fass, J. M. Berger, and P. S. Kim.1997. Core structure of
gp41 from the HIV envelope glycoprotein. Cell89:263–273.
11.Chen, B., et al.2005. Structure of an unliganded simian immunodeficiency
virus gp120 core. Nature433:834–843.
12.Chertova, E., et al.2002. Envelope glycoprotein incorporation, not shedding of surface envelope glycoprotein (gp120/SU), is the primary determinant of SU content of purified human immunodeficiency virus type 1 and simian
immunodeficiency virus. J. Virol.76:5315–5325.
13.Davis, K. L., et al.2009. Human immunodeficiency virus type 2 (HIV-2)/ HIV-1 envelope chimeras detect high titers of broadly reactive HIV-1
V3-specific antibodies in human plasma. J. Virol.83:1240–1259.
14.Derby, N. R., et al.2006. Antibody response elicited in macaques immunized with human immunodeficiency virus type 1 (HIV-1) SF162-derived gp140 envelope immunogens: comparison with those elicited during homologous simian/human immunodeficiency virus SHIVSF162P4 and heterologous
HIV-1 infection. J. Virol.80:8745–8762.
15.Doms, R. W., and J. P. Moore.2000. HIV-1 membrane fusion: targets of
opportunity. J. Cell Biol.151:F9–F13.
16.Edinger, A. L., et al.2000. Characterization and epitope mapping of
neu-tralizing monoclonal antibodies produced by immunization with oligomeric
simian immunodeficiency virus envelope protein. J. Virol.74:7922–7935.
17.Feig, M., J. Karanicolas, and C. L. Brooks III.2004. MMTSB Tool Set: enhanced sampling and multiscale modeling methods for applications in
structural biology. J. Mol. Graph. Model.22:377–395.
18.Forsell, M. N., W. R. Schief, and R. T. Wyatt.2009. Immunogenicity of
HIV-1 envelope glycoprotein oligomers. Curr. Opin. HIV AIDS4:380–387.
19.Frank, J., et al.1996. Spider and Web: processing and visualization of images
in 3D electron microscopy and related fields. J. Struct. Biol.116:190–199.
20.Gorny, M. K., et al.2005. Identification of a new quaternary neutralizing epitope on human immunodeficiency virus type 1 virus particles. J. Virol.
79:5232–5237.
21.Grundner, C., et al.2005. Analysis of the neutralizing antibody response elicited in rabbits by repeated inoculation with trimeric HIV-1 envelope
glycoproteins. Virology331:33–46.
22.Gzyl, J., et al.2004. Effect of partial and complete variable loop deletions of the human immunodeficiency virus type 1 envelope glycoprotein on the
breadth of gp160-specific immune responses. Virology318:493–506.
23.Helseth, E., U. Olshevsky, C. Furman, and J. Sodroski.1991. Human im-munodeficiency virus type 1 gp120 envelope glycoprotein regions important
for association with the gp41 transmembrane glycoprotein. J. Virol.65:2119–
2123.
24.Ho, D. D., et al.1991. Another discontinuous epitope on glycoprotein gp120 that is important in human immunodeficiency virus type 1 neutralization is
identified by a monoclonal antibody. Proc. Natl. Acad. Sci. U. S. A.88:8949–
8952.
25.Hohn, M., et al.2007. SPARX, a new environment for cryo-EM image
processing. J. Struct. Biol.157:47–55.
26.Huang, C., et al.2005. Structure of a V3-containing HIV-1 gp120 core.
Science310:1025–1028.
27.Huang, C. C., et al.2007. Structures of the CCR5 N terminus and of a
tyrosine-sulfated antibody with HIV-1 gp120 and CD4. Science317:1930–
1934.
28.Hwang, S. S., T. J. Boyle, H. K. Lyerly, and B. R. Cullen.1992. Identification of envelope V3 loop as the major determinant of CD4 neutralization
sensi-tivity of HIV-1. Science257:535–537.
29.Iyer, S. P. N., et al.2007. Purified proteolytically mature HIV type 1 SOSIP
gp140 envelope trimers. AIDS Res. Hum. Retroviruses23:817–828.
30.Jin, T., F. Guo, Y. W. Chen, A. Howard, and Y. Z. Zhang.2009. Crystal
structure of Ara h 3, a major allergen in peanut. Mol. Immunol.46:1796–
1804.
31.Johnson, M. J. W. E., et al.2002. A replication-competent, neutralization-sensitive variant of simian immunodeficiency virus lacking 100 amino acids of
envelope. J. Virol.76:2075–2086.
32.Kang, S. M., et al.2005. Modified HIV envelope proteins with enhanced
binding to neutralizing monoclonal antibodies. Virology331:20–32.
33.Kang, Y. K., et al.2009. Structural and immunogenicity studies of a cleaved,
stabilized envelope trimer derived from subtype A HIV-1. Vaccine27:5120–
5132.
34.Koito, A., G. Harrowe, J. A. Levy, and C. Cheng-Mayer.1994. Functional role of the V1/V2 region of human immunodeficiency virus type 1 envelope glycoprotein gp120 in infection of primary macrophages and soluble CD4
neutralization. J. Virol.68:2253–2259.
35.Kolchinsky, P., E. Kiprilov, P. Bartley, R. Rubinstein, and J. Sodroski.2001. Loss of a single N-linked glycan allows CD4-independent human immuno-deficiency virus type 1 infection by altering the position of the gp120 V1/V2
variable loops. J. Virol.75:3435–3443.
36.Krachmarov, C., et al.2005. Antibodies that are cross-reactive for human immunodeficiency virus type 1 clade A and clade B V3 domains are common in patient sera from Cameroon, but their neutralization activity is usually
restricted by epitope masking. J. Virol.79:780–790.
37.Kwong, P. D., et al.2002. HIV-1 evades antibody-mediated neutralization
through conformational masking of receptor-binding sites. Nature420:678–
682.
38.Kwong, P. D., et al.1998. Structure of an HIV gp120 envelope glycoprotein in complex with the CD4 receptor and a neutralizing human antibody.
Nature393:648–659.
39.Kwong, P. D., R. Wyatt, Q. J. Sattentau, J. Sodroski, and W. A. Hendrickson.
2000. Oligomeric modeling and electrostatic analysis of the gp120 envelope
glycoprotein of human immunodeficiency virus. J. Virol.74:1961–1972.
40.Laakso, M. M., et al.2007. V3 loop truncations in HIV-1 envelope impart resistance to coreceptor inhibitors and enhanced sensitivity to neutralizing
antibodies. PLoS Pathog.3:e117.
41.Lake, J. A., and K. R. Leonard.1974. Structure and protein distribution for
the capsid ofCaulobacter crescentusbacteriophageCbK. J. Mol. Biol.86:
499–518.
42.LaRosa, G. J., et al.1990. Conserved sequence and structural elements in the
HIV-1 principal neutralizing determinant. Science249:932–935.
43.Letvin, N. L., et al.2006. Preserved CD4⫹central memory T cells and
survival in vaccinated SIV-challenged monkeys. Science312:1530–1533.
44.Li, M., et al.2005. Human immunodeficiency virus type 1 env clones from
on November 7, 2019 by guest
http://jvi.asm.org/
acute and early subtype B infections for standardized assessments of
vaccine-elicited neutralizing antibodies. J. Virol.79:10108–10125.
45.Lin, G., et al.2007. Replication-competent variants of human immunodefi-ciency virus type 2 lacking the V3 loop exhibit resistance to chemokine
receptor antagonists. J. Virol.81:9956–9966.
46.Liu, J., A. Bartesaghi, M. J. Borgnia, G. Sapiro, and S. Subramaniam.2008.
Molecular architecture of native HIV-1 gp120 trimers. Nature455:109–113.
47.Liu, J., Y. Deng, A. K. Dey, J. P. Moore, and M. Lu.2009. Structure of the HIV-1 gp41 membrane-proximal ectodomain region in a putative prefusion
conformation. Biochemistry48:2915–2923.
48.Lu, M., S. C. Blacklow, and P. S. Kim.1995. A trimeric structural domain of
the HIV-1 transmembrane glycoprotein. Nat. Struct. Biol.2:1075–1082.
49.Ly, A., and L. Stamatatos.2000. V2 loop glycosylation of the human immuno-deficiency virus type 1 SF162 envelope facilitates interaction of this protein with CD4 and CCR5 receptors and protects the virus from neutralization by anti-V3
loop and anti-CD4 binding site antibodies. J. Virol.74:6769–6776.
50.Malvoisin, E., M. P. Kieny, and F. Wild.1997. Self-association of truncated
forms of HIV-1 gp120. Virus Res.49:163–172.
51.Mascola, J. R., et al.2005. Recommendations for the design and use of standard virus panels to assess neutralizing antibody responses elicited by candidate
human immunodeficiency virus type 1 vaccines. J. Virol.79:10103–10107.
52.Meissner, E. G., K. M. Duus, F. Gao, X. F. Yu, and L. Su.2004. Character-ization of a thymus-tropic HIV-1 isolate from a rapid progressor: role of the
envelope. Virology328:74–88.
53.Mikhailov, A. M., and B. K. Vainshtein.1971. Electron microscope deter-mination of the three-dimensional structure of the extended tail of the T6
bacteriophage. Soviet Phys. Crystallogr.16:428–436.
54.Moore, J. P., and J. Sodroski.1996. Antibody cross-competition analysis of the human immunodeficiency virus type 1 gp120 exterior envelope
glycop-rotein. J. Virol.70:1863–1872.
55.Moore, P. L., et al.2006. Nature of nonfunctional envelope proteins on the
surface of human immunodeficiency virus type 1. J. Virol.80:2515–2528.
56.Morikita, T., et al.1997. The V1/V2 region of human immunodeficiency virus type 1 modulates the sensitivity to neutralization by soluble CD4 and
cellular tropism. AIDS Res. Hum. Retroviruses13:1291–1299.
57.Myszka, D. G., et al. 2000. Energetics of the HIV gp120-CD4 binding
reaction. Proc. Natl. Acad. Sci. U. S. A.97:9026–9031.
58.Nabatov, A. A., et al.2004. Intrapatient alterations in the human immuno-deficiency virus type 1 gp120 V1V2 and V3 regions differentially modulate coreceptor usage, virus inhibition by CC/CXC chemokines, soluble CD4, and
the b12 and 2G12 monoclonal antibodies. J. Virol.78:524–530.
59.Owens, R. J., and R. W. Compans.1990. The human immunodeficiency virus type 1 envelope glycoprotein precursor acquires aberrant intermolecular disulfide bonds that may prevent normal proteolytic processing. Virology
179:827–833.
60.Pan, Y., B. Ma, and R. Nussinov.2005. CD4 binding partially locks the bridging sheet in gp120 but leaves the B2/3 strands flexible. J. Mol. Biol.
350:514–527.
61.Pancera, M., et al.2005. Soluble mimetics of human immunodeficiency virus type 1 viral spikes produced by replacement of the native trimerization domain with a heterologous trimerization motif: characterization and ligand
binding analysis. J. Virol.79:9954–9969.
62.Pancera, M., et al.2010. Structure of HIV-1 gp120 with gp41-interactive region reveals layered envelope architecture and basis of conformational
mobility. Proc. Natl. Acad. Sci. U. S. A.107:1166–1171.
63.Pantophlet, R., and D. R. Burton.2006. Gp120: target for neutralizing HIV-1
antibodies. Annu. Rev. Immunol.24:739–769.
64.Parren, P. W., J. P. Moore, D. R. Burton, and Q. J. Sattentau.1999. The neutralizing antibody response to HIV-1: viral evasion and escape from
humoral immunity. AIDS13:S137–S162.
65.Pettersen, E. F., et al.2004. UCSF Chimera—a visualization system for
exploratory research and analysis. J. Comput. Chem.25:1605–1612.
66.Phogat, S., and R. Wyatt.2007. Rational modifications of HIV-1 envelope
glycoproteins for immunogen design. Curr. Pharm. Des.13:213–227.
67.Pinter, A., et al.2004. The V1/V2 domain of gp120 is a global regulator of the sensitivity of primary human immunodeficiency virus type 1 isolates to neutralization by antibodies commonly induced upon infection. J. Virol.
78:5205–5215.
68.Poignard, P., et al.2003. Heterogeneity of envelope molecules expressed on primary human immunodeficiency virus type 1 particles as probed by the
binding of neutralizing and nonneutralizing antibodies. J. Virol.77:353–365.
69.Recht, B., B. Frangione, E. Franklin, and E. Loghem.1981. Structural stud-ies of a human gamma3 myeloma protein (Goe) that binds staph protein A.
J. Immunol.127:917–923.
70.Rusche, J. R., et al.1988. Antibodies that inhibit fusion of human immuno-deficiency virus-infected cells bind a 24-amino acid sequence of the viral
envelope, gp120. Proc. Natl. Acad. Sci. U. S. A.85:3198–3202.
71.Sanders, R., et al.2002. Stabilization of the soluble, cleaved, trimeric form of the envelope glycoprotein complex of human immunodeficiency virus type 1.
J. Virol.76:8875–8889.
72.Saunders, C. J., et al.2005. The V1, V2, and V3 regions of the human
immunodeficiency virus type 1 envelope differentially affect the viral
pheno-type in an isolate-dependent manner. J. Virol.79:9069–9080.
73.Saxton, W. O., W. Baumeister, and M. Hahn.1984. Three-dimensional
recon-struction of imperfect two-dimensional crystals. Ultramicroscopy13:57–70.
74.Schu¨lke, N., et al.2002. Oligomeric and conformational properties of a proteolytically mature, disulfide-stabilized human immunodeficiency virus
type 1 gp140 envelope glycoprotein. J. Virol.76:7760–7776.
75.Schweighardt, B., et al.2007. Development of an HIV-1 reference panel of subtype B envelope clones isolated from the plasma of recently infected
individuals. J. Acquir. Immune Defic. Syndr.46:1–11.
76.Srivastava, I. K., et al.2008. Comparative evaluation of trimeric envelope glycoproteins derived from subtype C and B HIV-1 R5 isolates. Virology
372:273–290.
77.Srivastava, I. K., K. VanDorsten, L. Vojtech, S. W. Barnett, and L. Stama-tatos.2003. Changes in the immunogenic properties of soluble gp140 human immunodeficiency virus envelope constructs upon partial deletion of the
second hypervariable region. J. Virol.77:2310–2320.
78.Stamatatos, L., and C. Cheng-Mayer.1998. An envelope modification that renders a primary, neutralization-resistant clade B human immunodeficiency virus type 1 isolate highly susceptible to neutralization by sera from other
clades. J. Virol.72:7840–7845.
79.Stanisz, A. M., R. Lieberman, and J. M. Davie.1984. IgA polymorphism in mice: individual light chains can pair covalently with some alpha heavy chains
and non-covalently with others. Mol. Immunol.21:815–818.
80.Sullivan, N., et al.1998. CD4-induced conformational changes in the human immunodeficiency virus type 1 gp120 glycoprotein: consequences for virus
entry and neutralization. J. Virol.72:4694–4703.
81.Walker, L. M., et al.2009. Broad and potent neutralization antibodies from
an African donor reveal a new HIV-1 vaccine target. Science326:285–289.
82.Wei, X., et al.2003. Antibody neutralization and escape by HIV-1. Nature
422:307–312.
83.Weissenhorn, W., A. Dessen, S. C. Harrison, J. J. Skehel, and D. C. Wiley.
1997. Atomic structure of the ectodomain from HIV-1 gp41. Nature387:
426–430.
84.Winkler, H., and K. A. Taylor. 1999. Multivariate statistical analysis of three-dimensional cross-bridge motifs in insect flight muscle.
Ultramicros-copy77:141–152.
85.Winkler, H., and K. A. Taylor.2006. Accurate marker-free alignment with simultaneous geometry determination and reconstruction of tilt series in
electron tomography. Ultramicroscopy106:240–254.
86.Winkler, H., et al.2009. Tomographic subvolume alignment and subvolume classification applied to myosin V and SIV envelope spikes. J. Struct. Biol.
165:64–77.
87.Wolk, T., and M. Schreiber.2006.N-Glycans in the gp120 V1/V2 domain of the HIV-1 strain NL4-3 are indispensable for viral infectivity and resistance
against antibody neutralization. Med. Microbiol. Immunol.195:165–172.
88.Wyatt, R., et al.1997. Analysis of the interaction of the human immunode-ficiency virus type 1 gp120 envelope glycoprotein with the gp41
transmem-brane glycoprotein. J. Virol.71:9722–9731.
89.Wyatt, R., et al.1998. The antigenic structure of the HIV gp120 envelope
glycoprotein. Nature393:705–711.
90.Wyatt, R., et al.1995. Involvement of the V1/V2 variable loop structure in the exposure of human immunodeficiency virus type 1 gp120 epitopes
in-duced by receptor binding. J. Virol.69:5723–5733.
91.Yang, X., et al.2002. Highly stable trimers formed by human immunodefi-ciency virus type 1 envelope glycoproteins fused with the trimeric motif of T4
bacteriophage fibritin. J. Virol.76:4634–4642.
92.York, J., and J. H. Nunberg.2004. Role of hydrophobic residues in the central ectodomain of gp41 in maintaining the association between human immunodeficiency virus type 1 envelope glycoprotein subunits gp120 and
gp41. J. Virol.78:4921–4926.
93.Yuan, W., J. Bazick, and J. Sodroski.2006. Characterization of the multiple conformational states of free monomeric and trimeric human immunodefi-ciency virus envelope glycoproteins after fixation by cross-linker. J. Virol.
80:6725–6737.
94.Yue, L., L. Shang, and E. Hunter. 2009. Truncation of the membrane-spanning domain of human immunodeficiency virus type 1 envelope glyco-protein defines elements required for fusion, incorporation, and infectivity.
J. Virol.83:11588–11598.
95.Zanetti, G., J. A. G. Briggs, K. Grunewald, Q. Sattentau, and S. D. Fuller.
2006. Cryo-electron tomographic structure of an immunodeficiency virus
envelope complexin situ. PLoS Pathog.2:e83.
96.Zhou, T., et al.2007. Structural definition of a conserved neutralization
epitope on HIV-1 gp120. Nature445:732–737.
97.Zhu, P., et al.2003. Electron tomography analysis of envelope glycoprotein trimers on HIV and simian immunodeficiency virus virions. Proc. Natl. Acad.
Sci. U. S. A.100:15812–15817.
98.Zhu, P., et al.2006. Distribution and three-dimensional structure of AIDS
virus envelope spikes. Nature441:847–852.
99.Zhu, P., H. Winkler, E. Chertova, K. A. Taylor, and K. H. Roux.2008. Cryoelectron tomography of HIV-1 envelope spikes: further evidence for
tripod-like legs. PLoS Pathog.4:e1000203.