JOURNALOFVIROLOGY,Apr. 1994,p. 2708-2719 Vol.68, No.4 0022-538X/94/$04.00+0
Copyright © 1994,AmericanSociety for Microbiology
The
Bell
Protein
of
Human
Foamy
Virus Contains One
Positive
and
Two
Negative Control
Regions
Which Regulate
a
Distinct
Activation Domain of
30 Amino
Acids
CHANG WOO LEE,1 JUN
CHANG,'
KI JEONGLEE,'
AND YOUNG CHULSUNG'
2* Departmentof LifeScience,'
CenterforBiofunctional Molecules,2 Pohang University ofScience and Technology, Pohang 790-784, Korea Received 12October1993/Accepted 14 January 1994
The Belltransactivatorisessentialfor thereplicationofhumanfoamyvirus(HFV).To define the functional domainsof HFVBell,wegenerated random missense mutations throughout the entire coding sequence of Bell. Functional
analyses
of 24 missense mutationshaverevealed the presence ofatleasttwofunctional domains in Bell. One domain corresponds to abasic amino acid-rich motif which acts as abipartite nuclear targeting sequence.Asecond,central domaincorrespondsto apresumedeffectorregion which, when mutated, leads to dominant-negative mutants and/or lacks transactivating ability. In addition, deletion analyses and domain-swappingexperiments further showedthatBellprotein containsastrongcarboxy-terminal activation domain. The activating region is also capable offunctioning as a transcription-activating domain in yeast cells,althoughit does not bearanysignificant sequencehomologytothewell-characterized acidic activation domain whichisknowntofunction
only
inyeastandmammaliancells. We alsodemonstrated thattheregionsof Bell fromresidues 1 to 76and from residues 153 to 225repressed transcriptional activation exertedbythe Bell activation domain.In contrast,theregionfrom residues 82 to 150appears to overcome an inhibitory effect. Theseresults indicate that Bell contains onepositiveand twonegative regulatory domains that modulate a distinct activation domain ofBell. These regulatorydomains of Bell cannot affect thefunction of theVP16 activation domain, suggesting that these domains specifically regulate the activation domain of Bell. Furthermore, in vivocompetitionexperiments showedthat thepositive regulatorydomain acts in trans.Thus, ourresultsdemonstratethatBell-mediated transactivation appearstoundergoacomplexregulatorypathway whichprovides a novelmode ofregulation foratranscriptional activation domain.Replication of human foamy virus (HFV) requires the functional expression of the viral nuclear regulatory protein,
Bell (23, 45). The Bell protein is a potent transactivator required for transcription from theHFVlong terminal repeat
(LTR)and the humanimmunodeficiency virus type 1 (HIV-1) LTRpromoter(22,25,27).Inboth theHFVand HIV-1 LTRs, the targetsequencefor Bellwasmappedto the U3 sequence of the LTR upstream of the transcription initiation site, indicating that Bell functions via a DNA element. Transacti-vation of Bell is mediated by a mechanism which increases the steady-state level ofviral transcripts(45, 46). This property of HFVBell is reminiscent of the HIV-1 Tat and human T-cell leukemia virus type I (HTLV-I) Tax proteins. However, the targetsequences fortransactivation by Bell are known to be as manyasfive distinct elements in the U3 domain of the HFV LTR (27). In addition, the bell gene product of HFV lacks
significant homology with the HIV-1 Tat and HTLV-I Tax proteins. Thus, the regulation mechanism of HFV Bell-medi-ated transactivation appears to be fundamentally different from that of other human retroviruses such as HIV-1 and
HTLV-I.In this regard, it islikely that the regulation mecha-nism of Bell-mediated transactivation is important to under-stand the lifecycle of HFV and the transcriptional regulation mechanism of eukaryotic cells.
Typical eukaryotic transcription activators have a modular design with separate DNA-binding and activating domains (11, 12, 28, 30,37). The DNA-binding domain directs the activating
*Correspondingauthor. Mailing address: Department of Life
Sci-ence,Pohang University of Science and Technology, Pohang 790-784, Korea.Phone:82-562-279-2294. Fax: 82-562-279-2199.
domaintoits target promoters, whereastheactivatingdomain interacts with some component of the transcription initiation complex. Transcriptional activation domains can be divided into three major classes on the basis of their amino acid sequences, including those that are glutamine rich (5, 8,51), proline rich(39),and acidic (19, 41).When Bell wasfusedto the yeastGal4DNAbinding motif, carboxy-terminalregionsof Bell fromresidues 255to266aswellasfrom 272to300acted asanactivation enhancement domain and autonomous activa-tion domain, respectively (53). Throughout this report, we have further defined thetranscriptional activation domain of Bell. Thecarboxy-terminal regionof Bell fromresidues 263 to 292 is sufficient for functioning as a minimum activation domain of 30 amino acids. Interestingly, this region does not contain predominantlyanyparticular type of the amino acids observed in the three major classes of transcriptional activa-tors, suggesting that the activation domain of Bell is an additional and distinct family of activation domainmotif.
Transcription factors regulating gene expression in the nu-cleus are synthesized in the cytoplasm and are therefore
subject tomechanismsdirecting the transcriptional regulatory
proteins into the nucleus. These transcription factors have a
specific nuclear localization signal (NLS) sequence required for active transport ofproteins to the nucleus. Typically, NLSs are eithershort sequences of 8 to 10amino acids containing a high proportion of positively charged residues (lysine and
arginine) orseveral sequences rich in basic amino acids which actsynergistically (7, 47).TheNLS of Bell was reported to be localized between residues 209 to 226 (17) and/or 211 to 225
(54) as asingle basic amino acid stretch.
Inaddition, many transcriptional activators appear to have a 2708
on November 9, 2019 by guest
http://jvi.asm.org/
multimerization domain required forthe biological activity of regulatory proteins and a negative regulatory domain which
regulates the activity of an activation domain (1-3, 21). The antioncogene product p53 is a transcriptional activator in
which theregion from residues92to 160negatively regulates thefunction of the strong activationdomain from residues 20 to92 (9, 33). The c-Rel protein which is closely related tothe Drosophila dorsal proteinand the mammalian NF-KB protein also contains a strong carboxy-terminal activation domain which isrepressedbyitsamino-terminal domain (4).
In this report,wehave furthercharacterized the functional
domain of the bell gene product to gain insights into the regulatorymechanism ofBell-mediated transactivation. First,
we constructed 24 missense mutants by oligonucleotide-di-rectedmutagenesis and tested fortheir abilities to transacti-vatetheHFV LTR,HIV-1LTR,and BREb in transientassays.
Next, we generated a chimeric Bell-VP16 and a series of
Gal4-Bell chimeric proteins that contained the amino-termi-nal 147 residues of a yeast transactivator, Gal4, to which a
portion of Bell wasfused. Thesechimeric constructs allowed
usto identify the minimumtranscriptional activation domain of30 amino acids and several regulatory regions which posi-tively or negatively modulate the activity of the activation
domain. Finally, we used in vivo competition assays to show thatthepositive regulatory domain of Bell actsintrans.These findings reveal a newmode ofregulation foratranscriptional
activation domain.
MATERIALS ANDMETHODS
Plasmid constructions. Constructions of thereporter CAT plasmidssuchas HFVLTR-, HIV-1LTR(NF-KB-)-, BREa-, BREb-, BREc3-, BREd-, and BREe-CAT were previously
described (25, 27). Other target promoter plasmids, G5ElbCAT and ElbCAT, contain the minimal promoter of the adenovirus Elb transcriptional unit with or without five
Gal4 DNA-bindingsites (26).
Missense mutations were introduced into the bell gene
present in pRC/CMV Bell by oligonucleotide-directed
mu-tagenesiswiththeAmershammutagenesiskit(24).pRC/CMV Bellwasgenerated by insertingthe bell openreadingframeof
HFV (nucleotides +9441 to +10711) into theHindIII-ApaI site of pRC/CMV (In Vitrogen). The specific amino acid changes introduced by oligonucleotide primers are listed in Table 1. All mutationswereconfirmedbya
dideoxy-sequenc-ing method (48). The carboxy-terminal deletion mutant of Bell,Bell(1-260),wasgenerated by cloningtheHindIII-ApaI
fragment of Bell-M259 into thesame site of thepRC/CMV
vector. Bell(1-278)wasgeneratedbyoligonucleotide-directed mutagenesis with a primer (5'-CTC`TL''TcIAAAGGGTC
CACCTGACATGTG-3') containing an early translational termination codon. Bell(1-260)IVP16, whichcontains the
ac-tivation domain ofherpes simplexvirustype1(HSV-1)VP16,
wasobtainedby insertingtheSacI-Apal fragment(aminoacids 423 to 490) ofVP16 into theApaI sites introduced in Bell-M259. Gal4-VP16 and Gal4(1-881) were kindly provided by
S. K.Jang(9).
Gal4 fusion plasmids were made by using pSG424, which
containsaDNAbindingdomain (aminoacids1 to 147)of the yeast transcriptional activator, Gal4,under the control of the simian virus 40(SV40)early promoterandaflanked polyclon-ingsite (26, 35).Theparental plasmid, Gal4-Bell(1-300),was
constructed as follows. The entire bell sequence from pHSRV (34)wasamplified by PCR, usingtwooligonucleotides
(5'-CrGGACATATGGATTCCTACGAAAAAGAAG-3' and
[image:2.612.318.560.82.373.2]5'-TCTAGACTGCGATTCAGCGTTGGATGTCAGAG-3')
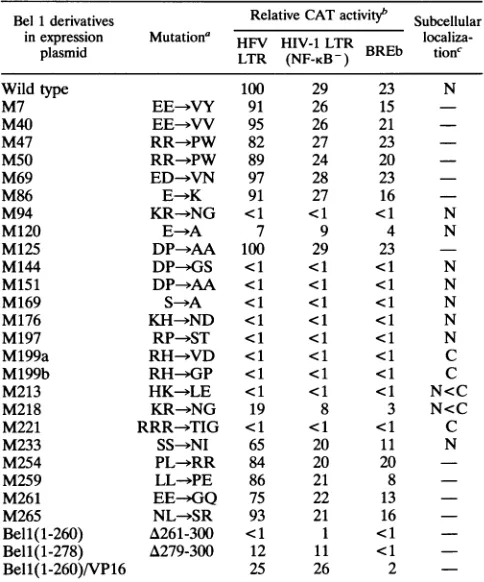
TABLE 1. Mutational analysis of the Bell protein Bel1derivatives Relative CATactivityb Subcellular
inexpression Mutationa HFV HIV-1 LTR
localiza-plasmid LTR (NF-KB- ) BREb tionc
Wild type 100 29 23 N
M7 EE-&VY 91 26 15
M40 EE-*VV 95 26 21
-M47 RR--PW 82 27 23
M50 RR--PW 89 24 20
M69 ED-->VN 97 28 23
M86 E->K 91 27 16
M94 KR--NG <1 <1 <1 N
M120 E- A 7 9 4 N
M125 DP--AA 100 29 23
M144 DP--GS <1 <1 <1 N
M151 DP->AA <1 <1 <1 N
M169 S-->A <1 <1 <1 N
M176 KH->ND <1 <1 <1 N
M197 RP->ST <1 <1 <1 N
M199a RH->VD <1 <1 <1 C
M199b RH--GP <1 <1 <1 C
M213 HK->LE <1 <1 <1 N<C
M218 KR--NG 19 8 3 N<C
M221 RRR->TIG <1 <1 <1 C
M233 SS-*NI 65 20 11 N
M254 PL-)RR 84 20 20
M259 LL->PE 86 21 8
M261 EE--GQ 75 22 13
M265 NL->SR 93 21 16
Bell(1-260) A261-300 <1 1 <1 Bell(1-278) A279-300 12 11 <1
Bell(1-260)/VP16 25 26 2
aSubstitutionsanddeletions ofamino acids introduced into Bell are
indi-cated.
bEach missensemutantofBell wascotransfectedwith reporter CAT con-structscontainingHFVLTR,HIVLTR(NF-KB ),orBREb intoBHK-21cells. The transactivationabilities ofvarious Bell missensemutants on BREa,BREc3, BREd, andBREe were notdetermined.CATactivitywasgivenrelativetothe transactivating ability ofHFV LTRbywild-typeBell, whichisarbitrarilyset at 100 from threeindependent experiments within lessthan15%variation.
cSubcellular localizations of the defectiveBellderivativesshown inFig.2 are indicated as N,C,andN<C,which denotenuclear,cytoplasmic,and predomi-nantlycytoplasmic distribution, respectively.-, mutants were notassayed.
as primers. Theamplified DNA fragmentwas fused in frame downstream of the Gal4 DNA-binding domain of pSG424
afterrestrictionenzymedigestionwithNdeIand Xba I.Various
Bell truncating mutants,Gal4-Bell(77-300), -(153-300), -(226-300),
-(237-300),
-(1-81/151-300),and-(1-226)weregenerated bydeletion ofEcoRI-PstI,
BamHI, ClaI, EcoRI-NcoI, BamHI-BamHI, and ClaI-XbaI fragments fromGal4-Bell(1-300), respectively.Gal4-Bell(1-260)wasconstructedby replacingtheClaI-HincII fragmentsof
Gal4-Bell(1-300)
with the ClaI-MscI fragments of Bell-M261.Gal4-Bell(1-82)
wasgenerated by deletion of the
BamHIl fragment
fromGal4-Bell(1-300) and thenbyreligation
(which
introducedanearly
translationaltermination
codon).
Gal4-Bell(82-150)
wascon-structed by
replacing
the BamHI-SacIfragments
ofpSG424
with theBamHI-PvuII fragment ofBell-M151. The
plasmids
Gal4-Bell(226-292),
-(226-278),
-(226-269),
and-(237-278)
wereconstructedintwosteps.First,DNAfragments
encoding
Bell amino acids 1to292,1 to278,and 1to269were
amplified
by PCR with a 5' primer with the sequence 5'-CTGGACATATiGGATTCCTACGAAAAAGAAG-3'
and 3'primers
5'-CTGATCTAGAGGGGGGACCCAAAATTGAG-3',
5'-CTC(5ThIXJA-iAAGGGTCCACCTGACATGTG-3',
and5'-GTACTCCCAAGTTTGGTAGGTTGCTGG-3',
on November 9, 2019 by guest
http://jvi.asm.org/
2710 LEE ET AL.
tively. Second, fragments ClaI-XbaI
(amino
acids 226 to 292 and 226 to278),
ClaI-SacI(amino
acids 226 to269),
and NcoI-XbaI (aminoacids 237 to278)
eachwere inserted into the SmaI-XbaI or SmaI-Sacl site of pSG424, respectively.Gal4-Bell(263-300)
was constructedby replacing
theEcoRI-AccI fragment of
Gal4-Bell(1-300)
with the MscI-AccI frag-ment of Bell-M261. When theincompatible
restriction en-zyme sites were used for cloning, blunt-endligation
wasperformed by treatment with mung bean nuclease or the Klenow fragment of DNA polymerase I. To construct
Gal4-Bell(263-292),
theDNAfragmentencoding
Bell amino acids263 to 292 was amplified by PCR,
using
twooligonucleo-tides
(5'-CTTf.GAATTCTCCAGCAACCTACCAAACT-3'
and5'-
CTGATCTAGAGGGGGGACCCAAAATTGAG-3')
asprimers.The
amplified
EcoRI-XbaIfragment
wasfusedtothe EcoRI-XbaI site ofpSG424.Gal4-Bell(1-260)/VPl6
was gen-eratedby replacingtheSacI-Apal fragmentofGal4-VP16 withthe
SacI-Apal
fragment ofBell(1-260)/VP16.The yeastexpression plasmids, yGal4,andyGal4wt
contain-ingthe entiregal4gene(aminoacids 1to881)andaselectable
marker,
Leu,were kindly provided by 0.K. Song(10).
yGal4contains the Gal4DNA-bindingdomain under the control of the yeast alcohol dehydrogenase promoter 1
(ADH1),
an autonomous replication origin, and a nutritional selectablemarker, TrpI. To generate yGal4-Bell(226-300), the
Clal-Hindlllfragment containing Bell amino acids 226to300was inserted in frame into thesamesite of theyGal4vector.
Cellculture,transfection,andCAT assay.
COS-7,
BHK-21,and HeLa cells were grown in Dulbecco's modified Eagle's
medium supplemented with 10% fetal calf serum. Plasmid transfections were carried out bythe DEAE-dextran method
(43).
Cells(106)
wereplatedona 100-mm-diameterdish 24 hbefore transfection and transfected with 2 ,ug of each of the reporter and activator plasmids. For the squelching
experi-ments, 1
pLg
of reporter, 0.5 ,ug of activator, and 5 ,ug ofcompetitor
plasmidswereused. At 48 h aftertransfection,thecellswere harvested and assayedfor chloramphenicol
acetyl-transferase (CAT) activity as described previously (15). The difference in transfection efficiencywas normalizedbyusinga second reporter
plasmid
containingtheluciferase
gene,pGL2-control
(Promega),
whichwascotransfected withactivator and reporterplasmids into cells. Luciferase activitywasmeasuredby
using the luciferase assay system (Promega) according tothe
suppliers'
recommendation. CAT enzyme reactionswerecarriedoutwith 50
jxg
ofprotein lysatesfor 2 h ofincubation. In cases in which the level ofacetylation ofchloramphenicolwas more than 90%, the protein lysates were diluted for the reaction and the CAT valueswere correctedby the dilution factor. AllCAT assay datareported in this articlewerefrom
points
in the linear range of the assay. Theproteinconcentra-tionwasdeterminedby usingaBio-Radproteinassaykit with bovineserumalbumin as astandard.
Preparationof anti-Bellrabbitantibody.ADNAfragment
of humanspumaretrovirusfrom nucleotides +9441to +10706,
corresponding to the entirebell open reading frame and the amino-terminal part of thebel2 gene,was amplified by PCR,
using
two oligonucleotides (5'-CTGGACATATGGATTCCTACGAAAAAGAAG-3' and 5'-TCTAGACTGCGATTCA
GCG1TTGGATGTCAGAG-3')
as primers, and then byre-striction endonuclease digestion with
NdeI
and XbaI. Theresulting DNAfragment was inserted into the
EcoRI-HindlIl
site ofpMAL-cRI,which isabacterial maltose-binding protein
(MBP)
hybridexpression vector (New England Biolabs), afterfilling
inwith Klenowfragment.The MBP-Bell fusion antigenwas overexpressed in Escherichia coli and purified by sugar
affinity
chromatography as described previously (36). Thepurified fusion protein (100 ,ug) was emulsified with Freund's complete adjuvant, and then the mixture was injected intra-dermally into a rabbit. The immunization was boosted four times at20-day intervals. The antiserumwasusedas aprimary antibody in the immunofluorescence experiments.
Immunofluorescence. Subcellular distribution of wild-type and mutant Bell proteins in COS-7 cells was analyzed by
indirect immunofluorescencemicroscopy. After 60 h of trans-fection, the transfected cells were fixed with 3.5% paraform-aldehyde, permeabilized with methanol, and then reacted with anti-Bell rabbit antibody (1/50 dilutions in Dulbecco's modi-fiedEagle's medium-1% fetal calf serum) and by reaction with goat anti-rabbit immunoglobulin G (heavy and light chains) coupledcovalentlytofluoresceinisothiocyanate (1/80 dilutions in Dulbecco's modified Eagle'smedium-1% fetal calf serum) (GIBCO-BRL).
Yeasttransformationand l-galactosidaseassay.Cellswere transformed, as described previously (10, 18), with 5 ,ug of yGal4, yGal4wt, and yGal4-Bell(226-300). Transformants wereselectedon supplementedsynthetic dextrose plates lack-ing tryptophan for yGal4 and yGal4-Bell(226-300)or onthose lacking leucine for yGal4wt. The resulting colonies were inoculated into syntheticmedium containinga nonrepressing sucrose as a carbon source and incubated until the culture reached mid-log phase(optical density at600 nm, 1.0). Then 0.1 mlof each culturewasaddedto0.7 mlof Zbuffer,and 50 ,ul of chloroform-50
[lI
of 0.1% sodium dodecyl sulfate was added and vortexed. The enzyme reactionwas carriedout at 30°C for 1 hafter the addition ofa ,B-galactosidasesubstrate,O-nitrophenyl-3-D-galactopyranoside. The optical density at 420 nm ofsamples was measured after removal of the cell
debris bycentrifugation.The 3-galactosidase activitywas rep-resented in unitsasdefined byJ. H. Miller (40).
RESULTS
Functional analysis of Bell protein. To determine the critical amino acids in the function of the Bell protein, we introduced 24 distinct missense mutations by site-directed mutagenesis at various positions within the bell gene. Each mutant was namedaccordingto theposition of the first Bell amino acid changed by the mutation (Fig. 1). The
transacti-vating abilityof thevarious Bell missense mutants was deter-minedbycotransfection with CATconstructscontainingHFV LTR, HIV-1 LTR(NF-KB-), and BREb into BHK-21 cells. The target sequence for transactivation of Bell was previously mappedtofive distinct regions in the U3 domain of the HFV LTR, BREa, BREb, BREc, BREd, and BREe (27). Since BREb showed the highest activity for Bell-mediated
transac-tivation,itwasusedas asingle target site within the HFV LTR for thefollowingstudies. The amino terminus of the bell gene
product has two regions from residues 5 to 44 and from residues 67 to87with ahigh content of acidic amino acids, a property observed in the "acid blob" of the transactivation domainexamplified by VP16 (6, 50). This sequence property suggeststhat theseregionsconstitute the activation domains of Bell. However, missense mutations introduced into these
regions (M40 and M69) had no significant effect on Bell-mediatedtransactivation, whereas M7 and M86 showed small reductions in CAT activity (approximately 33%) on the BREb promoter(Table1). These results are in good agreement with
previous reports that the amino-terminal portion of Bell is a nonessential region or a shared domain dispensable for Bell transactivation(17,54).Inaddition, missense mutations intro-ducedintoastretchof basic aminoacids (M47 and M50) had little effect on Bell-mediated transactivation. In contrast, J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
M7
M D S Y E KH S V A S
M40
A G E G E L T I AE
P
M69
S Y H A Y K E
IE
K HM94
S L C L I L C G L Y
M125
Q S D Tu D C F I V S
M151
E V G I W V K Y K P
T S G I Q D L Q T
M47 M50
E E N P
[P
R
P Q H I K L Q D W S A E K A S E I L Y T C I F C D A V
Ml 69 L R G I V G A9V,
M197 99a,b
30
L S E L V G P E N Y T K R E V
M86 I P T PW E
K C V 90
M S K
M120
r- ,120
R M P F T V S
WW
M1"
150
I H D P M P I R W M176
130
F I M H
KHQ
R N210
C S L V K P S T S C S E G P K P|
R
P||R H| D P V L R C D M F EM213 M218 M221 M233
K
HH
P R QPi
R R F SID
N E S C A SjjD
T M A N EM254 M259 M261 M265
P G S L C T N P L W M P G L S
GjujHJ
S
SN
P N L E240
270
300
V H M S G G P F W E E V Y G D S I L G P P S G S G E H S V L
FIG. 1. Summary of mutations introduced into the HFV bell gene. The HFV bell geneencodes a protein with the predicted 300-amino-acid sequence shown. The clustered point mutations (M) introduced byoligonucleotide-directed mutagenesis are indicated by the boxed residues. These mutations were named according to the changed first amino acid of their location within Bell.
various missense mutations within Bell central region (M94, M144, M151, M169, M176, M197, M199a, M199b, M213, and M221)completely abrogated Bell function on all of the CAT constructs containing the HFV LTR, HIV-1 LTR(NF-KB ), and BREb (Table 1). Other Bell missense mutants such as M120 and M218 showed reduced transactivating ability, whereas M125 only retained the wild-type activity of Bell. Missense mutations introduced at the carboxy terminus, such as M233,M254, M259, M261, and M265, had a little negative effect on CAT activities. Especially, the M233 and M259 missense mutants showed more than 50% reduced CAT activityonthe BREbpromoter.However, mostof themissense Bell mutants exhibited a similar pattern of transactivation abilityonall three reporter constructs tested. Takentogether, these results suggest that the central region ofBell plays a criticalrole forBell-mediated transactivation.
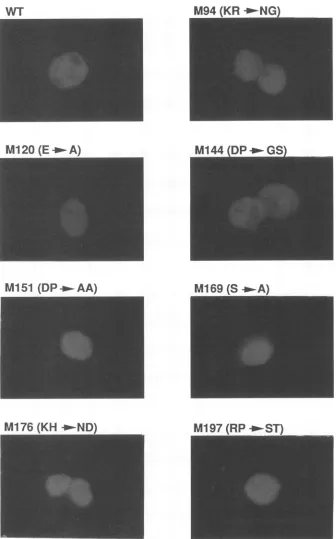
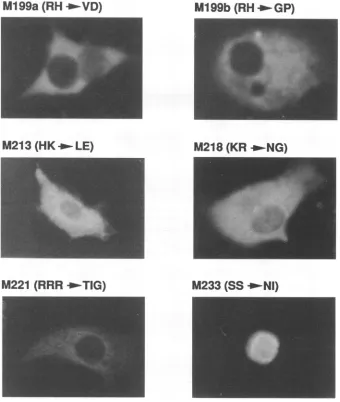
Inevaluating the functional activity of variousmutantforms ofatransactivatorprotein, it is obviously importanttoconfirm that theexpected protein displaysanormal intracellular local-ization pattern. To investigate the subcellular localization of the nonfunctional Bell missense mutants, immunofluores-cence staining of transiently transfected COS-7 cells was carried out. Immunofluorescence staining of cells expressing wild-type or mutant Bell is shown in Fig. 2, and the data obtained are summarized in Table 1. Some of the nonfunc-tional Bell mutants displayed asubcellular localization
indis-tinguishablewith that of the wildtype, indicatingthat lack of transactivation by Bell mutants such as M94, M144, M151, M169, M176, and M197 is not due to the lack of nuclear localization. However, M199a, M199b, and M221 were con-centrated in thecytoplasmof transfected COS-7cells, indicat-ing that the lack of transactivation by these Bell mutants is causedbyabnormal intracellular localization.Inaddition,both M213 and M218 were distributed diffusely throughout the expressed cells, but localized preferentially in the cytoplasm
around the nucleus. These results indicate thattwoclusters of basic amino acids within Bell, from residues 199 to 200 and 211 to 223, are essential for the nuclear localization of Bell,
suggestingthat the nucleartargetingsequenceof Bell protein is composed of a bipartite basic amino acid motif. These observations are not consistent with previous reports which showed that the single basic amino acid cluster of Bell from
residues209 to 226 and/or 211 to 225 was both necessary and sufficient for the nuclear localization ofBell protein (17, 54). In contrast to the results obtained by missense mutations introduced at the carboxy-terminal region, deletion of the carboxy-terminal region of Bell from residues 261 to 300 completely abolished transcriptional activationby Bell (Table 1).Deletion of22amino acidsatthe carboxyterminus partially retained transactivation ability by 12, 40, and <1% ofthatof wild-type Bell on HFV LTR, HIV-1 LTR(NF-KB
),
and BREb, respectively (Table 1). To determine whether this carboxy-terminal region of Bell functions as an activation domain for Bell-mediated transactivation, we replaced the carboxy-terminal region of Bell from residues261 to300with thetranscriptional activation domainfrom HSV-1 VP16 (ami-no acids423 to490). Fusion ofthe VP16 activatingregion to the carboxy terminus of Bell(I-260) to create Bell(I-260)/ VP16partiallyrestoresthe ability ofwild-type Belltoactivate its own target HFV promoter, suggesting that the region of Bell from residues I to 260 functions as a promoter-binding domain that directs Bell to its target HFV promoter. In particular, these chimeric proteins displayed a full activity whentestedon HIV-1 LTR(NF-KB-) (Table 1). Theseresults indicate that the VP16 activating domain can activate tran-scription from thetargetsequenceof Bell throughthe context ofthe Bell binding motif.Identification ofdominant-negative mutants. Several mis-sense mutants, which completely lacked any transactivation capability, were tested for the ability to interfere with the transactivating activity of the wild-type Bell protein in trans. Bothwild-type Bell anda10-fold molarexcessof thedefective Bellexpressionplasmidswerecotransfected intoBHK-2 I cells with CAT indicatorplasmidscontaining theHFVLTR, HIV-1
LTR(NF-KB-), BREb, or Rous sarcoma virus
(RSV)
pro-moter as a control. Mutants such as M94, M144, M151, and M176 reduced the CAT activity directed by wild-type Bell. MutantM120 hadno significant inhibitory effect onwild-type
Bell-mediated transactivation, whereas two mutants (M169 andM197)appeartohaveatleastapartialdominant-negative
effect on wild-type Bell (Table 2). The inhibitory effect was most dramatic inmutants M144 and M151,
suppressing
wild-type Bell-mediated CAT activity by 70 to 90%(Table
2).
However, these nonfunctional Bell mutantshad no effect onon November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.147.476.77.256.2]2712 LEE ET AL.
WT
M1
51
(DP
_
AA)
[image:5.612.135.471.70.609.2]M197
(RP -_-ST)
FIG. 2. Subcellular localizationof wild-type anddefective Bell mutantsby indirect immunofluorescence. Immunofluorescence photographs
wereobtainedbytransienttransfectionwith theplasmid expressingthe wild type or mutants of Bell into COS-7 cells.Cellswerestainedat60 h posttransfection with rabbitpolyclonal antiserumspecifictoBell and thentreated withfluorescein-conjugated goatanti-rabbitimmunoglobulin Gasdescribed in Materials and Methods.
the gene expression directed by the RSV promoter. These resultsdemonstrated that the interference observed here was a
specific effect by defective Bell mutants andwas not simply dueto apromotercompetitionphenomenon.
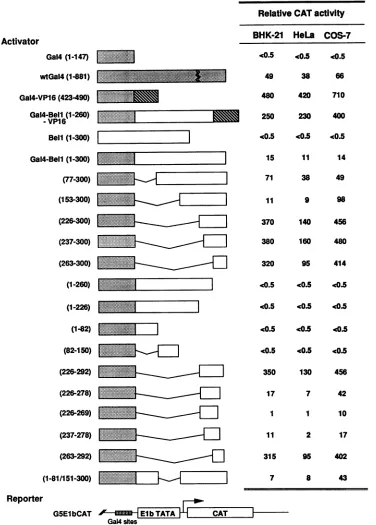
HFVBell contains a distinct activation domain as well as
one positive and two negative regulatory domains for the functionofthe Bellactivationdomain. Todetermine whether Bell contains a transcriptional activation domain, we con-structed several Gal4-Bell chimeric
proteins
thatcontain the amino-terminal147residues of Gal4towhichaportion
of Bell J. VIROL.on November 9, 2019 by guest
http://jvi.asm.org/
M199a (RH
_VD)
M1199b (RH _'GP)
M213
(HK_-LE)
M218 (KR
_NG)
M221 (RRR
_TIG)
M233 (SS
_'NI)
FIG. 2-Continued.
was fused. These fusion constructs were generated in the
context of theeukaryotic expression plasmid pSG424 (26). As
a control,we used a Gal4(1-147) containing the Gal4 DNA-binding domain, awild-type Gal4, and aGal4-VP16 chimeric
constructcontainingthe acidicactivation domain of the HSV-1 VP16 transactivator (residues 423 to 490) fused to the Gal4 DNA-binding domain. The transactivating ability of the chi-meric Gal4-Bell proteins was determined by measuring the
CAT activity following cotransfection with a G5ElbCAT
re-porter plasmid into BHK-21, HeLa, and COS-7 cells. The G5ElbCAT plasmidcontains anoligonucleotide consistingof both Gal4 DNA-bindingsites and the adenovirusElb TATA box inserted immediatelyupstreamof thecatgene. Results of representative CATassaysareshown inFig.3. Asexpected, a
wild-type Gal4andGal4-VP16 showed respectiveCAT activ-itiesatleast 76- to 132-fold and 840-to1,420-fold higherthan that of Gal4(1-147), depending on the cell line used. Gal4-Bell(1-300),inwhich Bell is boundartificiallytothe
[image:6.612.130.470.73.475.2]heterol-ogous promoterthroughtheDNA-binding regionof theyeast activatorGal4, exhibited an enhancing activityat least 22- to
TABLE 2. trans-dominance of severalnonfunctional Bellmutants
Indicator(CATactivity)"
Competitoi' RSV HFV LTR BREb (NF B-)
CMVC 80 90 21 23
M94 78 23 4 6
M120 84 90 25 15
M144 72 8 7 3
M151 80 8 2 3
M169 91 85 21 9
M176 76 22 7 3
M197 85 80 16 14
"Bel lexpression plasmid (0.5pLg)wascotransfectedwith 10-fold molarexcess
ofdefective Bell derivatives(M94,M120, M144, M151,M169,M176,andM197)
as acompetitorand 1 ,ugofareporterCATconstructcontainingRSVLTR,
HFVLTR,BREb,orHIV-1LTR(NF-KB-)into BHK-21 cells.
"CATactivitywasderived from three independent experimentswithin the
errorrange of about 10% standard deviation. 'CMV,cytomegalovirus.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.320.561.553.666.2]J. VIROL. 2714 LEE ET AL.
Activator
Gal
(1-147)
wtGal4(1-881)
---Gal4-VP16(423-490)
RelativeCATactivity BHK-21 HeLa COS-7
-0.5
40.5
40.5
49 38 66
480 420 710
Gal4-Bell(1-260)
-VP16
Bell (1-300) Gal4-Bell(1-300)
LIZZIIIZI
I
250 230 400
40.5
40.5
40.5
15 11 14
(77-300)
LF=
(153-300) (226-300) (237-300) (263-300)
Ii l
(82-1 50) (226-292)
(226-278)
(226-269) (237-278) (263-292) (1-81/151-300)
71 38 49
11 9 98
370 140 456
380 160 480
320 95 414
40.5
40.5
40.5
40.5
40.5
0c.540.5
40.5
40.5
40.5
40.5
40.5
350 130 456
17 7 42
1 1 10
11 2 17
315 95 402
7 8 43
Reporter _
G5E1bCAT = E CAT
[image:7.612.123.499.100.625.2]Gal4sites
FIG. 3. Transactivationpotential of plasmids encoding thewildtypeandGal4-Bellproteinderivatives. Theabilityof each fusionprotein,Gal4 DNA-binding domain (stippled box), andapartofBell (open box) totransactivatethe reporter constructG5E1bCAT in transientassayswith BHK-21, HeLa, and COS-7 cells is indicatedonthe right by CAT activity. The transcriptional abilities ofGal4-Bellchimeraswerecomparedwith those ofawild-typeGal4andGal4-VP16(positive controls) andGal4(1-147) (negative control).The CATactivityvaluesareaveragesofatleast fourindependent experiments, witha relative standard deviation less than14%. Inaddition,weusedaluciferasegeneas asecond reporterto
detect thedifference intransfectionefficiency,asdescribedin Materials andMethods. When thepercentacetylationofchloramphenicolwasmore
than90%, CATassayswererepeatedwith 1/5-to1/10-diluted cell lysatestoobtain data within the linearrangeoftheexperimentandthe CAT valuesobtainedweremultiplied byan appropriate dilution factor.
(1-260)
(1-226)
on November 9, 2019 by guest
http://jvi.asm.org/
HFVLTR CAT
A
B
9
HFVLTRCAT + Bell
-e
_
I
;
X
3
;_
j
.-
t
v-@0
I@
@
*-.
.,e.@
@
41 <1 <1 1.5 <1 <1 <1
CATactivity <1 89 90 88 64 5 62 38 7
[image:8.612.67.557.70.271.2](%conversion)
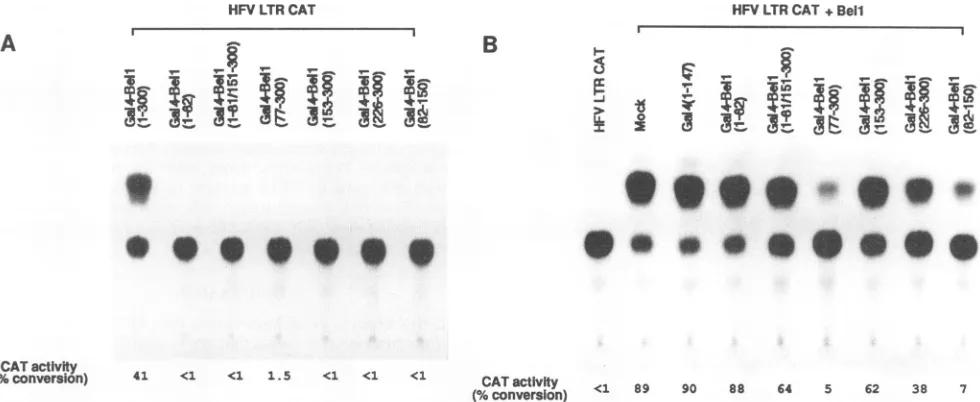
FIG. 4. Effect of Gal4-Bell fusion proteins on HFV LTR-directed gene expression in the absence or presence of Bell. (A) One microgram of variousGal4-Bell hybridplasmids wascotransfected with1 ,ug of HFV LTR CAT construct into BHK-21 cells. (B) As an activator, 0.5 ,ug ofBell expressionplasmid (45)wascotransfected with 10-fold excess amounts ofGal4-Bellhybrid plasmids (ascompetitors) and1 ,ug of HFV LTR CAT. All of theactivator and competitor plasmids were expressed under the control of the SV40 early promoter. Relative CAT activity was measured aspercent acetylation as indicated at the bottom.
30-fold higher than that of Gal4(1-147), indicating that Bell may function naturally in the vicinity of the promoter and contain a transcriptional activating region. Deletions ofBell from residues 1 to76and from 1 to225resultedin anincrease of CAT activity by approximately 3- to 5-fold and 13- to 32-fold, respectively, suggestingthat the regions from residues 1 to76and153 to225have aninhibitoryeffect on the function of the Bell activation domain. In contrast, deletion of the amino-terminal 76amino acids from Gal4-Bell(77-300) obvi-ously reduces the abilitytoactivate CATexpression by four-to sixfold in BHK-21 and HeLa cell lines, indicating that the region from residues77 to 152 has apositive regulatory effect on the function of the activation domain. Also, Gal4-Bell(1-81/151-300) showed similar, but alittle lower, levels of trans-activation activity when compared with those of
Gal4-Bell(153-300). However, thesameconstructs, Gal4-Bell( 153-300) and Gal4-Bell(1-81/151-300), did not show any decrease in CAT activity compared with Gal4-Bell(77-300) in COS-7 cells, suggesting that the positive effect of the region from residues 77 to 152wasnotapparent. Instead, deletion of Bell residues 77 to 152 showed an increase in CAT activity of twofold. It seemslikelythatthe different effects dependingon cell lines are caused either by replication of the activator plasmid containing the SV40 origin in COS-7 cells or by the interference of the large T antigen of SV40 constitutively
expressed in COS-7 cells. In fact, the CAT activities of Gal4-Bell fusion constructs ingeneral were higher in COS-7 cells than those in BHK-21 and HeLa cells.
Further analyses of amino-terminal deletion mutants showed that thecarboxy-terminalregionof Bell from residues 263 to 300 is enough to activate cat gene expression. As expected, carboxy-terminal deletion mutants such as
Gal4-Bell(1-260), Gal4-Bell(1-226), and Gal4-Bell(1-82) com-pletely eliminated transactivating ability. Aseries of
carboxy-terminal deletion mutants from Gal4-Bell(226-300) revealed that the region of Bell from residues 293 to 300 is not
necessaryforactivity, buttheregionfrom residues 270to292 isimportant forthetransactivation ability of Bell. It is likely
that theregion of Bell from residues263 to 292 issufficientfor functioningas aminimum transactivation domain. Ourresults are partially consistent with thoseofprevious studies inwhich thecarboxy-terminal portions (residues247to296 in He et al. [17] and residues 255 to 266aswell asresidues 272 to 300 in Venkateshetal. [53]) arecrucial for transcriptional activation of Bell. The activity ofGal4-Bell(263-292) is approximately twofold lowerthanthat of Gal4-VP16.Theactivating region of VP16 isstrongest among the reported activating regions. The activating region ofBell in the Gal4 fusion issixtimesstronger than that ofwild-type Gal4.
Todetermine whether the aboveregulatory regions play an importantroleinHFVLTR-directedgene expressionbyBell in trans, we cotransfected the Bell expression plasmid with 10-fold excess amounts of various Gal4-Bell derivatives into BHK-21 cells. The Gal4-Bell derivatives did not show any significant effectsonthebasal geneexpression directed bythe HFV LTR (Fig. 4A). However, Gal4-Bell derivatives Gal4-Bell(77-300) and Gal4-Bell(82-150) severely inhibited Bell-mediatedtransactivation of the HFV LTRbyatleast 13-fold, whereas derivatives Gal4-Bell(1-82), Gal4-Bell(1-81/15 1-300), and Gal4-Bell(153-300) did not have any significant effect on Bell-mediated gene expression (Fig. 4B). However,
Gal4-Bell(226-300), which contains the strong activation do-main ofBell, weakly inhibitedtranscriptional activation by the Bell protein. These results indicate that the region of Bell fromresidues82to150containsaregulatory domain actingin trans. To rule out the possibility that the regulatory region
binds totarget DNA sequences of Bell directly or
indirectly,
therebyinhibitingthe function ofBell,wetested the effect of nonfunctional Bell protein lackingthe transactivation domain on transcriptional activation directed by G5E1bCAT as a heterologous promoter by Gal4-Bell fusion derivatives. For this purpose,we cotransfected the defectiveBell(1-260)
with variousGal4-Bell derivatives intoBHK-21 cells.ThedefectiveBell(1-260) inhibited transactivation directed
by
Gal4-Bell(77-300) but not that by
Gal4-Bell(153-300)
andGal4-Bell(226-300) with the
G5E1bCAT
promoter(Fig.
5).
ToCATactivity (% conversion)
qp- v- 9-9- v- v- v-
9p-r.
9
3,
AV
v-9-
A
i
...w %.O 0
s
6
s
--.Oon November 9, 2019 by guest
http://jvi.asm.org/
2716 LEE ET AL.
GSE1bCAT upon nonrepressing carbon source induction, whereas Gai4-kl(77-300) - + + - - - yGal4wt, containing the entireGal4 domain (amino acids 1 to
Ga4-BOI7(153-30)
- - - + + -881),
showedaP-galactosidase
activity
2,300-fold
higher
thanGaI4-Bell(225-300)
- - - + + that ofyGal4. The chimeric Gal4-Bell(226-300) showed anSBl1(1-260)
+ - + - + - + increased activity of about 230-fold (Fig. 6). We thereforeconcludethat the activation domain of Bell, albeit at a lower activity, also functions in yeast cells, suggesting that the 9 9le9; 9 *_ function of the
activating
domainwithin Bell is well conserved acrossspecies
boundaries. Sinceonly
the acidic activation domain observed in VP16 appears to function efficiently in yeast cellsaswellasmammaliancells,it will beveryinteresting* to determine how the nonacidic activation domain of Bell functions inyeastcells.
+~~~~~~
CATactivity
(% conversion) <1 56 15 8 9 165 174 FIG. 5. Effect ofdefective Bell(1-260) on the transactivation of G5E1bCATby variousGal4-Bell derivatives.Asanactivator,0.5 jig
ofGal4-Bell derivativesand 10-fold excess amounts ofBell(1-260) (as competitor)werecotransfectedwith 1 ,ug ofG5E1bCATinto BHK-21 cells. Relative CATactivity wasmeasured aspercent acetylation as
indicated atthe bottom. Lysates of transfectantwith Gal4-Bell(226-300)weredilutedtoobtain CAT values within the linear rangeof the assay, andthe CATvalues obtained weremultiplied byanappropriate dilution factor.
examine whether these positive and negative regulatory do-mains of Bell also function with the heterologous activation domain, we constructed Gal4-Bell(1-260)/VP16 by inserting
theregion of Bell from residues1 to260into Gal4-VP16.The resulting construct showedatransactivation ability comparable tothat of Gal4-VP16 (Fig.3), suggestingthattheseregulatory
regions arespecific to the activation domain of Bell.
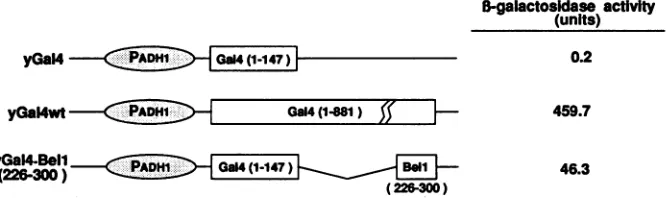
Transcriptional activating domainofBell functions in yeast cells.The fact thatseveral acidic transactivators suchasHSV-1 VP16and Gal4efficientlyfunction in both yeast and mamma-lian cells indicates that the pathway oftranscriptional activa-tion may be mediatedby a commoncellularfactor(s) ubiqui-tously existing in eukaryotic cells (9, 13, 42, 44). To test whether the transcriptional activation domain of Bell also functions inSaccharomyces cerevisiae, we constructed
yGal4-Bell(226-300) in whichGal4-Bell(226-300) was placed under the control of the yeast alcohol dehydrogenase gene promoter. As a control, yGal4, which contains only the Gal4 DNA-binding domain, did not induce any
3-galactosidase
activityDISCUSSION
In this report, we demonstrated that HFV Bell contains multiplefunctional domains thatareessentialfor itsbiological
activity in vivoand whose functionsarepossiblyregulated(Fig.
7). Extensive mutationalanalysesidentifiedthedistinctregions
of Bell requiredforHFV LTRtransactivation. Any missense mutation within the central region of Bell from amino acids 144 to213resulted in the total loss of functional Bellactivity,
indicating that this region plays a critical role for Bell-mediated transactivation. It was recently suggested that the loss oftransactivatingability bythe missense mutationwithin the Bell centralregionwasduetothe defect in thefunctional
binding domain(17).The regionof Bell from residues 263 to 292is capable offunctioning as astrongminimum activation domain in the Gal4-Bell fusion construct. The activating
region contains ahighly conserved sequence (residues 273 to 287) among the three transactivators of foamy viruses se-quenced so far (17, 38) (Fig. 7). In addition, the activating
domain of Bellcanbefunctionallyreplaced,atleast in part,by
the acidic activation domain of the HSV-1 VP16 transactivator at the carboxy terminus, indicating that the wild-type Bell contains a promoter-binding domain which directly or indi-rectly makes a contact with its own target DNA sequence elements. These results are partiallyconsistent with previous
reports that the carboxy-terminal region of Bell contains an activation domain that can be replaced by the activation
domain of VP16(17).Wepreviouslydomonstrated that trans-activation by Bell is mediated by at least five different
cis-acting elements present in theHFV LTRandbyonespecific
element in the HIV-1LTR(25, 27). Therefore,wesuggestthat Bell indirectly interacts with its target sites via DNA-bound proteins and containsadiscrete potenttransactivation domain. HFV Bell may bind to some proteins avidly and to other cellular proteins with lower affinity,which would account for
13.galactosidase activity (units)
0.2
yGa4
yGal4wt
F-yGal4Beil1 (226-300)
(226 300)
459.7
46.3
FIG. 6. Transcriptional activating domain of Bell functions in yeast cells. The region of Bell from residues 226 to 300 was fused in frame to theGal4DNA-binding domain within a yeast expression vector, as described in Materials and Methods. StrainCTY1wastransformed with yGal4, yGal4wt,oryGal4-Bell(226-300) hybrid construct. The
P-galactosidase
activity in each transformant was obtained from two different experiments, with lessthan15%variation.J. VIROL.
GaM4(I-88 )
Mf
on November 9, 2019 by guest
http://jvi.asm.org/
[image:9.612.60.298.72.260.2] [image:9.612.142.475.581.680.2]HUMAN FOAMY VIRUS Bell PROTEIN 2717
[image:10.612.138.488.77.165.2]Regulatory / Promoter bindingregion NLS
F... |
::::::
...::.::::.:::::::.:::.::::::::::.::-.::::..,.:::::::.::.:::::::::::::::::::::...::::::::::::::::: ::::::::_
~~~~~~~~~~~~~~~~~._
_~~~~~~~~~~~~~~~~~~... .. ....soe
94 144 151 176 trans-dominant negative region
TAD
0 Qc-m
nr
conserved
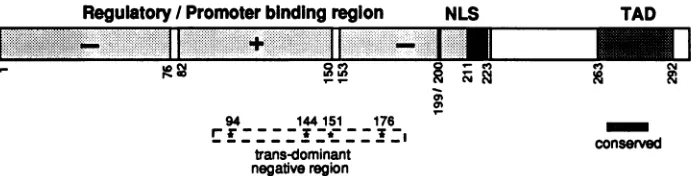
FIG. 7. Diagrammatic representation of the functional region of the Bell transactivator. The regions of Bell from residues 1 to 76 and 153 to 225 contain two negative regulatory domains (-); the central region of Bell from residues 82 to 150 contains a positive regulatory domain (+). The promoter-binding region from residues 1 to 260 appears to be overlapped with these regulatory domains. NLS and TAD denote a bipartite NLS sequence (amino acids 199 to 200 and 211 to 223) and a discrete transcriptional activation domain (amino acids 263 to 292), respectively. The asterisks represent transdominant-negative regions (M94, M144, M151, andM176), and the small hatched box designates a highlyconserved stretchof amino acidsequencesbetween HFV Bell and simian foamy virus Taf.
the variable effects of Bell on different Bell-responsive ele-ments (Table 1).
Surprisingly, the full-length Bell protein in the Gal4-Bell fusionwas10- to30-fold less active than the carboxy-terminal region. Maximum transcriptional activity of the Bell protein in theGal4-Bell fusion was revealed only if the carboxy-terminal portion of Bell was retained (Fig. 3).Aunique feature of Bell is that in the full-length protein containing one positive and two negative regulatory domains, the negative regulatory do-mains are dominant, as judged by lower transcriptional activa-tion. When the amino terminus of Bell containing one nega-tive and one positive domains was deleted, the effect of the
positive regulatory domain was revealed. Since these regula-tory domains did not down-regulate the activity of the VP16 activation domain(Fig.3),these domains specifically modulate the activation domain of Bell. The competition experiments showed that the positive regulatory domain acts in trans, suggesting that thisregion is involved either in the oligomer-ization of Bell monomers or in the interaction with cellular factor(s) required for Bell-mediated transactivation. Itislikely that the function of Bellproteinis modulated by the positive regulatory domain, thereby activating transcription by exposing thecarboxy-terminal activation domain masked by the nega-tive regulatory domain. These results indicate that Bell-mediated transactivation involvesa complex regulatory path-waywhich provides a novel mode of action for transcription regulators.
Nuclear targeting sequences are essential for the transport of transcription factors into the nucleus. The targeting se-quences are usually short stretches with many positively chargedamino acids (7, 49). Previous reports suggest that the basic amino acid sequences located between positions 209 to 226 and/or 211 to 225 are necessary and sufficient for the nuclear localization of Bell (17, 54). However, our results showed that Bell requirestwobasic amino acid clusters from residues 199 to200 and 211 to223 inorder totransportBell into the nucleus, suggesting that the NLS sequence of Bell functionsas abipartite
motif,
which is similartothatof other nuclearproteins (7,
47,49).
Thediscrepancy
between our resultsandprevious reports maybe due toeither the difference in the position of amino acids changed by site-directedmu-tagenesisortheartificial
generation
ofashortbasic amino acid cluster upstream of Bellfragments
in theBell/p-galactosidase
fusion plasmid constructedby
previous
studies(17, 54).
We haveconfirmed that both clusters ofpositively charged amino acids are requiredfordirecting
Bell to the nucleusby
fusing
theseclusterstothe
3-galactosidase
andinserting 12-bp
linkers betweentwobasicamino acid clusters(data
notshown).
Takentogether,weconclude thatHFVBell appearstobea
multido-mainprotein, containing multiple control regions, a
promoter-binding region, a bipartite NLS sequence, and a distinct
transcriptional activation domain (Fig. 7).
A detailed mutational analysis of several transcriptional activation domains has indicated that they can be classified into several classes, including the acidic motifs found in VP16,
Gal4, and Rta (6, 16, 31, 32, 35,50), the proline-rich domain observed in CTF/NF-1 (39), and the glutamine-rich motif found in Spl (5). The negatively charged residues in acidic activators are shown to contain an amphipathic helix in which theacidic amino acids are positioned on one side of the helix, presumably to allow them to come into contact with another protein (14,41). The minimum activation domain of Bell from residues 263 to 292 does not contain predominantly any particular type of amino acid observed in the above transacti-vators (13% acidic residues, 13% proline residues, and no glutamine residues). In contrast, it contains a high percentage of hydrophobic amino acids (33%). It was recently reported that the mutations ofspecific hydrophobic residues within the activation domain of HSV-1 VP16 and Epstein-Barr virus Rta severely impaired the transactivation ability (6, 16, 20, 44),
suggesting that either specific hydrophobic interaction with
protein partners or intramolecular folding is important for transactivation. In addition, a computer analysis (Microgenie
sequence analysis program) predicted that the minimum acti-vation domain of Bell predominantly forms a ,B-sheet or a
(3-turn
in the secondary structure of the protein (data notshown). It wasrecently demonstrated that ,B-sheetstructurein the activation domain is essential for the properfunction of Gal4 rather than theamphipathica-helicalstructure (29,52). Itremainstobe determined whether the activationdomain of Bellfunctions viaanovelmechanism in which both
hydropho-bic residues and,8-sheetstructure are involved in thefunction of the Bell activationdomain.Tofurtherassessthe contribu-tion ofspecificresidues indirecting transcriptional
activation,
adetailed mutationalanalysiswithin theactivation domain will be necessary.ACKNOWLEDGMENTS
We thank Y. D. Yun (Mogam Institute of Biotechnology) for providing anti-Gal4-polyclonal antibody and for plasmids pSG424, Gal4, G5ElbCAT, and EIbCAT.
This work was supported by Research Institute of Science and Technology grant R93035 and Center for Biofunctional Molecules grant CBM-93-07.
REFERENCES
1. Baichwal,V.R., A.Park,andR.Tjian. 1992.The celltype-specific activator region of c-Jun juxtaposes constitutive and negatively regulateddomains. Genes Dev.6:1493-1502.
VOL.68,1994
on November 9, 2019 by guest
http://jvi.asm.org/
2718 LEE ET AL.
2. Bogerd, H. P., R. A. Fridell, W. S. Blair, andB. R.Cullen. 1993. Genetic evidence that the Tat proteins of human immunodefi-ciencyvirus types 1 and 2can multimerize in the eukaryotic cell
nucleus. J. Virol. 67:5030-5034.
3. Bogerd, H. P., and W. C. Greene. 1993. Dominant-negative
mutants of human T-cell leukemia virustype 1 Rex and human immunodeficiencyvirustype1 Revfail tomultimerize in vivo. J.
Virol. 67:2496-2502.
4. Bull, P., K. L. Morley, M. F. Hoekstra, T. Hunter, andI. M.
Verma. 1990. Themousec-Relprotein hasanN-terminal
regula-tory domain and a C-terminal transcriptional transactivation
do-main. Mol. Cell. Biol. 10:5473-5485.
5. Courey,A. J., and R.Tjian. 1988. Analysis ofSpl in vivoreveals multipletranscriptionaldomains,includinganovelglutamine-rich
activation motif. Cell 55:887-898.
6. Cress, W. D., and S. J. Triezenberg. 1991. Critical structural elementsof the VP16 transcriptional activation domain. Science 251:87-90.
7. Dingwall, C., and R. A. Laskey. 1991. Nucleartargeting sequenc-es-aconsensus?Trends Biochem. Sci. 16:478-481.
8. Driever, W., J. Ma, C.Nusslein-Volhard, andM.Ptashne. 1989. Rescue of bicoid mutant Drosophila embryos by bicoid fusion proteins containing heterologous activating sequences. Nature
(London)342:149-154.
9. Fields, S.,andS. K. Jang.1991. Presenceofapotenttranscription activatingsequencein the p53 protein. Science249:1046-1049.
10. Fields, S., and0.K.Song. 1989. Anovelgeneticsystemtodetect
protein-proteininteractions. Nature (London) 340:245-246. 11. Flemington,E. K., A. M. Borras,J. P. Lytle,and S. H.Speck. 1992.
Characterizationof the Epstein-Barrvirus BZLF1 protein
trans-activationdomain. J. Virol.66:922-929.
12. Frankel,A. D., and P.S. Kim. 1991. Modular structure of
tran-scriptionfactors:implications forgeneregulation. Cell 65:717-719.
13. Gill, G.,I. Sadowski, and M. Ptashne. 1990. Mutations that
increase the activity of a transcriptional activator in yeast and mammaliancells. Proc.Natl. Acad. Sci. USA87:2127-2131. 14. Giniger,E., and M. Ptashne.1987.Transcription inyeastactivated
byaputative amphipathicao-helixlinkedtoaDNAbinding unit.
Nature (London)330:670-672.
15. Gorman,C. M., L. F.Moffat, and B. H. Howard. 1982.
Recombi-nantgenomeswhichexpresschloramphenicol acetyltransferase in
mammaliancells. Mol.Cell.Biol. 2:1044-1051.
16. Hardwick,J. M., L. Tse, N. Applegren, J. Nicholas, and M. A.
Veliuona. 1992. The Epstein-Barr virus R transactivator (Rta) contains a complex, potent activation domain with properties differentfrom those ofVP16. J.Virol.66:5500-5508.
17. He, F.,J. D. Sun,E. D. Garrett, and B. R.Cullen.1993.Functional
organizationofthe Bell transactivator of human foamy virus. J.
Virol. 67:1896-1904.
18. Hill, J.,K. A. Donald, and D. E.Grifliths. 1991. DMSO enhanced
wholecellyeasttransformation. Nucleic Acids Res. 19:5791.
19. Hope, I. A., and K. Struhl. 1986. Functional dissection of a
eukaryotictranscriptional activatorprotein, GCN4 ofyeast. Cell
46:885-894.
20. Ingles, C.J., M. Shales,W. D. Cress, S. J. Triezenberg, and J.
Greenblatt.1991. Reduced binding of TFIIDtotranscriptionally
compromisedmutantsofVP16. Nature (London)351:588-590.
21. Jones,N. 1990. Transcriptional regulation by dimerization: two
sidestoanincestuousrelationship. Cell 61:9-11.
22. Keller,A., E.D. Garrett,and B. R.Cullen. 1992. The Bell protein
ofhuman foamy virus activates human immunodeficiency virus
type 1 gene expression via a novel DNA target site. J. Virol.
66:3946-3949.
23. Keller,A., K.M. Partin,M. Lochelt, H. Bannert, R. M. Flugel, and
B. R. Cullen. 1991. Characterization of the transcriptional
trans-activator ofhuman foamy retrovirus.J. Virol. 65:2589-2594.
24. Kunkel,T. A.,J. D. Roberts,and R. A. Zakour. 1987. Rapidand
efficient site-specific mutagenesis without phenotypic selection.
MethodsEnzymol. 154:367.
25. Lee,A. H., K.J. Lee, S.Kim, andY.C. Sung.1992. Transactivation
of human immunodeficiency virus type 1 long terminal
repeat-directed geneexpression by the human foamy virus Bell protein
requiresaspecificDNAsequence.J.Virol. 66:3236-3240.
26. Lee, C. Q., Y. Yun, J. P.
Hoeffler,
and J. F. Habener. 1990. Cyclic-AMP-responsive transcriptional activation of CREB-327 involves interdependent phosphorylated subdomains. EMBO J. 9:4455-4465.27.Lee,K. J., A. H. Lee, and Y. C. Sung. 1993. Multiplepositive and negative cis-acting elements that mediate transactivation by Bell in the long terminal repeat of human foamy virus. J. Virol. 67:2317-2326.
28. Lee, W. S., C. C. Kao, G.0.Bryant, X. Liu, and A. J. Berk. 1991. Adenovirus Ela activation domain binds the basic repeat in the TATA box transcription factor. Cell67:365-376.
29. Leuther, K. K., J. M. Salmeron, and S. A. Johnston.1993. Genetic evidence that an activation domain of
Gal4
does not require acidity and may form a P-sheet.Cell72:575-585.30. Lieberman, P. M., and A. J.
Berk
1991. The Zta transactivator protein stabilizesTFIIDassociation with promoter DNAby direct protein-protein interaction. Genes Dev. 5:2441-2454.31. Lin,Y. S., and M. R. Green. 1991. Mechanism of action of an acidic transcriptional activator in vitro. Cell 64:971-981. 32. Lin,Y. S.,I.Ha, E. Maldonado, D. Reinberg, and M. R. Green.
1991. Binding of general transcription factor TFIIB to an acidic activating region. Nature (London) 353:569-571.
33. Liu, X., C. W. Miller, P. H.
Koefflier,
and A. J.Berk.
1993.The p53 activation domain binds the TATA box-binding polypeptide in holo-TFIID, and a neighboring p53 domain inhibitstranscription. Mol. Cell. Biol. 13:3291-3300.34. Lochelt, M., H. Zentgraf, and R. M. Flugel. 1991. Construction of an infectious DNA clone of the full-length human spumaretrovirus genome and mutagenesis of the bell gene. Virology 184:43-54. 35. Ma, J., and M. Ptashne. 1987. Deletion analysis of GAL4 defines
two transcriptional activating segments. Cell 48:847-853. 36. Maina, C. V., P. D. Riggs, A. G. Grandea, B. E. Slatko, L. S.
Moran, J. A. Yagliamonte, L. A.McReynolds, and C. Guan. 1988. A novel vector to express and purify foreign protein in
E.
coliby fusion to, and separation from, maltose binding protein. Gene 74:365-373.37. McKnight, S., and R. Tjian. 1986. Transcriptional selectivity of viral genes in mammalian cells. Cell 46:795-805.
38. Mergia, A., L. W. Renshaw-Gegg, M. W. Stout, R. Renne, and0. Herchenroeder. 1993. Functional domains of the simian foamy virus type 1 transcriptional transactivator (Taf). J. Virol. 67:4598-4604.
39. Mermod, N., E. A. O'Neill, T. J. Kelly, and R. Tjian. 1989. The proline-rich transcriptional activator of CTF/NF-1 is distinct from the replication and DNA binding domain. Cell 58:741-753. 40. Miller, J. H. 1972. Experiments in molecular genetics. Cold Spring
Harbor Laboratory, Cold Spring Harbor, N.Y.
41. Ptashne, M. 1988. How eukaryotic transcriptional activators work. Nature (London)335:683-689.
42. Ptashne, M., and A. F. Gann. 1990. Activators and targets. Nature (London) 346:329-331.
43. Queen, C., and D. Baltimore. 1983. Immunoglobulin gene tran-scription is activated by downstream sequence element. Cell 33:741-748.
44. Regier, J. L., F. Shen, and S. J. Triezenberg. 1993. Pattern of aromatic and hydrophobic amino acids critical for one of two subdomains of the VP16 transcriptional activator. Proc. Natl. Acad. Sci. USA 90:883-887.
45. Rethwilm, A., 0. Erlwein, G. Baunach, B. Maurer, and V. ter Meulen. 1991. The transcriptional transactivator of human foamy virus maps to the bell genomic region. Proc. Natl. Acad. Sci. USA 88:941-945.
46. Rethwilm, A., K. Mori, B. Maurer, and V. ter Meulen. 1990. Transacting transcriptional activation of human spumaretrovirus LTR in infected cells. Virology 175:568-571.
47. Robbins, J., S. M. Dilworth, R. A. Laskey, and C. Dingwall. 1991. Two interdependent basic domains in nucleoplasmin nuclear targeting sequence: identification of a class of bipartite nuclear targeting sequence. Cell 64:615-623.
48. Sanger, F., and A. R. Coulson. 1975. A rapid method for deter-mining sequences in DNA by primed synthesis with DNA poly-merase. J. Mol. Biol. 94:444-448.
49. Silver, P. A. 1991. How proteins enter the nucleus. Cell 64:489-497. J.VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
50. Stringer, K. F., C. J. Ingles, and J. Greenblatt. 1990. Direct and selective binding ofan acidic transcriptional activation
do-maintothe TATAbox factorTFIID. Nature (London) 345:783-786.
51. Tanaka,M.,and W. Herr. 1990. Differential transcriptional acti-vation by Oct-1 and Oct-2: interdependent activation domains induce Oct-2phosphorylation. Cell 60:375-386.
52. Van Hoy, M., K. K. Leuther, T.Kodadek, and S. A. Johnston. 1993. The acidic activation domains of the GCN4 and GAL4
proteinsarenota-helicalbut form 1-sheets.Cell72:587-594. 53. Venkatesh,L.K., and G. Chinnadurai. 1993. Thecarboxy-terminal
transcriptionenhancementregionofthe humanspumaretrovirus
trans activator contains discrete determinants of the activator function. J. Virol.67:3868-3876.
54. Venkatesh,L.K.,C.Yang,P.A.Theodorakis,and G. Chinnadu-rai. 1993. Functional dissection of the human spumaretrovirus transactivator identifies distinct classes ofdominant-negative
mu-tants.J.Virol. 67:161-169.