JOURNALOF
VIROLOGY,
Dec. 1993,p. 6937-6944 0022-538X/93/126937-08$02.00/0Copyright © 1993. American Society for Microbiology
Increased Spacing
between Spl and
TATAA
Renders
Human
Immunodeficiency Virus Type
1
Replication
Defective:
Implication
for Tat Function
LI-MIN HUANGAND KUAN-TEH JEANG*
Laboratory of Molecular Microbiology, Building
4, Room306,
NationalInstitulte
of
Allergy andInfectious
Diseases,
Bethesda,
Maryland
20892
Received 18 June 1993/Accepted23August 1993
Expression of the human immunodeficiency virus type 1 (HIV-1) is strongly activated byTat. The proper
action of Tat requires three elements: TATAA, TAR, andupstream motifs intheHIV-1 longterminal repeat. We show here that the correct spatial arrangement among Tat, Spl, and TATAA crucially influences HIV expression. Under conditions inwhich basal promoter activity is unperturbed, distancing
Spl
from TATAA markedly affected Tattrans activation. An increase in the Spl-TATAA distance from 18 to 101 nucleotides (depending on the inserted sequence) rendered HIV-1 either partially or wholly replication defective. This critical dependence onspacing suggests that Tat-, Spl-, and TATAA-binding factors mustcorrectly contact each other foroptimal expression and replication of HIV-1.Human immunodeficiency virustype 1 (HIV-1) is the
etio-logicalagentofAIDS (11, 25). Like other retroviruses, HIV-1
reverse transcribes an RNA genome into double-stranded
DNA, which integrates into the host chromosome. Under normalconditions, the integrated provirus expressesits genes
and replicates productively (21, 44). All HIV-1 genes are
controlledbyasinglepromoterlocated in the 5' long terminal repeat (LTR). The transcriptional activity of this LTR is determined by cellular factors (13, 14) thatcooperatewith the viralTat protein (22, 23, 29, 34, 39). In the presence of Tat, transcription from the HIV-1 LTR is increased 100- to 1,000-fold. Without Tat, HIV-1 is defective for expression and replication (9, 30).
Although the precise mechanism of Tat function is being investigated, three elements in the LTRare required for Tat
action:TAR, TATAA, andupstream motifs (2, 4, 35, 37, 40). TARisanRNAelement found in Rwhich formsa
stem-bulge-loop configuration after it is transcribed nascently (3, 5, 32, 37). Tat attaches to TAR RNA and is thereby tethered to the vicinity of the LTR promoter (2, 38, 42). The other two elements, TATAA (12) andupstreammotifs (18, 24, 33),are
sites that Tat (once bound to TAR) interacts with in the
processoftransactivation.
In the HIV LTR, the three Spl binding sites and the two NF-KBbinding sites (18, 24, 33)arefunctionally important in
basaland intrans-activated expression (4). In the subgenomic context,NF-KB contributesmoretobasal expression, andSpl is more important for Tat responsiveness (4). Tat poorly activatesaminimal HIV-1 promoterthatdoesnothave either
Spl
orNF-KB motifs (2, 4, 41). Addition of enhancer-bindingsitescritically reconstitutes responsiveness toTat (2, 4). Two roles can be considered for enhancers in Tat trans
activation. First, enhancersmaysimply prime thepromoterto provide a basal level of transcription to make TAR RNA available for Tat attachment. Second, enhancer-binding
pro-teinscould be apartofamultiprotein complex which includes
cellular factors and Tat. Correct formation of this complex could thus leadto enhancedtranscription. In this setting, Tat
*Correspondingauthor.
trans activation occurs with the participation of cellular
tran-scription factors. One test of this is to determine whether spacing changes between enhancers and TATAA that donot affect basal activity canaffectTat-inducedtransactivation. We have therefore analyzed HIV-1 promoters (both in sub-genomic settings and in the context of infectious virus) with different distances between Spl and TATAA. We found spacings that maintained thebasalactivityof the promoterbut dramatically reduced Tat-inducedtransactivation.Thus, there is aspacing requirement for the Spl-TATAA-Tat interaction
which is distinct from that for the Spl-TATAA interaction. We also found that the correctorientation ofTat relative to Spl and TATAA is important. These observationsare compatible
withan adaptor-coactivator role for Tat in transcription
com-plex formation.
MATERIALS AND METHODS
Plasmid constructions. All of the plasmids used were
de-rived from p-43CAT (4), which contains HIV-1 LTR se-quences from -43 to
+78,
positioned upstream of the cat gene.Threerestriction sites(from 5'to3', PstI,Sall,
andXbaI)are presentaroundposition -43. Anatural Spl triplet (from HIV-1 LAI)wasinserted into theSall sitetomakep-43Sp/26. Inournomenclature, the number following the slash indicates the distance (in base pairs) between the Spl binding site and TATAA, with the Spl-TATAA distance in HIV-1 clone pNL4-3 (1) being 18 bp. p4GSpTm/26 was modified from
p-43Sp/26
by insertion of four copiesofthe Gal4binding site (5'CGGAAGACTCTCCTCCG3' [15])atthe PstI site and hasa mutated TAR. TARwas mutated by changing the
nucleo-tidesfrom +24to +32 from TGAGCCTGGtoCCTCGGACC, eliminating the normal bulge and loop. pNL4G/101 was de-rived from HIV-1 molecular clonepNL4-3 (1), in which four copiesofthe Gal4binding site (total size, 78nucleotides)were
inserted at the -43 position into both LTRs. pNLUC/101 is similarto pNL4G/101 except that the insertedsequenceis 78 bpofpUC19sequence. Intact molecular clones of HIV-1were
prepared and propagated in Escherichia coli as described previously (25). Gal4-Tatwasobtained from MichaelR.Green (41). Gal4-VP16-expressing plasmid pCMVGal65 was a gift
6937
Vol.67, No. 12
on November 9, 2019 by guest
http://jvi.asm.org/
6938 HUANG AND JEANG
A
TAR
T A A CAT
[image:2.612.66.562.77.449.2]Sp x3 l -
.
---
JL
...'
...-.
-
.---Desig nation
p-43Sp/26
B4.0
3.6
3.2
r-as n/Ra
p-43Sp/47
0
-Fucw
p-43Sp/65 a
m
p-43Sp/84
_u
p-43Sp/1 04 c p-43Sp/1 86
2.8
2.4
2.0
1.6
1.2
0.8
0.4
0.0
3 2
103
= 3
o 2
CF 102
s
0
L) 2
cot
I-0 2
LL
I
~ 00
1 2 I0-1 26 36 47 65 84 104 186
0.5
A
%%
I.,N
A 2 * 4 T
_ A
8 -;
! i
S1V
a
26 36 47 65 84 104 186
Distance between Spl bindingsites and TATA(bp)
p-43Sp/104 p-43Sp/l86
ToT(/Ig)
0 0.5 1 2 4 0 0.5 1 2 4 0 0.5 1 2 4 0 0.5 1i
_0
__io
_,_.
AcCm _
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
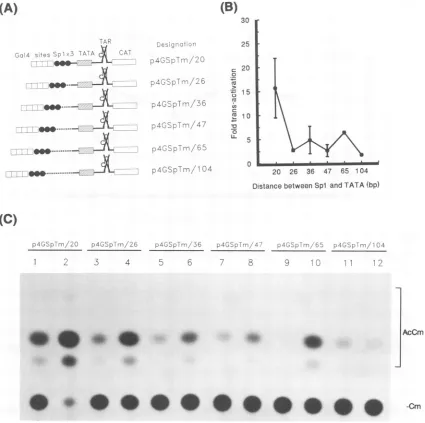
FIG. 1. Increased spacingbetweenSplbinding sitesandTATAA decreases responsiveness ofapromoter toTat. (A) Plasmidconstructs.All
constructscontain HIV-1 (clone pNL4-3)promotersequence upto -43 (mRNAstart site is +1)and threecopiesofSpl binding sites(solid
circles).Sequences frompUC19 of different lengths (dotted lines)wereinserted between the Spl sitesand TATAA.(B)Basalactivities andtrans
activation responsiveness of different constructions. (Left) Relative basal activities of differentpromoters.The activity ofp-43Sp/26wasset at 1. The basal activities of otherpromoters wereexpressed relativetothat ofp-43Sp/26. (Right) Responsiveness of differentpromoters tovarious
amountsofTat,asindicated (inmicrograms). Themeansand standarderrors werederived from threetosix experiments. (C) Representative titration
seriescomparing the trans-activationresponsiveness of p-43Sp/26 (lanes 1to5),p-43Sp/84 (lanes6to10), p-43Sp/104(lanes 11to15),andp-43Sp/186 (lanes 16to20). Subconfluent HeLa cellsweretransfected with 1.5 jigofreporteraloneorwith 0.5, 1, 2,or4 jigof Tat-producing plasmid.Acetylation
of['4C]chloramphenicolwasresolvedby thin-layer chromatography. Cm, chloramphenicol;AcCm, acetylatedchloramphenicol. from PeterO'Hare
(8).
Allplasmid
sequences wereconfirmed
by
DNAsequencing.
Cellculture andtransfection.
HeLa
cells
werepropagated in
Dulbecco's
modified
Eagle's medium with
10%fetal
bovine
serum.The continuous humanT-leukemia cell lines C8166 and
12D7 were grown in RPMI 1640
medium
with 10% fetal
bovine serum. Human
peripheral blood lymphocytes (PBL)
werestimulated
with
phytohemagglutinin (750
ng/ml)
for 4days prior
toinfection
andthen maintained in
RPMI 1640with 10% fetal bovine serumand 10%
interleukin-2
(Pharmacia).Transfections
of
HeLacells
wereperformed
with calciumphosphate (17).
CAT and RT assays.
Chloramphenicol
acetyltransferase(CAT)
assays wereperformed
asdescribed previously (16).
After resolution
by thin-layer chromatography,
radioactivitywas
quantitated
with aFuji
phosphorimager.
For optimalquantitations,
allCAT activities
were reanalyzedin
the linearrange of
acetylation.
Alltransfections
and correspondingas-sayswere donethree tosixtimes.
Reverse
transcriptase (RT)
assays wereperformed
asde-scribed
previously (36).
Each reaction mix contained 5 p.l of viralsupernatant in50
,ul of
RTcocktail [60
mM Tris(pH 8),
75 mM
KCl,
5 mMMgCl2, 0.1%
NonidetP-40,
1 mMEDTA,
5
,ug
ofpoly(rA)
perml,
0.16,ug of
oligo(dT)
perml]
andwasincubatedat
37°C for
1h. Fromeach reaction,
5 p.lwasspotted
ontoDEAE paper, whichwaswashed four
times
in 2xSSC
(1
x SSC is0.15
MNaCl plus 0.015
Msodium
citrate), and
radioactivity
wascounted
inscintillant.
Western
(immunoblot)
analysis
and HIVinfection.
Three molecularclones,
pNL4-3, pNL4G/101, and pNLUC/101 (7
jigof
each),
weretransfectedseparately
into HeLa cells. After 36 to 48h,
cells and supernatants were harvested.Cells
wereanalyzed
forHIV-1proteins, and the
supernatantswere used asvirus stocks forsubsequent infections.For Western
analysis (6),
cellswerewashed withphosphate-buffered saline and
resuspended
in sodiumdodecyl
sulfate(SDS)
solubilization buffer(50
mMTris[pH 7],
2%SDS,
5%glycerol,
10%2-mercaptoethanol). Proteins
wereresolved
inaC
p-43Sp/26 p-43Sp/84
2 4
I
I
I-J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
CRITICAL SPACING FOR
Spl,
TATAA, AND Tat INTERACTION 6939(A)
-.~~~~X
X-
4.
. ._...
_.
......
(B)
30 25
a 20
cu
- 15
~0
.5cU
U0
U-r
c
~.
-:'- -.-. . /- 50
F
F~ -./ ½.Ci'7,rl
/,A 20 26 36 47 65 104
Distance betweenSpi and TATA (bp)
(C)
ApmiUp4Sprn/_O
p4CSp>|ri/6c4QCCSQJrr, T6 s I/2Tn.. t_ c r; ;-' 5;
1 2 3 4 5 6 77 8 9 C I 2
S.
.|
_#*.
*
*S-
-
-
-
-
-AcCm
[image:3.612.96.521.82.505.2]-Cm
FIG. 2. Targeting of Tat to promoter via DNA tethering is less efficient and is sensitive to the distance separating Spl and TATAA. (A) Constructions usedtoassayactivationbyGal4-TatwithGal4sitespositioned upstream of Spl binding motifs. These constructs are derivatives of thep-43Sp plasmids shown in Fig. 1 except that in the starting plasmid, the distance separating Spl and TATAA is 20 instead of 26 nucleotides. Thereare twoothermodifications: (i) four copies of theGal4binding site (open squares) were inserted upstream of Spl and (ii) the TAR sequence wasmutated,rendering it Tat nonresponsive. (B) Behavior of plasmids in response toGal4-Tatfusion protein. Each construction wastransfected aloneorwithasecondplasmid expressingGal4-Tat.Ineach experiment, 1, 2, or 4 ,ug ofGal4-Tat-producingplasmid was used, and the foldtrans activationfor themostoptimalratio of reporter to effector was used. A total of three to six experiments were done. The means and standard errors are shown. (C) Typical thin-layer chromatogram showingtransactivation in HeLacells. Odd-numbered lanes, basal activities; even-numbered lanes, cotransfections withGal4-Tat. Cm, chloramphenicol; AcCm, acetylated chloramphenicol.
10% polyacrylamide-SDS gel
andtransferred
toImmobilon-P(Millipore), which
wasthen blocked with
TENbuffer (10
mM Tris[pH 8],
1 mMEDTA, 50 mMNaCI)
containing2%
nonfatdried
milk.The filters
werethenincubated with
hyperimmune
patient
serumovernight, washed,
andsubsequently
reacted with'25I-labeled
protein
Atovisualize
antibody-antigencom-plexes. Autoradiography
andquantitation
with aFuji
phosphor-imager
wereperformed.
For HIV-1
infections,
viral supernatants were passedthrough
0.45-p.m
filters and normalized for RTactivity.
Cells(4
x106) of
C8166 or12D7
or PBL(3 x106)
in a1-ml
volume wereabsorbed with virus supernatant which contained variousamounts of RT at
37°C. After
Ih,
3.5 ml ofmedium
wasadded. Cell
supernatants weresampled
every 2 or3days for
RT
determination.
Forthe PBLinfections,
1.5 x106
newcells were added every 10days.
RESULTS
Increased spacing between
Spl
and TATAAdramatically
reduces Tat transactivation without
affecting
basalpromoteractivity.
Spi
motifs are foundproximal
to TATAA in the HIV-1 promoter. Since Tat transactivation
is crucial for theexpression
of HIV-1 genes, thisspatial
arrangementsuggestsa-~~~~~~.mdmm
VC)L.67, 1993
-1- - - .:
---t--- --' ..-' ...
-::I1 .- A-7
5
on November 9, 2019 by guest
http://jvi.asm.org/
6940 HUANG AND JEANG
(A)
0
-.
ctI c)
4-0 CZ ct
-o
11
2
1 03
2
102
2
1 01
1
*
Gal4-VP1
6
Gal4-Tat
*
Tat
20 26 36 47 65 104
Distance between Spl and TATA
(bp)
.'4G'3pTmn/;".3 5'Ip rn/I. p4'33pTr%''D '--b4.3.1, 4i ?'_!rn' ',' p4.SSpTn/l 10
, 3 4 45 1T07 8 9 1 1 1 2
0
_
_
.
0
_
_VI
.* *
AcCrr
-Cm
FIG. 3. tranis-activationabilityofGal4-VP16isnotdistance sensitive.(A)Schematic comparisonof tr-ans-activation responsivenessof HIV-1
promoters todifferentversions of Tat. Aplasmid expressingGal4-VP16wascotransfectedwithplasmids containingdifferentspacingbetweenSpl
and TATAA, asshown in Fig. 2A. In each set ofexperiments, 1, 2,or4 pLgofGal4-VPI6-producing plasmidwasused to derive the optimal conditions for transfection.Thegraphshows themeansandstandarderrorsfor threetosixsetsofexperiments.TocompareGal4-VP16withTat andGal4-Tat,the trans-activation resultsfromFig. lBand 2BaresuperimposedontheGal4-VPl6 data.(B) RepresentativeCATassay,showing trans-activation responsiveness toGal4-VPI6. The reporterplasmids areindicated. Odd-numbered lanes,basalactivities; even-numberedlanes, cotransfections with Gal4-VP16. For thevalueusedinpanel A,theassayswererepeatedtoobtain results in the linear range ofacetylation. Cm, chloramphenicol; AcCm,acetylatedchloramphenicol.
role forSpl in Tat function. Indeed,manystudies have shown that
Spl
cooperateswith Tat(4, 27, 41).Tobetter understand the Spl-Tat interaction, we determined whether there is aspatial constraintonSpl withrespecttoTATAA.We
progres-sively increased the distance separatingSpl from TATAA (see constructs in Fig. IA). In this manner, we hoped to differen-tiate between the contribution of Spl to basal transcription
versus Tattransactivation (i.e., whethera distance separating
Spl
from TATAA that affects the latter but not the former could be achieved).Basal activities from our constructions did not change uniformly as the distance separating Spl and TATAA was
increased (Fig. 1B,left). In fact,the highest basal activitywas
seenwhen the distance between the twoelements was65bp. Overall, fluctuations in basal activities from all of the con-structs were less than threefold different from that of the starting construct, p-43Sp/26. Basal activity actually rose
slightly as the distance increased from 104 to 186 bp. These results suggest that the effect of Spl on the basic HIV-1 TATAA promoterremains relatively constant overthe range
of distances tested. However, when the same promoters were
cotransfected with aTat-producing plasmid,asteep declinein Tat-mediatedtransactivationwas seen asthe distance between
Spland TATAAincreased (Fig. IB, right [note log scale]; Fig. IC).Therewas a 103-fold difference inactivityforspacingsof 26 and 186 nucleotides (Fig. lB, right; Fig. IC). Thus, these
(B)
I I I i I I
J. VIRC)I..
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.101.518.82.503.2]CRITICAL SPACING FOR Spl, TATAA, AND Tat INTERACTION 6941
(A)
e 4
O" 7L:- 4
(B) pSp4GTm/1 04 pSp2GTm/65 pSpl GTm/47
1 2 3 4 5 6
]AcCm
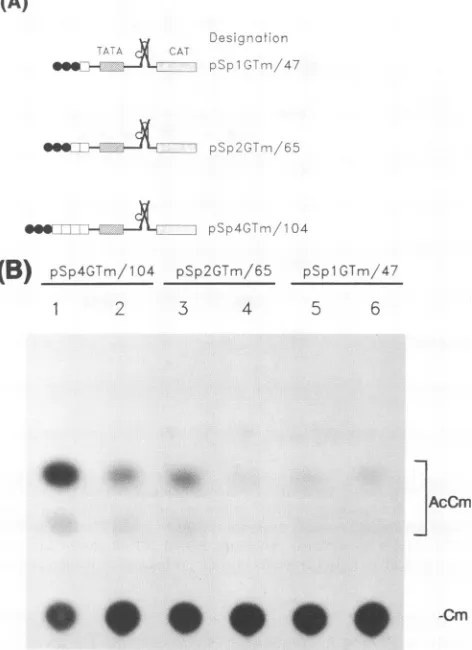
FIG. 4. Ga14-Tat is not functionalwhen placed between Spl and TATAA. (A) Constructions used toassayGal4-Tatactivitywhen the Gal4site(s)wasinserted between SplandTATAA.These constructs
are similar to those shown in Fig. 2A except that one, two, orfour
copies of the Gal4binding site (O) were inserted between Spl and TATAA. (B) Representative thin-layer chromatogram, showinglack
oftrans-activation responsiveness. Odd-numbered lanes,basal activi-ties;even-numberedlanes, cotransfectionswith Gal4-Tat.Cm,
chlor-amphenicol; AcCm, acetylated chloramphenicol.
constructionsclearly segregated the effectsofspacingbetween
Spl
and TATAAfor basalversus Tat-inducedtranscription.Distancebetween
Spl
and TATAAis alsocritical when Tatis targeted to the promoter via DNA binding. In the above experiments,Tatwasintroduced to thepromoterviatethering
to TARRNA. In suchanapproach,whenwemoved
Spl
awayfromTATAA,wealsodistanced
Spl
from Tat. Tocontrol forthe latter variable, we used a second approach that keptthe
distance between Tat and
Spl
constant while varying thedistance between
Spl
and TATAA. We positioned Gal4bindingsitesnext to the
Spl
motifs anddirectedGal4-Tat(41)tothe promoter by usingthe Gal4bindingsites (Fig. 2A).To
make sure that Gal4-Tat functioned only throughGal4 sites,
TAR was mutated so that it would be nonfunctional (see
Materials andMethods).We hadobserved thatwhen Gal4-Tat
was targetedto thepromotervia TARRNA,itwas asefficient
as Tat in trans activation (data not shown). However, when
Gal4-Tat was targeted by using DNA, the efficiency oftrans
activation was decreased 10'- to 102-fold (Fig. 2). This is in
agreementwithourprevious findingswithanalternative DNA
tether (2). Nevertheless, Gal4-Tat did maintain a moderate
level of trans activation (Fig. 2B; Fig. 2C, lanes 1 and 2) that
could be tested for
spatial sensitivity.
In thisinstance,
when weincreased the
distance between Spl and TATAA,
wedid
notaffect the distance between Tat and Spl. This lengthening of
distance did
compromise
Tat transactivation
(Fig. 2B and C),
and this result
supports the idea that correctspacing
betweenSpl and TATAA is important for optimal Tat function.
However,
because there is
noclear linear
relationship between
distance and loss of
transactivation, it is possible that the
relative
positioning of Spl and
TATAA onthe
DNAhelix is
also
important.
To
control
for the possibility that the Gal4 binding sites
might be
inherently malpositioned,
wechecked for the ability
of Gal4-VP16
toactivate the
plasmids shown
inFig. 2A. Potent
trans
activation by Gal4-VP16
wasobserved, and this activation
was not
sensitive
tothe distance between
Spi
and
TATAA(Fig. 3). This result
suggeststhat
Gal4-Tat and Gal4-VP16
differ in their
spatial
restrictions
(Fig. 3A)
and
is consistent
with observations from others that both
Tatand VP16
caninitiate
transcription (but in
nonidentical manners) (43).
Orientation
of Tatrelative
toSpl
isimportant.
The aboveexperiments
address the issue of the influence of spacingbetween
Spl and
TATAA on Tat tranis activation. Wewon-dered whether
the orientation of Tat relative
toSpl and
TATAA
could be
similarly important.
Wehave demonstrated
previously that
aspecific orientation of
Tatwhen
presented
from
TAR RNAis
crucial for function
(2).
Tofurther
explore
this from
aDNA-binding perspective, instead of positioning
Gal4-Tat
upstreamof
Spl,
weinterposed
Gal4
binding
sites
between
Spl
and TATAA
(Fig. 4A). Interestingly,
wefound
that
in this
configuration, Gal4-Tat
wascompletely inactive
(Fig. 4B, lanes I and 2).
Inthe
pSp4GTm/104 plasmid, the
distance between
Spl and
TATAAis
104bp.
Wedecreased the
Spl-TATAA
distance
to47
and 65 nucleotides
(Fig.
4A,
pSplGTm/47 and pSp2GTm/65,
respectively). Gal4-Tat still
did
notactivate these
two constructs(Fig. 4B, lanes
3 to6).
Control
experiments with
aGal4-VP16
plasmid
demonstratedthat
pSp4GTm/104, pSplGTm/47,
andpSp2GTm/65
were allfully
activatedby
VP16(data
notshown). Thus,
Tatactivation
of
the HIV-1 promoteris
orientation
specific for Spl
andTATAA, whereas
VP16activation is
not.Spacing
betweenSpl
and TATAA is critical for the virus.The
distance between Spl and
TATAAprofoundly influences
Tat
responsiveness in the subgenomic
HIV-1 context (Fig. 1 and2).
Toextrapolate this
finding
tothe
virus,
weengineered
two
molecular clones of
HIV-1clone
pNL4-3,
inwhich the
distance between Spl and
TATAAinboth the 5' and 3'
LTRs wasincreased
to 101bp (Fig. 5A).
Twodifferent
DNAs wereinserted
asspacers.pNL4G/101 contained
fourcopies of
Gal4
binding sites positioned between
Spl and TATAA, whilepNLUC/101 contained
arandomly selected
78-bp sequencefrom
pUC19.
Other thanthis
difference,
the twoclones
wereisogenic.
pNLUC/101, pNL4G/101,
andpNL4-3
wereindependently
transfected into
HeLacells.
Viralprotein synthesis,
superna-tantviral
RTproduction,
andthe
ability
to propagate aspreading infection
wereassessed
for each of the three
molec-ular
clones(Fig.
5and6).
At48 hafter transfection into
HeLacells,
wefound
(on
apermicrogram
ofinput
DNAbasis)
lessproduction
of viralproteins from pNLUC/101
andpNL4G/101
than from
pNL4-3. Supernatant
RTproduction
wasfound at relativeratios
of1,
3.5,
and 14for
pNLUC/101, pNL4G/101,
and
pNL4-3,
respectively (Fig. 5B);
relativeprotein (p24
andp55)
ratios
were1,
3.5,
and7.2,
respectively (Fig.
5B).
We next
assayed
theinfectivity
of the three viruses. Threedifferent cell lines
(PBL,
12D7[CEM],
andC8166)
wereused
(Fig. 6). Despite normalization of input virus
at the startof
VOL.67. 1993on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.63.299.84.409.2]6942 HUANG AND JEANG
W%
.Q Q C5 K
I.\
..v I
?,
,q
B
!.
ir
A
Designation U3 RU5
pNL4-3 U3 RU5
puc 19
sequence
seqIMNdIuI l|I pNlUC//101
69.8-No_
oI
<--p55
43.3-
28.3-*-p24
Gal4 binding
sites
"i
!
pNL4G/10118.1-RT--
p [image:6.612.104.513.80.318.2]1 2 3
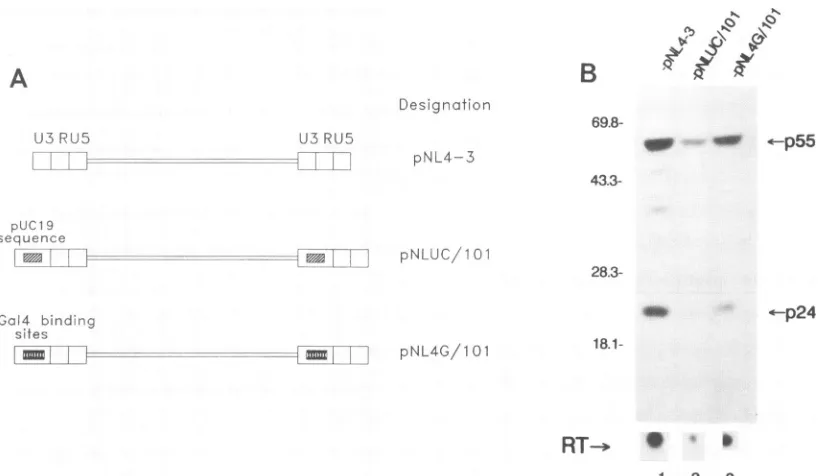
FIG. 5. HIVswith increased Spl-TATAA intervals show poor expression. (A) Schematics of threeHIV-1molecular clones. In bothpNL4G/101 and
pNLUC/101,
the distance between Spl and TATAA was increased to 101 bp. The spacer was either four copies of the Gal4binding site(pNL4G/101)
or 78bpof pUC19 sequence (pNLUC/101). (B) Increased Spl-TATAA spacing reduces viral expression. A typical Western analysis is shown. Positions of molecular size markers are indicated to the left of the gel (in kilodaltons). The supernatant RT produced for each transfection is shownunder the lanes. Thepositionsofp55and p24 proteins are indicated. Equal amounts of totalproteins were loaded in each lane.infection in all three cell lines,
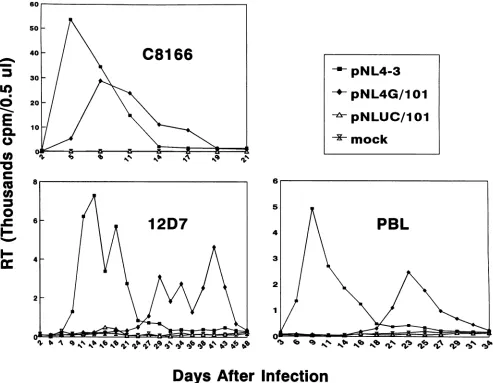
pNLUC/101
was severelydelayed in peak RTproduction (no peak RT detected during the course of the experiment), while pNL4G/101 was
moder-ately delayed (3 to 15 days) (Fig. 6). Reinfection of cells with virus harvested at peak RTproduction (for
pNL4G/101)
and with medium supernatant at the end of 6 weeks of infection (forpNLUC/101) reproduced the same kineticpatterns (data notshown). Polymerase chain reaction analysis of thepNL4G/ 101 virus alsodemonstrated that the insertedspacersequence was retainedstably (19a).DISCUSSION
Although it is well accepted that Tat is a transcriptional
activator, the mechanism of Tatactionremainsunclear(20-22, 31, 34, 39). One way to appreciate the mechanism is to understand the immediate interactive sites for Tat in the promoter(4). For example, it has been shown thatTatpoorly activates an HIV-1 minimal promoter which has intact TATAAand TAR elements (2, 4). Providing enhancer bind-ing-sites conferred optimal Tat responsiveness on the pro-moter (4, 27, 41). Inparticular, Spl sites have been shown to most effectively augment Tat function (4, 27). Our current study extends these observations by characterizing the spacing and relative orientation requirements for
Spl,
TATAA, and Tat for function. We also show that correct spacing between SpIandTATAAin theproviral LTR isveryimportant for viralinfectivity.
How do Spl, TATAA, and Tat interact? One possibility is that these three componentscoalesce to form a multiprotein
transcription complex. The fact that Spl and TATAA interact functionally (4, 18) isnotsurprising, since Spl motifsarefound
ubiquitously upstream ofmanyTATAA elements (26). How Tat interplays with this process is intriguing. In our
experi-ments, we
found
adramatically
steepdrop in
Tat-mediated
trans
activation when
weprogressively separated
Spl farther
from
TATAA(Fig.
1B, right).
Atthe
sametime, the effect of
Spl
onthe basal
activity of
the promoterremained
relatively
unimpaired
(Fig.
IB, left).
These
observations
suggestthat
Tat"bridges" SpI and
TATAA(similar
tothe
function of
anadaptor-coactivator [10]). The fact that
afunctional
Spl-Tat-TATAA
interaction is orientation sensitive
(Fig.
4) is also
consistent with
abridging
rolefor
Tat.Biochemical evidence
that
Tatphysically binds Spl
(22a)
and that
Tatalso
binds
TFIID
(Sa)
supportsthis
bridging
hypothesis.
The bridging
hypothesis
alsoexplains why
Tatand VP16 canboth initiate
transcription
but indifferent
ways(43).
We suggestthat while
VP16is
adirect activator
of
aminimal
TATAA promoter, Tat does not
activate
directly
butinstead
"adapts"
the
effectof
an upstream factor(e.g.,
Spl)
tothe
TATAA promoter.
Thus,
Tatrequires Spl (4, 27,
41),
aspecific
spacing between Spl and
TATAA(Fig.
1),
and
aspecific
orientation relative
toSpl
(Fig.
4). VP16, in
contrast,needs
noneof
these.Finally,
weverified
thespacing
requirements
in the context of HIV-1. Wetransferred
ourSpl-TATAA moieties into
infectious molecular clones of
HIV-1(Fig.
6) and found that,
in
thesetting of virus
replication,
a distance of 101 bases betweenSpl
and TATAAdramatically
changed
virusgrowth.
In three
T-lymphocyte
cell types(PBL,
12D7,
and
C8166),
thereplication
ofviruses
with anSpl-TATAA distance of
101bp
was
moderately
(pNL4G/101)
orseverely
(pNLUC/101)
de-layed (no
replication detected by
supernatant RT at 6weekspostinfection).
The reasonsfor
thedifferences
seenwhen the spacer was fourcopies
of the Gal4binding
site or 78bp
of
pUC19
sequence are notclear.
However, both results
areclearly consistent
with theinterpretation that increased
spacing
J. VIROL.
"--.4 = ..I
on November 9, 2019 by guest
http://jvi.asm.org/
CRITICAL SPACING FOR Spl, TATAA, AND Tat INTERACTION 6943
~~~~~~~~~~~~p
NL4-3LO
so
pNL4G/1
01
020-E
10 +<pNLUC/lO0
-~mock
1 i
C 8
co
0 ~~~~~~~~~~~~~5
0
12D7 PBL
I-=
4 3
Days After
Infection
FIG. 6. Viruseswith increased Spl-TATAA spacing in U3 replicate poorly. Replication kinetics of thethreeviruses fromFig. 5Aareshown.
Cellularsupernatantscontaining5 x 105cpmof RTactivitywereusedtoinfect 12D7and C8166cells.For PBLinfection,supernatantscontaining 106cpmof RTwereused. Althoughnotshown, allinfectionswerecarriedoutfor 6weeksbefore termination. The growthcurvesfor pNL4-3 and
pNLUC/101inC8166 and 12D7 cellswererepeatedtwice.
between
Spl
andTATAA
negatively affects virus replication.
Thecrucial
consideration
ofspacing for
HIV-1 LTRfunction
is
important
inlight
ofrecentcontroversies
overthe need for
NF-KB
and/or Spl
elementsfor
virus viability (28,
30,36).
Differences in
competing studies
arelikely
explainable by the
factthat
mutations
inthe
respective
NF-KBand
Spl sites did
notconservespacing relative
toTATAAin the
provirus clones.
Itis
important
tonotethat insertions of
sequencesinto
theHIV-1 U3
could affect
polyadenylation.
Wehave
examined
transcripts
produced from
thepNL4G/101 virus and found that
they
werepolyadenylated
at theexpected position (data
notshown).
Inaddition,
recentstudies by
Valsamakis
et al.(45)
and
Cherrington
andGanem (7)
suggestthat
ourparticularinsertions in
U3
wouldnothave
amajor
effectonpolyadenyl-ation.
ACKNOWLEDGMENTS
WethankAnneGatignol, Oliver Semmes, and Lung-Ji Chang for readingsof themanuscript.
This work was supported in part by the Council for Tobacco Research-USA, Inc.,andbytheAIDStargeted antiviralprogramfrom
the Officeof the Director of the NIH. REFERENCES
1. Adachi, A.,H. E.Gendelman, S. Koenig, T. Folks,R.Willey, A. Rabson,and M. A. Martin. 1986. Productionofacquired immu-nodeficiencysyndrome-associated retrovirus in human and
non-human cells transfected with an infectious molecular clone. J.
Virol.59:284-291.
2. Berkhout,B.,A.Gatignol,A. B.Rabson,and K.-T.Jeang. 1990. TAR-independent activation of theHIV-1LTR: evidencethat Tat requires specific regions ofthepromoter.Cell62:757-767. 3. Berkhout, B., and K.-T. Jeang. 1989. trans activation of human
immunodeficiency virustype 1 is sequencespecific for both the single-stranded bulge and loop of the trans-acting-responsive hairpin:aquantitative analysis. J. Virol. 63:5501-5504.
4. Berkhout, B., and K-T. Jeang. 1992. Functional roles for the
TATApromoterand enhancers in basal and Tat-induced
expres-sionof humanimmunodeficiencyvirustype1longterminalrepeat.
J.Virol.66:139-149.
5. Berkhout, B., R. H. Silverman, and K.-T. Jeang. 1989. Tat trans-activates the human immunodeficiencyvirus through a
na-IrI VOL.
67,
1993on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.66.559.85.468.2]6944 HUANG AND JEANG scent RNA target. Cell 59:273-282. Sa.Brady, J. Personal communication.
6. Burnette, W. N. 1981. Western blotting: electrophoretic transferof proteins from sodium dodecyl sulfate-polyacrylamide gels to un-modified nitrocellulose and radiographic detection withantibody andradioiodinated protein A. Anal. Biochem. 112:195-203. 7. Cherrington, J., and D.Ganem. 1992. Regulation of
polyadenyl-ation in human immunodeficiency virus (HIV): contribution of promoter proximity and upstream sequences. EMBO J.
11:513-1524.
8. Cousens, D. J., R. Greaves, C. R. Goding, and P. O'Hare. 1989. The C-terminal 79 amino acids of the herpes simplex virus regulatory protein, Vmw65, efficiently activate transcription in yeast and mammalian cells in chimeric DNA-binding proteins. EMBO J.8:2337-2342.
9. Dayton, A. J., G. Sodroski, C. A. Rosen, W. C. Goh, and W. A. Haseltine.1986. The trans-activator gene of human T-cell lympho-tropic virus type III isrequiredfor replication. Cell 44:941-947. 10. Dynlacht, B. D., T. Hoey, and R. Tjian. 1991.Isolation of cofactors
associatedwith the TATA-binding protein thatmediate transcrip-tionalactivation. Cell 66:563-576.
11. Fauci, A. S. 1988. The human immunodeficiency virus: infectivity andmechanisms of pathogenesis. Science 239:617-622.
12. Garcia, J. A., D. Harrich, L. Soultanakis, F. Wu, R. Mitsuyasu, and R. B. Gaynor. 1989. Human immunodeficiency virus type 1 LTR TATA and TAR region sequences required for transcrip-tionalregulation. EMBO J. 8:765-778.
13. Gatignol, A., C. Buckler, and K.-T. Jeang. 1993. Relatedness of an RNA-bindingmotif in human immunodeficiency virus type I TAR RNA-binding protein TRBP to human PI/DsI kinase and Dro-sophilastaufen. Mol. Cell. Biol. 13:2193-2202.
14. Gatignol, A., A. Buckler-White, B. Berkhout, and K.-T. Jeang. 1991. Characterization of a human TAR RNA-binding protein that activates the HIV-1 LTR.Science 251:1597-1600.
15. Giniger, E., S. M. Varnum, and M. Ptashne. 1985. Specific DNA binding of Gal4, a positive regulatory protein of yeast. Cell 40:467-74.
16. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982. Recombi-nant genomeswhich expresschloramphenicolacetyltransferase in mammalian cells. Mol. Cell. Biol. 2:1044-1051.
17. Graham, F. L., and A. J. Van der Eb. 1973.A newtechnique for the assay of infectivity of human adenovirus 5 DNA. Virology 52:456-467.
18. Harrich, D., J. Garcia, F. Wu, R. Mitsuyasu, J. Gonzalez,and R. B. Gaynor. 1989.Role ofSPI-binding domains in in vivo transcrip-tional regulation of the humanimmunodeficiencyvirus type Ilong terminal repeat. J. Virol. 63:2585-2591.
19. Ho, D. D., R. J. Pomerantz, and J. C. Kaplan. 1987.Pathogenesis ofinfectionwith humanimmunodeficiency virus. N. Engl.J.Med. 317:278-286.
19a.Huang, L.-M. Unpublished data.
20. Jeang, K.-T., and B. Berkhout. 1992. Kinetics of HIV-1 long terminal repeat trans-activation: use of intragenic ribozyme to assessrate-limiting steps. J. Biol. Chem. 267:17891-17899. 21. Jeang, K.-T., B. Berkhout, andB.Dropulic.Effects ofintegration
and replication on transcriptionof theHIV-1 LTR.J.Biol.Chem., in press.
22. Jeang, K.-T., Y. Chang, B. Berkhout, M. Hammarskjold, and D. Rekosh. 1991.Regulation ofHIVexpression: mechanism of action of Tat and Rev. J. Acquired Immune Defic. Syndr. 5(Suppl. 2):S3-S14.
22a.jeang, K.-T., R. Chun, N. H. Lin, A. Gatignol, C.G.Glabe,and H. Fan. 1993. In vitro and in vivo binding of human immunodefi-ciency virus type 1 Tat protein and Spl transcription factor. J. Virol. 67:6224-6233.
23. Jeang, K.-T., P. R. Shank, and A. Kumar. 1988. Transcriptional activation ofhomologous viral longterminal repeats by the human immunodeficiency virus type 1 orthe human T-cellleukemia virus type I Tat protein occurs in the absence of de novo protein synthesis. Proc. Natl. Acad. Sci. USA 85:8291-8295.
24. Jones, K. A., J. T. Kadonaga, P. A. Luciw, and R. Tjian. 1986. Activation of the AIDS retrovirus promoter bythe cellular
tran-scription factor, Spl. Science 232:755-759.
25. Joshi, A., and K.-T. Jeang. 1993.Reduction in growth temperature minimizes instability of large plasmidscontaining HIV-1 proviral genomes.
Biotechniques
14:883-884.26. Kadonaga, J. T., K. A. Jones, and R. Tjian. 1986. Promoter-specific activation of RNA polymerase II transcription by Spl. Trends Biochem.Sci. 11:20-23.
27. Kamine, J., and G. Chinnadurai. 1992. Synergisticactivation of human immunodeficiency virus type 1 promoter by the viral Tat protein and cellular transcription factor Sp]. J. Virol. 66:3932-3926.
28. Kim, J. Y., F. Gonzalez-Scarano, S. L. Zeichner, and J. C. Alwine. 1993. Replication of type I human immunodeficiency viruses containing linker substitution mutations in the -201 to -130 region of the longterminal repeat. J. Virol. 67:1658-1662. 29. Laspia, M. F., A. P. Rice, and M. B. Mathews. 1989. HIV-1 Tat
protein increases transcriptional initiation and stabilizes elonga-tion. Cell 59:283-292.
30. Leonard, J., C. Parrott, A. J. Buckler-White, W. Turner, E. K. Ross,M. A.Martin, and A. B. Rabson. 1989.The NFKB binding sites in the human immunodeficiencyvirus type 1 long terminal repeatarenotrequiredfor virus infectivity. J. Virol. 63:4919-4924. 31. Marciniak, R. A., B.J. Calnan, A. D. Frankel, and P. A. Sharp. 1990. HIV-l Tat proteintrails-activates transcriptioninvitro.Cell 63:791-802.
32. Muesing, M. A., D. H. Smith, and D. J. Capon. 1987. Regulation of mRNA accumulation by a human immunodeficiency virus trans-activator protein. Cell 48:691-701.
33. Nabel, G., and D. Baltimore. 1987. An inducible transcription factoractivatesexpression of humanimmunodeficiency virus in T cells. Nature(London) 326:711-713.
34. Pavlakis, G. N., and B.K. Felber. 1990.Regulation ofexpression of humanimmunodeficiency virus. New Biol. 2:2t)-31.
35. Rosen, C. A., J. G. Sodroski, and W. A. Haseltine. 1985. The location of cis-acting regulatory sequences in the human T-cell lymphotropic virus type III (HTLV-III/LAV) long terminal re-peat.Cell 41:813-823.
36. Ross,E.K.,A.J. Buckler-White,A. B.Rabson, G.Englund,and M. A. Martin.1991.Contribution of NFKB and Splbindingmotifs tothe replicativecapacityof human immunodeficiencyvirus type 1:distinct patterns of viral growtharedetermined by T-cell types. J. Virol. 65:4350-4358.
37. Selby, M.J., E. S. Bain, P. A. Luciw, and B. M. Peterlin. 1989. Structure, sequence, and position of the stem-loop in TAR determine transcriptional elongation by Tat through the HIV-1 longterminal repeat. Genes Dev. 3:547-558.
38. Selby, M.J., and B. M.Peterlin. 1990.trans-activationbyHIV-1 Tat viaaheterologous RNAbinding protein.Cell 62:769-776. 39. Sharp, P. A., and R. A. Marciniak. 1989. HIV TAR: an RNA
enhancer? Cell 59:229-230.
40. Sodroski, J. G.,R.Patarca,C. A.Rosen,F.Wong-Staal,and W.A. Haseltine. 1985. Location of the trans-activating region on the genome of human T-cell lymphotropic virus type III. Science 229:74-77.
41. Southgate, C. D.,and M. R.Green. 1991.TheHIV-1 Tatprotein activatestranscription fromanupstreamDNA-bindingsite: impli-cations forTatfunction. Genes Dev.5:2496-2507.
42. Southgate,C.D., M. L.Zapp,andM. R. Green. 1990. Activation oftranscription by HIV-1 Tat protein tethered to nascent RNA through anotherprotein. Nature (London) 345:640-642. 43. Tiley, L. S.,S.J. Madore,M. H. Malim, and B. R.Cullen. 1992.
The VP16 transcription activation domain is functional when targeted to a promoter-proximal RNA sequence. Genes Dev. 6:2077-2087.
44. Vaishnav, Y. N., and F. Wong-Staal. 1991. The biochemistry of AIDS. Annu. Rev. Biochem. 60:577-630.
45. Valsamakis, A., S. Zeichner, S. Carswell, andJ.C.Alwine. 1991. The humanimmunodeficiencyvirustype 1polyadenylation signal: a 3' long terminal repeat element upstream of the AAUAAA necessaryforefficientpolyadenylation.Proc.Natl. Acad. Sci. USA 88:2108-2112.
J
![FIG.1.circles).constructsTheactivationseriesamountsof(lanes ['4C]chloramphenicol Increased spacing between Spl binding sites and TATAA decreases responsiveness of a promoter to Tat](https://thumb-us.123doks.com/thumbv2/123dok_us/1296748.82712/2.612.66.562.77.449/circles-constructstheactivationseriesamountsof-chloramphenicol-increased-spacing-decreases-responsiveness-promoter.webp)