0022-538X/82/100054-13$02.00/0
Structure
and
Biochemical
Functions of
Four
Simian Virus
40
Truncated
Large-T
Antigens
FURZANACHAUDRY,ROBERT HARVEY, AND ALAN E. SMITH*
Biochemistry Division,National Institute for Medical Research, Mill Hill, London NW7 IAA, United
Kingdom
Received 30 April 1982/Accepted 22 June 1982
Thestructureof four abnormalTantigens whichare presentin differentsimian
virus 40 (SV40)-transformed mouse cell lines was studied by tryptic peptide
mapping, partialproteolysis fingerprinting, immunoprecipitationwith monoclonal
antibodies, and in vitro translation. The results obtained allowedus todeducethat
theseproteins, which haveapparent molecular weights of15,000, 22,000, 33,000
and45,000, aretruncatedforms of large-T antigen extendingtodifferent amounts into the aminoacidsequencesuniquetolarge-T.Theproteinsareall
phosphory-lated,probablyatasite between amino acids106and123.ThemRNAscoding for the proteins probably contain the normal large-T splice butare shorter than the normal transcripts of theSV40 early region. The truncated large-Ts weretested for the ability to bindto double-stranded DNA-cellulose. This showed that the 33,000- and45,000-molecular-weight polypeptidescontained sequences sufficient forbindingunder theconditions used, whereas the 15,000- and 22,000-molecular-weight forms did not. Together with published data, this allows the tentative mapping ofaregion ofSV40large-T between amino acids 109 and 272 that is
necessaryandmaybesufficient for the bindingtodouble-stranded DNA-cellulose
in vitro. None of the truncated large-T speciesformeda stable complex withthe
host cell protein referred to as nonviral T-antigen or p53, suggesting that the
carboxy-terminal sequencesoflarge-Tare necessaryforcomplexformation.
The early region of simian virus 40 (SV40) codes fortwoproteinscalledlarge-Tantigenand small-tantigen,withapparentmolecularweights of 94,000 (94K) and 17K (7, 26, 33). Large-T playsapivotalrole in thebiologyofthe virus. It is required for the initiation of viral DNA
syn-thesis, for the stimulation of cellularDNA
syn-thesis, and for the regulation of viral DNA transcription (10, 36, 38). Large-T has also been implicated in both the initiation and maintenance of transformation (10,36,38).Theroleof small-t in bothproductive infection and transformation
isless clear.
Several biochemical activities have been
as-cribed to large-T. It has the ability to bind to
double-strandedDNA(dsDNA) (3),particularly
to aspecific regionneartheorigin ofreplication
on SV40 DNA (20, 21, 35). Large-T also
inter-actswithahostcellphosphoprotein, referredto as nonviralT-antigen (NVT) or p53, toform a
high-molecular-weight complex (13, 15, 19). In
addition,anATPaseactivity of large-Thas been
demonstrated (37), and an associated protein kinaseactivity hasbeenreported (9).
Thesedata suggest thatlarge-T isa multifunc-tional protein and raise the possibility that the proteinmaybe composed ofanumberof
func-tional domains,eachassociatedwithaparticular activity. Suchaview is supported by work with
mutants which are temperature sensitive for
replication and which have lesions mapping in the early region. Under certain conditions it is possible to separate the transforming function from the temperature-sensitive lesion (27). To date it is not known which of the biochemical activities oflarge-Tand,inconsequence, which of theputativefunctional domainsare associat-ed withitsabilitytotransform cells. The results presented hereare anattempt todefine
function-aldomains onthe large-T molecule.
Immunoprecipitation ofextractsof cells
trans-formedby SV40 withserumfromtumor-bearing animals shows thatthecellsoftencontain
abnor-mal forms of large-T, in addition to large-T, small-t, andNVT(4, 12,18, 32,34).Oneclass of these abnormal T-antigens consists ofproteins considerably larger than normal large-T andare
referredto as super-T(4, 12, 18, 34;M. Lovett, C.E.Clayton,D.Murphy,P. W.J.Rigby,A. E.
Smith, and F. Chaudry, submitted for publica-tion). Another class is composed of truncated forms oflarge-T (18, 32, 34). In this study we
characterized four such truncatedlarge-T
mole-cules and showedthat they containthe normal
54
on November 10, 2019 by guest
http://jvi.asm.org/
SV40 TRUNCATED LARGE-T ANTIGENS 55
amino-terminal region of large-T and differing
amounts of the amino acid sequences uniqueto
large-T. We also measured the ability of the
truncated large-T molecules to bindtodsDNA in
vitro and assayed for their presence in a stable
complex with NVT. Inthiswaywe wereableto
deduce a tentative functional map of a DNA
binding domain on SV40 large-T and to locate
sequences necessary forNVT binding.
MATERIALS ANDMETHODS
Cells. FourSV40-transformedmousecell lineswere
used in this study: MES 2006, obtained from W. C.
ToppatColdSpringHarborLaboratory, ColdSpring
Harbor, N.Y., and SWSV3T3, BALB/c 3T3C120, and
BALB/c 3T3 ClM, which have been described
previ-ously (5, 33, 34).
Antisera. The anti-SV40 tumor cell (anti-T) serum
used was apool of sera obtained fromtumor-bearing
golden hamsters as described previously (33). The
monoclonalantibodies to large-T and NVT werethe
kindgiftof Ed Harlow(11).
Labeling, extraction, andimmunoprecipitation.
Con-ditions for the growth, labelingwith
32Pi,
andextrac-tion oftransformed cells and forimmunoprecipitation
have been described (33, 34). Sodiumdodecyl
sulfate-polyacrylamideelectrophoresiswas on10or15%gels.
Driedgelswereautoradiographedfor 2to3days.
RNA preparation andpurification withSV40
DNA-ceUlulose. Preparation of RNA from cells grown in
culture was exactly as previously described (22).
Polyadenylated RNA was partially purified on SV40
DNA-cellulose aspublished (23).
In vitro translation. The nuclease-treated
reticulo-cytelysatewaspreparedby the procedure of Pelham
and Jackson (24). Conditions for cell-free synthesis
andimmunoprecipitation were asdescribed(22).
Tryptic peptide mapping. [355]methionine-labeled
extracts were immunoprecipitated with anti-Tserum
andsubjectedtoelectrophoresisin12.5%
preparative-scalepolyacrylamide gels. Individual polypeptides in
unfixed dried gels were located by autoradiography.
Elution of polypeptides from gels and subsequent
tryptic digestion and two-dimensional fingerprinting
onthin-layercellulose sheets were aspreviously
de-scribed (33, 34).
Partialproteolysisanalysis.32P-labeledextracts were
immunoprecipitated, and theproteinswereseparated
on a10% polyacrylamide gel. Large-T and the
truncat-edlarge-Tmoleculeswerelocatedbyautoradiography
of the wet gels, excised, and eluted in 100 mM
ammoniumbicarbonate-0.1%sodiumdodecyl sulfate.
The eluted materialwasdiluted,toreduce the
concen-tration of sodiumdodecyl sulfate, and reprecipitated
asdescribed by Lane and Robbins (14). The washed
immunoprecipitatesweresuspended in 120 mM
NaCl-50 mMTris-hydrochloride (pH 8.0)-0.5%Nonidet
P-40and treated with limited amountsof trypsin at 0°C
for 60 minasdescribedby Schwyzeretal. (30). The
32P-labeled digestionproductswereseparatedby
elec-trophoresis and autoradiographed.
DNA-cellulose chromatography. Calf thymus
dsDNA-cellulose (native) was eitherpurchased from
PL Biochemicals or made by a modification of the
method of Albertsetal.(1). Thewetslurry (PL)orthe
dry DNA-cellulose powder was suspended
immediate-ly before use in pH6.2bufferwhich contained 5 mM
potassium phosphate (pH 6.2)-0.1 M NaCI-1 mM
dithiothreitol-0.5% Nonidet P-40-10% (vol/vol)
glyc-erol. The columnwaspoured ina5-mlsyringe (0.5ml,
packed volume) andequilibratedwith 5 volumes ofpH
6.2buffer (3).
32P-labeled extracts (500,ul)wereadjustedtopH 6
with 2 to 5 ,ulof 1 M acetic acid and applied to the
column.Nonboundmaterial wascollected, and
wash-ing was continued until no furtherradioactivity was
detected. The column was then eluted with pH 8.0
elution buffer containing 10 mM Tris-hydrochloride
(pH 8.0), 1 mMdithiothreitol, 10%(vol/vol)glycerol,
and 0.1, 0.3, or 1.0 M NaCl. All fractions were
adjusted to pH 8.0 and 0.15 MNaCl.Portions of each
fraction were takenfor determination of radioactivity,
and 0.5-ml samplesfrom the peaks of eluted material
wereimmunoprecipitated with anti-Tand control sera
and separated onapolyacrylamide gel.
Sedimentation analysis of cellextracts. Samples (0.4
ml)of32P-labeledcellextracts werecentrifuged ona
4.7-mllinear 5 to20%o sucrosegradientwitha0.4-ml
60%osucrosecushion under conditions similartothose
described byCarrolletal.(3).Thegradientscontained
10 mM Tris-hydrochloride (pH 8.0), 10 mM
dithio-threitol, and 0.14 MNaCl and were centrifuged for 15
h at25,000 rpmin an SW50.1 Beckman rotor at4°C.
The gradient fractions (0.24 ml) were collected and
immunoprecipitatedasdescribed(18, 19).
RESULTS
We(18, 34; Lovettetal.,submittedfor
publi-cation) and others (4, 12, 32) have previously
shownthatcells transformed bySV40contain a
numberof proteins that arespecifically
immuno-precipitatedby SV40 anti-T serum. One class of
theseproteinsaresmaller thanlarge-T and have
beenshown to be related to it byanalysisof their
methionine tryptic fingerprints. These proteins
weretentatively identifiedastruncatedformsof large-T (32, 34). Here we analyze the structure offourtruncatedlarge-T antigens inmoredetail. Immunoprecipitationof truncatedlarge-T
anti-gens. Figure 1 shows the phosphoproteins
spe-cifically immunoprecipitated by anti-T serum
from32P-labeledextractsof four differentmouse
cell lines transformed by SV40. In addition to
super-T(140K), large-T(94K), and NVT (53K),
which have all been characterized previously (12, 15, 17, 18, 34; Lovett et al., submitted for publication), a number of other polypeptides
withmolecular weights in the range from 45K to
15K are specifically immunoprecipitated. Four
such proteins with apparent molecular weights of15K, 22K, 33K, and 45K, each of which is synthesized in adifferent cell line,were investi-gated further.Theproteinsarereferredto asthe
15K, 22K, 33K, and 45K species, respectively.
Two-dimensional tryptic fingerprint analysis.
Tocompare the structureoftheseproteins with
thoseoflarge-Tandsmall-t,they wereanalyzed
by two-dimensional tryptic peptide
fingerprint-VOL. 44,1982
on November 10, 2019 by guest
http://jvi.asm.org/
0
Super -
e
Lz-arit: -r--NV
---451K
-3K
15K-s
FIG. 1. Immunoprecipitat tracts of fourmouse celllini
Theextractswereseparated
gel after immunoprecipitati(
serum (C) and hamster anti
BALB/c 3T3CIM;2,BALB/I
4, SWSV3T3, M,molecular
ing. Previous work has s
of large-T and small-t
[35S]methionine-labeled
that each has additionalp
(23, 33). This occurs bec
early region ofSV40DN)
alternative ways, resultir
large-T and small-t. Smal
contiguous sequenceofS'
large-T mRNA is spliced
coding for the carboxy-te
including the termination
peptides common to smE
natefromtheirshared am
tides unique to small-t ar
spliced from the large-T
tides unique to large-T
quences downstream frc
region (23, 34).
The SV40-transformed
with [35S]methionine, anc
tation ofcell extracts, th
ratedon preparative-scal
Theproteinswereeluted:
with performic acid, dige
gerprinted by electrophc
finally chromatographed
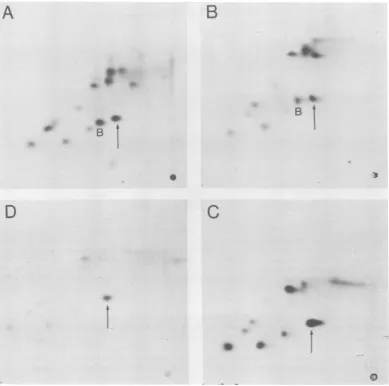
Figure 2 shows the f
large-T and small-t togel
22Kandthe 45Kproteins
the common N-terminal I
Asp-Lys) (18) as well as all the otherpeptides
shared between large-T and small-t. The 22K
protein contains none of the peptides unique to
either small-t or large-T, nor any additional
peptidesnot normallypresentintheT-antigens.
r* -ir><- The 45K
protein
contains severalpeptides
withmobility identical to those of large-T unique
iss^ peptides, including a peptidepreviously referred
to as peptide B (18, 34).
Wehave already shown that the fingerprint of
the 33K polypeptide contains the peptides
sharedbetween large-T and small-t and at least
two large-T unique peptides (34). It does not
containpeptide B. These data establish that the
33K and 45K polypeptides are truncated forms
of large-T, since the three proteins share
com-monpeptides.
As the 22K protein only contains methionine
peptides sharedby bothlarge-Tand small-t,it is
not possible to conclude on the basis of the
tion
3of2
Plabelefingerprint
data that it contains sequences fromson
of P-labeled ex- thelarge-T unique region. This was notsurpris-os
transf
olrmed bylam40ing,
asthe sequence of SV40 DNApredicts
thatonf
with normal hamster fewif
any soluble,methionine-containing
tryptici-T
serum (T). Lanes: 1, peptides are coded for by the region of the'c 3T3C120; 3, MES2006; mRNA fromimmediatelyafter thesplice.We do
weight size markers. know, however, that the 22K polypeptide
con-tains neither small-t unique peptides (23) nor a
methionine-rich tryptic peptide coded in an
al-,hown that fingerprints ternative reading frame in the sequences around
share a number of the splice junction and present in an abnormal
tryptic peptides and small-t coded for by the deletion mutant d12122
eptidesthat are unique (39; M. Kress and F. Chaudry, unpublished
ause transcripts of the data).
kcan be spliced in two To examine further the structure of the 22K
ng in the mRNAs for polypeptide, the
32P-labeled
degradationprod-l-t is translated from a ucts generated by partialproteolysis ofthe 22K
V40DNA,whereasthe protein were compared with those of
normal-to remove sequences sized large-T.
-rminal half of small-t, Partial proteolysis analysis. Partial protdolysis
codon (7). Thus, the fingerprints of SV40 large-T have previously
all-t and large-T origi- been generated by methods devised by Cleve-iino terminus, the pep- land et al. (6, 18, 33) and bySchwyzeretal.(30).
re coded by the region The latter method is particularly useful for the
mRNA, and the pep- analysis of32P-labeled large-T molecules, since
are coded for by se- the major proteolysis product is a 17K
N-termi-)m the small-t coding nal fragment which hasbeenshownbysequence
analysistoresultfrom the cleavage oflarge-Tat
celllines were labeled a lysine-rich region ending at amino acid 131
J afterimmunoprecipi- (30). Thus, the presence of the authentic 17K
e products were sepa- fragment in alarge-T-related molecule indicates
e polyacrylamide gels. that the protein is normalforatleast thefirst131
from the gels, oxidized amino acids. Since the sequences coding for
-sted withtrypsin, fin- these amino acids span the splice joint in the
)resis at pH 2.1, and large-T mRNA (30), the presence of the
frag-(33). ment also impliesthat the mRNAcoding forthe
fingerprints of normal proteinhas been spliced normally.
ther with those of the Figure 3 shows the partialproteolysisanalysis
,.Both proteins contain of the 33K and 22Kpolypeptides. In both cases
peptide (N-acetyl-Met- a major
32P-labeled
fragment which comigratedJ. VIROL.
.111.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.496.52.243.72.282.2]SV40 TRUNCATED LARGE-T ANTIGENS 57
A
B
1
*^
I
D
C
[image:4.496.58.448.74.460.2]T
FIG. 2. Comparisonofmethionine tryptic peptidemapsoflarge-T and small-t withthose of 22K and 45K
polypeptides. Peptideswereseparated byelectrophoresisatpH2.1andascendingchromatographyin
butanol-aceticacid-water-pyridine. (A) Large-TfromBALB/c 3T3CIM; (B)45KfromBALB/c 3T3C120;(C)22Kfrom
MES2006;(D)small-tfrom BALB/c 3T3 CIM. Thearrowsindicate the amino-terminalpeptide
(N-acetyl-Met-Asp-Lys), and BindicatespeptideB.
with the 17K peptide from normal large-T was
produced. This resultimplies that although we
did not detect any methionine-labeled tryptic peptides unique to large-T in the fingerprint of the 22K protein, it is nevertheless a truncated
large-T moleculeand it is normal at leastuntil
amino acid131.Theresultalso indicatesthat the
mRNAfor the 33K protein is spliced normally
and that the protein is a contiguous, truncated large-T molecule.
The15Kprotein was notanalyzedbytryptic fingerprinting, since wecouldnotprepare suffi-cient methionine-labeled material. It could not be analyzedby partial proteolysis because it is
smaller thanthe 17Kfragment. Toinvestigateits
structureweusedmonoclonal antibodies to
im-munoprecipitate the protein.
Immunoprecipitationwith monoclonal
antibod-ies. A number of monoclonal antibodies that
reactwith SV40large-T have beenisolated,and
the regions of the molecule with which they
interact havebeenmapped (11).Wehave used a
series ofsuchmonoclonal antibodies isolatedby Harlow et al. (11) to immunoprecipitate 32p_ labeledextractsof the transformedcells.
Figure4Ais anexampleof the immunoprecip-itationof the15Kpolypeptide. Antibodiesto the
amino-terminal region of large-T recognize the
VOL.44, 1982
4w
0. i
on November 10, 2019 by guest
http://jvi.asm.org/
2 3 4 5
L -lrge T-_
33.R----m
-A*
S
M
94
*-3
*-53
0 ---0
.A4,
7K
fragmrlet
:0S
-c--21 [image:5.496.108.394.74.305.2]cm --*- 14
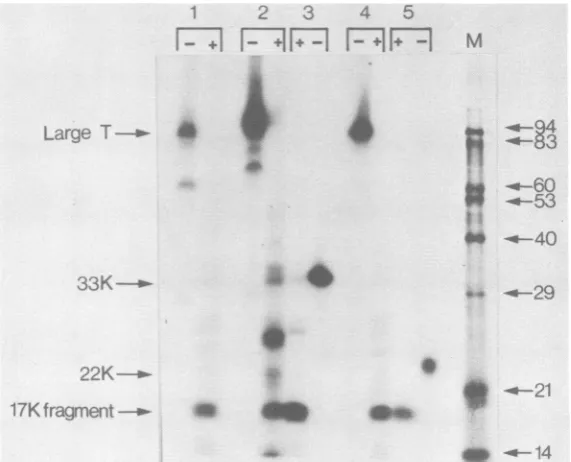
FIG. 3. Comparison of the partialproteolysisproducts of large-T, 33K, and 22K polypeptides. 32P-labeled
extractsoftransformed cells wereimmunoprecipitatedandseparated on15% polyacrylamide gels. The proteins
wereelutedfrom the gel,re-immunoprecipitated, and digested with trypsin. The fragments generated (+) were
analyzed alongsideuntreated material (-) on a 15% polyacrylamide gel. Lanes: 1, Large-T from SV40-infected
CV1 cells;2,large-Tfrom SWSV3T3;3, 33Kfrom SWSV3T3;4,large-T from MES 2006; 5, 22K from MES
2006; M, molecularweightsize markers.
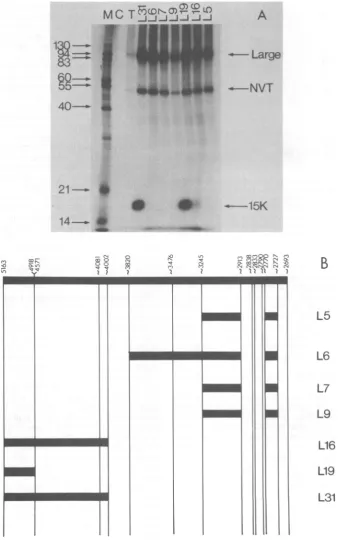
15K protein, but those against determinants mappingin the carboxy-terminalend oflarge-T (seeFig. 4B) do not. Experiments onthe other proteins yielded similar results; L19 interacted strongly with theproteins, L16 interacted weak-ly,and all other monoclonals tested(L5,L6, L7, L9)were negative.
Themonoclonal data, therefore, provide evi-dence that the 15K protein contains sequences
relatedtothe amino terminus of SV40large-T.
In vitro translation. Toestablish whether the
truncatedlarge-Tantigenswereprimary transla-tionproducts ordegradation products of other forms oflarge-T, we studied their synthesis in
vitro. mRNA was isolatedfrom each of the cell
lines and partially purified by chromatography
onSV40 DNA-cellulose(23). Figure5shows the proteins immunoprecipitated from a nuclease-treated rabbit reticulocyte lysate programmed withthepartially purifiedmRNAsfromBALB/c 3T3C120andClM.Proteins withmobility
identi-cal tothatof the45Kand15Kpolypeptideswere
detected, indicating that both are synthesized directly from virus-codedmRNA. The 33Kand
22Kproteins werealso detectedinthe cell-free translation products ofpartially purified mRNA
fromtheirrespective cell lines(datanotshown).
ThemRNAscodingforthepolypeptideswere
fractionated on formamide-containing sucrose
gradients, and samples from the different
frac-tions were translated in vitro.Figure 6 shows the
gradient analysis of thepartially purified mRNA
from MES 2006 cells. The mRNA coding for the 22K polypeptide sedimented at approximately 11 to12S, whereas the mRNA for normal
large-T sedimented at about 18S. Similar analysis of the mRNAs forthe45K, 33K,and 15K
polypep-tides showed that they sedimented in therange
11 to 16S. This result indicated that the four
truncated large-T antigens studied here are all synthesized fromtruncated mRNAs.
Predicted structure of the truncated large-T
molecules. Taken together, the data presented
hereallowusto deduce atentative structurefor the 15K, 22K,33K, and45Kpolypeptides (sum-marized inFig. 7). There isstrongevidence that theproteins consist of the normal amino-termi-nal region of SV40 large-T, and in three cases
there isgood evidence thatthe slicejunctionis
normal.
The methioninetrypticfingerprintofthe 45K
truncatedlarge-T containspeptideB.This
pep-tide isduplicatedin the 115Kform(16, 17)and
the 145Kform (18; Lovettet al., submitted for
publication) of super-T. Since the duplicated region ofthese molecules has been sequenced,
we can deduce that peptide B corresponds to
one of the two predicted methionine tryptic
J. VIROL.
FS29
on November 10, 2019 by guest
http://jvi.asm.org/
SV40 TRUNCATED LARGE-T ANTIGENS 59
1I
40-40
---O----MC
!~
..e
21-14-.o
0
-14
z-00
2oOo Iq
I ?
0 x.
T
I5i'
t.
e^:
--':M
0
co en00Nn
;0 1 1:2 1-XCbrsN N ;>
v .N > CODNN
N INNN No-
-I i I2 I1I 2I 2
- I I - I I 11PI II
-i-Il
-I-I
[image:6.496.87.429.73.614.2]- I
FIG. 4. Immunoprecipitation of 32P-labeledextractof BALB/c3T3 CIM cells. Extracts(50 jil)weremixed
with approximately 20 ±II of tissue culture supernatant from cultures of secreting hybridoma cells (11).
Immunoprecipitateswerecollected,and thepolypeptideswereseparatedon a15%polyacrylamide gel.(A) M,
Molecularweightsizemarker;C, normal hamsterserum;T,hamster anti-Tserum.Theremaining sampleswere
immunoprecipitatedwith the Lseries of monoclonal antibodiesasindicated.(B)Location of the determinants
recognized bythe monoclonal antibodies(takenfrom reference11).
A
Large
r'
u0
B
L5
L6
Li
L9
L16
L19
L31
VOL.44, 1982
on November 10, 2019 by guest
http://jvi.asm.org/
60 AND
130--
40-1 2 3
M
FCTIrCT
CTIf.w
'Nb*
21-
-FIG. 5. Cell-free synthesis of T-ai
tidesfrom different cell lines. The in vi
products made inresponseto(1) mRN
CV1cells, (2) purified mRNA from BW
cells, or (3) purified mRNA from BA
cells were immunoprecipitated with r
serum(C) and anti-Tserum(T). M, M
size markers.
peptides, which end at amino acii
Thissuggests thatthe45K species
the unique region of large-T at h
amino acid 281. This has been
recent sequencing data on the int
template for the 45K protein. TI
quencehas been shown toextend 3713 (correspondingtoaminoacid
pointthere isapresumedvirus-ho
termination codon isreached after
tional codons (C. E. Clayton, D.
Lovett,and P. W. J.Rigby, Nature press).
The fingerprint data indicate I protein is atruncated form ofthe
since they share at least two la methionine tryptic peptides. Alth( notidentified each ofthe labeled I ent in the fingerprints, tocode fo
methionine tryptic peptides woulc
the 33Kspecies includesequences asamino acid236.
The partial proteolysis data pr( 22K polypeptide must extend t(
cleavage siteatamino acid131, bi
no large-T unique methionine pel
not extend as far as the predi
endingatamino acid 178.
Thereisnodirect evidencethatI encodingthe 15Kproteinextend
siteinto theunique portionoflarg
tentative datasupporting this inte the immunoprecipitation of the p
L16 and L31 monoclonals, which
sites that may map in this region (11), and the findingthattheprotein isphosphorylated.
Phosphoaminoacid analysis of the 17K frag--
--Super
T mentgenerated by partial proteolysis
shows that~-LargeT it contains predominantly phosphoserine with
lesser amounts of phosphothreonine (data not
shown).This is in agreementwith the results of Schwyzer et al. (30) and allows the tentative 45K mapping of at least one phosphorylation site in SV40large-T. Small-t isnotphosphorylated,nor
is an 8K truncated T-antigen characterized by Spangleretal.(32). Ifweassumetherefore that the phosphorylation site is not one of the two
serine residues in the sequences shared by small-t, large-T, and the 8K protein but that it is
Sm
al5
t presentin thelarge-T unique
region,aphosphor-ylationsitecanbemapped tentativelytoone or
ntigen polypep- moreof thefive serine residues between amino itro-synthesized acids 106 and 123 in
SV40
large-T.Afrominfected If the 15K truncated large-T species is phos-kLB/c 3T3 C120 phorylated at the normal site, we can predict iLB/c 3T3 CIM that theprotein extends into the large-T unique normal hamster region atleast as far as amino acid 106. lolecularweight Since it is clear that the
15K
to 45Kpolypep-tidesform afamily of truncated forms of large-T, we used them to study properties associated id 281 or 364. with large-T, with the hope that this would allow sextends into the mapping of functional domains on the pro-east as far as tein.
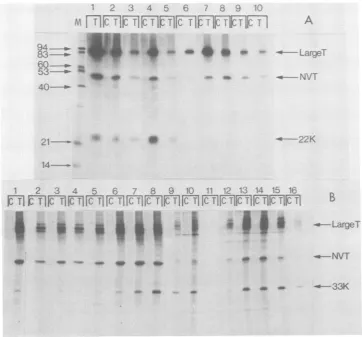
confirmed by DNA-cellulosechromatography. Large-T binds :egrated DNA to calfthymus dsDNAimmobilizedon cellulose he coding se- andcan be eluted by buffers ofincreased pH and tonucleotide ionic strength (3, 21). To test the ability of the 368), atwhich truncated forms of large-T to bind to DNA we stjunction. A subjected
32P-labeled
extracts of the trans-onlysix addi- formed cells to DNA-cellulose chromatography Murphy, M. andimmunoprecipitated the fractions obtained.D[London],in Figure 8 shows the elution profiles obtained
with the extracts from MES 2006 cells and SW that the 33K SV3T3 cells containing the 22K and 33K
trun-- 45K protein cated large-T antigens, respectively. Figure 8A
rge-T unique shows that all of the 22K proteinis present in the Dugh we have unbound fraction, whereas a large proportion of
peptides
pres- large-T is present in thefractionseluting atpHor two soluble 8.0 and pH 8.0 plus 1.0 M NaCl. Figure 8B
d require that shows that the 33K protein is present in the atleastasfar bound fraction and its presence parallels that of large-T. It has been argued that the different edict that the conditionsrequiredtoelute the various fractions
D the trypsin of large-T from
dsDNA-celiulose
reflect theut since ithas presence of more than one population oflarge-T ptides it may molecules andthateachpopulationhas a differ-cted peptide, ent affinity for dsDNA (3, 21). It is noticeable that the 33Kpolypeptidehas thesamecomplex thesequences elution profile as large-T.
past
thesplice Similaranalysisof the 15K and 45K truncated;e-T.
Theonly large-T proteins showed that the 15K protein rpretation are does not bind, whereas the 45K protein does. rotein by the These data therefore suggest that 33K truncated have binding large-T contains sequences that are necessaryJ. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.496.52.244.69.245.2]SV40 TRUNCATED LARGE-T ANTIGENS 61
a) -J
[
Lo~
6
cnc
LO
(iT,
CD
[
00~
v-~
-..I...'
*,::4 f_.'
# 4 ...*.'A,
*.
.1
t;&
.
.1
t
t tt
O tCO C)OcO
tt t OC)
Q( O nT C)
IN,1
'0 r.4)
C\M
~~~c 0
. Z
. E
C=
Z °d
co
4)4;._
'5 .~
tA
4.) cU
4.=
O.
_
... <
4. v Z
'0 &
cd;o 18D
'0 .c3CO
'0
a o.
00CU Z)
0.2 o H
CM~~~~~~C
O 2 -aYE
0
cm~~~~~~~c
VOL.44,1982
on November 10, 2019 by guest
http://jvi.asm.org/
62 CHAUDRY, HARVEY, AND SMITH
5 X100 200 300 400 500 690 7Q0
Large T
f
15K
m<>R
106
22K
A131
33K
236
45K
-_
FIG. 7. Structures deduced for the truncated large-T antigens. The shaded areas indicate regions where there is evidence that the amino acid sequences are coded for by SV40 DNA; the unshaded areas indicate regions for
which there is no direct evidence. The scale is amino acid number in normal large-T; othernumbers are referred
to in thetext.
4 6 b)
A,
FIIFT7IKIIFT
1K7IKVF7F7m
A83-
aw7- a4 e e al^--Large-:94
--_-
53--40--.-i
_~- h421----'-
1~~
* £9
1 2 3 4 5 6 A 8 9 1Q 1 1 1 1K3 14 1to V16
F~~~~~~~.-TT.r
_----33K
FIG. 8. Binding ofSV40 large-T and truncated large-Tmolecules (22K and 33K) to native calf thymus
DNA-cellulose. 32P-labeledextracts were bound to and eluted from aDNA-cellulose column. Selected fractions were
immunoprecipitated with anti-T (T) and normal (C) sera and analyzed bypolyacrylamide gel electrophoresis. (A)
MES2006: M,molecular weight sizemarkers; 1, starting material at pH 8.0; 2, starting material adjusted to pH
6.0; 3and4, flow-through fractions; 5 through 7, material eluted at pH 8.0 plus 0.1 M NaCl; 8 through 11,
material elutedatpH 8.0plus 1 MNaCl.(B) SWSV3T3: 1, starting material at pH 8.0; 2 through 4, flow-through
fractions;5through 8, material eluted at pH 8.0 plus 0.1 MNaCI;9through 11, material eluted at pH 8.0 plus 0.3
MNaCI; 12through 16, material eluted at pH 8.0plus 1.0 MNaCl.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.496.105.415.76.209.2] [image:9.496.73.436.263.600.2]VOL.44, 1982
and sufficient for binding to dsDNA, whereas the closely related 22K truncated large-T lacks these sequences.
Association with NVT. It has previously been shown that the complex formed between large-T and NVTin mouse cells transformed bySV40 is stable and can be readily detected by sucrose gradientanalysis (18, 19). The bulk of NVT and the fraction oflarge-T associated with it sedi-ment between 23 and 25S. Other multimeric forms of the complex can also be detected in the 16-18Srange (19).
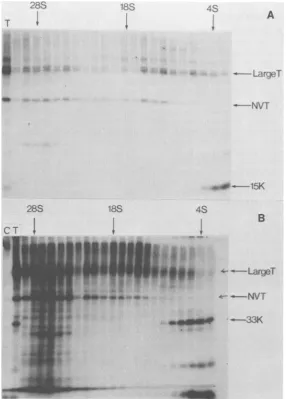
Figure 9 shows the gradient analysis of32p_
288 18S
SV40 TRUNCATED LARGE-T ANTIGENS 63
labeled extracts of the transformed cells after centrifugation and immunoprecipitation. The 15K and the 33Ktruncated large-Tantigens both remainnear the top of thegradients, whereas the large-T-NVT complexes sedimentrapidly.Both truncated large-T species sediment more slowly than the bulk of free large-T molecules. This may suggest that they are sedimenting in a monomeric form and do not associate stably with themselves, large-T, or NVT. Similar re-sults were obtained when the 22K and 45K species were analyzed on sucrose gradients (data not shown).
V-A
4 - - LargeT
"-NVT
_ 15K
18S 4S
B
4--'- LargeT
[image:10.496.107.394.220.620.2]_4 33K
FIG. 9. Sucrosegradient sedimentation oflarge-Tandtruncatedlarge-T molecules. 32P-labeledextractsof
BALB/c 3T3 CIM(A)and SWSV3T3 (B)cellswere subjectedtosucrosegradient sedimentation.A totalof 23
fractionswerecollected from eachgradientandimmunoprecipitatedwith anti-Tserumbeforepolyacrylamide gel
electrophoresis.The Svalueswereobtained from 32P-labeled rRNA markersruninaparallel gradient.Trackson
the left endof eachgel show the productsimmunoprecipitated fromanunfractionatedextractwith anti-T(T) and normal(C)sera.
28S
CT I
f.w
A4.
on November 10, 2019 by guest
http://jvi.asm.org/
An independent analysis of the possible asso-ciation between NVT and the truncated large-T
specieswas made by using monoclonal
antibod-ies directed against NVT. Although the
mono-clonal antibody only recognizes NVT, it immu-noprecipitates large-T because of its association
with NVT (13, 15, 19). However, none of the
truncated large-T molecules coprecipitated with NVT, inagreementwith the results of the
gradi-entanalysis.
Taken together, all these results indicate that
truncated large-T species extending as far as
amino acid 368 in large-T lacksequencesthatare
required for stable association with NVT.
DISCUSSION
The results presented here partially define the
structure of four species of truncated large-T. The analysis is summarized in Table 1, and the
structures deduced are presented in Fig. 7.
There is no doubt that the proteins share a
common amino-terminal amino acid sequence.
They extend tovarying degrees into the large-T
unique region. However, the data presented do
not allow us to identify precisely the carboxy
termini of the protein. Such information could
beobtained by characterizing the template DNA
for the polypeptides. This has only been done
for the 45K species (Clayton etal., in press).
No methionine tryptic peptides other than
thosepresentin normal large-Tweredetected in
thetruncated large-T species analyzed. In
addi-tion, the amount ofSV40-coded sequence that
each protein was predicted to contain
corre-sponded roughly with the apparent molecular
weights of the proteins. This suggests that the
proteins contain relatively fewnon-SV40-coded
amino acids. Indeed, DNA sequence data
pre-dict that the 45K polypeptide contains only six
host-coded amino acids (Clayton et al., in
press). Assuming that the non-SV40
carboxy-terminal sequencesof the other truncated
large-T antigens are notextensive and that the
addi-tional sequences have no deleterious effect on
the activity of the proteins, the truncated large-T
moleculescanbe used for the tentative mapping
of functional domains on large-T, as discussed
below.
Several mechanismscanbeenvisaged for the
generation of truncated large-T molecules from
integrated viral DNA, e.g., point mutations in
the viral DNA resulting in the generation of a
termination codon, rearrangements of the viral
DNA,orinterruption of the viral DNA by fusion
with host cell sequences. Examples of the last
twomechanisms have been described for
poly-oma virus (H. E. Ruley, F. Chaudry, and M.
Fried, Nucleic Acids Res., in press) and SV40
(5, 29). The mRNAs coding for the truncated
large-T species described here are all smaller
than thenormal large-T mRNA, indicating that
they probably arise by eitherrearrangement or
fusion.
DNAbinding siteon large-T. The DNA
bind-ing data obtained with the truncated large-T molecules is summarized in Table 1 and
indi-cates that the 33K polypeptide has all the
se-quences necessaryandsufficient for the binding
of SV40 large-T to dsDNA-cellulose, whereas
the 22Kspecies lacks someof thesesequences.
Studies witha33Kfragment of large-T coded for
by theSV40deletionmutantdlO001 showed that
it also bound to dsDNA-cellulose (28). Other
work with fragments ofSV40 large-T generated
by in vitro translation ofSV40 cRNA showed
that an 82K fragment containing sequences
ex-clusively from the large-T unique region also
bound to dsDNA-cellulose, but that a related
70Kfragment didnotbind(25).
Taken together, these data indicate that a
region ofSV40large-T representing the overlap
between the 33K truncated large-T
character-ized here and the 82K cRNA translationproduct
containssequencesthatarenecessaryand
[image:11.496.60.452.539.664.2]possi-bly sufficient for binding to dsDNA. If it is
TABLE 1. Structural features and biochemicalproperties ofthefour truncatedlarge-Tpolypeptides Structural features Biochemicalproperties
Size of Immunopre- Shared large- 17
Cell line truncated cipitation T and small-t Large-T d17,00 DNA binding Association T-antigen with N-ter- methionine unique haltoh (calfthymus with host
minal trypticpep- peptides
pepopdh
dsDNA) NVT(p53)monoclonal tides
antibodies
BALB/c3T3 15K + NDa ND -
-CIM
MES2006 22K + + 0 + -
-SWSV3T3 33K + + 2 + +
-BALB/c 3T3 45K + + 2 + pep- ND +
Cl20 tideB
aND, Not done.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
SV40 TRUNCATED LARGE-T ANTIGENS 65
Large
T
w22K
178
,---33K
6
dl 1001
(33K)
82K
70K
272
109
186
dsDNAbinding regin
(serine
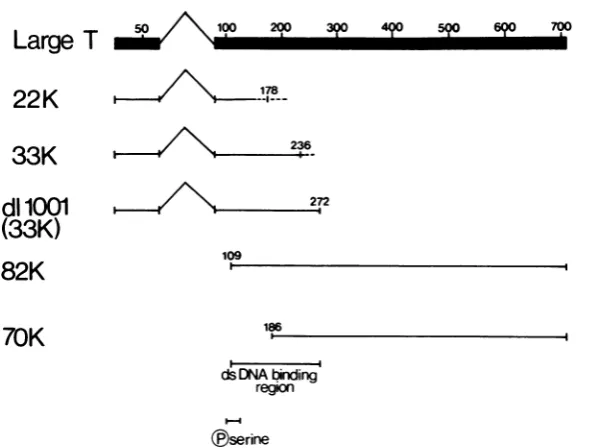
FIG. 10. A tentative mapof aphosphorylation site and a DNAbinding regionon SV40large-T, predicted
fromthe presenteddata.
assumed that the 82K protein results from the initiation ofprotein synthesis at the first AUG codon in the unique portion oflarge-T (21, 23) (corresponding to amino acid 109 in the whole molecule) and that the 33K truncated large-T extends no further than the dllO01 coding
se-quence(i.e.,toamino acid 272), thebinding site
canbe definedasbeing between amino acids109 and272(Fig. 10).
It must be emphasized, however, that this
resultrelatesonlytothebinding of SV40 large-T
to dsDNA-cellulose in vitro. No attempt has
beenmade to measureaccurately theaffinityof
suchbinding, noritsspecificity. Itisquitelikely that these activities would require additional large-T sequences.
Indeed,
recent results char-acterizing large-Trevertantsthat haveanaltered specificity ofbinding to the SV40 origin show that the mutations map within the sequencesbetween nucleotides 3459 and 3985, which are
outside the site defined here (31).
The influence of the secondary structure of large-T and the association between large-T and
NVT orbetween large-T molecules themselves
has also not been investigated in the studies
reported here. Such interactionsmay also influ-ence the affinity and specificity of binding.
In-deed, arecentreportsuggests thatlarge-Tmust
aggregate before it is able to bindto the origin
region ofSV40 DNA (2). We believe,however,
thatthe truncatedlarge-T moleculesthatbind to
dsDNA(thatis, the 45Kand 33Kspecies) bind
in their own right and not as a result of an
association with full-size large-T, because the
gradient analysis indicated that there was little detectableinteraction between thetwo.In addi-tion, several monoclonal antibodies that effi-ciently immunoprecipitated full-size large-T via carboxy-terminal determinants failed to coprecipitate the truncated large-T molecules.
NVTbinding site on large-T.Sincenoneof the
truncatedlarge-Tspecies studied here interacted stably withNVT it is notpossible todefine the region on large-T required for complex forma-tion. The results do show that the
amino-termi-nal sequences as faras amino acid 368 are not
sufficient for binding, implying that some
se-quencesfrom thecarboxy-terminalend mustbe
required. Since thelarge-T species coded for by the SV40 deletion mutants d11263 and d1265 form a complex with NVT, the extreme
car-boxy-terminal end of large-T wouldappear tobe
unnecessary(8). The results presented here
pre-dict that thesequencesbetweenamino acids 387
and662playanessential roleincomplex forma-tion with NVT. However, a more extensive study is necessary to determine whether all of
thisregion isrequired and whether it issufficient
fortheassociation with NVT.
ACKNOWLEDGMENTS
We thank Ed Harlow for the kind gift of monoclonal
antibodies; Peter Rigby forallowing us to see manuscripts
beforepublication;BenOostra, DanKalderon,andGraham Belsham for critical reading of the manuscript; and Lydia
Pearsonfor secretarial assistance.
F.C. is supported by an Imperial Cancer Rsearch Fund Bursarship.
4?0 590 690 700
mob
VOL.44, 1982
on November 10, 2019 by guest
http://jvi.asm.org/
[image:12.496.104.402.69.293.2]LITERATURE CITED
1. Alberts, B. M., F. J. Anodio, M.Jenkins, E. D. Gutmann, and F. L.Ferris. 1968. Studies with DNA-cellulose chro-matography. DNA binding proteins from E. coli. Cold Spring Harbor Symp. Quant. Biol. 33:289-305. 2. Bradley, M. K., J. D. Griffin, and D. M. Livingston. 1982.
Relationship of oligomerization to enzymatic and DNA binding properties of the SV40 large-T antigen. Cell 28:125-134.
3. Carroll, R. B., L. Hager, and R. Dulbecco. 1974. Simian virus T-antigen binds to DNA. Proc. Natl. Acad. Sci. U.S.A. 71:3754-3758.
4. Chang, C., D. T. Simmons, M. A. Martin, and P. T. Mora. 1979. Identification and partial characterization of new antigens from simian virus 40-transformed mouse cells. J. Virol. 31:463-471.
5. Clayton, C. E., and P. W. J. Rigby. 1981. Cloningand characterization of the integrated viral DNA from three lines of SV40 transformed mouse cells. Cell 25:547-556. 6.Cleveland, D. W., S. G. Fischer, M. W. Kirschner, and
U. K.Laemmli.1977.Peptide mapping by limited proteol-ysis in sodium dodecyl sulphate and analproteol-ysis by gel electrophoresis. J. Biol. Chem. 252:1102-1106. 7. Crawford, L. V., C. N. Cole, A. E. Smith, E. Paucha, P.
Tegtmeyer, K. Rundell, and P. Berg. 1978. Organization andexpression of early genes of SV40. Proc. Natl. Acad. Sci.U.S.A.75:117-121.
8. Denhardt, D. T., and L. V.Crawford. 1980. Simian virus 40T-antigen: identification oftryptic peptides in the C-terminal region and definition ofthe reading frame. J. Virol.34:315-329.
9.Griffin,J. D., G. J.Spangler,and D. M.Livingston. 1979. Protein kinase activity associated with SV40 T-antigen. Proc. Natl. Acad.Sci. U.S.A. 76:2610-2615.
10. Hand, R. 1981. Functions of T-antigens of SV40 and
polyomavirus. Biochim.Biophys. Acta 651:1-24. 11. Harlow, E., L. V. Crawford, D. C. Pim, and N. M.
Williamson.1981.Monoclonal antibodiesspecificfor sim-ian virus 40 tumorantigens. J. Virol. 39:861-869. 12. Kress, M., E. May, R. Cassingena, and P. May. 1979.
Simian virus 40-transformed cells express newspecies of proteinprecipitable by anti-simian virus40T-serum. J.
Virol.31:472-483.
13. Lane, D. P., and L.V.Crawford. 1979.T-antigenis bound to a host protein in SV40 transformed cells. Nature (London) 278:261-263.
14. Lane, D. P., and A. K. Robbins. 1978.Production,
proper-tiesand specificityofarabbitantibodytopurifiedSV40
large-T antigen. Virology87:182-193.
15. Linzer, D. H., and A. J.Levine.1979.Characterization of a 54K cellular SV40 tumor antigen present in SV40 transformed cells and uninfected embryonal carcinoma cells. Cell 17:43-52.
16. May, E., J.-M. Jeltsch,and F.Gannon.1981. Character-ization of agene encoding a 115K super-T antigen ex-pressed by an SV40 transformed rat cellline. Nucleic Acids Res. 9:4111-4128.
17. May, E.,M.Kress,L.Daya-Grosjean,R.Monier,andP. May. 1981.Mapping of the viralmRNAencodinga super-Tantigenof115,000 daltonsexpressedin simian virus 40-transformedratcell lines.J.Virol. 37:24-35.
18. McCormick, F., F. Chaudry, R. Harvey, R. Smith, P. W.J. Rigby, E.Paucha, and A. E. Smith. 1980. T-antigens of SV40 transformed cells. ColdSpringHarbor
Symp. Quant.Biol. 44:171-178.
19. McCormick, F.,and E. Harlow. 1980.Association ofa
murine53,000-daltonphosphoproteinwith simianvirus 40 large-Tantigen in transformed cells. J. Virol. 34:213-224. 20. Myers, R. M., D. C. Rio, A. K. Robbins,andR. Tjian. 1981. SV40 gene expression is modulated by the co-operative binding ofT-antigentoDNA.Cell 25:373-384. 21. Oren, M., E. Winocour,andC.Prives.1980. Differential affinities ofSV40 largeT-antigenfor DNA. Proc. Natl. Acad. Sci. U.S.A. 77:220-224.
22. Paucha, E., R. Harvey, and A. E. Smith. 1978. Cell-free synthesis of simian virus 40T-antigens.J. Virol. 28:154-170.
23. Paucha, E., and A. E. Smith. 1978. The sequences be-tween0.59 and 0.54 map units onSV40 DNA code for the unique region of small-t antigen. Cell 15:1011-1020. 24. Pelham, H. R. B., and R. J. Jackson. 1976. An efficient
mRNA dependent translation system from reticulocyte
lysates.Eur. J.Biochem. 67:247-256.
25. Prives, C., Y. Beck, and H. Shure. 1980. DNA binding properties ofsimian virus 40 T-antigens synthesized in vivo and in vitro. J. Virol.33:689-696.
26. Prives, C., E.Gilboa, M. Revel, and E. Winocour. 1977. Cell-free translation of SV40early mRNA coding for viral T-antigen. Proc. Natl. Acad. Sci. U.S.A. 74:457-461. 27. Rassoulzadegan, M., B. Perbal, and F. Cuzin. 1978.
Growth control insimian virus 40-transformed rat cells: temperature-independent expression of the transformed phenotype in tsA transformants derived by agar selection. J.Virol. 28:1-5.
28. Rundell, K., J. K. Collins, P. Tegtmeyer, H. L. Ozer, C. J. Lai, and D. Nathans. 1977. Identification of simian virus 40protein A. J. Virol. 21:636-646.
29. Sager, R., A. Anisowicz, and N. Howell. 1981. Genomic
rearrangementsina mouse cellline containing integrated SV40 DNA. Cell 23:41-51.
30.Schwyzer, M., R. Weil,G. Frank, and H. Zuber. 1980. Aminoacid sequence analysis of fragments generated by partial proteolysis from large SV40 tumor antigen. J. Biol. Chem. 255:5627-5634.
31.Shortle, D. R., R. F. Margoiskee, and D. Nathans. 1979. Mutational analysis of the SV40 replicon: pseudorever-tantsof mutants with adefective replication origin. Proc. Natl. Acad. Sci. U.S.A.76:6128-6131.
32. Spangler, G. J., J. D. Griffin, H. Rubin, and D. M. Livingston. 1980. Identification andcharacterization of a new low-molecular-weight virus-encoded T-antigen in a line of simian virus40-transformedcells. J. Virol. 36:488-498.
33. Smith,A.E.,R.Snith,and E. Paucha.1978. Extraction andfingerprint analysis of simian virus 40 large and small
T-antigens.J.Virol.28:140-153.
34. Smith,A.E.,R.Smith, and E. Paucha. 1979. Character-ization ofdifferenttumorantigens present in cells trans-formedbySV40. Cell 18:335-346.
35. Tjian,R.1978. Thebindingsite on SV40 DNA fora T-antigen related protein. Cell 13:165-179.
36. Tjian,R.1981.Regulation of viraltranscriptionandDNA
replication by SV40 large-T antigen. Curr. Top. Micro-biol.Immunol.93:5-24.
37. Tjian, R., and A. Robbins. 1979. Enzymatic activities associated withapurified SV40 T-antigen relatedprotein. Proc.Natl. Acad.Sci. U.S.A. 76:610-614.
38. Tooze, J. 1981. Molecularbiology oftumorviruses,part2: DNAtumorviruses, 2nd ed. ColdSpring Harbor Labora-tory, ColdSpring Harbor,N.Y.
39. Volckaert,G., J. Feunteun,L. V.Crawford,P.Berg,and W.Fiers.1979. Nucleotidesequence deletions within the coding region for small-t antigenof simian virus 40. J. Virol. 30:674-682.
on November 10, 2019 by guest
http://jvi.asm.org/