JOURNALOF VIROLOGY, May 1977, p. 402-411
Copyright ©D 1977 AmericanSocietyforMicrobiology
Vol.22,No. 2
Printed in U.S.A.
Adenovirus Type
2
Coded Single-Stranded DNA Binding
Protein:
In
Vivo
Phosphorylation and Modification
YUN-HUA JENG,* WILLIAM S. M. WOLD, KENJI SUGAWARA,' ZVEE GILEAD, AND
MAURICE GREEN
InstituteforMolecular Virology, Saint Louis UniversitySchoolofMedicine, St. Louis, Missouri 63110
Received for publication 30 November 1976
The adenovirustype2-coded single-strandedDNAbinding protein (DBP) was shownto be aphosphoprotein and to exist in at least two forms that differ in mobility by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. After a 30-min pulse with [35S]methionine or 32PO4, 35S- or 32P-labeled DBP had a nominal molecularweightof 74,000, whereas aftera30-minlabel followed bya 20-h chase, 35S- and 32P-labeled DBP hadanominalmolecular weight of77,000. Both large and small forms of 35S-and32P-labeledDBPboundtosingle-stranded
DNA-cellulose columns andwereeluted by0.4 to 0.6 MNaCl; both forms also wereimmunoprecipitated byantiserumagainst adenovirustype 1-simian virus 40-inducedtumorcells(thisantiserumcontainsantibodiesagainstDBP) andby
monospecific antiserum against95 to 99% purified DBP. Withhighlypurified
32P-DBPlabeled7 to 10 hpostinfection, it wasshownthat the32p radioactivity was firmly associated with protein material (i.e., not contaminating nucleic acids or phospholipids) and had properties expected of a phosphoester of an aminoacid;paperelectrophoresisof acidhydrolysatesof thispreparation identi-fied phosphoserine but notphosphothreonine. Phosphoserine but not phospho-threonine was also identified in acid hydrolysates of another preparation of 32P-DBP labeled for30 min, chased for20 h, and then immunoprecipitated by adenovirus type 1-simianvirus 40 antiserum.
Human adenovirus 2 (Ad2) is an oncogenic DNA virus that replicates in the nucleus of permissive human cells (reviewedinreference 18 and in W. S. M. Wold, M. Green, and W. Buttner,In D. P.Nayak, ed.,Molecular Biology
ofAnimal Viruses, in press). The productive infection proceedsin an"early"stage,when23 to 32%of the asymmetricgenome isexpressed asmRNA, andin a"late"stage,which follows the initiation of viral DNA replicationat 6 to 7 h postinfection (p.i.). Virus-coded or -induced proteinssynthesized earlyafter infection
prob-ably are involved in viral DNA replication, regulation of viral gene expression, and trans-formation ofnonpermissive cells. A protein of 75,000 (75K) daltons, which binds to single-stranded DNAbutnotto native DNA, is pro-ducedinlarge quantities in early stages of Ad2 infection(24, 27). Ananalogous Ad5DNA bind-ing protein (DBP) of 72K was isolated from abortively infectedmonkey cells (28). Recently anAdl2DBPof 58K to 60K has beendescribed (29). Ad2 and Ad5 DBPs of 41K to 45K are also
IPresentaddress: Department ofMicrobiology,Faculty ofMedicine, Kyoto University,Konoe-cho,Yoshida, Sakyo-hu,Kyoto, Japan606.
synthesized, butpeptide analyses (20), genetic
studies (29), and immunoprecipitation studies (27a) indicate that these are derived from the larger DBP (possibly degradation products). Ad2 DBP has been showntobe viral codedby cell-free translation of hybridization-purified Ad2 earlymRNA into a 72Kpolypeptidewith a peptide map similar to that of DBP isolated from infectedcells, and theDBPgenehasbeen
mapped in Ad2 EcoRI-B fragment (Ad2 map position, 0.59 to 0.71) (16). Recently, Ad2 DBP has been purified to 95 to 99% homogeneity; sucrosegradientandgel filtration studies indi-cate that it does not have a typical globular protein structure (27).
DBPisprobably involved in viral DNA repli-cation, since itis found in both nuclear mem-brane and soluble Ad2 DNA replication com-plexes (24, 33, 34; M. Arens, T. Yamashita, R. Padmanabhan, T. Tsuruo, and M. Green, man-uscript in preparation). Genetic studies indi-cate that DBP functions in the initiation of viral DNAreplication (30). DBP issynthesized inlargequantities, which suggests that its role is stoichiometric rather thancatalytic (28), as inthecase of T4gene 32protein (1). Many Ad2-402
on November 10, 2019 by guest
http://jvi.asm.org/
PHOSPHORYLATED Ad2 DNA BINDING PROTEIN 403
transformed cells neither containtheDBPgene (23) nor synthesize DBP (15, 27a; P. H. Galli-more, personal communication), indicating that DBP does not function to maintain cell transformation.
Many cellular processes are controlled by modifications, such as adenylation (10), meth-ylation (4), and phosphorylation (22), of key regulatory proteins. We, therefore, investi-gatedwhether Ad2 DBPundergoes posttransla-tional modification. Inthiscommunication, we demonstrate that Ad2 DBP is phosphorylated atserineresidues, and that at least two forms of DBPexist that differ inmobility by sodium dodecyl sulfate (SDS)-polyacrylamide gel elec-trophoresis.
MATERIALS AND METHODS
Chemicals. [35S]methionine (Met) (400Ci/mmol),
[3H]leucine (40Ci/mmol), and inorganic32P04 (car-rier-free) were obtained from New England Nuclear Corp.;
1-,f-D-arabinofuranosylcytosine
(ara-C),phos-phoserine, phosphothreonine, pancreatic DNase
(EC 3.1.4.5), and Pronase were secured from Sigma Chemical Co.; acrylamide came from
East-man Kodak Co.; N,N-methylenebisacrylamide,
N,N,N',N'-tetramethyladenediamine (TEMED),
and ammoniumpersulfate were procured from
Bio-Rad Laboratories; SDS was bought from BDH
Chemical, Ltd.; pancreatic RNase A (EC 2.7.7.16)
and Escherichia coli alkaline phosphatase (EC
3.1.3.1) wereboth obtained from Worthington
Bio-chemical Corp.; and pancreatic RNase T, was
ac-quiredfromCalbiochem.
Preparation of labeled protein extracts. Human
KBcells (clonal line) were grown at370CinEagle
minimal essential medium (MEM) containing 5%
horse serum. Cells were infected at 6 x 106cells/ml
with Ad2 (strain 38-2) at 100 PFU/cell in MEM
without horse serum. The purification of Ad2 has
beendescribed (9). After1hofadsorption,cells were
suspendedat 3.5 x 105cells/mlinMet-free or phos-phate-free MEM with 5% horse serum. At 4 h p.i.,
ara-Cwasadded to aconcentrationof 20
gg/ml.
At0.5 h priorto labeling, cells werecentrifuged and
resuspended at 6.5 x 105cells/ml in warm Met-free
orphosphate-freeMEMwith5%horseserumand20
,ugof ara-Cper ml. Cellswerepulse-labeledfrom 7
to 7.5 h p.i. with 25 ,uCi of [35S]Metor0.5 mCi of
32PO4 per ml. Chased samples were prepared by
washing cells twice in warm, complete MEM
con-taining 5% horse serumand20,.gof ara-C perml,at
the endof thelabelingperiod, andthenincubating
cellsinfresh washmedium for20h.Harvested cell
pellets were washed twice with cold
phosphate-buffered saline lackingCa2+ and Mg2+. Cellswere
thensuspendedat 107cells/mlinreticulocyte
stan-dardbuffer (10 mM Tris-hydrochloride[pH 7.4], 10
mMNaCl, 1.5mMMgC12), placedinanicebath for
15min, andsonicallydisrupted. Cellextracts were
digestedwith 25 Mugof DNaseperml at37°Cfor1h
in 5 mM MgCl2. EDTA wasadded to 10mM, and
extractswere furtherdigested with 20 ,ug each of
RNase A andT1 permlat370C for 30 min.Extracts
were thendialyzed for 16 h at40C against 10 mM Tris-hydrochloride (pH 7.5) and 1 mM 2-mercapto-ethanol, and the samples wereeither analyzed di-rectlyorstoredat -70°C. Mock-infected protein
ex-tracts were similarly prepared, except that virus
wasnotadded.
Analytical and preparative SDS-polyacrylamide gel electrophoresis. SDS-polyacrylamide gel experi-ments were prepared as described (11, 14, 26).
Gra-dientgels of8to 21%polyacrylamide with 0.1%SDS
were formed as slabs between glass plates; slabs
were 1.5 mm thick and 100 or 270 mm long. A
stacking gel of 5% polyacrylamide and 0.13% bis-acrylamide (analytical gels contained up to 20 sam-ple wells) was cast above the separating gel.
Elec-trophoresis was performed at 7°C with a constant
currentof20mA. After electrophoresis, analytical
gelsweredried under vacuum and autoradiographed
with Kodak single-coated panoramic dental X-ray
film (DF-85). Apparent molecular weights of
pro-teinswereestimatedby comparison of their
migra-tion rate in SDS-polyacrylamide gels with known
molecular weight standards. Cylindrical gels (0.6 by
10cm) containing 6%polyacrylamide were
electro-phoresed at 2 mA/gel for 3 h.Gels were fractionated
into 2-mm slices with aGilson gelfractionator and
counted in 5 ml ofAquasol in a liquid scintillation
counter.
To localize protein bands in preparative gels, a
lightly loaded analytical gel with labeled protein
markers was run in parallel with each batch of
preparative gels. The gels were kept wet during
autoradiography, and thelocalized phosphoprotein
bands were cut out and elutedfrom gels with
agita-tion at 37°C in 10volumes of 5mMNaHCO3
contain-ing 0.05% SDS. After 16 h, acrylamide fragments
wereremoved by centrifugation, and the
superna-tant was dried. A small portion ofprotein from
preparative gels was checked forpurity by
analyti-calSDS-polyacrylamide gelelectrophoresis.
Immunoprecipitation of DBP from crude
ex-tracts and DNA-cellulose purified materials.
Indi-rect radioimmunoprecipitation was carried out as
described (7, 8) with minormodifications.
Immuno-globulin G (IgG)wasisolated fromanantiserum (a
gift from R. Gilden) against extracts of Adl-simian
virus 40(SV40)-induced hamstertumorcells(6); this
antiserum contains antibodiesto DBP(7, 8).
Sam-ples from infected and mock-infected extractswere
mixed with 20 Mg of Adl-SV40 IgGornonimmune
IgG in 240-,ul reaction mixtures and incubated for 18
hat4°C.Then45 ul of goat serumanti-hamster IgG
wereadded and incubation continued for2hat37°C.
Immunoprecipitates were collected by
centrifuga-tion for 1.5 min at 10,000 x g in a Beckman 152
microfuge, washed threetimeswith150M1 of
phos-phate-buffered saline containing0.5 M urea, 0.5%
Nonidet P-40, and1%deoxycholate, suspendedinan
equal volume of gel electrophoresis sample buffer
(11), and electrophoresed on SDS-polyacrylamide
gels.
Someimmunoprecipitations wereconductedwith
guineapigIgG against95to 99%purifiedDBP
(anti-DBP) (27a); these utilized a modified
radioimmuno-precipitation buffer (10mM sodiumphosphate [pH
VOL. 22, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
404 JENG ET AL.
7.2], 150 mM NaCl, 1% Triton X-100, 1% sodium
deoxycholate, and 0.1% SDS) for both incubation
and washing (5). Portions of DBP-containing
elu-atesfrom DNA-cellulosewereadjustedto1x
radio-immunoprecipitationbuffer and mixed with 60 Ag
ofanti-DBP guineapig IgGornonimmune guinea
pigIgG; the reactionswere thenincubated for 18 h
at4VC. Then 60 pul of goat serum anti-guinea pig
IgG were added to form theimmunoprecipitates.
Purification of32P-labeled DBP. Adetailed
de-scription of the DBP purification method is
pre-sented elsewhere (27). Briefly, infected cells were
simultaneously labeled with 2 jtCi of [3H]leucine
and 0.5mCi Of32PO4per mlfrom7to10hp.i.inthe
presence ofara-C. Cytoplasmic extracts were
pre-pared and chromatographed on a single-stranded
DNA-cellulose column (24). Bound proteins were releasedfrom the columnby stepwise elution with
buffer A (20 mMTris-hydrochloride[pH 8.1],50mM
NaCl, 5 mMEDTA, 1 mM2-mercaptoethanol, and 10%glycerol)containing0.05M, 0.2 M, 0.4 M, 0.6 M,
and 2.0 MNaCl,respectively.Fractionseluted with
0.6 M NaCl were pooled, treated with 1% sodium
deoxycholate for 1 h at 40C, and dialyzed against
buffer B (10 mMTris-hydrochloride[pH7.5], 5mM
NaCl, 1 mM 2-mercaptoethanol, 0.02% Nonidet
P-40, 0.4 M urea, and 10% glycerol). The dialyzed
material wasthenappliedto aDEAE-Sephadex
col-umnequilibrated withbufferB, andproteins were
eluted withalinear gradientofbufferBand 0.3 M
NaCl inbufferB.The major elutedradioactivepeak
containing DBP was pooledand analyzed by SDS-polyacrylamidegelelectrophoresis.
Identification of phosphorylated amino acids. 32P-labeled DBP purified by DNA-cellulose and DEAE-Sephadexchromatographywasfurther puri-fied by electrophoresis in preparative SDS-poly-acrylamidegels asdescribedabove. The32P-labeled
DBPband was eluted from the gel and dried, and
thedriedresidue washydrolyzedin 1mlof 6 N HCl at 1100C for2 h. A 2-h period was selected as the hydrolysistime tomaximize hydrolysis ofthe
pep-tide bondwhilelimitinghydrolysisof the
phospho-rylated amino acid. Under theconditionsofpartial
acid hydrolysis used, about 20% of the liberated phosphoserine would be hydrolyzed (12). The hy-drolysatewasdriedanddissolved in 16pjlofwater containing 40 ,ug ofunlabeled phosphoserine and phosphothreonineeach andsubjectedtopaper elec-trophoresis (2). Electrophoresis wascarried out on
Whatman no. 1filterpaper with a formicacid-acetic
acid-water (30:90:880)mixture (pH 1.9) at 600 V for
16 h at 40C. The paper was dried, and unlabeled
phosphoamino acidmarkers developed with
ninhy-drinreagent (Sigma). Theposition ofradioactivity
wasdeterminedwith aPackardradiochromatogram
scanner.Rechromatographywasperformedby
elut-ing theradioactive spots from the paper
chromato-gram withwater andsubjectingthe driedmaterial
topaperelectrophoresisasdescribed above.
RESULTS
Pulse-chase experiments showing that
35S-and 32P-labeled DBP decrease in mobility in
SDS-polyacrylamide
gel
electrophoresis
dur-ing
chaseperiods.
Ad2-infected and mock-in-fected KB cellswere labeled with [35S]Met or32PO4
for 30min,
orfor 30 minfollowedby
a20-h chase. To prevent the transition to late
stages
ofinfection,
ara-C was added 4 hp.i.,
and cellswerelabeled in thepresenceof ara-C. Whole cell extracts were
prepared,
andpoly-peptides
wereresolvedby
SDS-polyacrylamide
gradient
slabgel
electrophoresis. Figure
1 shows anautoradiogram
ofone suchexperi-ment. After a 30-min
pulse,
35S-labeled DBPmigrated
withan apparent molecularweight
of 74K (lane
A),
but after a 20-h chase theapparent
molecularweight
of DBP had in-creasedto 77K (lane C). The35S-labeled
DBP band was notpresent
in lanesrepresenting
pulse
(lane B)- orpulse-chase
(laneD)-la-beled,
mock-infectedcellextracts.Analyses
ofproteins
labeled with32P04
indicated thatDBP,
and no other Ad2-inducedpolypeptide,
was associated with thephosphate
label (inFig.
1 cf. lane E withF,
and lane G withH).The 32P-labeled DBP behaved like35S-labeled
DBP in
pulse-chase
experiments:
32P-labeledDBP hada nominal molecular
weight
of 74Kaftera30-min
pulse
(lane E) and 77K aftera20-h chase (lane G). In
Fig.
1,
there are noapparent
virus-specific
32P-labeled
polypep-tides of 41Kto45K
daltons, i.e.,
representing
the DBP
subspecies
oftheseapparent molecu-larweights
(20).Apparently
the 32P-labeledDBP
subspecies
were present in insufficient amounts tobeobserved in theautoradiogram,
because in other experiments these
polypep-tides were shown to be
phosphorylated
(data
not
shown).
Immunoprecipitation by
Adl-SV40IgG
ofpulse-labeled
andpulse-chase-labeled
35S-and32P-DBP.
Antiseraagainst
cell extracts ofAdl-SV40-andAd2-SV40-induced hamster tu-morcells containantibodiestoAd2 DBP(7,8).
Asshownin
Fig.
2,
IgG
fromAdl-SV40antise-rum
precipitated
35S-labeled DBP from cells labeled for 30 min(lane
F),
or for 30 min followedby
a20-h chase (lane J). Theimmu-noprecipitated DBPs showed the same appar-ent molecular
weight change
as untreatedsamples
(lanes A andC,
pulse-labeled andpulse-chase-labeled
infected cells); i.e., the chased DBP hadaslightly
reducedmobilityin thegels.
In control experiments, IgG from nonimmunehamstersdidnot immunoprecipi-tate 35S-DBP from pulse-labeled (lane E) orpulse-chase-labeled
(lane I) infected cellex-tracts;
also,
no 35S-labeled polypeptides wereimmunoprecipitated
from mock-infected cell extractsby
Adl-SV40IgG
(pulse-lane
H,
chase-lane L) or normal hamster
IgG
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
PHOSPHORYLATED Ad2 DNA BINDING PROTEIN 405
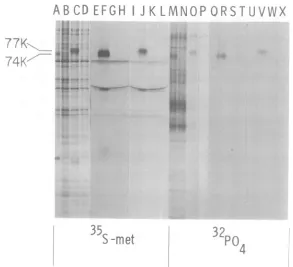
ABC
D E F
G
H
4
I.
4
77K>
s4
74KI
*Pd
Hi9*
1"n s * P;4
_¢ ¢ 9* <>
n"= z
i__
V
b__* ..*
u * * is ^
u
"g2.:
_ b ^.,-_
u__*w .w.
_
h __
-=-S0|i -
wt-~ ~ V
IMI
mlI
M I MC
35 32
S -met PG 4
FIG. 1.Autoradiogram of SDS-polyacrylamide
gel electropherogram showing two electrophoretic forms(anapparentmolecularweight of74K,aftera
30-mmn pulse,and77K,aftera30-mmn pulse followed
bya20-h chase) of35S- and 32P-labeled DBP. Ad2-infected (I) and mock-infected (M) KB cells were
pulse-labeled with [358]Metor32P04, 7 to 7.5 hp~i. (p),or7to 7.5hp.i.followed bya20-hchase in thepresenceofara-C. Total cellproteinextractswere
prepared andanalyzed by SDS-polyacrylamide gel
electrophoresis followed by autoradiography. (AB)
us;(C,D) pulse-chase; (EF)3) pulse;
(0 pulse-chase.
(pulse-lane G, chase-lane K). Adl-SV40 IgG also precipitated32P-DBP from both pulse (lane R)- and pulse-chase-labeled (lane V) in-fected cell extracts. Again, the chased 32P-la-beledDBPhadaslightlyreducedmobility. As with 35S-labeled DBPs, the 32P-labeled DBPs immunoprecipitated from pulsed and pulse-chased infected cellextractshad the same ap-parentmolecularweight as 32P-labeledDBPin samplesnotimmunoprecipitated(lanesMand 0). Controlimmunoprecipitationexperiments performedwith32P-labeledinfected and mock-infected cell extracts revealed no detectable 32P-labeledDBP (lanesQ, S,T, U, W, andX). Thus DBP-specific antisera immunoprecipi-tated74Kand 77K formsof both 35S- and32p_ labeled DBP. High-titer monospecific guinea pig antiserum against 95 to 99% purified DBP also precipitated 35S- and 32P-labeled DBP (data notshown).
Pulse-labeled and pulse-chase-labeled 32S-and 32P-DBPs that bind to single-stranded DNA-cellulose. Ad2 DBP binds to single-stranded DNA and canbe eluted from single-stranded DNA-cellulose with0.6 M NaCl (24). Totestwhether both74K and 77K forms of 35S-and32P-labeledDBPhavea similaraffinityfor DNA, infectedand mock-infected KB cellswere labeled with V35S]Metor32PO4for 30min(or for 30minfollowedby a 20-h chase), and cytoplas-micextracts were chromatographed on single-stranded DNA-cellulose columns. About 90 to 95% of the labeledproteins passed throughwith
the void volume in 50 mM NaCl. The column was washed with buffers of increasing ionic
strength(seeabove). Proteins eluted with0.6 M NaCl were electrophoresed on SDS-polyacryl-amide gels. As shown in Fig. 3, pulse-labeled 35S-DBP(lane A) and 32P-DBP (laneE), aswell as pulse-chase-labeled iS-DBP (lane C) and 32P-DBP (lane G), boundto DNA-celluloseand were eluted by0.6 MNaCl. Both pulse-chased 35S- and32P-labeled DBPs had lower mobilityon
the gel. Thus both 74K and77K forms of ",S-and 32P-labeled DBPs bind to single-stranded DNA and are eluted from the column at the sameNaCl concentration.
Purification of 32P-labeled DBP by DNA-cellulose and DEAE-Sephadex chromatogra-phy and preparative SDS-polyacrylamide gel electrophoresis. The experiments described above demonstrate that 32p labelisapparently firmly associated with DBP (i.e., notremoved by SDS and by chromatographyon DNA-cellu-lose). For further studiesonthephosphorylated component, 32P-labeled DBP was purified by sequential DNA-cellulose and DEAE-Sephadex
chromatography (which yieldsDBP of95to99% purity) (27), followed by preparative SDS-gel
VOL. 22, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.50.242.57.550.2]3EKFIH
K
LMNO,
1>
~~~~~~~~~A.a
ii~.~ ~ ~ ~ ~ ~ ~ * 4
FIG. 2. Autoradiogram ofSDS-polyacrylamide gel electropherogram showingthatAdl-SV4OIgG
immu-noprecipitatesboth 74K(seen after30-mmnpulse)and 77K(seenafter30-mmnpulse followedby20-h chase) forms of35S- and32P-labeledDBP.Ad2-infectedandmock-infectedcellswerelabeled with[35S]Met or32O (from7to7.5hp.i.,or7to 7.5hfollowedbya20-hchase),totalcellextractswereprepared,and immuno-precipitationexperimentswereperformedwithAdl-SV4Oand nonimmunehamsterIgGs.AthroughDand M throughP,respectively,show 35S- and32P-labeledextractselectrophoresed before immunoprecipitation:
pulse-labeledextractsare inA(Ad2 infected),B(mockinfected), M(Ad2infected),and N(mock infected);
pulse-chase-labeledextractsareinC(Ad2infected),D(mockinfected),0(Ad2infected),and P(mockinfected).E
throughL(35S-labeledextracts) andQthroughX(32P-labeledextracts)show the resultsofthe
immunoprecipi-tationexperiments. (EandQ)Infected pulse-labeledcellsversusnonimmunehamsterIgG;(FandR)infected
pulse-labeledcells versus Adl-SV4OIgG; (GandS) mock-infected, pulse-labeled cells versus nonimmune IgG; (Hand T) mock-infected,pulse-labeledcells versusAdl-SV4O IgG;UIand U) infectedchase-labeled
cellsversusnonimmuneIgG; (J andV)infectedchase-labeled cellsversusAdl-SV4OIgG;(KandW)
mock-infected,chase-labeled cellsversus nonimmuneIgG;and(LandX)mock-infected,chase-labeled cellsversus Adl-SV4OIgG.
electrophoresis. Infected cell proteins were
la-beled lightly with [3H]leucine and copurified with 32P-labeledproteins. Each stage of purifi-cation was monitoredby gel
electrophoresis.
A labeling time of7 to 10 h p.i. was chosen to obtainmaximal synthesis of DBP,asindicated by preliminary experiments. The cytoplasmic extract was adsorbed to a single-strandedDNA-cellulose column, and bound proteins were eluted with buffers of increasing ionic strength. 32P-labeled polypeptides eluted with each NaCl concentrationwereanalyzedbygel
electrophoresis,
and the results are shown in Fig. 4. Lanes A and B, respectively, show marker 35S-labeled polypeptides from infected and mock-infected cell extracts; these extracts werepreparedby the"cycloheximideenhance-ment" procedure to increase the levels of la-beledAd2-inducedearly polypeptides(11). Ad2-induced
polypeptides
of 74K (pulse form ofDBP), 21K, 19K,
17.5K (faint), 15K,11.5K,
11K, and 8K are apparent in this autoradi-ogram.
Figure
4shows the32P-labeled polypep-tides presentin totalcell(laneC)and cytoplas-mic (lane D)extracts, and thefollowing NaCleluates from DNA-cellulose: 0.05 M (lane
E),
0.2 M(laneF), 0.4M (lane G), 0.6M (laneH),
and 2.0 M NaCl(lane
I). 32P-labeledDBPwas elutedmainlywith 0.6 MNaCl (lane H)andto alesserextentwith0.4MNaCl (laneG). Thus32P-labeled DBP is eluted from DNA-cellulose
atthesameNaClconcentrationsas3H-labeled DBP reported previously (24, 28). 32P-labeled
DBP is notapparent inlanes C and D,
repre-406 JENG ET AL. J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.504.114.407.71.338.2]PHOSPHORYLATED Ad2 DNA BINDING PROTEIN
A
BCD E F
G
H
0
--#1
.4DEAE-Sephadex. 32P-labeled proteins co-chro-matographed with the 3H-labeled proteinsthat wereused to monitor the purification. The peak fractions were pooled and analyzed by electro-phoresis on cylindrical gels. The 3H- and 32P_ labeled DBP comigrated in the cylindrical gel
(datanotshown), suggesting the covalent asso-ciation of 32p label with purified DBP. The re-maining material was electrophoresed on a pre-parative slab gel; the band correspondingto32p_ labeled DBP was cut from the gel, and the 32p_ labeled DBP was eluted and chemically ana-lyzed as described below.
DBPcontains phosphoserine but not phos-phothreonine. The phosphate linkage proper-ties of the 32P-labeled DBP band isolated by preparative gel electrophoresis, as described
M
I
MlMi
I
Mp c p IC
[image:6.504.50.241.61.383.2]35S
-met
32P04
FIG. 3.Autoradiogram of SDS-polyacrylamide
gel electropherogram showingthatboth74K(pulse)
and 77K(chase) forms of 35S-and 32P-labeledDBP
bindtosingle-stranded DNA-cellulose columnsand
are elated by 0.6 MNaCl. Ad2-infected (I)
mock-infected (M) cellswerepulse-labeled with [35S]Metor
32PO47to 7.5 hp.i. (p),orthesameperiod followed
bya20-h chase(c). Cytoplasmic extracts were
pre-pared and chromatographed on single-stranded
DNA-cellulose columns. Bound proteinswereelated
with 0.05M,0.4M,and 0.6 MNaCl. Theproteins
elated by 0.6 M NaCl were analyzed by
SDS-poly-acrylamide gel electrophoresisandautoradiography.
(A,B) 35S pulse; (C,D), 35Spulse-chase; (E,F) 32p
pulse; (G,H) 32ppulse-chase.
senting total cell and cytoplasmic extracts (which contain DBP),becauseequal
radioactiv-ity was applied to each lane, and 32P-labeled
DBPwas enriched in the0.4 and 0.6 M NaCl
eluates from DNA-cellulose. The 74K 32P-la-beled polypeptide eluted from DNA-cellulose
was showntobe DBPby immunoprecipitation of both the 0.4 and 0.6 M NaCl eluates with monospecific IgG against purified DBP (data not shown).
The 0.6 M NaCl eluate from DNA-cellulose was further purified by chromatography on
74K
21
K
19K
17. 5K
. t
15K
11.5K
K
1K
-I
8K-A B
C
D
E
F G
H
I
ii
* * 0
_T.
_., I
S
[image:6.504.255.449.256.485.2]I M
FIG. 4. Autoradiogram of SDS-polyacrylamide
gel electropherogram showing the 32P-labeled
poly-peptides elated from a single-stranded
DNA-cellu-lose column by various NaCl concentrations.
Ad2-infectedcellswerelabeled with32PO47 to 10 hp.i.,
and thecytoplasmicextractwaschromatographedon
asingle-strandedDNA-cellulose column. The bound
proteinswereelated withincreasingNaCl
concentra-tions.(A)Ad2early infectedKB cells extracts labeled
with[35S]Met 7to8 hp.i. in thepresenceofara-C
after cycloheximidepretreatment(i.e.,Ad2early
pro-teinmarkers); (B) mock-infected,35S-labeled cell
ex-tractstreated thesame asinfected cells. (C through I)
show 32P-labeled polypeptides as follows: (C) total
cell extracts; (D) cytoplasmic extract before
DNA-cellulose; (E)DNA-celluloseflow-through and0.05
MNaCl wash; (F) 0.2 M NaCl eluate; (G) 0.4 M
NaCI eluate;(H) 0.6MNaCleluate;and (I)2.0M
NaCleluate.
77K
74K
lt
VOL. 22, 1977 407
d,.>
42
I
I
d
w .
on November 10, 2019 by guest
http://jvi.asm.org/
408 JENG ET AL.
above, were analyzed. As shown in Table 1,
treatment with hot trichloroacetic acid or
or-ganicsolvents didnotsolubilize significant 32p counts,suggesting that the 32p labelwas
associ-ated with protein andnotwith RNAorDNAor
phospholipidscontaminatingtheDBP
prepara-tion. Pronasetreatmentcompletely solubilized the32P counts,which establishesthatthe radio-activity was associated with DBP. The
phos-phate linkage was at most 20% resistant to treatmentwith alkali, anditwas90%resistant
toacid,asexpected foraphosphoester of serine orthreonine; this excludedphosphohistidineas a major component of DBP, because
phos-phoimide linkages are stable to alkalai and sensitive toacid (10, 19). Consistentwiththis,
alkaline phosphatase partially solubilized 32P
counts, indicating a phosphomonoester bond
(17).
To identifythe phosphoamino acid, thesame
preparation of 32P-labeled DBP was partially
hydrolyzed bytreatmentwith6NHClat110°C for 2 h, mixed with 40 ,g each of "carrier" phosphoserine andphosphothreonine, and elec-trophoresedonpaperatpH 1.9. Phosphoserine andphosphothreonine markers,notmixed with sample,werealso electrophoresed. The
electro-pherogramwas developed with ninhydrin and
then autoradiographed. The carrier phospho-serineandphosphothreonine inthesample mi-grated slightly behind the marker phospho-aminoacids, possibly becausetrace amountsof materials (e.g., oligoacrylamide) had retarded the mobility of phosphoamino acidsinthe
sam-ple. Tworadioactivespotsappearedonthe
au-toradiogram of thiselectropherogram:one
coin-cided with inorganic phosphate and the other with the carrier phosphoserine ninhydrin spot inthe sample; thissuggeststhat serine butnot threonineresidues ofDBParephosphorylated.
To verify that the radioactivity was in
phos-phoserine, the carrier phosphoserine spot was
eluted, evaporated repeatedly, and reelectro-phoresed on paper. As shown in Fig. 5, the
radioactivity again coincided with phosphoser-ine andinorganic phosphate. In the reelectro-phoresisexperiment,themobility of the carrier phosphoserine inthe samplecoincided exactly with the markerphosphoserine. These results indicate thatserine butnotthreonine residues of DBP arephosphorylated. The 32p04 spot on
thepaperelectropherogramspresumablyarose
by hydrolysisof32P-labeledphosphoserine resi-dues of DBP, because the first electrophoresis step should have eliminated inorganic phos-phate from thecarrierphosphoserine.
In the above experiment, DBP was labeled
from7 to10hp.i.,sothatitisnotclear whether
TABLE 1. Propertiesof the phosphatelinkage
associated withhighly purified 32P-labeled DBPa
Trichloroacetic Treatment acid-insoluble 32p
counts(%)
Control 100
10%Trichloroacetic acid, 20 min at 100
900C
10%Trichloroacetic acid, 20 min at 100
90'C plus ethanol-ether (3:1)
ex-traction, 30 min at 60'C
0.1 N HCl, 15 min at 1000C 90
0.1 N KOH, 15 minat1000C 25
Pronase, 1 h at370C 0
Alkaline phosphatase, 1hat370C 30
a Portions corresponding to about 500 cpm were treated with trichloroacetic acid, trichloroacetic acid
plus organic solvents, HCl, KOH, Pronase, or
alka-line phosphatase (19). One hundred ,ug of bovine
serum albumin wasadded at the end of the
incuba-tion, and the proteins were precipitated with cold
10% trichloroacetic acid. The acid-insoluble
materi-als were collected on glass-fiber filters, washed,
dried,and counted forradioactivity.
both the pulse and pulse-chase forms of DBP contain phosphoserine. To demonstrate that the chased form contains phosphoserine, Ad2-infected cells were labeled with32P04,from7to 7.5 hp.i., thenchasedwith normal MEM for20 h. Total cellular proteins were extracted and immunoprecipitated by Adl-SV40 IgG; then the immunoprecipitates were electrophoresed on SDS gels. As described above, the 32P-labeled DBPband was cutfrom the gel, the DBPwas eluted andwasacid hydrolyzed, andthe hydrol-ysatewassubjectedtopaperelectrophoresis.As shown inFig. 6, the 32P countscoincided with phosphoserine and inorganic phosphate. Thus
the chased formof DBP containsphosphoserine
but notphosphothreonine.
DISCUSSION
Wehave shown that Ad2DBPis
phosphoryl-ated as indicated by (i) identification of 32p_
labeled DBPininfected cell extracts, (ii) bind-ingand elution of32P-labeledDBPfrom
single-strandedDNA-cellulose, (iii) immunoprecipita-tion of32P-labeled DBP by Adl-SV40IgG and monospecific anti-DBPIgG, and (iv) identifica-tionof32P-labeledserine inpurifiedand immu-noprecipitated DBP. Pulse-chase experiments also indicated that DBP exists in atleasttwo forms that differinmobility by
SDS-polyacryl-amide gel electrophoresis. After a short (30 min) pulse, DBP had an apparent molecular
weightof 74K, but, afterapulse followedby a 20-h chase, the apparent molecular weight of DBPincreased to77K, and the74Kformwas no
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.504.268.459.95.219.2]PHOSPHORYLATED Ad2 DNA BINDING PROTEIN 409
MI g 9 P-serine P-ttreonire
FIG 2
FIG. 5. Identification of32P-labeledphosphoserine inhighlypurified32P-labeledDBP.Ad2-infectedcells
werelabeled with 32PO4from 7 to 10 h p.i., and32P-labeled DBP was purified from cytoplasmic extracts by
sequential chromatography on DNA-cellulose and DEAE-Sephadex (27), followed by preparative
SDS-polyacrylamide gel electrophoresis. The32P-labeledDBPband was elated from the gel, partially hydrolyzed in
HCl,mixed withunlabeled "carrier" phosphoserine and phosphothreonine, and electrophoresed on paper as
described inthetext.Phosphoserine andphosphothreonine"markers" (i.e.,notmixed with the 32P-labeled
DBPsample) were also electrophoresed. The "carrier"phosphoserine and phosphothreonine inthe sample
migrated slightly behind the marker phosphoserine and phosphothreonine. The radioactivity coincided with phosphoserineinthe sample and with inorganic phosphate. The phosphoserine radioactive spot was elated
fromthe paper chromatogram and electrophoresed a second time. The results of the second electrophoresis are
shown in this figure. Ninhydrin spray was used to locate the position of unlabeled amino acids, and a Packard radiochromatogram scanner was used to determine the position of radioactivity. The origin is located to the
extremeright of the paper scan.
+ pPi P-serine P-threonine
4 e | *4-is
I t
---
-
.3
_---_-#S
_L
---
'3-
-FIG. 6. Identification of 32P-phosphoserine in pulse-chase labeled 32P-DBP after immunoprecipitation by
Adl-SV4OIgG and preparativeSDS-polyacrylamide gel electrophoresis. Ad2-infected cells were labeled with 32PO4,from 7 to 7.5 h p.i., then chased in normal MEM for 20 h. 32P-labeled DBP was immunoprecipitated
from cytoplasmic extracts byAdl-SV4O IgO, then electrophoresed onSDS-polyaciylamidegels. 32P-labeled
DBP waselatedfrom the gel, acid hydrolyzed, mixed with carrier phosphoserine and phosphothreonine, and
subjected to paperelectrophoresis. The figure shows the correspondence between sample radioactivity and
carrier phosphoserine present in the sample. "Marker"phosphothreonine and phosphoserine (i.e., not mixed
with the sample) had the same electrophoretic mobilities as"carrier"phosphoserine and phosphothreonine in
the sample.
longerobserved.Both74Kand 77Kforms
of35S-and 32P-labeled DBPboundto single-stranded
DNA and were immunoprecipitated by Adl-SV40IgG.
Other workers have commented on altera-tions ofAd2 DBP mobility on
SDS-polyacryl-amide gels. Anderson et al. (3) reportedthat late after infection Ad2 DBP appeared as a diffuseband, and occasionallyas adoublet. In studies of viral-specific proteins during late stages ofinfection, by Walter andMartin (31) withAd2-SV40hybrids,and Weber(32)withan
Ad2latepolypeptide-processingmutant, a sim-ilar reduction inthe mobility of this polypep-tide (presumably) was observed. Lewis et al. (16) found that DBP synthesized by cell-free translation of early Ad2 mRNA occasionally appearedas adoublet. Both formswere identi-calby peptide analysis andwereverysimilarto DBPisolated from infected cells. The results of our pulse-chase experiments, together with data of other workers, may indicate that 74K DBP isthe primary translationproduct thatis then phosphorylated and further modified in VOL. 22, 1977
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.504.49.452.327.442.2]410 JENG ET AL.
some unknown manner that affects the DBP mobility in gel electrophoresis. Both 74K and 77Kforms of35S-and 32P-DBPcouldbe detected when cellswerepulse-orpulse-chase-labeledin the presence of ara-C, which indicates that DBP can be phosphorylated and converted to the apparentlargerform in the absence of viral DNA replication. Highly purified DBP has a frictional ratio of 1.86 and tends to aggregate unless treated with detergent, which suggest that DBP does nothavetypicalglobularprotein structure and has hydrophobicsurfaceregions (27). Possiblyphosphorylation and other post-translational modifications contribute tothese physical properties of DBP.
The roles ofphosphorylation and other possi-ble posttranslational modifications in DBP function areunknown. Both 74K and77Kforms of 35S- and 32P-labeled DBP bind to single-stranded DNA, but we have no accurate esti-mateoftheaffinityofeach formtoDNA.Also, it is not known whether nonphosphorylated
DBP will bind to DNA. Protein phosphoryla-tion isbelievedtoplayakeyregulatoryrolein
eukaryotic cell metabolism. The role of phos-phoproteins (modulated by adenosine
3',5'-cyclicmonophosphate)inintermediary metabo-lism iswell established, and protein phospho-rylation may beinvolvedinregulation of mem-brane function (22). Particularly germane to Ad2 DBP is that phosphorylation ofhistones
and especially acidic DNA binding proteins mayfunction in regulation of gene expression (13, 22,25). The SV40 T-antigen is phosphoryl-ated (R.Carroll, D. Livingstone, P. Tegtmeyer,
personal communication),andthis protein may
havemanybiologicalfunctions,including initi-ationof viral DNA replication and expression of late genes, as well as maintenance of the trans-formed phenotype of SV40-transformed cells. Ad2 DBP seems to be an excellent model sys-temtostudy the general rolethat phosphoryla-tion plays in the biochemistry of eukaryotic DNA binding proteins.
ACKNOWLEDGMENTS
Wethank H. Thornton for cell culture assistance and R. V.Gilden for a gift of the Adl-SV40 antiserum. This work
was supported by Public Health Service grant AI-01725 fromthe National Institute of Allergy and Infectious
Dis-eases andbyresearch contractNO1 CP 43359 within the Virus Cancer Program ofthe National Cancer Institute. W.S.M.W. is a research fellow of the Canadian Medical Research Council. M.G. is a Research Career Awardee of the National Institute of Health (Public Health Service grant 5K6-A14739 from the National Institute of Allergy andInfectious Diseases).
LITERATURE CITED
1. Alberts, B. M., and L. Frey. 1970. T4 bacteriophage gene32: astructural protein inthereplicationand
recombination of DNA. Nature (London) 227:1313-1318.
2. Allerton, S.E., and G. E. Perlmann. 1965. Chemical
characterization of thephospho-protein phosvitin.J. Biol. Chem.240:3892-3898.
3. Anderson, C.W.,P. R. Baum, and R. F.Gesteland.
1973.Processing of adenovirus 2-induced proteins. J.
Virol. 12:241-252.
4. Cantoni, G. L. 1975. Biological methylation: selected
aspects.Annu. Rev. Biochem.44:435451.
5. Gielkens, A. L. J., D. Van Zaane, H. P. J.Bloemers,
and H.Bloemendal. 1976.SynthesisofRauscher mu-rine leukemia virus-specific polypeptides in vitro. Proc. Natl. Acad. Sci. U.S.A. 73:356-360.
6. Gilden,R. V., J. Kern, A. E. Freeman, C. E. Martin, R.
C.McAllister,H. C.Turner,andR. J. Huebner. 1968.
Tand tumorantigens of adenovirus group C-infected andtransformed cells. Nature (London)219:517-518. 7. Gilead, Z., M. Q. Arens, S. Bhaduri, G. Shanmugam,
and M.Green. 1975. Tumourantigen specificityofa
DNAbinding protein fromcells infected with adeno-virus 2.Nature (London) 254:533-536.
8. Gilead, Z., K. Sugawara, G. Shanmugam, and M. Green. 1976.Synthesis of the adenovirus-coded DNA binding protein ininfected cells. J.Virol.18:454460. 9. Green, M., and M. Pina. 1963. Biochemical studies on
adenovirus multiplication. IV. Isolation,purification
and chemical analysis of adenovirus. Virology
20:199-208.
10. Gumport, R.I.,and I. R. Lehman. 1971.Structure of
the DNA ligase-adenylate intermediate lysine
(e-amino)-linked adenosinemonophosphoramidat. Proc. Natl. Acad. Sci. U.S.A.68:2559-2563.
11. Charter, M. L., G. Shanmugam, W. S. M. Wold, and M. Green. 1976. Detection of adenovirus type 2-induced earlypolypeptides usingcycloheximide pretreatment toenhance viral protein synthesis. J. Virol. 19:232-242.
12. Kabat, D. 1971. Phosphorylationofribosomalproteins inrabbit reticulocytes. A cell-free system with ribo-somal protein kinase activity. Biochemistry 10:197-203.
13. Kleinsmith, L. J., J. Stein, and G. Stein. 1976. Dephos-phorylation of nonhistoneproteinsspecifically alters
the pattern ofgene transcription in reconstituted chromatin. Proc. Natl. Acad. Sci. U.S.A. 73:1174-1178.
14. Laemmli, U. K. 1970. Cleavageof structural proteins
duringtheassemblyofthehead ofbacteriophageT4.
Nature(London) 227:680-685.
15. Levinson, A., A. J. Levine, S. Anderson, M. Osborn, B. Rosenwirth, and K. Weber. 1976. The relationship
betweengroupCadenovirus tumor antigen and the
adenovirussingle-strand DNA-binding protein. Cell
7:575-584.
16. Lewis, J. B., J. F. Atkins, P. R. Baum, R. Solem, R. F.
Gesteland, and C. W.Anderson. 1976.Location and
identification of the genes for adenovirus type 2 early
polypeptides. Cell 7:141-151.
17. Perlmann, G. E. 1955. The nature of phosphorus link-ages inphosphoproteins. Adv. Protein Chem. 10:1-30.
18. Philipson, L., U. Pettersson, and U. Lindberg. 1975.
Molecular biology of adenoviruses, p. 1-115. In S. Gard and C. Hallauer (ed), Virology monographs, vol. 14.Springer-Verlag, New York.
19. Rahmsdorf, H. J., S. H. Pai, H. Ponta, P. Herrlich, R.
Roskoski, Jr., M. Schweiger, and F. W. Studier.
1974.Proteinkinase induction in Escherichia coli by
bacteriophage T7. Proc. Natl. Acad. Sci. U.S.A.
71:586-589.
20. Rosenwirth, B., C. Anderson, and A. J. Levine. 1976.
Trypticfingerprint analysis of adenovirus types 2, 5,
and12DNA-binding proteins. Virology 69:617-625.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
PHOSPHORYLATED Ad2 DNA BINDINGPROTEIN
21. Rosenwirth, B., K. Shiroki, A. J. Levine, and H. Shi-mojo. 1975.Isolation and characterization of adenovi-rus type 12 DNAbinding proteins. Virology 67:14-23. 22. Rubin, C. S., and0.M. Rosen.1975. Protein
phospho-rylation.Annu. Rev. Biochem. 44:831-887.
23. Sambrook,J.,M. Botchan,P.Gallimore,B.Ozanne,
U.Pettersson, J. Williams, and P. A. Sharp. 1974. Viral DNA sequences in cellstransformed by simian virus 40, adenovirus type 2 and adenovirus type 5. Cold Spring Harbor Symp. Quant. Biol. 39:615-632. 24. Shanmugam, G.,S. Bhaduri, M. Arens, and M. Green.
1975.DNAbinding proteins in the cytoplasm and in a
nuclearmembrane complex isolated from uninfected
andadenovirus2infected cells. Biochemistry
14:332-337.
25. Stein, G. S., T. C. Spelsberg, and L. J. Kleinsmith. 1974. Nonhistone chromosomal proteins and gene
regulation. Science 18:817-824.
26. Studier, F. W. 1973. Analysis of bacteriophage T7 early RNAs and proteins on slabgels. J. Mol. Biol. 79:237-248.
27. Sugawara, K., Z. Gilead, and M. Green. 1977. Purifica-tion and molecular characterization of adenovirus type2DNAbinding protein. J. Virol.21:338-46.
27a. Sugawara, K., Z. Gilead, W. S. M. Wold, and M. Green. 1977. Immunofluorescencestudyof the adeno-virus type 2single-strandedDNA-binding proteinin
infectedandtransformedcells. J. Virol. 22:527-539.
28. van der Vliet, P. C., and A. J. Levine. 1973. DNA binding proteins specific for cells infected by adenovi-rus. Nature(London) 246:170-174.
29. vanderVliet,P. C., A. J. Levine, J. M. Ensinger, and
H. S. Ginsberg. 1975. Thermolabile DNA binding proteinsfrom cellsinfectedwitha
temperature-sensi-tive mutant of adenovirus defective in viral DNA synthesis. J. Virol. 15:348-354.
30. vander Vliet, P. C., andJ. S. Sussenbach. 1975. An adenovirus type 5 gene functionrequired for initia-tion ofviral DNA replication. Virology 67:415-426. 31. Walter, G.,and H. Martin. 1975. Simian virus
40-spe-cificproteins inHeLa cells infected with nondefective
adenovirus 2-simian virus 40hybridviruses. J. Virol. 16:1236-1247.
32. Weber, J. 1976.Geneticanalysis of adenovirustype 2. III. Temperature sensitivity ofprocessingofviral proteins. J.Virol. 17:462-471.
33. Yamashita, T., M. Arens, and M.Green. 1975.
Adenovi-rusdeoxyribonucleic acid replication.II.Synthesisof
viral deoxyribonucleicacid in vitrobyanuclear mem-brane fractionfrominfectedKBcells.J.Biol.Chem. 250:3273-3279.
34. Yamashita, T., and M. Green. 1974. AdenovirusDNA replication. I. Requirement for protein synthesis and isolation of nuclear membrane fractionscontaining
newlysynthesized viralDNAandproteins.J.Virol. 14:412-420.
411 VOL. 22, 1977
on November 10, 2019 by guest
http://jvi.asm.org/