Evolving gene regulatory networks for cellular morphogenesis
Full text
Figure
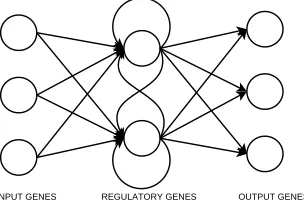
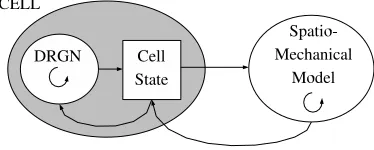
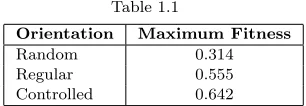

![Fig. 1.4Fittest phenotypes at key stages (generation G) in artificial evolution,where f is fitness:[1.4(a)-1.4(d)] Random growth/division, [1.4(e)-1.4(h)] Regulargrowth/division, [1.4(i)-1.4(l)] DRGN control](https://thumb-us.123doks.com/thumbv2/123dok_us/8513205.350732/14.612.135.442.179.543/phenotypes-generation-articial-evolution-tness-division-regulargrowth-division.webp)
Related documents
Because of the recommended procedure for evaluating reliability (and validity) is to have bilingual participants fill out the questionnaire in both languages and assess the
There are infinitely many principles of justice (conclusion). 24 “These, Socrates, said Parmenides, are a few, and only a few of the difficulties in which we are involved if
Of those who responded, up to 6 respondents did not answer certain questions (1 for years in practice; 6 for number of patients; 2 for frequency of asking patients about
At the turn of the century, a director's compensation essentially con- sisted of the value he received from the enhancement of his equity invest- ment through the
The Houston (First) Court of Appeals agreed with HBU. 59 The court noted that a promise of permanent or lifetime employment must be reduced to writing to be
(A and B) A129 mice were vaccinated with various doses of CHIKV/IRESv1 vaccine or PBS and then challenged 4 weeks later with Asian/American CHIKV strain YO123223, and weight (A)
In conclusion, the finding of this cross-sectional study of pre and post menopausal women show no evidence of a significant difference in BMD between Ocs users and never user
Twenty-five percent of our respondents listed unilateral hearing loss as an indication for BAHA im- plantation, and only 17% routinely offered this treatment to children with
