0022-538X/05/$08.00⫹0 doi:10.1128/JVI.79.17.11014–11021.2005
Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Multiple Gene Segments Control the Temperature Sensitivity and
Attenuation Phenotypes of
ca
B/Ann Arbor/1/66
Erich Hoffmann,
1Kutubuddin Mahmood,
1Zhongying Chen,
1Chin-Fen Yang,
1Joshua Spaete,
1Harry B. Greenberg,
1M. Louise Herlocher,
2Hong Jin,
1and George Kemble
1*
MedImmune Vaccines, 297 North Bernardo Ave., Mountain View, California 94043,1and
Department of Epidemiology, 109 Observatory St., Ann Arbor, Michigan 481092
Received 3 August 2004/Accepted 10 February 2005
Cold-adapted (ca) B/Ann Arbor/1/66 is the influenza B virus strain master donor virus for FluMist, a live, attenuated, influenza virus vaccine licensed in 2003 in the United States. Each FluMist vaccine strain contains six gene segments of the master donor virus; these master donor gene segments control the vaccine’s repli-cation and attenuation. These gene segments also express characteristic biological traits in model systems. Unlike most virulent wild-type (wt) influenza B viruses,caB/Ann Arbor/1/66 is temperature sensitive (ts) at 37°C and attenuated (att) in the ferret model. In order to define the minimal genetic components of these phenotypes, the amino acid sequences of the internal genes ofcaB/Ann Arbor/1/66 were aligned to those of other influenza B viruses. These analyses revealed eight unique amino acids in three proteins: two in the polymerase subunit PA, two in the M1 matrix protein, and four in the nucleoprotein (NP). Using reverse genetics, these eight wt amino acids were engineered into a plasmid-derived recombinant ofcaB/Ann Arbor/ 1/66, and these changes reverted both thetsand theattphenotypes. A detailed mutational analysis revealed that a combination of two sites in NP (A114 and H410) and one in PA (M431) controlled expression ofts, whereas these same changes plus two additional residues in M1 (Q159 and V183) controlled theattphenotype. Transferring this genetic signature to the divergent wt B/Yamanashi/166/98 strain conferred both thetsand the
attphenotypes on the recombinant, demonstrating that this small, complex, genetic signature encoded the essential elements for these traits.
Influenza virus can be isolated from avian and mammalian species, and new strains emerge each year from this gene pool (29). These new strains have the potential to create epidemics that cause approximately 40,000 deaths annually in the United States or pandemics that can result in tens of millions of deaths. Influenza B virus has been isolated from both humans and seals, and the primary mechanisms driving emergence of new type B strains are reassortment among human isolates and antigenic drift (23). Influenza B virus causes excess morbidity and mortality and has been at least partially responsible for 16 epidemics during the past 70 years (31).
Vaccination is the main method of control for influenza epidemics. FluMist, a live, attenuated, trivalent, influenza vac-cine has been shown to be safe, well tolerated, and effective in controlled clinical studies in both children and adults and was recently licensed for the prevention of influenza A and B (for a review, see reference 3). FluMist contains three different 6:2 genetic reassortants, each containing the hemagglutinin (HA) and neuraminidase (NA) gene segments of a currently circu-lating wild-type (wt) strain (i.e., H3N2, H1N1, and B) com-bined with the six internal gene segments (PB2, PB1, PA, NP, M, and NS) of the master donor virus (MDV) derived from eithercaA/Ann Arbor/6/60 (MDV-A) orcaB/Ann Arbor/1/66 (MDV-B).
The properties that attenuate the vaccine and control its replication in humans, thereby conferring its safety and
con-tributing to its effectiveness, are inherited from the six internal gene segments of the MDVs. MDV-B was produced by serial passage of the parental wt B/Ann Arbor/1/66 isolate at succes-sively lower temperatures in eggs and primary chicken kidney (PCK) cells, which resulted in a vaccine strain that differed genetically from the parent isolate. These genetic differences resulted in the phenotypic expression of three characteristic markers that differentiate MDV-B from wt B/Ann Arbor/1/66. These characteristic phenotypes for MDV-B are as follows: (i) cold adaptation (ca), efficient growth at 25°C; (ii) temperature sensitivity (ts), restricted replication at 37°C; and (iii) attenu-ation (att), restricted replication in the lower respiratory tracts of ferrets (21). In addition, these phenotypes are genetically stable; multiple plaque isolates of an MDV-B derived vaccine retained theca, ts, andattphenotypes after prolonged repli-cation in the lower respiratory tract of immunosuppressed hamsters (28).
There is limited information describing which gene segments are necessary and sufficient for expression of the characteristic phenotypes for MDV-B. By using classical reassortment tech-niques, it was reported that the MDV-B PA gene segment could transfer thets andatt phenotypes to the heterologous B/Hong Kong/1732/76 strain (5, 6). Mutation of methionine 431 to isoleucine in this reassortant reverted thetsphenotype (i.e., non-ts) of the virus, demonstrating the critical nature of M431. However, introduction of the PA I431 mutation into MDV-B did not revert thets phenotype (5), suggesting that other loci on the same and/or other MDV-B gene segments contributed to thetsphenotype.
The types of genetic analyses described above using classical * Corresponding author. Mailing address: MedImmune Vaccines,
Inc., 297 North Bernardo Ave., Mountain View, CA 94043. Phone: (650) 603-2356. Fax: (650) 603-3356. E-mail: kembleg@medimmune .com.
11014
on November 8, 2019 by guest
http://jvi.asm.org/
reassortment techniques and heterologous strains are time-consuming. Interpretation of these data are confounded by the fact that gene segments from different strains generally differ from each other by many other sequence differences in addi-tion to the individual residues being targeted for study. The generation of infectious influenza B virus from cloned cDNA, however, facilitates the genetic analysis, since mutations in all eight gene segments can be specifically introduced by recom-binant DNA methods. No selection system is needed to obtain the appropriate recombinant virus, and interpretation of the data is not confounded by other strain differences. Previously, we reported the generation of wt B/Yamanashi/166/98 by transfection of eight plasmids by using the RNA pol I/pol II bidirectional transcription system (8, 9). Here, we generated MDV-B entirely from plasmids. This approach allowed us to generate an isogenic non-ts, non-att recombinant virus that differed by eight amino acids on three gene segments (PA, NP, and M) from MDV-B. The minimal genetic loci responsible for expression of the ts and att phenotypes were identified by constructing specific recombinant viruses with changes in a subset of these eight differences. Introducing a six-amino-acid subset of these differences into a divergent wt strain trans-ferred thetsandattphenotypes, confirming their role in the expression of these phenotypes.
MATERIALS AND METHODS
Cells and virus isolates.caB/Ann Arbor/1/66 (MDV-B) was originally ob-tained from H. F. Maassab at the University of Michigan. MDCK cells were originally obtained from the European Collection of Cell Cultures. Primary chick kidney (PCK) cells were derived in house by trypsinization of the primary tissue and suspension in MEM (Earle’s) medium containing 5% fetal calf serum. PCK cells were seeded in 96-well cell culture plates for 48 h in order to prepare
monolayers with⬎90% confluency.
Cloning of plasmids.Viral RNA was isolated and reverse transcription-PCR was performed as described previously with modifications (8). Briefly, the PCR fragments were digested with BsmBI (or BsaI for the NP segment) and inserted into the BsmBI sites of pAD3000 in a two- or three-fragment ligation reaction. Two to four clones of each plasmid were sequenced and compared to the consensus sequence of MDV-B. Plasmid sequences were corrected, when nec-essary, by routine cloning or utilizing the QuikChange site-directed mutagenesis kit (Stratagene, La Jolla, CA) such that the coding capacity was identical to that of MDV-B. The resultant plasmids were designated pAB121-PB1, pAB122-PB2, pAB123-PA, pAB124-HA, pAB125-NP, pAB126-NA, pAB127-M, and pAB128-NS. The PB1 and HA plasmids, pAB121-PB1 and pAB124-HA, each had two silent nucleotide changes and pAB128-NS had one silent nucleotide change compared to the consensus sequence (PB1, A924G; C1701T; HA, T150C; and T153C NS, A416G). These nucleotide changes did not result in amino acid alterations and were retained to facilitate identification of the recombinant viruses.
Construction of plasmids with wt nucleotide substitutions in the PA, NP, PB2, and M gene segments were constructed by QuickChange mutagenesis of pAB123-PA, pAB125-NP, pAB122-PB2, and pAB127-M plasmids or by ampli-fication of the target region with primers containing the desired mutation and subsequent ligation into the plasmid. The resulting plasmids were sequenced to ensure that the cDNA did not contain unwanted mutations.
The sequence of template DNA was determined by using Rhodamine or dRhodamine dye-terminator cycle sequencing ready reaction kits with AmpliTaq DNA polymerase FS (Perkin-Elmer Applied Biosystems, Inc., Foster City, CA). Samples were separated by electrophoresis and analyzed on PE/ABI model 373, model 373 Stretch, or model 377 DNA sequencers.
Temperature sensitivity assays in PCK cells and MDCK cells.For plaque assays, virus dilutions were incubated for 30 to 60 min at 33°C on confluent MDCK monolayers. The cells were overlaid with 0.8% agarose and incubated at
33°C or 37°C in 5% CO2cell culture incubators. Three days after infection the
cells were stained with 0.1% crystal violet solution or immunostained with chicken anti-MDV-B antiserum, and the number of plaques was determined.
Thetsassay was also performed by titration of the virus samples at 33 and 37°C
on 96-well plates of PCK cells, and the median infectivity was determined [50%
tissue culture infective dose(s) (TCID50)] by examining the cytopathic effect.
Serial 10-fold dilutions of the virus samples were prepared in 96-well blocks. The diluted virus samples were then transferred to the washed PCK monolayer in the 96-well plates. At each dilution of the virus sample, replicates of six wells were used for infection with the diluted virus, and the titer of each virus sample was determined in two to four replicates. Each assay included uninfected control cells
and atscontrol virus. In order to determine thetsphenotype of the virus
samples, the plates were incubated for 6 days at 33 and 37°C in 5% CO2cell
culture incubators, and the virus titers were calculated by the Karber method and
reported as the mean (n⫽4) log10TCID50/ml⫾the standard deviation. The
difference in virus titer at 33 and 37°C was used to determine thetsphenotype.
A virus was classified as temperature sensitive (ts) if its titer at 37°C was at least
2 logs (100-fold) lower than its titer at 33°C. The results shown in the figures are derived from at least two replicates.
Attenuation assay in ferrets.Recombinant viruses obtained after transfection were passaged one time in embryonated chicken eggs to produce a virus stock. Nine-week-old ferrets (Triple F Farms, Syre, PA) were lightly anesthetized with isoflurane and inoculated intranasally by droplets using 0.5 ml per nostril of virus. Three days after infection ferrets were euthanized, and their lungs and nasal turbinates were harvested. A 10% lung tissue suspension was prepared by using OptiMEM I, serially diluted, and 0.1 ml was injected into 10- or 11-day-old embryonated chicken eggs, followed by incubation for 3 days at 33°C. The allantoic fluids were harvested, the presence of virus in the lung tissues was detected by the hemagglutination assay, and the titer was calculated as 50% egg
infectious dose per gram of tissue (log10EID50/g⫾the standard deviation).
Virus replication in nasal turbinates was determined by plaque assay and
ex-pressed as log10PFU per gram of tissue (log10PFU/g).
RESULTS
Generation of MDV-B from eight plasmids. To generate MDV-B entirely from cloned cDNA, either cocultured 293T-MDCK or COS7-293T-MDCK cells were transfected with pAB121-PB1, pAB122-PB2, pAB123-PA, pAB124-HA, pAB125-NP, pAB126-NA, pAB127-M, and pAB128-NS, representing each of the eight gene segments. At 7 days after transfection, su-pernatants of transfected cells were titrated by plaque assay on MDCK at 33°C. The virus titers in the supernatants of cocul-tured 293T-MDCK and COS7-MDCK cells were 5.0⫻106and
7.6⫻106PFU/ml, respectively. The regions encompassing the
five silent nucleotide substitutions were sequenced, confirming that the virus in the supernatant of the infected cells was indeed derived from the plasmids. These results demonstrated that plasmid derived, recombinant MDV-B, designated R1 (rMDV-B), was efficiently recovered after transfection of the eight plasmids.
In order to demonstrate the fidelity of plasmid rescue R1 (rMDV-B) was evaluated for the characteristictsphenotype by measuring the TCID50of the virus on PCK cells at two
differ-ent temperatures. The differences in virus titer between 33 and 37°C for the parental MDV-B and R1 (rMDV-B) were 3.4 and 3.7 log10, respectively (Table 1). Thus, R1 (rMDV-B)
ex-pressed thetsphenotype that was indistinguishable from the parental, nonrecombinant MDV-B.
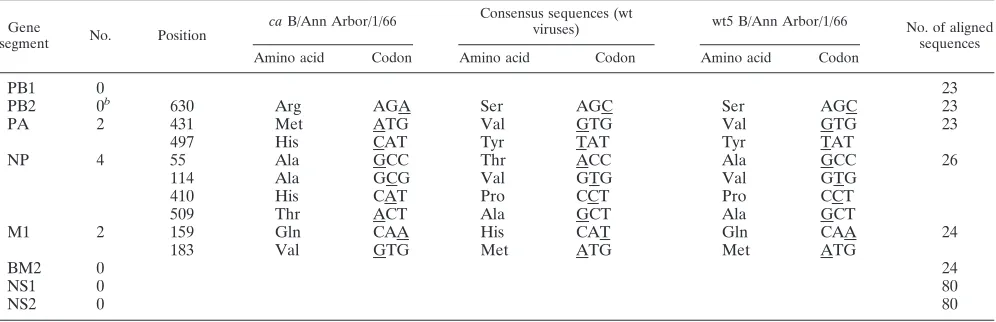
Eight amino acid loci in the internal gene segments are unique to MDV-B.As the first step in elucidating the molecular changes responsible for the characteristic biological properties of MDV-B, including thetsandattphenotypes, the nucleotide and the deduced amino acid sequences of the internal genes of MDV-B were compared to sequences of wt influenza B viruses deposited in the Los Alamos influenza database using the BLAST search algorithm (1, 22). This analysis identified eight amino acids unique to MDV-B that were not present in any other influenza B strain (Table 2); these changes were located
on November 8, 2019 by guest
http://jvi.asm.org/
in the PA (two changes), NP (4) and M1 (2) open reading frames. In addition, one change in PB2 was observed in only one other wt influenza B virus. Sequence analysis of wt5, a virus derived from the original wt B/Ann Arbor/1/66 by three passages in eggs at 25°C (4–6), revealed that this isolate was dissimilar to MDV-B at seven of these nine amino acid posi-tions (Table 2). The wt5 isolate, which expresses non-tsand non-att phenotypes, was identical to the consensus wt se-quences at seven loci, indicating that the progenitor of MDV-B had a genetic constitution reflective of other wt influenza B viruses. These sequence analyses indicated that one or more of the PA, NP, and M gene segments were likely involved in controlling thetsandattphenotypes of MDV-B.
Altering eight amino acid positions of MDV-B reverts thets phenotype.A set of plasmids was constructed in which the eight residues of the PA, NP, and M1 genes were changed by site-directed mutagenesis to the consensus wt amino acids and the resulting recombinant viruses are listed in Table 3. A recom-binant with all eight consensus wt changes, designated R8, was generated by cotransfection of the appropriate plasmids onto cocultured COS7-MDCK cells.
In order to characterize the effect of these genetic changes on temperature-dependent replication, viruses in the superna-tants of the transfections were titrated on MDCK cells by plaque assay and PCK cells by TCID50at 33 and 37°C. As
shown in Fig. 1, R1 (rMDV-B) expressed thetsphenotype in both MDCK and PCK cells, the observed difference in titer between the two temperatures was ⬎3.0 log10. In contrast,
introduction of all eight wt amino acids into MDV-B reverted thetsphenotype; the difference in titer between 33 and 37°C for R8 wasⱕ1.0 log10in both PCK and MDCK cells (Fig. 1).
The R8 recombinant expressed the non-tsphenotype (Fig. 1) and, in sharp contrast to R1 (rMDV-B), R8 produced large and clear plaques at 37°C (Fig. 2). These results showed that alteration of the eight amino acids within the PA, NP, and M gene segments was sufficient to revert thets phenotype and generate a non-tsvirus.
Three of the eight amino acid differences are required to express thetsphenotype.To identify the gene segments that controlled the ts phenotype, a series of recombinants were constructed. Recombinant viruses harboring only one of the wt gene segments were generated, and all of these recombinants exhibited growth restriction at 37°C in MDCK cells and in PCK cells (Fig. 1), indicating that wt amino acid changes in any one gene segment were not sufficient to revert thetsphenotype.
Recombinant viruses containing multiple wt gene segments were constructed to define which combination of segments could revert thetsphenotype. Recombinant viruses that car-ried both the wt NP and wt M (R6) gene segments also re-tained thetsphenotype (Fig. 1). These data demonstrated that the MDV-B PA gene segment was sufficient to express thets
phenotype even in the presence of two other wt gene segments. Similarly, the R5 recombinant harboring both the wt PA and wt M gene segments also expressed thetsphenotype, indicat-ing the MDV-B NP gene segment was also sufficient for ex-pression of thetsphenotype.
[image:3.585.43.285.97.160.2]Only the combination of wt PA and wt NP gene segments together resulted in reversion of the tsphenotype. Both re-combinants R7 and R8 were no longer temperature sensitive in either PCK or MDCK cells. The R8 recombinant virus con-taining the wt PA NP and M gene segments was less restricted at 37°C in PCK cells (Fig. 1) and had a slightly larger plaque size on MDCK cells (Fig. 2) than R7, indicating that the M TABLE 1. Temperature sensitivity phenotype for parental,
nonrecombinant MDV-B and R1 (rMDV-B) generated from eight plasmidsa
Virus
Mean log10TCID50/ml
⫾SD Difference
(log10)
Phenotype
33°C 37°C
caB/Ann Arbor/01/66 8.5⫾0.1 5.1⫾0.1 3.4 ts
R1 (rMDV-B) 7.0⫾0.1 3.3⫾0.1 3.7 ts a
PCK cells were infected with the parent virus MDV-B and the plasmid-derived recombinant virus R1 (MDV-B). The virus titer was determined at two
different temperatures. A difference of 2.0 log10or greater indicates expression
of thetsphenotype.
TABLE 2. Amino acids unique to the internal genes of MDV-B compared to wt influenza B virus sequencesa
Gene
segment No. Position
caB/Ann Arbor/1/66 Consensus sequences (wt
viruses) wt5 B/Ann Arbor/1/66 No. of aligned
sequences
Amino acid Codon Amino acid Codon Amino acid Codon
PB1 0 23
PB2 0b 630 Arg AGA Ser AGC Ser AGC 23
PA 2 431 Met ATG Val GTG Val GTG 23
497 His CAT Tyr TAT Tyr TAT
NP 4 55 Ala GCC Thr ACC Ala GCC 26
114 Ala GCG Val GTG Val GTG
410 His CAT Pro CCT Pro CCT
509 Thr ACT Ala GCT Ala GCT
M1 2 159 Gln CAA His CAT Gln CAA 24
183 Val GTG Met ATG Met ATG
BM2 0 24
NS1 0 80
NS2 0 80
aThe sequence of the internal gene segments of MDV-B was used in a BLAST search. Amino acid positions different between MDV-B and the aligned sequences
are shown. The nucleotides in the codons that are underlined represent the substituted positions.
bThe residue Arg630 of PB2 did not meet the criteria of unique to MDV-B; this amino acid was present in one wt influenza strain, B/Harbin/7/94 (AAF89735). All
other wt strains, including wt5, encoded Ser at this position.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:3.585.44.542.531.692.2]gene segment may have had some impact on expression of the
tsphenotype. The results of these studies demonstrated that the PA and NP gene segments were the major controlling elements of thetsphenotype. The MDV-B PA and NP seg-ments could independently confer the ts phenotype, and a combination of wt amino acids in both these segments was required for reversion of this phenotype.
To determine the contribution of each residue of the four amino acids in the NP protein and two in the PA protein to the
tsphenotype, a detailed mutational analysis of the PA and NP gene segments was performed (Fig. 3). A set of recombinant viruses was constructed in which each virus contained a PA gene with the two wt consensus amino acids and one of the four wt amino acids in the NP gene. Recombinant viruses R9 and R12 with single wt amino acids in NP, T55 or A509, respectively, continued to express thetsphenotype, indicating that these amino acid positions had little role in controlling expression of this phenotype. In contrast, both R10 and R11 containing single wt residues in NP, V114 and P410, resulted in a non-ts recombinant, indicating that these two amino acid positions were critical for control of thetsphenotype. A re-combinant virus (R13) which combined both of these changes (V114 and P410) along with the two wt residues in PA also expressed thetsphenotype.
A similar strategy was used to dissect the contribution of the two amino acids in the PA gene. A set of recombinants was constructed, each harboring an NP gene segment with four wt consensus amino acids and a PA gene with only one of the two consensus wt amino acids. Substitution of H497 with Y497 (R15) remainedts(Fig. 3), demonstrating that this locus had little impact on expression of the phenotype. In contrast, sub-stitution of M431 with V431 (R14) resulted in reversion of the
[image:4.585.46.541.81.320.2]tsphenotype. These results show that amino acids A114 and
[image:4.585.308.536.368.566.2]FIG. 1. Temperature sensitivity of recombinant viruses. Recombi-nant viruses were generated by cotransfection of eight plasmids. R1 (rMDV-B) was constructed by transfection of eight plasmids corre-sponding to the consensus sequence of ca B/Ann Arbor/1/66. The gene segment listed below each set of bars denotes the recombinant virus designation (see Table 3) and wt gene segment(s) in the recombinant virus containing wt consensus nucleotide replacements. The colored bars beneath each label represent the derivation of the PA, NP, and M gene segments in each recombinant (red, wt residues; blue, MDV-B). Each recombinant virus was titrated in PCK cells (dark gray) by TCID50and in MDCK cells (light gray) by plaque assay at 33 and 37°C. The difference in the log10titer at these two temperatures (titer log10 33°C–titer log1037°C) is displayed on theyaxis. The dotted horizontal line represents a difference in which the titer at 33°C is 2.0 log10greater than the titer at 37°C. The error bars represent⫾the standard devi-ation from the mean for the TCID50assays; the light gray bars (plaque assay) represent a typical experiment.
TABLE 3. Genetic composition of recombinant viruses
Designation
Amino acida
PA NP M PB2
431 497 55 114 410 509 159 183 630
MDV-B M H A A H T Q V R
R1 (MDV-B) M H A A H T Q V R
R2 (WT PA) V Y A A H T Q V R
R3 (WT NP) M H T V P A Q V R
R4 (WT M) M H A A H T H M R
R5 (WT PA/M) V Y A A H T H M R
R6 (WT NP/M) M H T V P A H M R
R7 (WT PA/NP) V Y T V P A Q V R
R8 (WT PA/NP/M) V Y T V P A H M R
R9 V Y T A H T Q V R
R10 V Y A V H T Q V R
R11 V Y A A P T Q V R
R12 V Y A A H A Q V R
R13 V Y A V P T Q V R
R14 V H T V P A H M R
R15 M Y T V P A H M R
R16 V H A V P T Q V R
R17 V H A V P T H V R
R18 V H A V P T Q M R
R19 V H A V P T H M R
R20 V Y T V P A H M S
aThe amino acid at the indicated position is given for each gene segment. Wild-type residues are indicated in boldface.
on November 8, 2019 by guest
http://jvi.asm.org/
H410 in MDV-B NP and M431 in MDV-B PA are the major determinants controlling temperature sensitivity of MDV-B.
In order to confirm that these three residues are critical in controlling the expression of the ts phenotype, recombinant
R16 was constructed containing only PA (V431) and NP (V114 and P410) in MDV-B. This recombinant replicated efficiently at both 33 and 37°C; the difference in titer was 0.3⫾0.1 log10
PFU/ml on MDCK cells. Therefore, R16 did not express thets
phenotype. These data demonstrated that these three residues were the minimal requirements for reverting thetsphenotype of MDV-B.
The att phenotype of MDV-B requires loci in addition to those for expression ofts.MDV-B and its 6:2 derivatives are not detected in lung tissues of ferrets after infection by the intranasal route, whereas nonattenuated influenza B viruses are detected in lungs and may occasionally cause other signs of influenza-like illness in these animals (24). To determine whether the eight wt amino acid changes had an impact on the
att phenotype, ferrets were infected intranasally with R1 (rMDV-B) or R8 (wt PA/NP/M). Three days after infection, lung tissues were harvested, and virus was detected by using an egg infectivity assay. As expected, no virus was detected from the lung tissues of ferrets infected with R1. In contrast, the lungs of animals infected with R8 contained approximately 3 log10EID50/g virus (Table 4). Thus, changing the eight unique
amino acids in PA, NP, and M gene segments to wt consensus residues was sufficient to revert theattphenotype to a non-att
phenotype.
In order to determine the minimal number of gene segments required to revert theattphenotype, ferrets were infected with recombinants R3 (wt NP) and R4 (wt M) or R7 (wt PA and NP). Virus was recovered from the nasal turbinates of all of the animals, indicating that each animal had been successfully in-fected; however, in contrast to R8, virus was not detected in the lung tissues (Table 4). These data demonstrated that none of the single gene segments was capable of reverting theatt
[image:5.585.85.494.69.285.2]phenotype, neither was the combination of PA and NP to-gether. All three gene segments—PA, NP, and M—required FIG. 2. Plaque morphology of recombinant MDV-B viruses. Recombinant viruses with wt amino acids in NP, PA, and M were plated on MDCK cells and incubated at either 33 or 37°C for 3 days; the cell monolayer was fixed and immunostained with anti-MDV-B sera. Refer to Table 3 for the designation of the recombinant viruses.
FIG. 3. Contribution of amino acid residues in NP and PA for temperature sensitivity. Viruses were generated by cotransfection of six MDV-B plasmids, together with PA and NP plasmids with the indicated amino acid changes. Gray boxes represent wt amino acid residues present in each recombinant. The TCID50 of the different recombinants was measured at both 33 and 37°C and the difference between the log10values of these titers was plotted on theyaxis. The dotted horizontal line represents a difference in which the titer at 33°C is 2.0 log10greater than the titer at 37°C. The error bars represent⫾ the standard deviation from the mean for the TCID50assays.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:5.585.47.279.410.631.2]the presence of wt amino acid residues to revert theatt phe-notype
To further define the minimal number of changes required to revert theattphenotype, a second study was performed with recombinant viruses containing subsets of wt residues in the PA, NP, and M gene segments. Recombinant R16 contained one wt residue in PA (V431) and two wt residues in NP (V114 and P410); these three changes together were sufficient to revert thetsphenotype. As expected from the previous exper-iment, R16 was not detected in the lung tissues of ferrets 3 days after infection. Recombinant viruses R17 and R18 were con-structed such that each contained one of the wt amino acids found in M in addition to the changes in R16. Both R17 and R18 were detected in the lung tissues of ferrets; however, only two or three of the animals had detectable virus in the lung, respectively. Recombinant R19, which contained both wt M amino acids (H159 and M183) in addition to PA (V431) and NP (V114 and P410), was detected in the lung tissues of all four inoculated animals (Table 4) and replicated to levels only slightly lower than R20 that contained all nine wt amino acids.
Transferring the genetic signature forattto a heterologous influenza strain.Our results with the MDV-B backbone indi-cated that five amino acids in PA (V431), NP (V114 and P410), and M (H159 and M183) were sufficient to revert both thets
andattphenotypes. In order to determine whether changes at these loci could transfer the biological traits to a heterologous nonattenuated wt influenza B virus, the five MDV-B amino acids PA (M431), NP (A114 and H410), and M (Q159 and V183) plus an additional residue in PA (H497) were intro-duced into wt B/Yamanashi/166/98.
Recombinant wt B/Yamanashi/166/98 (recYam) (8) repli-cated well at both 37 and 33°C; the difference in titer was 0.17 log10between these two temperatures. In contrast, rec6Yam,
the derivative recombinant virus containing the six MDV-B amino acids, was clearlyts: the difference in virus titer between 37 and 33°C was 4.6 log10. Both viruses replicated efficiently in
the nasal turbinates of ferrets; however, only wt recombinant recYam was recovered from the lung tissues (Table 4). Thus,
introducing the ts/att loci from MDV-B into a divergent wt strain was sufficient to transfer thetsandattphenotypes.
DISCUSSION
These studies demonstrated that the PA and NP gene seg-ments control the expression of thetsphenotype of MDV-B. Previously, the PA M431 amino acid of MDV-B had been shown to contribute to the control of thets phenotype, but those studies concluded that other residues were also likely to contribute to expression and reversion of this trait (5, 6). The findings reported here show that three amino acid loci of MDV-B—PA M431, NP A114, and NP H410—were the min-imal elements that controlled thetsphenotype, and reversion of all three loci in combination was required to revert thets
phenotype. Although the exact mechanistic roles of PA and NP have not been fully elucidated, there are multiple stages in viral replication that depend on either PA or NP proteins, including those mediated by the viral RNA polymerase complex such as vRNA3mRNA (transcription), vRNA3cRNA (first step in replication), and cRNA3vRNA (second step in replication) or the switch between transcription and replication (18, 26). Since the PA and NP segments were shown to independently impart thets phenotype, viral replication may be blocked at several different points of the replication cycle at 37°C. Restriction of activity at multiple points may explain the several order of magnitude reduction in replication at this higher temperature. Ferrets have body temperatures ranging from 38 to 40°C. Although the inability to replicate at the higher temperatures certainly may contribute to expression of the att phenotype, this defect may not be the sole mechanism for restriction of replication in the lung tissues of these animals. In order to revert theattphenotype, the three changes required to revert the ts phenotype were required in addition to at least one change in the M gene segment. Both changes in the M gene segment encoded changes in the M1 protein. The M1 protein has many different motifs, including a highly conserved zinc finger, and multiple activities during viral replication and pack-TABLE 4. Attenuation studies of recombinant viruses in ferrets
Study no.a Recombinant virus Phenotype Nasal turbinatesb
(log10PFU/g)
Lung tissue
(log10EID50/g⫾SD) (n/N)c
A R1 (MDV-B) ts 4.0 ⬍1.5 (0/4)
R3 (WT NP) ts 4.1 ⬍1.5 (0/4)
R4 (WT M) ts 4.2 ⬍1.5 (0/4)
R7 (WT NP/PA) Non-ts 4.7 ⬍1.5 (0/4)
R8 (WT NP/PA/M) Non-ts 4.7 2.8⫾0.6 (4/4)
B R16 Non-ts 5.0 ⬍1.5 (0/4)
R17 Non-ts 5.3 2.2⫾0.9 (2/4)
R18 Non-ts 5.7 2.8⫾0.5 (3/4)
R19 Non-ts 6.2 2.3⫾0.6 (4/4)
R20 Non-ts 6.1 3.6⫾0.4 (4/4)
A recYam Non-ts 4.9 3.7⫾1.1 (4/4)
rec6Yam ts 4.0 ⬍1.5
aGroups of four ferrets were inoculated with 6 log
10PFU/ml (study A) or 7 log10PFU/ml (study B) Recombinant virus designations are listed in Table 3.
bThree days after infection virus titers of the nasal turbinates and lung tissues were determined; the average titer is shown.
c“⬍1.5” indicates that no virus was detected.n/Ndenotes number of animals with virus detected in the lung tissues (n) compared to the total number of inoculated
animals (N).
on November 8, 2019 by guest
http://jvi.asm.org/
[image:6.585.42.542.80.237.2]aging (2, 7, 12, 19, 25, 32). In addition, the M1 and BM2 products of the influenza B M gene segment are critical for efficient budding of virus particles at the cell membrane (10), the presence of Q159 or V183 in the M1 protein may result in a less efficient production of progeny viruses, resulting in a less efficient spread of virus in the infected host; however, the mechanism for the M gene’s contribution to theattphenotype remains to be investigated.
Studies from a variety of negative-sense RNA viruses dem-onstrate that the genes important for replication and/or tran-scription can be associated with temperature sensitivity and attenuation. Several previous studies and a recent study using recombinant derivatives of the FluMist influenza A MDV (MDV-A) have mapped the amino acids responsible for ex-pression oftsto five sites: three in PB1, one in PB2, and one in NP (11). For RNA viruses with nonsegmented genomes, such as human parainfluenza virus and respiratory syncytial virus, attenuating lesions were found in the phosphoprotein P and the large polymerase L (13–15, 20, 30). Both proteins are components of the viral RNA polymerase. The precise map-ping of the specific residues in PA and NP for MDV-B may now facilitate the elucidation of the underlying molecular mechanisms of those temperature-dependent defects.
The genetic complexity of these MDV-B phenotypes poten-tially reveals the mechanism of genetic stability of thetsandatt
phenotypes. Studies in immunocompromised hamsters, as well as seronegative chimpanzees and humans, have shown that the
tsand att phenotypes of MDV-B and its 6:2 derivatives are stable after many rounds of replication in the host (24, 28). In addition, the uniqueness of these amino acid changes in MDV-B and the observed stability may have resulted from the constant selection of viruses that replicated efficiently as the growth temperatures were lowered. Further genetic and bio-chemical analyses could elucidate the mechanism of action of these vaccine strains. The results reported here demonstrated that expression of the att phenotype is more complex and partially separable fromts.
Transferring the genetic loci responsible for ts and att of MDV-B to the non-ts, non-attrecombinant wt B/Yamanashi/ 166/98 resulted in transferring theattandtsbiological traits to the resulting recombinant. These data confirm that the mini-mal changes required to express the ts and att phenotypes reside within these loci. The genomes of ca B/Ann Arbor/1/66 and B/Yamanashi/166/98 are 96% identical and nucleotide alignments of these two viruses revealed⬎350 nucleotide dif-ferences (⬎50 amino acid differences) between these two strains. Nevertheless, introduction of the MDV-Btsandattloci was tolerated and conferred the appropriate biological prop-erties.
The sequence analysis strategy used here to find unique residues in influenza type B sequences important fortsandatt
should be helpful to find yet-unknownatt/tsresidues in other attenuated viruses, such as ca A/Leningrad/134/57 or B/USSR/69 (16, 17, 27). It is noteworthy that the two wt resi-dues (NP P410 and M1 H159) are conserved within both wt influenza type A and B viruses. The conservation of these residues across this evolutionary distance (⬃30 to 40% amino acid identity between type A and B proteins) may underline the potential importance of these residues for protein structure and function. Our results demonstrate the utility of reverse
genetics in combination with bioinformatics tools to find and characterize the effect of specific residues for the growth of influenza B virus. This genome-wide analysis is important be-cause most of the determinants for biological properties of influenza A and B viruses are multigenic. Thus, this approach provides not only a powerful framework for understanding and potentially enhancing master donor viruses but can also be used for detecting and testing residues of all eight segments important for virus growth, pathogenesis, and transmission.
ACKNOWLEDGMENTS
We thank Jocelyn Ibanez, Hui Ju, and other members of tissue culture facility at MedImmune Vaccines for supplying cells. We thank the animal facility for performing the ferret experiments. We are grateful to Hui Liu and Amy Aspelund for excellent technical assis-tance.
REFERENCES
1.Altschul, S. F., T. L. Madden, A. A. Schaffer, J. Zhang, Z. Zhang, W. Miller, and D. J. Lipman.1997. Gapped BLAST and PSI-BLAST: a new generation
of protein database search programs. Nucleic Acids Res.25:3389–3402.
2.Arzt, S., F. Baudin, A. Barge, P. Timmins, W. P. Burmeister, and R. W. Ruigrok.2001. Combined results from solution studies on intact influenza virus M1 protein and from a new crystal form of its N-terminal domain show
that M1 is an elongated monomer. Virology279:439–446.
3.Belshe, R. B., and W. C. Gruber.2001. Safety, efficacy and effectiveness of cold-adapted, live, attenuated, trivalent, intranasal influenza vaccine in
adults and children. Philos. Trans. R. Soc. Lond. B Biol. Sci.356:1947–1951.
4.DeBorde, D. C., A. M. Donabedian, M. L. Herlocher, C. W. Naeve, and H. F. Maassab.1988. Sequence comparison of wild-type and cold-adapted B/Ann
Arbor/1/66 influenza virus genes. Virology163:429–443.
5.Donabedian, A. M., D. C. DeBorde, S. Cook, C. W. Smitka, and H. F. Maassab.1988. A mutation in the PA protein gene of cold-adapted B/Ann Arbor/1/66 influenza virus associated with reversion of temperature
sensi-tivity and attenuated virulence. Virology163:444–451.
6.Donabedian, A. M., D. C. DeBorde, and H. F. Maassab.1987. Genetics of cold-adapted B/Ann Arbor/1/66 influenza virus reassortants: the acidic poly-merase (PA) protein gene confers temperature sensitivity and attenuated
virulence. Microb. Pathog.3:97–108.
7.Harris, A., F. Forouhar, S. Qiu, B. Sha, and M. Luo.2001. The crystal structure of the influenza matrix protein M1 at neutral pH: M1–M1 protein
interfaces can rotate in the oligomeric structures of M1. Virology289:34–44.
8.Hoffmann, E., K. Mahmood, C. F. Yang, R. G. Webster, H. B. Greenberg, and G. Kemble.2002. Rescue of influenza B virus from eight plasmids. Proc.
Natl. Acad. Sci. USA99:11411–11416.
9.Hoffmann, E., G. Neumann, Y. Kawaoka, G. Hobom, and R. G. Webster.
2000. A DNA transfection system for generation of influenza A virus from
eight plasmids. Proc. Natl. Acad. Sci. USA97:6108–6113.
10.Imai, M., S. Watanabe, A. Ninomiya, M. Obuchi, and T. Odagiri.2004. Influenza B virus BM2 protein is a crucial component for incorporation of viral ribonucleoprotein complex into virions during virus assembly. J. Virol.
78:11007–11015.
11.Jin, H., B. Lu, H. Zhou, C. Ma, J. Zhao, C. F. Yang, G. Kemble, and H. Greenberg.2003. Multiple amino acid residues confer temperature sensitiv-ity to human influenza virus vaccine strains (FluMist) derived from
cold-adapted A/Ann Arbor/6/60. Virology306:18–24.
12.Judd, A. K., A. Sanchez, D. J. Bucher, J. H. Huffman, K. Bailey, and R. W. Sidwell.1997. In vivo anti-influenza virus activity of a zinc finger peptide.
Antimicrob. Agents Chemother.41:687–692.
13.Juhasz, K., B. R. Murphy, and P. L. Collins.1999. The major attenuating mutations of the respiratory syncytial virus vaccine candidate cpts530/1009 specify temperature-sensitive defects in transcription and replication and a
non-temperature-sensitive alteration in mRNA termination. J. Virol. 73:
5176–5180.
14.Juhasz, K., S. S. Whitehead, C. A. Boulanger, C. Y. Firestone, P. L. Collins, and B. R. Murphy.1999. The two amino acid substitutions in the L protein of cpts530/1009, a live-attenuated respiratory syncytial virus candidate vac-cine, are independent temperature-sensitive and attenuation mutations.
Vaccine17:1416–1424.
15.Juhasz, K., S. S. Whitehead, P. T. Bui, J. M. Biggs, J. E. Crowe, C. A. Boulanger, P. L. Collins, and B. R. Murphy.1997. The temperature-sensitive (ts) phenotype of a cold-passaged (cp) live attenuated respiratory syncytial virus vaccine candidate, designated cpts530, results from a single amino acid
substitution in the L protein. J. Virol.71:5814–5819.
16.Klimov, A. I., N. J. Cox, W. V. Yotov, E. Rocha, G. I. Alexandrova, and A. P. Kendal.1992. Sequence changes in the live attenuated, cold-adapted
vari-ants of influenza A/Leningrad/134/57 (H2N2) virus. Virology186:795–797.
on November 8, 2019 by guest
http://jvi.asm.org/
17.Klimov, A. I., A. Y. Egorov, M. I. Gushchina, T. E. Medvedeva, W. C. Gamble, L. G. Rudenko, G. I. Alexandrova, and N. J. Cox.1995. Genetic stability of cold-adapted A/Leningrad/134/47/57 (H2N2) influenza virus: se-quence analysis of live cold-adapted reassortant vaccine strains before and
after replication in children. J. Gen. Virol.76(Pt. 6):1521–1525.
18.Lamb, R. A., and R. M. Krug.2001. Orthomyxoviridae: the viruses and their
replication, p. 1487–1530.InD. M. Knipe, P. M. Howley, et al. (ed.), Fields
virology, 4th ed. Lippincott/The Williams & Wilkins Co., Philadelphia, Pa. 19.Liu, T., and Z. Ye.2002. Restriction of viral replication by mutation of the
influenza virus matrix protein. J. Virol.76:13055–13061.
20.Lu, B., R. Brazas, C. H. Ma, T. Kristoff, X. Cheng, and H. Jin.2002. Identification of temperature-sensitive mutations in the phosphoprotein of respiratory syncytial virus that are likely involved in its interaction with the
nucleoprotein. J. Virol.76:2871–2880.
21.Maassab, H. F., and D. C. DeBorde.1985. Development and
characteriza-tion of cold-adapted viruses for use as live virus vaccines. Vaccine3:355–369.
22.Macken, C. L., H., J. Goodman, and L. Boykin.2001. The value of a database in surveillance and vaccine selection. Elsevier Science, Amsterdam, The Netherlands.
23.McCullers, J. A., G. C. Wang, S. He, and R. G. Webster.1999. Reassortment and insertion-deletion are strategies for the evolution of influenza B viruses
in nature. J. Virol.73:7343–7348.
24.Murphy, B. R., and K. Coelingh.2002. Principles underlying the develop-ment and use of live attenuated cold-adapted influenza A and B virus
vaccines. Viral Immunol.15:295–323.
25.Nasser, E. H., A. K. Judd, A. Sanchez, D. Anastasiou, and D. J. Bucher.1996.
Antiviral activity of influenza virus M1 zinc finger peptides. J. Virol.70:8639–
8644.
26.Portela, A., and P. Digard.2002. The influenza virus nucleoprotein: a mul-tifunctional RNA-binding protein pivotal to virus replication. J. Gen. Virol.
83:723–734.
27.Rudenko, L. G., A. N. Slepushkin, A. S. Monto, A. P. Kendal, E. P. Grigor-ieva, E. P. Burtseva, A. R. Rekstin, A. L. Beljaev, V. E. Bragina, N. Cox, et
al.1993. Efficacy of live attenuated and inactivated influenza vaccines in
schoolchildren and their unvaccinated contacts in Novgorod, Russia. J.
In-fect. Dis.168:881–887.
28.Snyder, M. H., W. T. London, H. F. Maassab, and B. R. Murphy.1989. Attenuation and phenotypic stability of influenza B/Texas/1/84 cold-adapted
reassortant virus: studies in hamsters and chimpanzees. J. Infect. Dis.160:
604–610.
29.Webster, R. G., W. J. Bean, O. T. Gorman, T. M. Chambers, and Y. Kawaoka.1992. Evolution and ecology of influenza A viruses. Microbiol.
Rev.56:152–179.
30.Whitehead, S. S., K. Juhasz, C. Y. Firestone, P. L. Collins, and B. R. Murphy.1998. Recombinant respiratory syncytial virus (RSV) bearing a set of mutations from cold-passaged RSV is attenuated in chimpanzees. J. Virol.
72:4467–4471.
31.Wright, P. F., and R. G. Webster.2001. Orthomyxoviruses, p. 1534–1577.In
D. M. Knipe, P. M. Howley, et al. (ed.), Fields virology, 4th ed. Lippincott/ The Williams & Wilkins Co., Philadelphia, Pa.
32.Ye, Z., T. Liu, D. P. Offringa, J. McInnis, and R. A. Levandowski.1999. Association of influenza virus matrix protein with ribonucleoproteins. J.
Vi-rol.73:7467–7473.