Copyrightq1997, American Society for Microbiology
Adeno-Associated Virus Type 2 DNA Replication In Vivo:
Mutation Analyses of the D Sequence in Viral
Inverted Terminal Repeats
XU-SHAN WANG,1,2KEYUN QING,1,2SELVARANGAN PONNAZHAGAN,1,2
ANDARUN SRIVASTAVA1,2,3*
Division of Hematology/Oncology, Department of Medicine,1Department of Microbiology and Immunology,3
and Walther Oncology Center,2Indiana University School of Medicine, Indianapolis, Indiana 46202
Received 27 November 1996/Accepted 16 January 1997
The adeno-associated virus type 2 (AAV) genome contains inverted terminal repeats (ITRs) of 145 nucleo-tides. The terminal 125 nucleotides of each ITR form palindromic hairpin (HP) structures that serve as primers for AAV DNA replication. These HP structures also play an important role in integration as well as rescue of the proviral genome from latently infected cells or from recombinant AAV plasmids. Each ITR also contains a stretch of 20 nucleotides, designated the D sequence, that is not involved in HP structure formation. We have recently shown that the D sequence plays a crucial role in high-efficiency rescue, selective replication, and encapsidation of the AAV genome and that a host cell protein, designated the D sequence-binding protein (D-BP), specifically interacts with this sequence (X.-S. Wang, S. Ponnazhagan, and A. Srivastava, J. Virol. 70:1668–1677, 1996). We have now performed mutational analyses of the D sequences to evaluate their precise role in viral DNA rescue, replication, and packaging. We report here that 10 nucleotides proximal to the HP structure in each of the D sequences are necessary and sufficient to mediate high-efficiency rescue, replication, and encapsidation of the viral genome in vivo. In in vitro studies, the same 10 nucleotides were found to be required for specific interaction with D-BP, but viral Rep protein-mediated cleavage at the functional terminal resolution site is independent of these sequences. These data suggest that AAV replication and terminal resolution functions can be uncoupled and that the lack of efficient replication of AAV DNA may not be a consequence of impaired resolution of the viral ITRs. These studies further illustrate that the D sequence– D-BP interaction plays an important role in the AAV life cycle and indicate that it may be possible to develop the next generation of AAV vectors capable of encapsidating larger pieces of DNA.
Adeno-associated virus type 2 (AAV) is a nonpathogenic human parvovirus which contains a single-stranded DNA ge-nome of 4,680 nucleotides (37). Optimal replication of the AAV genome requires coinfection with a helper virus such as adenovirus or herpesvirus (2–4). In the absence of a helper virus, the wild-type (wt) AAV establishes a latent infection in which the viral genome integrates into host chromosomal DNA in a site-specific manner (6, 17–20, 32). When a latently infected cell is superinfected with a helper virus, the proviral AAV genome undergoes rescue and proceeds through a nor-mal lytic infection (2, 24, 25). The viral genome can also be rescued from recombinant plasmids containing the wt AAV genome by transfecting plasmid DNA into adenovirus-infected human cells (29, 31). Thus, recombinant plasmids serve as a useful model for studying rescue and replication of the latent proviral AAV. Two AAV sequences are required for viral DNA replication. The first is the viralrepgene, which codes for four nonstructural proteins that are synthesized from a single open reading frame by the use of alternate promoters and splicing (37), and the second is the viral origin of DNA repli-cation, which consists of a 145-nucleotide (nt) inverted termi-nal repeat (ITR) sequence (9, 21). Two of the viral Rep pro-teins (Rep78 and Rep68) are site-specific and strand-specific endonucleases that specifically bind to and cleave at the
ter-minal resolution site (trs) within the AAV ITRs (1, 14–16, 35). AAV genomes with mutations in the repgenes are defective for viral DNA replication (10, 27, 38). The terminal 125 nt in the ITR form a palindrome that can fold back on itself to form a T-shaped hairpin (HP) structure, and the remainder of the ITR consists of a domain, designated the D sequence, which is not involved in HP formation. The terminal HP structure is used as a primer for initiation of viral DNA replication (2, 21, 23, 26, 36). ITRs are required incisfor AAV DNA replication as well as for rescue, or excision, from prokaryotic plasmids (8, 12, 13, 29, 31, 33). Following rescue of the AAV genome from plasmid sequences, only the viral sequences undergo selective DNA replication (12, 29, 44). Our recent studies have demon-strated that the D sequence plays a crucial role in the efficient rescue, selective replication, and encapsidation of the AAV genome (39, 40).
Which of the 20 nucleotides in the D sequence mediate these diverse functions is not known. In addition, thetrsis at the junction of the D sequence and HP sequence, and when a substitute (S) sequence was used to replace the D sequence, it was not clear whether a functionaltrssite had been restored. In order to address these questions, we constructed partial-sub-stitution mutations in the D sequence and studied in detail the effects of these mutations on AAV DNA rescue, replication, and encapsidation in vivo and on D sequence-binding protein (D-BP) interaction andtrsfunctions in vitro. In this report, we present evidence that the proximal 10 nt in the D sequence are necessary and sufficient for the AAV DNA rescue, replication, and encapsidation functions in vivo. However, whereas in vitro D-BP interaction studies closely mimic these functions, the * Corresponding author. Mailing address: Department of
Microbi-ology and ImmunMicrobi-ology, 635 Barnhill Dr., Medical Science Building, Room 231-B, Indiana University School of Medicine, Indianapolis, IN 46202-5120. Phone: (317) 274-2194. Fax: (317) 274-4090. E-mail: arun [email protected].
3077
on November 9, 2019 by guest
http://jvi.asm.org/
Rep-mediated cleavage at thetrsappears to be independent of the D sequence. Thus, an altered trs function may not be consequential for high-efficiency rescue and selective replica-tion of the AAV genome. These studies also suggest the pos-sibility of the development of the next generation of AAV vectors capable of encapsidating larger pieces of DNA.
MATERIALS AND METHODS
Cells, viruses and plasmids.Human nasopharyngeal carcinoma cell line KB was obtained from A. C. Antony, Indiana University School of Medicine, Indi-anapolis, Ind., and maintained as monolayer cultures in Iscove’s modified Dul-becco’s medium supplemented with 10% fetal bovine serum, penicillin, and streptomycin as previously described (36). AAV and Ad2 virus stocks were obtained, respectively, from K. I. Berns, Cornell University Medical College, New York, N.Y., and K. H. Fife, Indiana University School of Medicine, and were propagated as previously described (24, 25). The recombinant AAV plas-mid pSub201 (30) was supplied by R. J. Samulski, University of North Carolina, Chapel Hill, N.C.
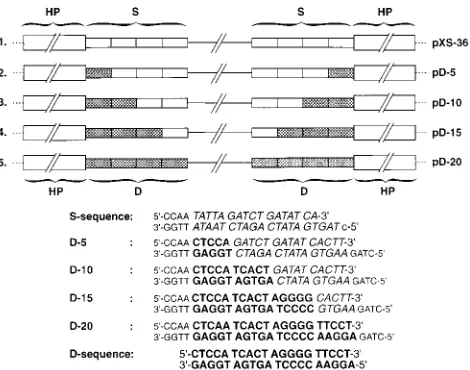
Construction of the recombinant AAV plasmids.Standard cloning techniques were used for constructing all recombinant plasmids (28). The construction of plasmid pXS-36 has been described recently (39, 40). To generate D sequence mutant plasmids containing five successive nucleotide substitutions, the synthetic oligonucleotides shown in Fig. 1 were inserted between theXbaI andBalI sites of plasmid 22 (39) to generate recombinant plasmids 64D5, pXS-64D10, pXS-64D15, and pXS-64D20. Plasmids pD-5, pD-10, pD-15, and pD-20 were generated by ligating the bluntedClaI-PvuII insert from plasmids pXS-64D5, pXS-64D10, pXS-64D15, and pXS-64D20 between theClaI-XbaI sites of plasmids pXS-64D5, pXS-64D10, pXS-64D15, and pXS-64D20, respectively.
AAV DNA rescue and replication assays.DNA-mediated transfections were carried out by the DEAE-dextran procedure (29, 31) with 4mg of each plasmid per 100-mm-diameter dish of 50% confluent KB cells. The transfection mixture also contained 10 PFU of Ad2. At various times posttransfection, low-MrDNA
samples were isolated by the procedure described by Hirt (11), digested exten-sively withDpnI, and analyzed on Southern blots by using32P-labeled AAV
coding sequence-specific DNA probes.
AAV-packaging assays.DNA transfections were performed by the calcium phosphate method (28). Ad2-infected cultures were incubated at 378C in a CO2
incubator for 60 h, and cells were harvested. Cell pellets were subjected to three cycles of freezing and thawing, CsCl was added to a final density of 1.4 g/cm3, and
the mixture was centrifuged in an SW41Ti rotor at 35,000 rpm for 48 h at 208C. Fractions with refractive indexes between 1.3740 and 1.3710 were pooled and dialyzed in phosphate-buffered saline, pH 7.0, followed by exhaustive digestion with DNase I. Clarified supernatants were heated at 568C for 30 min to inactivate Ad2. Equivalent amounts were analyzed on quantitative DNA slot blots with a
32P-labeled AAV DNA probe as previously described (40). Human 293 cells
were infected with these culture supernatants in the presence of Ad2, and the low-MrDNA samples were analyzed on Southern blots as described above.
Preparation of WCE.Whole-cell extracts (WCE) were prepared from HeLa cells in accordance with the method of Muller (22). Total protein concentrations were determined, and the extracts were frozen in liquid N2and stored at2808C.
EMSA.Electrophoretic mobility shift assays (EMSA) were performed in ac-cordance with the method of Carthew et al. (5).32P-labeled oligonucleotides
containing the sequences specific for the S sequence (59-AAGTGATATCAGA TCTAATA-39), the D-5 sequence (59-AAGTGATATCAGATCTGGAG-39), the D-10 sequence (59-AAGTGATATCAGTGATGGAG-39), the D-15 se-quence (59-AAGTGCCCCTAGTGATGGAG-39), and the D-20 sequence (59 -AGGAACCCCTAGTGATGGAG-39) were used as probes. DNA-binding reac-tions were performed in a volume of 20ml with 2mg of poly(dI-dC) and 2mg of bovine serum albumin. The complexes were separated from the unbound probe on low-ionic-strength 4% polyacrylamide gels with recirculating Tris-acetate-EDTA buffer (pH 7.9) containing 6.72 mM Tris-HCl, 3.3 mM sodium acetate, and 1 mM EDTA. In competition experiments, increasing concentrations of unlabeled oligonucleotides were added to the reaction mixtures 10 min prior to the addition of the radiolabeled S sequence, D-5, D-10, D-15, and D-20 probes; this was followed by incubation with the labeled probes and separation as de-scribed above. When D-20 was used as a probe, increasing concentrations of unlabeled S sequence, D-5, D-10, D-15, and D-20 were added to the reaction mixtures as described above.
In vitro Rep-mediated cleavage assays.Appropriate DNA substrates for Rep-mediated cleavage assays were obtained as follows. Plasmid pSub201 was di-gested withXbaI, and, following treatment with shrimp alkaline phosphatase, DNA was digested byPvuII. Plasmids 36, 64D5, 64D10, pXS-64D15, and pXS-64D20 were digested withBsaI and treated with shrimp alkaline phosphatase, followed by digestion withPvuII. ITR fragments were separated on 6% polyacrylamide gels, excised from gels, and eluted. The 59termini of ITRs were labeled with [g-32P]ATP (3,000 Ci/mmol) by using T4 polynucleotide
ki-nase. The labeled ITRs were boiled and quickly chilled, and the 59overhangs were repaired with the Klenow fragment. DNA fragments were separated on 6% polyacrylamide–7 M urea denaturing gels and were eluted as described above. In vitro cleavage assays were carried out in a 20-ml reaction mixture containing 25
mM HEPES–KOH (pH 7.5), 5 mM MgCl2, 1 mM dithiothreitol, 0.5 mM ATP,
0.2 mg of bovine serum albumin, 1mg of poly(dI-dC),;1 ng of the32
P-end-labeled ITRs (105cpm), and 10 ng of purified Rep68 protein, generously
sup-plied by N. Muzyczka, University of Florida, Gainesville, Fla. The reactions were initiated with the addition of MgCl2, and the reaction mixtures were incubated
for 1 h at 378C. The reactions were terminated by the addition of 50ml of phenol and 30ml of Tris-EDTA buffer containing 10mg of tRNA. Following extraction with phenol, the reaction mixture was precipitated with ethanol. The precipitates were dissolved in 90% formamide containing 0.1% bromophenol blue and 0.1% xylene cyanol and were electrophoresed on 6% polyacrylamide–8 M urea se-quencing gels, as previously described (15, 26, 34).
RESULTS
Mutations in the D sequence affect rescue, replication, and
packaging of AAV.The recombinant AAV genomes into which
various mutations were introduced within the D sequence are depicted in Fig. 1. Plasmid pXS-36, in which the D sequences have been replaced by a 20-nt S sequence, has been described recently (39). In plasmid pD-5, nt 6 to 20 in the D sequence were replaced by 15 nt from the S sequence, and in plasmid pD-10, nt 11 to 20 in the D sequence were replaced by 10 nt from the S sequence. Similarly, in plasmid pD-15, nt 16 to 20 in the D sequence were replaced by 5 nt from the S sequence. Plasmid pD-20 contains 20 nt of the authentic D sequence and is similar to plasmid pSub201 except that nt 146 to 190 and nt 4485 to 4530 have been deleted. These plasmids were trans-fected separately into Ad2-intrans-fected KB cells, and low-MrDNA
samples were isolated at various times posttransfection, di-gested with DpnI (45) to degrade unreplicated input plasmid DNA, and analyzed on Southern blots by using 32P-labeled
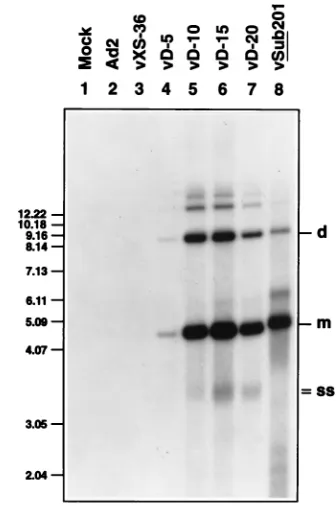
[image:2.612.320.555.68.253.2]probes specific for the AAV-coding sequences. The results of rescue and replication are shown in Fig. 2. It is evident that rescue and replication of the AAV genome from plasmid pXS-36 (lanes 1 to 3) were significantly less than those from plasmid pSub201 (lanes 16 to 18), an observation consistent with our previous studies (39). It is also evident that whereas inclusion of 5 nt of the D sequence augmented rescue and replication of the AAV genome from plasmid pD-5 (lanes 4 to 6), the extent of accumulation of AAV DNA replicative inter-mediates from plasmids pD-10 (lanes 7 to 9), pD-15 (lanes 10 to 12), and pD-20 (lanes 13 to 15) was nearly the same as that from plasmid pSub201. These results suggest that 10 nt of the D sequence proximal to the HP structures are necessary and
FIG. 1. Schematic representation of AAV genomes containing sequential substitutions of the authentic D sequence (hatched boxes, boldface type) with the S sequence (open boxes, italics). Five nucleotides were replaced successively in each of the recombinant plasmids.
on November 9, 2019 by guest
http://jvi.asm.org/
sufficient for optimal rescue and replication of the AAV ge-nome. These results also indicate that nt 46 to 191 present in plasmid pSub201 are not necessary for efficient replication of the AAV genome.
Since we have recently documented that the D sequence serves as a packaging signal for AAV (40), we wished to ex-amine whether the proximal 10 nt in the D sequence were also sufficient for efficient encapsidation of the viral genomes in progeny virions. Various recombinant plasmids were trans-fected into Ad2-intrans-fected 293 cells by the calcium phosphate procedure, and progeny virions were purified on CsCl equilib-rium density gradients. Following digestion with DNase I to degrade any unencapsidated DNA, equivalent amounts of vi-rus stocks were deproteinized to release the AAV DNA and were analyzed on quantitative DNA slot blots by using a32
P-labeled AAV DNA probe as previously described (40). Such a blot is presented in Fig. 3. It can be seen that viral genomes rescued from plasmid pXS-36 failed to undergo encapsidation into viral particles (vXS-36) and that inclusion of the first 5 nt of the D sequence in plasmid pD-5 partially restored encapsi-dation of viral genomes in progeny virions (vD-5). Strong hy-bridization signals were detected with progeny virions vD-10, vD-15, and vD-20 produced, respectively, from plasmids pD-10, pD-15, and pD-20. These signals were comparable to that detected with vSub201 generated from plasmid pSub201. These results further suggest that the first 10 nt in the D sequence are necessary and sufficient for optimal packaging of the AAV genome.
We next examined whether, following encapsidation, the progeny AAV were infectious. Human 293 cells were infected at a multiplicity of infection of 1 in the presence of Ad2. Because of inefficient rescue, replication, and encapsidation of the AAV genome from plasmids pXS-36 and pD-5, approxi-mately 100-fold more vXS-36 and approxiapproxi-mately 60-fold more vD-5 virus stocks were used in these experiments. Equivalent amounts of low-MrDNA isolated 48 h postinfection were
an-alyzed on Southern blots with a32P-labeled AAV DNA probe.
[image:3.612.61.294.67.252.2]These results are shown in Fig. 4. It is evident that the extent of replication of virions produced from plasmids pD-10, pD-15, and pD-20 was roughly the same as that from plasmid pSub201 since monomeric and dimeric viral DNA replicative interme-diates could be readily observed. However, vD-5 viral DNA replication efficiency was significantly less under identical con-ditions. These data provide strong support for the conclusion that the first 10 nt of the D sequence are crucial for efficient rescue and replication of the AAV genome.
FIG. 2. Southern blot analysis of rescue and replication of AAV genomes containing D sequence substitutions. Each of the D sequence-mutated plasmids, along with plasmids pSub201 and pXS-36 as appropriate controls, was trans-fected into Ad2-intrans-fected KB cells, and low-MrDNA isolated at 24 (lanes 1, 4, 7,
10, 13, and 16), 48 (lanes 2, 5, 8, 11, 14, and 17), and 72 (lanes 3, 6, 9, 12, 15, and 18) h postinfection was digested withDpnI and analyzed on Southern blots with a32P-labeled AAV DNA probe as described under Materials and Methods.
[image:3.612.360.511.72.245.2]d and m, dimeric and monomeric viral replicative DNA intermediates, respec-tively.
FIG. 3. DNA slot blot analysis for encapsidation of AAV genomes contain-ing various substitutions in the D sequence. Twofold serial dilutions of equivalent amounts of viral stocks were analyzed with a32P-labeled AAV DNA probe as
described in Materials and Methods.
FIG. 4. Southern blot analysis of replication of progeny AAV containing D sequence substitutions. Equivalent amounts of each of the virus stocks were used to infect Ad2-infected human 293 cells, and replication assays were performed as described in the legend for Fig. 2. Mock-infected or Ad2-infected cells were used as appropriate controls. d and m, dimeric and monomeric viral replicative DNA intermediates, respectively; ss, single-stranded progeny viral DNA strands.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.348.516.422.675.2]Replication of the D sequence mutants in vivo correlates
with interactions with D-BP in vitro.We have recently
dem-onstrated that cellular protein(s) D-BP interacts specifically with the D sequence (40). Since the D sequence mutation analysis indicated that the first 10 nt of the D sequence were responsible for the D sequence functions, we wished to exam-ine whether these mutations also correlated with D-BP inter-actions. WCE were prepared from HeLa cells, were incubated with a32P-labeled D-20 oligonucleotide probe in the absence
or presence of unlabeled D-5, D-10, D-15, D-20, and S se-quence-specific synthetic oligonucleotides, and were analyzed by EMSA as previously described (1, 40). The results are pre-sented in Fig. 5. It is apparent that the D-20 probe formed a specific complex with D-BP and that complex formation could be competitively inhibited by incubations with 10-, 50-, and 200-fold molar excesses of unlabeled D-10 (lanes 7 to 9), D-15
(lanes 10 to 12), and D-20 (lanes 13 to 15) oligonucleotides but not by incubations with the S sequence (lanes 1 to 3) or the D-5 (lanes 4 to 6) oligonucleotides. These data suggest that the first 10 nt in the D sequence play an important role in the interac-tion with D-BP.
These results were further corroborated by performing EMSA with32P-labeled D-5, D-10, D-15, and D-20
oligonu-cleotides under self- and cross-competition conditions. The results are shown in Fig. 6. It is evident that whereas D-10, D-15, and D-20 oligonucleotides formed complexes with D-BP, complex formation with the D-5 oligonucleotide was very inefficient (panel A). Furthermore, complex formation with D-10, D-15, and D-20 oligonucleotides was specific since the binding could be effectively competed with 200-fold molar excess amounts of unlabeled D-10 (panel D), D-15 (panel E), and D-20 (panel F) but not with the S sequence (panel B) or the D-5 oligonucleotide (panel C) under identical conditions. These results establish that the first 10 nt in the D sequence are indispensable for specific interaction with D-BP.
Thetrs function is not affected by mutations in the D
se-quence.Because terminal resolution is a key step during AAV
DNA replication and because it remained possible that the observed lack of optimal rescue and replication of the AAV genome from plasmid pD-5 was due to impairedtrsfunction, we next examined whether mutations in the D sequence af-fected Rep-mediated cleavage at the trs. HP structure DNA substrates were synthesized from each of the recombinant plas-mids, were radiolabeled at the 59 ends with 32P, and were
[image:4.612.65.292.67.267.2]incubated in an in vitro reaction with and without purified Rep68 protein as previously described (15, 34). The data are shown in Fig. 7. Plasmid pSub201 was used as a positive con-trol, and as can be seen, DNA substrate prepared from this plasmid could be effectively cleaved by Rep68 to generate a DNA product with a predicted size of 73 nt (lane 12). Inter-estingly, Rep68-mediated cleavage also occurred with DNA substrates prepared from plasmids pD-5 (lane 8), pD-10 (lane 9), pD-15 (lane 10), and pD-20 (lane 11), and the efficiencies of these cleavages were not significantly different among different D sequence mutations, nor were they significantly different from that with the wt substrate. Furthermore, DNA substrate prepared from plasmid pXS-36, in which the D sequence has been replaced by the S sequence, could also be cleaved by Rep68, albeit less efficiently (lane 7). No cleavage occurred in
FIG. 5. EMSA with the D-20 oligonucleotide. Radiolabeled D-20 probe was incubated with WCE in the presence of up to a 200-fold molar excess of the unlabeled S sequence, D-5, D-10, D-15, and D-20 as competitor oligonucleo-tides. The assays were carried out as described in Materials and Methods. The upper arrow indicates D-BP.
FIG. 6. EMSA with the D sequence-substituted oligonucleotides. The D-BP reaction with each individual radiolabeled oligonucleotide was carried out either in the absence of competitor oligonucleotides (panel A) or in the presence of a 200-fold molar excess of unlabeled S sequence (panel B), D-5 (panel C), D-10 (panel D), D-15 (panel E), and D-20 (panel F) oligonucleotides as competitors. The assays were carried out as described in the legend for Fig. 5. The upper arrow indicates D-BP.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.154.463.524.698.2]the absence of the Rep68 protein (lanes 1 to 6). These data, though not quantitative, suggest that the lack of optimal rep-lication of the AAV genome may not be a consequence of impaired cleavage at thetrs.
DISCUSSION
Recent studies from our laboratory have suggested that the D sequence in AAV ITRs is crucial for efficient rescue, selec-tive replication, and successful encapsidation of the AAV ge-nome (39, 40). In the present study, when 10 nt in the distal half of the D sequence were removed, the AAV genome could still undergo efficient rescue, replication, and encapsidation. However, when this deletion was extended to 15 nt in the D sequence, the efficiency of AAV genome rescue, replication, and packaging was severely compromised. Thus, it would ap-pear that only 10 nt in the D sequence proximal to the HP structures are sufficient to mediate these functions. Why must then the AAV genome carry 10 extra nt in each of its ITRs? Although we did not carry out DNA footprint analyses, on the basis of the data shown in Fig. 6, it would seem reasonable to propose that the proximal 10 nt in the D sequence are required for initiating contact with D-BP and that the remainder of the nucleotides might be involved in enhancing this interaction since there appeared to be a progressive increase in the effi-ciency of D-BP binding with D-10, D-15, and D-20 probes, respectively. These studies, nonetheless, add to the current
understanding of the requirements for an optimally functional
trs.
Correlation between rescue and replication functions.
Ter-minal resolution is a key step during AAV DNA replication. Several studies have suggested that rescue without replication or replication without rescue can occur (8, 12, 13, 25, 41, 44). However, it has been difficult to uncouple these two steps in the virus life cycle. Previous studies have shown that both the secondary structure element of the ITR and a specific se-quence at thetrsare required for Rep-mediated cleavage at the
trsand that 5 to 6 nt at thetrsare required for Rep recognition (34). However, since thetrsis located between the A sequence and the D sequence, i.e., between nt 124 and 125 in the ITR, it was conceivable that mutations in the D sequence led to destruction of the functionaltrs, resulting in impaired rescue and replication of the AAV genome. Interestingly, however, the experimental data did not appear to support this possibil-ity. For example, Rep-mediated cleavage assays in vitro docu-mented that none of the mutations significantly affected Rep protein recognition followed by cleavage at thetrs, regardless of the number of authentic nucleotides of the D sequence. In fact, Rep-mediated cleavage also occurred in the complete absence of the D sequence, suggesting that, at least in vitro, these two steps can be uncoupled and that the lack of optimal replication of the AAV genome is not a consequence of im-paired cleavage at thetrs. However, it remains possible that the efficiency of Rep-mediated cleavage at thetrsis enhanced by D-BP in vivo (39, 40). These data are, nonetheless, consistent with previous studies in which the absence of the D sequence did not prevent the initiation of AAV DNA replication in vitro (43) and in vivo (39, 40).
Implications of the D sequence mutations in the
develop-ment of AAV vectors for gene therapy.AAV has gained
atten-tion as a potentially useful vector for human gene therapy, primarily because of its nonpathogenic nature and its broad host range (23). However, one of the limitations of the cur-rently available vectors is the size of DNA molecules that can be successfully packaged at high efficiency. For example, the wt AAV DNA is 4.68 kb long, but the foreign DNA insert size is limited to about 4.3 kb since ITRs in pSub201-based vectors contain 191 nt each (;0.4 kb). Consequently, some of the larger inserts, such as the cystic fibrosis transmembrane con-ductance regulator cDNA, have been inserted into AAV vec-tors without authentic promoter sequences, and although low-level expression has been obtained (7), it would appear that the inclusion of a strong promoter element upstream of the trans-duced gene would be desirable. In pD-10-based vectors, on the other hand, we have engineered anEcoRV site in each ITR, and as a result, the ITRs contain only 138 nt. Thus, these vectors should allow an increase in packaging capacity of at least 106 nt. Indeed, it has been possible to include additional promoter sequences upstream of large cDNAs, such as the multidrug resistance-1 (MDR-1) and the retinoblastoma sus-ceptibility-1 (RB-1) cDNAs, in the next generation of AAV vectors (42, 46).
ACKNOWLEDGMENTS
We are grateful to Nicholas Muzyczka for his kind gift of the puri-fied Rep68 protein as well as for helpful suggestions. We also thank Kenneth I. Berns for supplying the AAV stock and Richard J. Samul-ski for providing pSub201 plasmid.
[image:5.612.90.268.65.375.2]This research was supported in part by Public Health Service grants (HL-48342, HL-53586, and DK-49218, Centers of Excellence in Mo-lecular Hematology) from the National Institutes of Health and by a grant from the Phi Beta Psi sorority. A.S. was supported by an Estab-lished Investigator Award from the American Heart Association.
FIG. 7. Rep68-mediated cleavage at thetrsin AAV ITRs containing various substitutions in the D sequences. HP DNA substrates prepared from each indi-cated plasmid were radiolabeled at the 59ends and were incubated in the absence (lanes 1 to 6) or the presence (lanes 7 to 12) of purified Rep68 protein in in vitro cleavage assays as described in Materials and Methods. The predicted sizes of DNA cleavage products from pSub201 (solid arrowhead), pD-5 through pD-20 (arrow), and pXS-36 (open arrowhead) are indicated.
on November 9, 2019 by guest
http://jvi.asm.org/
REFERENCES
1.Ashktorab, H., and A. Srivastava.1989. Identification of nuclear proteins that specifically interact with adeno-associated virus type 2 inverted terminal repeat hairpin DNA. J. Virol.63:3034–3039.
2.Berns, K. I., and R. A. Bohenzky.1987. Adeno-associated viruses: an update. Adv. Virus Res.32:243–306.
3.Berns, K. I., R. M. Kotin, and M. A. Labow.1988. Regulation of adeno-associated virus DNA replication. Biochim. Biophys. Acta951:425–429. 4.Buller, R. M. L., J. E. Janik, E. D. Sebring, and J. A. Rose.1981. Herpes
simplex virus types 1 and 2 completely help adenovirus-associated virus replication. J. Virol.40:241–247.
5.Carthew, R. W., L. A. Chodosh, and P. A. Sharp.1985. An RNA polymerase II transcription factor binds to an upstream element in the adenovirus major late promoter. Cell43:439–448.
6.Cheung, A. K. M., M. D. Hoggan, W. W. Hauswirth, and K. I. Berns.1980. Integration of the adeno-associated virus genome into cellular DNA in latently infected human Detroit 6 cells. J. Virol.33:739–748.
7.Flotte, T. R., S. A. Afione, R. Solow, M. L. Drumm, D. Maekakis, W. B. Guggino, P. L. Zeitlin, and B. J. Carter.1993. Expression of cystic fibrosis transmembrane conductance regulator from a novel adeno-associated virus promoter. J. Biol. Chem.268:3781–3790.
8.Gottlieb, J., and N. Muzyczka.1988. In vitro excision of adeno-associated virus DNA from recombinant plasmids: isolation of an enzyme fraction from HeLa cells that cleaves DNA at poly(G) sequences. Mol. Cell. Biol.8:2513– 2522.
9.Hauswirth, W. W., and K. I. Berns.1977. Origin and termination of adeno-associated virus DNA replication. Virology78:488–499.
10. Hermonat, P. L., and N. Muzyczka.1984. Use of adeno-associated virus as a mammalian DNA cloning vector: transduction of neomycin resistance into mammalian tissue culture cells. Proc. Natl. Acad. Sci. USA81:6466–6470. 11. Hirt, B.1967. Selective extraction of polyoma DNA from infected mouse cell
cultures. J. Mol. Biol.26:365–369.
12. Hong, G., P. Ward, and K. I. Berns.1992. In vitro replication of adeno-associated virus DNA. Proc. Natl. Acad. Sci. USA89:4673–4677. 13. Hong, G., P. Ward, and K. I. Berns.1994. Intermediates of adeno-associated
virus DNA replication in vitro. J. Virol.68:2011–2015.
14. Im, D.-S., and N. Muzyczka.1989. Factors that bind to adeno-associated virus terminal repeats. J. Virol.63:3095–3104.
15. Im, D.-S., and N. Muzyczka.1990. The AAV origin binding protein Rep68 is an ATP-dependent site-specific endonuclease with DNA helicase activity. Cell61:447–457.
16. Im, D.-S., and N. Muzyczka.1992. Partial purification of adeno-associated virus Rep78, Rep68, Rep52, and Rep40 proteins and their biochemical characterization. J. Virol.66:1119–1128.
17. Kotin, R. M., and K. I. Berns.1989. Organization of adeno-associated virus DNA in latently infected Detroit 6 cells. Virology170:460–467.
18. Kotin, R. M., J. C. Menninger, D. C. Ward, and K. I. Berns.1991. Mapping and direct visualization of a region-specific viral DNA integration site on chromosome 19q13-qter. Genomics10:831–834.
19. Kotin, R. M., M. Siniscalco, R. J. Samulski, X. Zhu, L. Hunter, C. A. Laughlin, S. McLaughlin, N. Muzyczka, M. Rocchi, and K. I. Berns.1990. Site-specific integration by adeno-associated virus. Proc. Natl. Acad. Sci. USA87:2211–2215.
20. Laughlin, C. A., C. B. Cardellichio, and H. C. Coon.1986. Latent infection of KB cells with adeno-associated virus type 2. J. Virol.60:515–524. 21. Lusby, E., K. H. Fife, and K. I. Berns.1980. Nucleotide sequence of the
inverted terminal repetition in adeno-associated virus DNA. J. Virol.34: 402–409.
22. Muller, M. T.1987. Binding of herpes simplex virus immediate-early gene product ICP4 to its own transcription start site. J. Virol.61:858–865. 23. Muzyczka, N.1992. Use of adeno-associated virus as a general transduction
vector for mammalian cells. Curr. Top. Microbiol. Immunol.158:97–129. 24. Nahreini, P., and A. Srivastava.1989. Rescue and replication of the
adeno-associated virus 2 genome in mortal and immortal human cells. Intervirology 30:74–85.
25. Nahreini, P., and A. Srivastava.1992. Rescue of the adeno-associated virus 2 genome correlates with alterations in DNA-modifying enzymes in human cells. Intervirology33:109–115.
26. Ni, T.-H., X.-H. Zhou, D. M. McCarty, I. Zolotukhin, and N. Muzyczka. 1994. In vitro replication of adeno-associated virus DNA. J. Virol.68:1128– 1138.
27. Owens, R. A., and B. J. Carter.1992. In vitro resolution of adeno-associated virus DNA hairpin termini by wild-type Rep protein is inhibited by a dom-inant-negative mutant of Rep. J. Virol.66:1236–1240.
28. Sambrook, J., E. F. Fritsch, and T. Maniatis.1989. Molecular cloning: a laboratory manual, p. 1.53–1.110. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
29. Samulski, R. J., K. I. Berns, M. Tan, and N. Muzyczka.1982. Cloning of adeno-associated virus into pBR322: rescue of intact virus from the recom-binant plasmid in human cells. Proc. Natl. Acad. Sci. USA79:2077–2081. 30. Samulski, R. J., L.-S. Chang, and T. Shenk.1987. A recombinant plasmid
from which an infectious adeno-associated virus genome can be excised in vitro and its use to study viral replication. J. Virol.61:3096–3101. 31. Samulski, R. J., A. Srivastava, K. I. Berns, and N. Muzyczka.1983. Rescue
of adeno-associated virus from recombinant plasmids: gene correction within the terminal repeats of AAV. Cell33:135–143.
32. Samulski, R. J., X. Zhu, X. Xiao, J. Brook, D. E. Houseman, N. Epstein, and L. A. Hunter.1991. Targeted integration of adeno-associated virus (AAV) into human chromosome 19. EMBO J.10:3941–3950.
33. Senapathy, P., J.-D. Tratschin, and B. J. Carter.1984. Replication of adeno-associated virus DNA. Complementation of naturally occurring rep2 mu-tants by a wild-type genome or an ori2mutant and correction of terminal palindrome deletions. J. Mol. Biol.179:1–20.
34. Snyder, R. O., D.-S. Im, T.-H. Ni, X. Xiao, R. J. Samulski, and N. Muzyczka. 1993. Features of the adeno-associated virus origin involved in substrate recognition by the viral Rep protein. J. Virol.67:6096–6104.
35. Snyder, R. O., R. J. Samulski, and N. Muzyczka.1990. In vitro resolution of covalently joined AAV chromosome ends. Cell60:105–113.
36. Srivastava, A.1987. Replication of the adeno-associated virus DNA termini in vitro. Intervirology27:138–147.
37. Srivastava, A., E. W. Lusby, and K. I. Berns.1983. Nucleotide sequence and organization of the adeno-associated virus 2 genome. J. Virol.45:555–564. 38. Tratschin, J.-D., I. L. Miller, and B. J. Carter.1984. Genetic analysis of
adeno-associated virus: properties of deletion mutants constructed in vitro and evidence for an adeno-associated virus replication function. J. Virol. 51:611–619.
39. Wang, X.-S., S. Ponnazhagan, and A. Srivastava.1995. Rescue and replica-tion signals of the adeno-associated virus 2 genome. J. Mol. Biol.250:573– 580.
40. Wang, X.-S., S. Ponnazhagan, and A. Srivastava.1996. Rescue and replica-tion of adeno-associated virus type 2 as well as vector DNA sequences from recombinant plasmids containing deletions in the viral inverted terminal repeats: selective encapsidation of viral genomes in progeny virions. J. Virol. 70:1668–1677.
41. Wang, X.-S., and A. Srivastava.1997. A novel terminal resolution-like site in the adeno-associated virus type 2 genome. J. Virol.71:1140–1146. 42. Wang, X.-S., and A. Srivastava.Unpublished results.
43. Ward, P., and K. I. Berns.1991. In vitro rescue of an integrated hybrid adeno-associated virus/simian virus 40 genome. J. Mol. Biol.218:791–804. 44. Ward, P., and K. I. Berns.1995. Minimum origin requirements for linear
duplex AAV DNA replication in vitro. Virology209:692–695.
45. Wobbe, C. R., F. Dean, L. Weissbach, and J. Hurwitz.1985. In vitro repli-cation of duplex circular DNA containing the simian virus 40 DNA origin site. Proc. Natl. Acad. Sci. USA82:5710–5714.
46. Woody, M. J., and A. Srivastava.Unpublished results.