Vol. 62, No.7 JOURNAL OFVIROLOGY, JUlY1988, p. 2373-2379
0022-538X/88/072373-07$02.00/0
Copyright ©) 1988, American Society for Microbiology
Structure and Complete Nucleotide
Sequence
of the
Marek's
Disease
Herpesvirus gp57-65
Genet
PAUL M. COUSSENSt* AND LELAND F. VELICER
Department ofMicrobiology and Public Health, Michigan State University,EastLansing, Michigan 48824 Received 9 December1987/Accepted 17 March 1988
The gene encoding the precursor polypeptide of the Marek's disease herpesvirus (MDHV) secretory
glycoprotein gp57-65 (formerly identifiedasAantigen) has been sequenced.Previous results had localized the
genetoa4.6-kilobase (kb)segmentof the BamHI Bfragmentintheunique long regionof the MDHVgenome.
S1 nuclease protection experiments were used tomore precisely locate the 5' initiation and approximate 3'
termination points of the approximately 1.8-kb MDHV gp57-65 mRNA within this segment. These results indicated that the entire MDHVgp57-65codingsequenceis contained withina2.35-kb PvuII-EcoRIfragment,
with the direction oftranscription from PvuII to EcoRI (5'to3'). Nucleotidesequenceanalysis of this region revealedasingleopenreading frame of 1,515 base pairs. The MDHV gp57-65 codingsequencehasanoverall
guanosine-plus-cytosine content of 41 %. Translation of the single open reading frame would produce a polypeptideof 505 aminoacids, withacalculated molecularweightof56,805.Theputative gp57-65precursor
polypeptide contains features common tomany glycoproteins. These includea hydrophobic amino-terminal region (aminoacids1to27) thatmayfunctionas asignal peptide and nine potential N-linked glycosylation sites (Asn-X-Ser/Thr). These two features, predicted from nucleotide sequence data, are consistent with the
published data showing that gp57-65 has a signal peptide and N-linked glycosylation (R. J. Isfort, R. A. Stringer, H.-J. Kung, and L. F. Velicer, J. Virol. 57:464-474, 1986). The predictedsequenceindicatesthat overall the polypeptide is relatively hydrophobic, with a possible 18-residue carboxyl-terminal membrane
anchorsequence. Thissequenceappearstobe less prominent than thosecommonly found in integral membrane glycoproteins. The lack of a strong hydrophobic anchor sequence may help to explain the predominantly secretory natureof MDHVgp57-65.
Marek's disease herpesvirus (MDHV) is the causative agentof Marek'sdisease (MD),alymphoproliferative
disor-der of chickens which results in T-cell lymphomas and
peripheralnervedemyelination (11, 14,15). Anantigenically related apathogenic herpesvirus of turkeys (HVT) has been used as a successful live-virus vaccine which prevents the symptoms ofMD (13) but does not prevent MDHV infec-tion. HVT represents the first successful vaccine against a naturallyoccurring tumor in any species. Thus, the MDHV-HVT system provides a unique natural host animal model system for the study of the immunoprevention and the
pathogenesis ofan oncogenic herpesvirus disease.
To more completely understand the nature of HVT-in-duced immunity to MDHV symptoms, we and others have
investigated antigens common toboth viruses (8, 9, 13, 17, 19, 23, 24). At least sixvirallyencodedantigenic proteins are
common between MDHV and HVT (4, 22). One of these proteins, previously designated the A antigen (2, 5, 8, 9), is
a predominantly secreted glycoprotein with an apparent molecularweightof 57,000to65,000 in itsfully glycosylated form (6), hereafter referredto asgpS7-65. Acombination of pulse-chase and cell-free translation studies have shown that
gp57-65 isfirst synthesized as aprecursorpolypeptide with
an apparent molecular weight of 44,000 (pr44). Previous estimates of molecular weightwere based on an ovalbumin marker size of 46,000 daltons and indicated that the size of thegp57-65precursorpolypeptide was 47,000 daltons (6). A
*
Corresponding
author.t Article no. 12456 of the Michigan Agricultural Experiment Station.
tPresent address:Departmentof AnimalScience,MichiganState University, EastLansing,MI 48824.
later revision of the ovalbumin marker size to 43,000 daltons suggests that the apparent molecular mass of the gp57-65 precursorpolypeptide is, infact, 44,000 daltons.
Aftercleavage ofasmallsignalpeptide, the polypeptideis
glycosylated and secreted from the cell in a precisely pro-grammedmanner(6). Althoughgp57-65 isoneof theprimary proteins recognized by sera from MDHV- or HVT-infected chickens,itspreciserole,ifany,intheimmunopreventionor
pathogenesis ofMD is unclear. It has been postulated that the extensively secreted gp57-65 may play an
immunoeva-siverole in the pathogenesisof MD(6), and it isacandidate for causing the early-stage immunosuppression that occurs after MDHVinfection(5).
Recently, the gene encoding the precursorpolypeptideof MDHVgp57-65 waslocalized to a4.6-kilobase(kb) PvuII-SmaI segment of the BamHI Bfragment(5), with reference
to the restriction map of the MDHV genome generated by Fukuchietal. (1). In the present report, we describea more
precise localization of the MDHV gp57-65 gene, presentits
complete nucleotide sequence, and analyze its predicted precursorpolypeptide, especiallyin relation to known prop-erties of the antigen.
MATERIALS ANDMETHODS
Cell cultures and viruses. The preparation, propagation, and infection of small-scale duck embryo fibroblast (DEF) cell cultures with MDHV were performed as described
previouslybyGlaubigeretal. (2) andmorerecently by Isfort
etal.(5). MDHV strain GAwasusedatpassage level 6after
itwasisolated from feathertipsobtainedfrom infected birds (21).
Restriction enzymes, generation of M13 subclones, and nucleotide sequencing. The MDHV (strain GA) BamHI
li-2373
on November 10, 2019 by guest
http://jvi.asm.org/
2374 COUSSENS AND VELICER
brary was a gift from M. Nonoyama, Showa University Research Institute (1). The MDHV DNA in each clone was propagated in Escherichia coli DH-5 and was extracted by standard methods (10). All restriction and modifying en-zymes werepurchased fromeither Bethesda Research Lab-oratories, Inc. (Gaithersburg, Md.) or Boehringer Mannheim Biochemicals (Indianapolis, Ind.) and used according to the instructions of the manufacturer. In preparation for nucleo-tide sequencing, the 2.35-kilobase-pair (kbp) PvuII-EcoRI segment of the BamHI B fragment was subcloned into plasmid pUC19 cleaved with SmaI and EcoRI to generate plasmid p19MDA2.35 (ATCC 40312). This cloning
proce-dure destroyed the PvuII site of the MDHV fragment but placed a convenient BamHI cleavage site from the multiple
cloningregion of pUC19 4 nucleotides 5' of theSmaI-PvuII
junction. Subclones of MDHV DNA were prepared in
M13mpl8-M13mp19 vectors by standard techniques (10).
Single-strand recombinant M13 DNA templates were
pre-pared by polyethylene glycol precipitation of virus from culture supernatants andextraction with phenol and
phenol-chloroform(1:1), followed byethanol precipitationof single-stranded viral DNA. Templates were sequenced by the
dideoxy chain-termination method (20), using the Klenow fragment of DNA polymeraseI,[a-32P]dATP,and an appro-priate ratio of dideoxynucleotides to deoxynucleotides.
Products of sequencing reactions were analyzed on 8% polyacrylamide gels containing8 M urea.
IsolationofcellularRNA.TotalcellularRNAwasisolated
from normal and MDHV-infected DEF cells essentially as
described previously (10). Briefly, cells were lysed in 4 M
guanidinium isothiocyanate. DNA wassheared by repeated
passageofthe celllysates through an18-gauge syringe. The final preparation was loaded on a cushion of5.7 M CsCl in 0.1 M EDTA. Centrifugation was carried out at a force of
174,000 x g for 16 h. RNA pellets were solubilized in a
solutioncontaining100 mMTrishydrochloride(pH 8.0) and 10 mM EDTA and extracted twice with phenol-chloroform
(1:1) and once with chloroform. After precipitation with
ethanol, RNAwassolubilized inwater at aconcentration of 5 Lg/,uland storedat -70°C untiluse.
Si
nucleaseprotection studies.Nucleaseprotection studieswere performed essentially as described elsewhere (10),
usingtheprobes described in thetext. Hybridizations were
routinelyperformedat50°Casrecommended forDNA, with
an averageG+C content ofapproximately 46% (10).
a P/SI
B} 1K
I
K BIKb 00 1.0
S\N
6
sv 201
2.0 E
I I I
ru
b 2 3 4 5
234
6 7 8 Size(bp)
- 1,353
* - 622
-527 - ~-404 - 309
.242 -238 -217
-201
-
180
FIG. 1. Finestructuremapping oftheMDHVgp57-65gene. The orientation andlimitsoftheMDHVgp57-65gene weredetermined by
Si
nuclease protection experiments, using probes I and 11 (a). Probestodeterminetheorientation and5'limitswere5'endlabeled with [-y-32P]ATP, using T4 polynucleotide kinase. After denatur-ation, hybridization to RNA, and S1 digestion, protected species wereanalyzedon5% denaturingpolyacrylamidegels(b). Lanes:1, Probe 11withoutRNAand S1; 2, probeI withoutRNAandSi;
3, probe II plus MDHV-infectedDEFRNAandS1 nuclease; 4,probe 11 plus uninfected DEF RNA and Si nuclease; 5, probe I plus MDHV-infected DEF RNA andSi
nuclease; 6, probe I plus uninfectedDEF RNA andSi
nuclease;7and8;sameaslanes 5and 6,respectively,except that thehybridizationtemperaturewas45°C instead of50°C.Todeterminetheapproximate3'limitofthe MDHVgp57-65 gene (panel a, dashed arrow), probe I1 was labeled with
[(a-32P]dGTP,
usingtheKlenowfragment ofE. coliDNA polymer-ase 1. Theapproximate5' initiation and 3' termination sites ofthe MDHVgp57-65mRNAareindicatedinpanelaby solid and dashed arrows, respectively. Restriction enzymes used in subcloningand thegeneration of probeswereBamHl(BI),Bglll(BII), KpnI (K),Sspl (S), NcoI (N), EcoRV (V), EcoRI (E), PvuII (P), andSmaI
(SI).
RESULTS
Precise mapping of the MDHV gp57-65 gene. Previous
results hadlocalized the MDHV gp57-65 (previous nomen-clature, MDHV A antigen) gene to a 4.6-kb PvuII-SmaI segment of the BamHI B fragment but had not precisely mapped the gene or determined its orientation 5'->3' (5). Beforenucleotidesequenceanalysis,weattemptedtodefine the5' and3' ends of thegene.Resultsfrom SP6
polymerase-and T7 polymerase-generated single-stranded RNA probes
had indicated that the direction oftranscription was 5'->3' PvuII-+EcoRI (P. M. Coussens and L. F. Velicer,
unpub-lished results). To confirm this result and to locate the
approximate RNAstart site, 5'-end-labeled probes I and II
(Fig. la)wereprepared.Afterdenaturation, hybridizationto MDHV-infected cell RNAoruninfected DEFRNA,and S1
nuclease digestion as described in Materials and Methods,
the protected fragments were sized on a 5% denaturing polyacrylamide gel and detectedbyautoradiography. Probe Iextended from theBamHI sitetotheBglIIsite(Fig. la)and
migratedwithanapparentsize ofapproximately650bp
(Fig.
lb, lane 2). Probe II extended from the BglII site to the
EcoRI site and was
approximately
1,400 bp long(Fig. lb,
lane 1). After denaturation and
hybridization
to MDHV RNA, two segmentsofprobeI,
a majorspecies
of approx-imately 486 nucleotides and a minor species ofapproxi-mately 364 nucleotides, were protected from S1 nuclease
digestion (Fig.
lb,
lane 5). As expected, uninfected DEF RNA was unable to protect probe I from S1 nucleasedigestion (Fig. lb, lane 6). Identical results were obtained
with probe I when the hybridization temperature was
re-duced to 45°C (Fig.
lb,
lanes 7 and 8). These results suggested that two possible transcriptional initiation sites existfor the MDHVgp57-65
gene andconfirmedthe5'->3'orientation
predicted
by the SP6 polymerase- and T7poly-merase-generated RNA probes. In addition, these results
indicated that the 5' end of the MDHV
gp57-65
mRNAmappedatmost486nucleotides5' of the
BglII
site. ProbeII was completely digestedbySi
nuclease in the presence ofJ. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.335.549.68.321.2]STRUCTURE AND SEQUENCE OF THE MDHV gp57-65 GENE
(Kbp)0
B T
I I
0.5
T T g T
I aI
1.0 T
1.5 20
TN T
0 .- it
2.5
TR E
.1 I
-4 __
4t-c .
*e
4~ ~ _
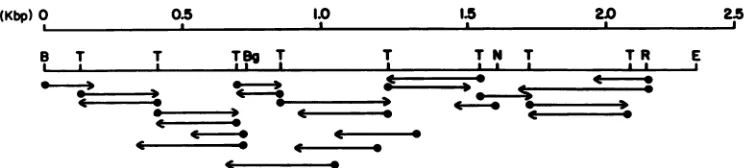
FIG. 2. Strategy of andclones prepared for nucleotide sequence analysis of the 2.35-kbPvuII-EcoRI fragment of the MDHV genome containing all of the MDHV gp57-65 coding region, determined by S1 nuclease digestionasdescribed in thelegendtoFig. 1.All cloneswere prepared in eitherM13mpl8orM13mpi9asappropriate. Arrowsindicate thedirection andextentofDNAsequenceanalysison aparticular fragment.Restrictionenzymesites shownareBamHI(B),BgIII(Bg),NcoI (N), TaqI(T), EcoRV (R), and EcoRI (E).
MDHV-infected cellRNA oruninfectedDEF RNA(Fig.lb, lanes 3 and 4, respectively), asexpected from our previous
results and theprotection of probe I in thepresent study. TheMDHVgp57-65 mRNA has been shown by Northern
(RNA) blot analysis to be approximately 1.8 kb long (5; P. M. Coussens and L. F. Velicer, unpublished results). Theseresultscombinedwith the 5'Simapping data strongly
suggested that the MDHV gp57-65 3' terminus was within
the 2.35-kb PvuII-EcoRI segment. However, the results of
Isfort et al. (5) indicated that the adjacent 2.4-kb
EcoRI-SmaI fragment could hybrid select gp57-65 mRNA and
hybridizeto asmallportion of gp57-65mRNAon Northern
blots (5).To morepreciselydefine the 3'limit ofthe MDHV
gp57-65 mRNA,Si nuclease protection studies using
3'-end-labeled probes I and II (Fig. la) were performed. As
ex-pected, probeI, whichwaslabeledonthecoding strandfor
3' endmapping, was notprotected from Si nuclease diges-tion by hybridization with MDHV-infected cell RNA (data notshown). However,fragments of probeII wereprotected (data not shown), which suggests that the 3' end of the
gp57-65 coding region was contained entirely within the
2.35-kb PvuII-EcoRI fragment. These results (Fig. la, dashed arrow) were in agreement with the interpretation obtained bycombining the 5'
Si
mapping data (Fig. 1)and datafrom Northern blots probed withsubfragments ofthisregion (Coussensand Velicer, unpublishedresults). Nucleotide sequence of the MDHV gp57-65 gene. The results obtained by
Si
nuclease protection analysis hadlocalizedthe MDHVgp57-65gene within the 2.35-kb PvuII-EcoRIsegment oftheBamHIB fragment. As apreliminary
step to nucleotide sequence analysis, an ordered set of
M13mpl9subclones for thisregionwasprepared by cloning
TaqI
digestion products intothe AccIsite ofM13mpl9. Thepreparation ofadetailed TaqI restrictionmapofthegpS7-65 coding regionalso allowedrapidcomparison ofthegpS7-65
genesisolated from variousMDHVandHVTisolates(P. M.
Coussens, R. A. Stringer, and L. F. Velicer, unpublished
results). Severalclones representing each of the TaqI frag-ments wereisolated. Nucleotide sequence analysisof these
clones allowedrapid bidirectional sequencing ofmostof the
2.35-kb PvuII-EcoRI segment (Fig. 2). Junctions were
se-quencedfromadditional clones generatedbyeitherBAL31
digestion from specific sites or directed cloning in M13 vectors with the restrictionsites indicated in Fig. 2. Terminal sequences were determined by cloning the entire 2.35-kb
fragment into both M13mpl8 and M13mpl9. Since this
represented the first MDHV gene to be sequenced in its
entiretyand it would be used as a standard forcomparison
with the
gpS7-65
genesofother MDHV strains and serotypes(including
HVT), many regions were sequenced several times in bothdirections (Fig. 2).The complete nucleotide sequence ofthe region
contain-ing the putative gpS7-65 coding region is shown in Fig. 3. There is asingle large openreading frame within this region extending from the ATG codon beginning307 bp 3' ofthe
BamHI site (nucleotide 1 in Fig. 3) ofp19MDA2.35 to the
termination codon TAG, startingatnucleotide1516ofFig.3.
Translationofthis 1,515-bpopenreading frame (Fig. 3,line
2) would produce a polypeptide of 505 amino acids, with a
calculated molecularweight of 56,805. There arefour poten-tial TATA sequencesinthe307bpimmediately upstreamof
the MDHVgpS7-65 initiation ATG. These occur at
positions
-49 to -52, -190 to -193, -239 to -242, and -276 to -279. There are threesequences which may act aspotential CAT-box regions, ranging from nucleotides -94 to -100, -161 to -172, and -207 to -219. The sequence AATAAA
from -157 to -163 represents a possible polyadenylation signal fora gene 5' to the MDHV gpS7-65gene. The same AATAAA sequence occurs onceafter thegpS7-65 termina-tioncodonbetweennucleotides1655 and 1661 (not shown in
Fig. 3) and is the probableMDHV
gpS7-65
poly(A) addition signal. Consistent with buoyant density measurements oftotal MDHV DNA (7), the overall guanosine plus cytosine (G+C) contentofthe
gpS7-65
coding regionis 41.6%.The predicted amino acid sequence of the precursor
MDHV
gpS7-65
polypeptide, translated from the DNA se-quence,exhibits featurescommon tootherglycoproteins.
A sequence of 19 hydrophobic amino acids (Fig. 3) in the amino-terminal region may function as part ofthegpS7-65
signal peptide (5). This region ends in a sequence that appears tofollow the rulesof Pearlman and Halvorson (16)
forpredicting signal peptide cleavage sites. Inthis case, the
peptide cleavage site would be the alanine (amino acid 28) whosecodon begins at nucleotide 82 (Fig. 3). There are also
ninepotentialN-linkedglycosylationsites(Asn-X-Ser/Thr),
which areunderlinedin Fig. 3.
Analysis ofthehydrophilicity ofthepredicted amino acid
sequence (Fig. 4) indicates that, overall, the protein is
relatively hydrophobic and contains an 18-residue (amino acids 461 to 479) hydrophobic sequence in the carboxyl terminus.Despite thepredominantly secretory natureofthe
gpS7-65
polypeptide, thisregion orothers of similar hydro-phobicity could function as possible membrane anchor se-quences similar to those found in glycoproteins which are known to interact with the plasma membrane. Thehydro-phobic sequence representing the putative
gpS7-65
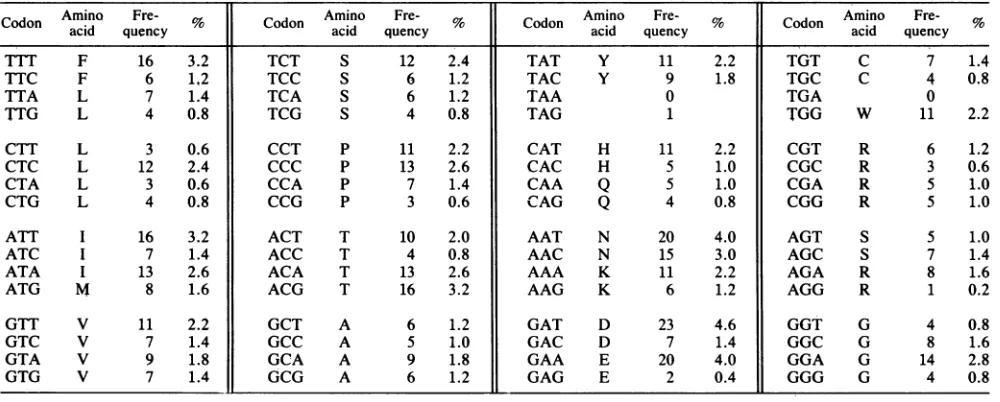
signal peptide, residues 1 to 27, is clearly visible in this analysis (Fig.4). Consistentwith the hydrophilicity plot (Fig.4), the codon usage table(Table1) shows that thepredictedgpS7-65
aminoacid sequencecontains 55.4%nonpolaramino
acids,
including tryptophan (8.6%) and isoleucine
(7.2%)
as the most abundant amino acids in the sequence.2375 VOL.62, 1988
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.117.493.70.154.2]-300 -290 -260 -270 -260 -250
* * * * * *
OCA TCC CCC TCC CTA TCT ACT CAAATC TAT ACC TCG CAT TTA TCA TAA CTA CTT TAC TTO
-240 -230 -220 -210 -200 -190
* * * * * *
Tm CTA TAT TCA CTA CCC CTA TCT TOA CCAMT COTTCT TCA CAT CTS CCC CAT ATA COT
-180 -170 -160 -150 .140 -130
* * * * * *
ATT OAT COT TCT TTC CAA COO COA ASAMA CTI TCA TAC ATA CTAMCOATCTOA OTT
-120 .110 -100 .90 -80 -70
* * * * * *
TIT TAT GAG COT TCA MA CAA AGO TAC CAT COO TTT AM ACT MC 170 CAT ATC CTA ATC
-60 -50 -40 -30 -20 -10
* * * * * *
CAC MA MT CAT TTT ATA CTCAT CCC CM GA A CCAMC OTA ACC CTC TAC ATA TCT
1 10 20 30 40 S0
* * * ,
-*---4---*-____-TCC CTCATO CTC AGO CCOC COT OT TA CCA OCT TIC0OC TOG ACT COA CTC TTT Tm TTC Not Lou Thr fto Arg Va1 Lou ArgAl Lu OlyTrp Thr Cly Lou Ph. Ph. Lou
60 70 t0 90 100 110
~~- ---* * * *
CTT TTA TCT CCC ACMC OTC CT OGA OCC ACC CTTACC OG CAT CTC CAA CAC CCC ATT Lou Lou Sor tro Ser As Va1 tLu Oly Ala Sr Lou Ser
Arg
Asp Lou Clu His Pro Ile120 130 140 1S0 160 170
* * * * * *
CTA TCC TIT CAT CCA TCC MC ATT TC ATT MCC¢C OCC CCT TIAACT GAG CO CCT CAT Lou S.r Ph. Asp Pro Ser AwnlIe Sewr Ile Asa ly Ala Pro Lou Thr GluVa1 Pro His
1S0 190 200 210 220 230
* * * * * *
CCA CCT TCC AC CM ACT OTC TCA ACA MTTCO CMA AGTACC MT CM CATACC ATA ACA Ala fto SrThr Clu SerVa1 Ser Thr Asa Ser Clu Ser Thr As. Clu His Thr IlI Thr
240 250 260 270 280 290
* * * * * *
CM ACO ACO 0C0 MC MC OCA TAC ATC CAC MC MTCC TCT ACO CAC MC CM MT CCC Clu Thr Thr Oly Lys Asn Ala Tyr Ile His s Asa Al Ser Thr Asp Lys Gln As. Ala
300 310 320 330 340 350
* * * * * *
MC OAC ACT CAT MAA ACO CCC MT ATA CTC TOC CAT ACO GM CM G01 117 01 TTC CTT AsnAsp Thr His Lys Thr Pro Asn Ile Lou Cys Asp Thr Clu Clu Val Ph. V-1 Ph. Lou
360 370 380 390 400 410
* * * * * *
MC GM ACG GGA mA TTT GTT TOT ACT CTC MA C01 OAC CCC CCC TCCOAT ACT CM TOG AsnGlu Thr Gly Ar; Ph. Va1 Cys Thr Lou Lys Va1 Asp Pro Pro Ser Asp Ser CluTrp
420 430 440 450 460 470
* * * * * *
TCC MC TTT GTT CTA GAT CTG ATC Tm MC CCA ATT GM TAC CAC CCC MC CM MC MT Ser Asn Phe Va1 LouAsp Lou Ile Ph. Awn Pro Ile luTyr His Ala Awn Glu Lys Asn
480 490 500 510 520 530
* * * * * *
T0 GCM GCG GCG CGT ATC GCT GGT CTC TAT GGA GTC CCC OCA TCA CAC TAT GCA TAC CCA Va1 Glu Ala AlaAr; Ile Ala Gly Lou Tyr GlyV.1 Pro Gly Ser Asp TyrAla lyr Pro
540 550 560 570 580 590
* * * * * *
COT CM TCT CM TTA ATT TCT TCC ATT CGA CGA OAT CCC CAC C ACA TT1 TOGACC AMC
Arg Cln S-r Clu Lou Ile Ser S.r IleArg Arg Asp Pro Gln Gly Thr Ph. Trp Thr Ser
600 610 620 630 640 650
* * * * * *
CCA TCA CCT CAT GGAMC AMG TAC TTC ATA TOG ATAAMC MA AGA ACC MT ACO ATO0CC
fto S.r Pro His Gly Awn Lys Tyr Ph. Ile Trp Ile Asn Lys Thr ThrAwn Ther Ht Gly
FIG. 3. DNA sequenceof theMDHVgp57-65gene. The sequenceis numbered with+1asthefirstbaseoftheinitiationATG codon of the 505-amino-acid open reading frame. The predicted aminoacid sequence of thegp57-65 precursor polypeptide is written below the nucleotide sequence,arrangedascodonsin segmentsofthreenucleotides. Thehydrophobicamino-terminalsequence that mayfunctionas
a signal peptide is highlighted by adashed line above the sequence (nucleotides +25 to +81). Potential sites ofN-linked
glycosylation
(Asn-X-Ser/Thr)areunderlined.
2376
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.146.483.37.694.2]660 670 680 690 700 710
* * * * * *
GTG GAA ATT AGAMT OTA GAT TAT GCT GAT MT OGC TAC ATG CM GTC ATT ATG CCT GAC Val Glu Ile Arg AsnVal Asp Tyr Ala Asp Asn Gly TyrHot Gln Val Ile Hot Arg Asp
720 730 740 750 760 770
* * * * * *
CAT TTT MT COG CCT TTA ATAOAT AA CATATE TAC ATACOT OTO TOT CM CGA CCTOCA His Ph. AsnArg Pro Lou Ile Asp Lys His Ile Tyr Il Arg Val Cys Gln Arg ProAla
780 790 800 610 820 830
* * * * * *
TCA GTG GAT GTA CTO GCCCCT CCA GTC CTC AGCGGA GM MT TAC MO GCA TCT TOT ATC Sor Vol Asp Val Lou Ala Pro Pro Val Lou Sor GlyGluAsn Tyr Lys Ala Ser Cys Ile
840 850 860 870 880 890
* * * * * *
GTT AGA CAC TEE TAT CCC CCT GGA TCT OTC TAT OTA TCT TGG AGA CAG MT GGA MC ATT Vol Arg His Ph. Tyr Pro Pro Gly Ser Vol Tyr Vol Ser Trp ArgGln Asn Gly Asn Ile
900 910 920 930 940 950
* * * * * *
GCA ACT CCT COG AMA AT COC OAT GGA ACT TET TGG TOG TTC GM TCT GGT AGA GGA GCT Ala Thr Pro Arg Lys Asp Arg Asp Gly Ser Ph. Trp Trp Ph. Glu Ser Gly Mrg Gly Ala
960 970 980 990 1000 1010
* * * * * *
ACG TTG GTT TCT ACA ATA ACA TTG GGA MT TCA GGA ATT OAT TTC CCC CCC MAATA TCT
Thr LouVal Ser Thr Ile Thr Lou GlyAsn Ser Gly Ile Asp Phe Pro Pro Lys Ile Ser
1020 1030 1040 1050 1060 1070
* * * * * *
TGT CTG GTT CCC TGG MG CAG GOT OAT ATO ATC AGC ACO ACO MT GCC ACA GCT ATC CCG Cys Lou Val Ala Trp Lys Gln Gly Asp Het Ile Ser Thr Thr JAs Ala Thr Ala Ile Pro
1080 1090 1100 1110 1120 1130
* * * * * *
ACG OTA TAT CAT CAT CCC CGT TTA TCC CTG GCT TTT MA GAT GGG TAT GCA ATA TGT ACT Thr Vol Tyr His His Pro Arg Lou Ser LouAla Ph. Lys Asp Gly Tyr Ala Ile Cys Thr
1140 1150 1160 1170 1180 1190
* * * * * *
ATA GM TOT GTC CCC TCT GAG ATT ACT OTA CGG TGG TTA GTA CAT GAT GM GCG CAG CCT Ile GluCys Vol Pro S-r Glu Ile Thr Val Arg Trp LouVal His Asp Glu Ala Gln Pro
1200 1210 1220 1230 1240 1250
* * * * * *
MC ACA ACT TAT MT ACT GTO GTT ACA GOT CTC TGC CGG ACC ATC GAT CGC CAT AGA MT Asn Thr Thr Tyr Asn Thr Vl Vol Thr Gly Lou Cys Arg Thr Ile Asp Arg His Arg Asn
1260 1270 1280 1290 1300 1310
* * * * * *
CTC CTC AOCCGC ATT CCA OTA TOG OAC MT TOG ACG AA ACA AMTAT ACO TGC AOA CTC Lou Lou Ser Arg Ile Pro Val Trp Asp An Trp Thr Lys Thr Lys Tyr Thr Cys Arg Lou
1320 1330 1340 1350 1360 1370
* * * * * *
ATA 0GC TAC CCC TTC GAT GM GAT MA TET CM OAT TCG GM TAT TAC OAT GCA ACT CCA Il Oly Tyr Pro Ph. Asp Glu Asp Lys Ph. Gln Asp Ser Glu Tyr Tyr Asp Ala Thr Pro
1380 1390 1400 1410 1420 1430
* * * * * *
TCT GCA AOA GGA ACA CCC ATO GTT ATT ACG GTT MC 0C0 ACT TEE 0G0 ATT GGC TGT MT Ser Ala Arg Oly Thr Pro Met Val Il. Thr Val Asn GI Ph. Gly Ile Gly Cys Asn
1440 1450 1460 1470 1480 1490
* * * * * *
TET AGO GTA GOG GAT MT CAT GAC TGC CCT ATO TmT ATA CAC TCC ACA COA AM ATA TTC Ph Arg Vol Oly Asp Asn His Asp Cys Pro Hot Ph. Ie His Sor Thr Arg Lys Ile Ph.
1500 1510 1520 1530 1540 1550
* * * * * *
OAT TAT MT CTC ATT GTTATO TAG TTG TGA TmT ATT MA CAT ATT TET TAT MC TCT ACT Asp Tyr Asn Lou Ile Vl Hot
---1560 1570 1580 1590 1600 1610
* * * * * *
[image:5.612.135.482.60.717.2]ATT CTC CGA GTE ACT TAT ATA TET ATE TOT CAG ACA ATA ATC CM TAG TGG AA MTCOT FIG. 3-Continued.
2377
on November 10, 2019 by guest
http://jvi.asm.org/
2378 COUSSENS AND VELICER
4
3
J
2
I
0
-1
I 100.8 200.8 300.4 400.2 500
[image:6.612.69.307.71.242.2]SEQUENCE POSITION
FIG. 4. Hydrophilicity plot of the predicted MDHV gp57-65 aminoacid sequence.The plotis basedonthealgorithmofHopp andWoods(3)forpredictingpossibleantigenic determinantsites.
DISCUSSION
The MDHV gp57-65genehas beenlocalized, molecularly
cloned, and sequenced. The results presented in this report show that the gp57-65geneis located nearthecenterof the BamHI B fragment and is contained entirely within the 2.35-kb PvuII-EcoRI fragment subcloned into plasmid pUC19togenerateplasmidpl9MDA2.35. Molecular hybrid-izations and nucleotide sequence analyses have indicated
that the analogous gene from the immunologically related vaccinevirus,
HIVT,
islocated inanidentical position in the HVTgenome(Coussens and Velicer, unpublished results).Nucleotidesequenceanalysisof the cloned 2.35-kb
PvuII-EcoRI segment of the BamHI B fragment revealed a long
openreading frame of 1,515 nucleotides which, when trans-lated, encodes a polypeptide of 505 amino acids, with a calculatedmolecular weight of 56,805. The predictedMDHV
gp57-65 polypeptide contains features common to other
known glycoproteins and consistent with previously re-ported properties of the protein (6). One such property is a
hydrophobic amino-terminal region (amino acids1to27)that endsinasequenceconsistentwith the rules of Pearlman and
Halvorson for predicting glycoprotein signal peptidase cleavage sites (16). The existence of a signal peptidase cleavage site in the gp57-65 precursorwas expected, since
incubation of the gp57-65 precursor synthesized in cell-free translation reactions with dog pancreatic microsomes re-sulted incleavage of the precursor(6). The precise site of
signal peptidase cleavage is being investigated by N-terminal
sequence analysis of the mature gp57-65 antigen isolated from MDHV-infected cells. The existence of potential sites for N-linked glycosylation (Asn-X-Ser/Thr) was also
ex-pected, since tunicamycin,aninhibitor of N-linked
glycosyl-ation, completely inhibits glycosylation of this antigen (6). Thepredicted polypeptide is relativelyhydrophobic, with
a possible carboxyl-terminal membrane anchor sequence
(amino acids 461to479).Additional hydrophobic amino acid stretches occurat positions throughout the predicted poly-peptide sequence (Fig. 4). Although the predominantly secretory nature of MDHV gp57-65 suggests that these
sequencesareinsufficienttoanchor theprotein in theplasma membrane, recent evidence (6; unpublished observations) indicates that a small amount of mature gp57-65 can be
detected in the plasma membranes of infected cells. At the present time, however, it is not clear whether this small amountofgp57-65 isactually anchored in the plasma mem-braneorheld by other interactions.
The predicted molecular weight of the gp57-65 precursor
polypeptide (56,805) is considerably
larger
than theapparentmolcularweight of 44,000 currently estimated from previous sodium dodecyl sulfate-polyacrylamide gel electrophoretic analysis of immune precipitation products from cell-free translation reactions or pulse-chase studies (6). With this
method, severedistortion ofprotein migrationinthisregion ofthegelmayresultfromthelargeamountsof immunoglob-ulin G heavy chains (Mr -50,000) present in the immune precipitation reactions. Experiments designed to measure
the apparent molecular weight of the gp57-65 precursor
polypeptide by immunoblotting sodium dodecyl
sulfate-polyacrylamide gelsoftunicamycin-treatedMDHV-infected
DEFculturemediumareinprogressand willprovideasize estimate withoutinterferencefromimmunoglobulin G.
TABLE i. Codon usage in thegp57-65 amino acid sequence
CodonAmino Fre- % Codon Amino Fre- Cd Amino Fre- Codon Amino
Fre-acid
quenccy
acid quencyoon acid quency acid quencyTTT F 16 3.2 TCT S 12 2.4 TAT Y 11 2.2 TGT C 7 1.4
TTC F 6 1,2 TCC S 6 1.2 TAC Y 9 1.8 TGC C 4 0.8
TTA L 7 1.4 TCA S 6 1.2 TAA 0 TGA 0
TTG L 4 0.8 TCG S 4 0.8 TAG 1 TGG W 11 2.2
CTT L 3 0.6 CCT P 11 2.2 CAT H 11 2.2 CGT R 6 1.2
CTC L 12 2.4 CCC P 13 2.6 CAC H 5 1.0 CGC R 3 0.6
CTA L 3 0.6 CCA P 7 1.4 CAA Q 5 1.0 CGA R 5 1.0
CTG L 4 0.8 CCG P 3 0.6 CAG Q 4 0.8 CGG R 5 1.0
ATT I 16 3.2 ACT T 10 2.0 AAT N 20 4.0 AGT S 5 1.0
ATC I 7 1.4 ACC T 4 0.8 AAC N 15 3.0 AGC S 7 1.4
ATA I 13 2.6 ACA T 13 2.6 AAA K 11 2.2 AGA R 8 1.6
ATG M 8 1.6 ACG T 16 3.2 AAG K 6 1.2 AGG R 1 0.2
GTT V 11 2.2 GCT A 6 1.2 GAT D 23 4.6 GGT G 4 0.8
GTC V 7 1.4 GCC A 5 1.0 GAC D 7 1.4 GGC G 8 1.6
GTA V 9 1.8 GCA A 9 1.8 GAA E 20 4.0 GGA G 14 2.8
GTG V 7 1.4 GCG A 6 1.2 GAG E 2 0.4 GGG G 4 0.8
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.70.570.524.725.2]STRUCTURE AND SEQUENCE OF THE MDHV gp57-65 GENE
The total G + C content of the gp57-65 coding region is 41%, which is relatively low compared with that of most
alphaherpesviruses, such asherpes simplex virus type1(18), but consistent with that of many othergammaherpesviruses,
such as herpesvirus saimiri (12). Other genes recently iden-tified in the MDHV genome show a slightly higher overall G+C content than the gp57-65 sequence presented in this report (I. Sithole, P. M. Coussens, and L. F. Velicer, unpublished results).
The data presented here represent the first nucleotide sequence of a known coding region within the MDHV genome. Current studies in our laboratory have indicated that while the overall nucleotide sequence of the HVT gp57-65 gene is 80 to 85%homologous tothatof the MDHV gp57-65 gene, most differences occur in the third base position of gp57-65 codons and are thustranslationallysilent (Coussens and Velicer, unpublished data). In addition to
determining the exact relationship between MDHV and HVT, nucleotide sequence analysis ofa growing body of isolated MDHV genes will allow us to examine the regula-torysequences involved in control of MDHV gene transcrip-tion. Itwillbe of particular interesttodeterminetherole of
suchsequences, ifany, intheloss of gp57-65 gene
expres-sion at the transcriptional level after continued passage in cellculture.Experimentstoprecisely locatethe mRNA start
site and to identify the promoter sequences ofthe gp57-65 gene are in progress. Thus, the molecule nowidentified as gp57-65 (formerly called A antigen) continues to be of interest and importance in this natural host animal model system. Furtheranalysis of gp57-65andthe geneencodingit will helpdetermine its precise role in theimmunobiologyand
pathogenesis of MDHV.
ACKNOWLEDGMENTS
This study was supported by grant 85-CRCR-1-1879 to L. F. Velicer under the Competitive Research Grants Program adminis-teredby the U.S. Department of Agriculture and byagrantfrom the Michigan State University Biotechnology Research Center to L. F. Velicer.
We thank Ruth A. Stringer for excellent technical assistance, Peter Brunovskis for help in mapping the TaqI fragments of the gp57-65 gene, Larry Snyder andCheng Kao for helpful technical advice on DNAsequencing, and IdahSithole for careful review of themanuscript and many helpful suggestions.
LITERATURECITED
1. Fukuchi, K.,M.Sudo, Y.-S. Lee, A. Tanaka, and M.Nonoyama. 1984.Structure of Marek's disease virusDNA:detailed restric-tionenzyme map. J. Virol. 51:102-109.
2. Glaubiger, C., K. Nazerian, and L. F. Velicer. 1983. Marek's disease herpesviruses. IV. Molecular characterization of Ma-rek'sdiseaseherpesvirusAantigen. J. Virol. 45:1228-1234. 3. Hopp, T. P., and K. R. Woods. 1981. Prediction of protein
antigenic determinants from amino acid sequences. Proc. Natl. Acad. Sci. USA78:3824-3828.
4. Ikuta, D., S. Ueda, S. Kato, and K. Hirai. 1983. Most virus-specific polypeptides in cells productively infected with Marek's disease virusor herpesvirus of turkeys possess cross-reactive determinants.J. Gen. Virol.64:961-965.
5. Isfort, R.J.,H.-J. Kung,and L. F. Velicer. 1987.Identification
of thegene encoding Marek's disease herpesvirus A antigen. J. Virol. 61:2614-2620.
6. Isfort,R.J.,R.A.Stringer, H.-J. Kung, and L. F. Velicer. 1986. Synthesis, processing, and secretion of the Marek's disease herpesvirus A antigen glycoprotein. J. Virol. 57:464-474. 7. Lee, L. F., E. D. Kieff, S. L. Bachenheimer, B. Roizman, P. G.
Spear, B. R. Burmester, and K. Nazerian. 1971. Size and composition of Marek's disease virus deoxyribonucleic acid. J. Virol. 7:289-294.
8. Long, P. A., J. L. Clark, and L. F. Velicer. 1975. Marek's diseaseherpesviruses. II. Purification and further characteriza-tion of Marek's disease herpesvirus A antigen. ,J. Virol. 15:1192-1201.
9. Long, P. A., P. Kaveh-Yamini, andL.F.Velicer. 1975.Marek's diseaseherpesviruses. I. Production andpreliminary character-ization of Marek's disease herpesvirus A antigen. J. Virol. 15:1182-1191.
10. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular cloning: a laboratory manual. ColdSpring Harbor Laboratory, ColdSpring Harbor, New York.
11. Marek, J. 1907. Multiple Nerventzuendung (Polyneuritis) bei Huehnern.Dtsch.Tieraerztl. Wochenschr. 15:417-421. 12. Melendez, L. V., M. D. Daniel, R.D.Hunt, andF. G. Garcia.
1968. An apparently new herpesvirus from primary kidney cultures of the squirrel monkey (Saimiri sciureus). Lab. Anim. Care 18:374-381.
13. Okazaki, W., H. G. Purchase, and B. R. Burmester. 1970. Protection against Marek's disease by vaccination witha her-pervirus ofturkeys (HVT). Avian Dis. 14:413-429.
14. Pappenheimer, A. M., L. C. Dunn, and V. Cane. 1929. Studies onfowlparalysis (neurolymphomatosis gallinarum). I. Clinical features and pathology. J. Exp. Med. 49:63-86.
15. Pappenheimer, A. M., L. C. Dunn, and S. M. Seidlin. 1929. Studiesonfowlparalysis(neurolymphomatosis gallinarum). II. Transmissionexperiments.J. Exp. Med. 49:87-102.
16. Pearlman, D., and H. 0. Halvorson. 1983. A putative signal peptidase recognition site and sequence in eukaryotic and prokaryotic signal peptides. J. Mol. Biol. 167:391-409. 17. Purchase, H. G., W. Okazaki, and B. R. Burmester. 1972.
Long-term field trials with the herpesvirus of turkeys vaccine againstMarek'sdisease. Avian Dis. 16:57-71.
18. Roizman, B. 1979. The structure and isomerization of herpes simplex virus genomes. Cell 16:481-494.
19. Ross, L.J. N., 0. Basarab, D. J. Walker, and B.Whitby. 1975. Serologicalrelationship betweenapathogenic strain of Marek's disease virus, its attenuated derivative and herpesvirus of turkeys.J. Gen.Virol. 28:37-47.
20. Sanger, F.,S.Nicklen,and A. R.Coulson. 1977. DNA sequenc-ing with chain terminating inhibitors. Proc. Natl. Acad. Sci. USA 74:5463-5470.
21. Sharma, J. M.,and H. A. Stone. 1972. Genetic resistance to Marek's disease. Delineation of the response of genetically resistantchickensto Marek'sdiseasevirus infection.AvianDis. 16:894-906.
22. Silva,R.F.,and L. F. Lee.1984. Monoclonalantibody-mediated immunoprecipitation ofproteins from cells infected with Ma-rek's disease virusorturkeyherpesvirus. Virology136:307-320. 23. Velicer, L. F., D. R. Yager, and J. L. Clark. 1978. Marek's disease herpesviruses. III. Purification and characterization of Marek'sdisease herpesvirusBantigen.J. Virol. 27:205-217. 24. Witter, R.L., K.Nazerian, H. Purchase, and G. H. Burgoyne.
1970. Isolation from turkeys of a cell-associated herpesvirus antigenically relatedtoMarek'sdisease virus. Am. J. Vet. Res. 31:525-538.
VOL. 62,1988 2379
![FIG.1.orientationbywithation,wereProbes6,probegp57-65ProbeMDHV-infecteduninfectedinsteadase11arrows,the[(a-32P]dGTP,SsplMDHV plus respectively, Fine structure mapping of the MDHV gp57-65 gene](https://thumb-us.123doks.com/thumbv2/123dok_us/1338727.87591/2.612.335.549.68.321/orientationbywithation-wereprobes-probegp-probemdhv-infecteduninfectedinsteadase-ssplmdhv-respectively-structure.webp)